


INTRODUCTION
Archinome rosacea (Blake, Reference Blake1985) was originally described as belonging to the genus Euphrosine Savigny in Lamarck, Reference Lamarck1818 in the family Euphrosinidae Williams, Reference Williams1852 and later redescribed by Kudenov (Reference Kudenov, Petersen and Kirkegaard1991), who transferred it to the newly erected genus Archinome Kudenov, Reference Kudenov, Petersen and Kirkegaard1991 and established the new family Archinomidae Kudenov, Reference Kudenov, Petersen and Kirkegaard1991. According to this author the new family differs in the morphology of the caruncle, the branchiae and the parapodia in comparison to the Euphrosinidae as well as the Amphinomidae Savigny in Lamarck, Reference Lamarck1818. However, some authors believe that Archinome falls within Amphinomidae and that the erection of a new family was not justified (e.g. Fauchald & Rouse, Reference Fauchald and Rouse1997; Pleijel, Reference Pleijel, Rouse and Pleijel2001; Wiklund et al., Reference Wiklund, Nygren, Pleijel and Sundberg2008). Molecular studies by Wiklund et al. (Reference Wiklund, Nygren, Pleijel and Sundberg2008) revealed a close phylogenetic relationship of Archinome rosacea to Chloeia flava and the two species form a sister group to the remaining Amphinomidae. Wiklund et al. (Reference Wiklund, Nygren, Pleijel and Sundberg2008) opt for inclusion of Archinome within Amphinomidae, while the other option would be to include Chloeia within Archinomidae. Since there is no new information to decide between the two possibilities, we still consider the position of the genus Archinome unsettled and follow the current state of systematic placement for the time being.
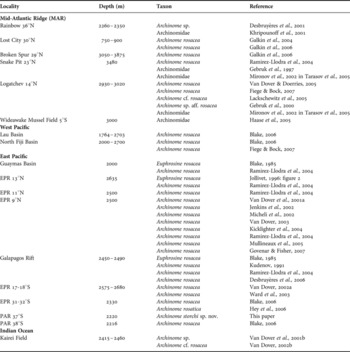
According to published records from various locations in the Pacific, Atlantic, and Indian Oceans (Table 3) Archinome rosacea appears to be a cosmopolitan species, in contrast to most hydrothermal vent species which usually show a much more restricted distribution. However, the majority of these records is not supported by a detailed description based on the respective specimens and should therefore be regarded as tentative or preliminary identifications. A thorough study of morphological characters of specimens available to us from various locations in the Pacific and on the Mid-Atlantic Ridge (MAR) revealed significant differences for specimens collected from the Pacific–Antarctic Ridge (PAR). Specimens of A. rosacea collected from the North Fiji Basin fit to the original description while those from the Logatchev site on the MAR showed slight variations in some characters. Archinome storchi sp. nov. is described as new to science based on specimens collected from the PAR at 37°S.
MATERIALS AND METHODS
The specimens examined in this study have been collected during various cruises of RV ‘Sonne’ and RV ‘Meteor’ to hydrothermal vent sites in the Pacific Ocean, i.e. So 99, Hyfiflux I: North Fiji Basin (Halbach et al., Reference Halbach, Auzende and Türkay1996); So 134, Hyfiflux II: North Fiji Basin (Halbach et al., Reference Halbach, Giere, Seifert and Seifert1998); So 157, Foundation 3: PAR (Stoffers et al., Reference Stoffers, Worthington, Petersen, Hannington, Türkay, Ackermand, Borowski, Dankert, Fretzdorff, Haase, Hekinian, Hoppe, Jonasson, Kuhn, Lancaster, Monecke, Renno, Stecher and Weiershäuser2001), and to the MAR, i.e. Me 60/3, Hydromar I: MAR (Kuhn et al., Reference Kuhn, Alexander, Augustin, Birgel, Borowski, Carvalho, Engemann, Ertl, Franz, Grech, Hekinian, Imhoff, Jellinek, Klar, Koschinsky, Kuever, Kulescha, Lackschewitz, Petersen, Ratmeyer, Renken, Ruhland, Scholten, Schreiber, Seifert, Süling, Türkay, Westernströer and Zielinski2004) (Table 1).
Table 1. List of stations (abbreviations: MAR, Mid-Atlantic Ridge; PAR, Pacific–Antarctic Ridge; Me, RV ‘Meteor’; So, RV ‘Sonne’; TVG, TV grab; ROV, remotely operated vehicle; HV, hot vent).

Benthic samples were taken using Van Veen grabs with integrated TV camera (TVG), and remotely operated vehicles (ROVs) and sieved on board. Specimens were fixed in 10% formaldehyde–seawater solution and later transferred to 70% ethanol. Preserved specimens were examined using stereo and compound microscopes.
For SEM investigations specimens were dehydrated via graded ethanol series, critical point dried using CO2, coated with Au–Pd and examined in a CamScan CS 24 SEM equipped with ORION digital image capture. Light microscopical pictures were taken with a Nikon Coolpix 4500 or Canon G7 camera adapted to a Leica/Wild MZ 8 stereomicroscope. Drawings of specimens were made using a camera lucida and finalized according to the method described by Coleman (Reference Coleman2003). Illustrations were assembled using Adobe Photoshop and Adobe Illustrator.
Specimens are indicated in the text as: cs (complete specimen) and af (anterior fragment). In this description we follow Fauchald & Rouse (Reference Fauchald and Rouse1997) and Pleijel (Reference Pleijel, Rouse and Pleijel2001) using the term palps for the slender, digitiform prostomial appendages called palpal cirri by Kudenov (Reference Kudenov, Petersen and Kirkegaard1991). Likewise we prefer the term peristomial lips for the flat pads around the mouth (s.a. Fauchald & Rouse, Reference Fauchald and Rouse1997; Pleijel, Reference Pleijel, Rouse and Pleijel2001) described as palps by Kudenov (Reference Kudenov, Petersen and Kirkegaard1991). For the definition of dorsal, lateral, ventral cirri and nuchal cirrus we follow Kudenov (Reference Kudenov, Petersen and Kirkegaard1991). Since it appears uncertain whether the darkly pigmented spots on the prostomium represent eyes (Kudenov, Reference Kudenov, Petersen and Kirkegaard1991: eyes non lenticulate) we follow Ward et al. (Reference Ward, Jenkins and Van Dover2003) in using ‘eyespots’.
Type and other specimens are deposited in the Senckenberg Museum Frankfurt (SMF).
Table 2. Synoptic table of characters of Archinome storchi sp. nov. and Archinome rosacea.

TYPE MATERIAL
Holotype (cs), SMF 17876, south-east Pacific. RV ‘Sonne’ 157, Foundation 3, PAR: Central Axial High, Station 30 TVG, 37°47.467′ S 110°54.868′ W, 2212 m, 28.6.2001, sample containing glassy silica-rich lava and other vent related fauna (crabs, mussels, barnacles and polychaetes).
Table 3. Records of Archinomidae from the literature, including tentative or preliminary identifications (abbreviations: EPR, East Pacific Rise; PAR, Pacific–Antarctic Ridge).
Paratype (cs) as SEM stub 884, with chaetae removed, SMF 17877, same locality as holotype. 1 additional specimen (cs), in two fragments as SEM stubs 872, 874, SMF 17878, same locality as holotype.
COMPARATIVE MATERIAL EXAMINED
Archinome rosacea (Blake, Reference Blake1985): Holotype (cs), USNM 81788, East Pacific, Galapagos Rift geothermal vents, Alvin Dive 984, Rose garden, 00°48.3′ N 86°13.5′ W, 2447–2460 m, 1.12.1979, mussel washings.
Archinome rosacea (Blake, Reference Blake1985): North Fiji Basin, RV ‘Sonne’ 99, Hyfiflux I: Station 48 TVG, 1 spm (SMF 17679); Station 115 TVG, 2 spms (SMF 17880)—RV ‘Sonne’ 134, Hyfiflux II: Station 33 TVG, 31 spms (SMF 17881); Station 35, many (SMF 17903); Station 66 TVG, 4 spms (SMF 17882); Station 99 TVG, many (SMF 17883)—MAR, RV ‘Meteor’ 60/3, Hydromar I: Station 23 ROV 10, 1 spm (SMF 17884), 1 spm on SEM stub 886 (SMF 17885); Station 35 TVG, many (SMF 17886), 3 spms on SEM stubs 870 (SMF 17887), 871 (SMF 17888), 877 (SMF 17889); Station 38 ROV, many (SMF 17890), 1 spm (SMF 17891), 3 spms on SEM stubs 883 (SMF 17892), 885 (SMF 17893), 887 (SMF 17894); Station 56 ROV, 11 spms (SMF 17895), 5 spms on SEM stubs 875 (SMF 17896), 876 (SMF 17897), 878 (SMF 17898), 880 (SMF 17899), 881 (SMF 17900); Station 66 ROV, >11 spms (SMF 17901), 1 spm on SEM stub 873 (SMF 17902).
DESCRIPTION
Holotype short, fusiform with a median row of ventral glandular pads. Length 15 mm for 23 chaetigers (including reduced posteriormost chaetiger, only visible in SEM); width 4.5 mm at widest part, appendages and chaetae not included. Body trapezoidal in cross-section (Figure 1A, B). Colour of body in alcohol light yellow; chaetae light yellow to transparent.

Fig. 1. Archinome storchi sp. nov. Holoype (SMF 17876): (A) Lateral view, right side; (B) dorsal view. Scale bar: 5 mm.
Prostomium extending anteroventrally into peristomial lips. Two pairs of ‘eyespots’; one pair of oblong mid-dorsal ‘eyespots’ in front of dorsomedial antennae and two elongate ‘eyespots’ situated ventrolaterally of chaetiger 1. One pair of dorsomedial antennae arising from a small pit posterior to dorsal pair of ‘eyespots’. A pair of slender ventrolateral palps present. Antennae and palps cirriform, similar in shape and size. Peristomium consisting of 2 large cushion-like lips separated by a midventral groove leading to mouth. Caruncle elongate extending posteriorly from chaetiger 1 to chaetiger 4, with median keel, lateral, longitudinal, ciliated ridges and ciliated groves (Figures 2A, 3A); caruncle of K-1 category according to Kudenov (Reference Kudenov1987). Long nuchal cirrus inserting at anterior end of caruncle, without cirrophore (Figure 2B).

Fig. 2. Archinome storchi sp. nov. Holotype (A, B: SMF 17876). (A) Anterior region, dorsal view; (B) detail of (A). Nuchal cirrus (arrow); Archinome rosacea. Holotype (USNM 81788); (C) anterior region, dorsal view. Nuchal cirrus (arrow); A. rosacea from Logatchev (MAR) (SMF 17893); (D) anterior region, dorsal view; upper part of chaetae removed. Nuchal cirrus (arrow). Scale bars: A–C, 1 mm.

Fig. 3. Archinome storchi sp. nov. Paratype (SMF 17877). (A) Anterior region, dorsal view; (B) left parapodium of chaetiger 4, posterior view; (C) notochaeta. Scale bars: A & B, 1 mm; C, 250 µm.
All parapodia biramous. Parapodia of first segment strongly reduced and projecting anteriorly. Parapodia of segment 2 and the following segments with low, conical notopodia, projecting dorsally (Figure 3B). Dorsal cirri long, slender, without cirrophores, inserting anteriorly to branchiae on dorsomedian border of notochaetal field (Figure 4A). Lateral cirri shorter, arising behind apex of notopodia; long, cirriform and basally coiled; thick cirrophore present (Figure 4B).

Fig. 4. Archinome storchi sp. nov. (A–C: SMF 17877; D & E: SMF 17878): (A) Dorsal cirrus and branchiae on chaetiger 8, dorsal view; (B) notopodium on chaetiger 10 with coiled lateral cirrus, anterior view; chaetae removed; (C) neuropodia with ventral cirri, chaetigers 4–6 from left to right, lateral view; chaetae removed; (D) anal cirrus, lateral view; (E) posterior end, dorsal view. Archinome rosacea from Logatchev (MAR) (SMF 17892); (F) posterior end, dorso-lateral view.
Neuropodia mound-shaped, dorsally pointed and projecting laterally. Ventral cirri slender, cirriform inserting midventrally, oriented posteriorly and upwards; with very short cirrophore. Chaetal field surrounded by epidermal bulge (Figure 4C).
Branchiae starting on chaetiger 3 with three filaments, palmate with short common trunk inserting on posterodorsal border of notopodial chaetal field (Figure 4A). Six filaments at chaetiger 4, seven filaments at chaetiger 7.
All chaetae simple and hollow. Neurochetae and notochaetae bifurcate with tines varying greatly in length. Ringent notochaetae absent. Notochaetae slightly stronger than neurochaetae (Figure 3C).
Single, thick, elongate, median pygidial cirrus with short transverse band of dark pigment on dorsal side (Figure 4D). Anus dorsally on chaetiger 19 (5th terminal chaetiger) surrounded by a thick, glandular epidermis (Figure 4E). Last chaetiger strongly reduced and projecting posteriorly, lacking cirri.
REMARKS
The species Archinome storchi sp. nov. differs from A. rosacea in the position of the anus, the length of the nuchal cirrus and the first appearance of branchiae (Table 2). The anus of A. storchi sp. nov. is positioned on the 5th terminal chaetiger (including reduced posteriormost chaetiger) while in A. rosacea it is situated on the 2nd terminal chaetiger with the pygidial cirrus arising directly behind the anus (Figure 4F) (reduced posteriormost segment not observed in holotype of A. rosacea under stereomicroscope). The nuchal cirrus of A. storchi sp. nov. is about twice as long as in A. rosacea (Figure 2B, C). Branchiae appear first on chaetiger 3 with three filaments in A. storchi sp. nov., while they appear on chaetiger 2 with 1–2 filaments in A. rosacea. Transverse band of dark pigment on dorsal side of pygidial cirrus is continuous in A. storchi sp. nov. and discontinuous in A. rosacea (s. Kudenov, Reference Kudenov, Petersen and Kirkegaard1991: figure 28). Moreover, appendages such as prostomial antennae, parapodial cirri and palps appear more delicate in A. storchi sp. nov. than in A. rosacea.
The specimens of Archinome collected from Logatchev (MAR) differ from A. rosacea and A. storchi sp. nov. by the presence of a shorter, minute nuchal cirrus (Figure 2D). Branchiae appear first on chaetiger 3 with 2–3 filaments. Whether this combination of characters indicates the presence of another undescribed species of Archinome on the MAR or simply represents intraspecific variability can only be solved by genetic studies of populations from various areas.
ETYMOLOGY
The species is named after Professor Dr Dr Volker Storch (Heidelberg), a distinguished German zoologist, who made important contributions to our knowledge of the anatomy, ultrastructure and taxonomy of Polychaeta.
DISTRIBUTION
Archinome storchi sp. nov. is only known from the type locality on the PAR at 37°S.
ACKNOWLEDGEMENTS
We thank our colleagues Thomas Jellinek, Michael Türkay and Jens Stecher (all Senckenberg, Frankfurt) for collecting and making the material available to us. Kristian Fauchald and Valorie Barnes (USNM, Smithsonian Institution, Washington, DC) kindly provided the holotype of Archinome rosacea on loan. Our thanks are also due to Melih Ertan Çinar and an anonymous referee for their valuable comments on the manuscript.







