INTRODUCTION
Internal defence in invertebrate species depends on an innate, non-lymphoid immune system. It consists of a variety of cell types and effector molecules, which interact to eliminate effectively foreign bodies. The haemocytes of molluscs play an important role in their defence against potential pathogens. Haemocytes are thought to be involved in many functions, such as shell repair (Sparks & Morado, Reference Sparks and Morado1988), digestion and transport of nutrients (Bayne, Reference Bayne, Saleuddin and Wilbur1983), excretion (Narain, Reference Narain1973) and immune defence (Bayne, Reference Bayne, Saleuddin and Wilbur1983). The most important role of haemocytes, however, is the internal defence (Cheng, Reference Cheng, Ratcliffe and Rowley1981). The haemocytes may also produce other soluble compounds as part of the defence strategies, including agglutinins, lectins (Renwrantz & Stahmer, Reference Renwrantz and Stahmer1983; Leippe & Renwrantz, Reference Leippe and Renwrantz1988) and antibacterial peptides (Mitta et al., Reference Mitta, Vandenbulcke, Hubert, Salzet and Roch2000).
Most studies on the morphological characteristics and functions of haemocytes in the gastropod have focused on the pulmonates Biomphalaria glabrata (Hahn et al., Reference Hahn, Bender and Bayne2000; Bender et al., Reference Bender, Broderick, Goodall and Bayne2005; Humphries & Yoshino, Reference Humphries and Yoshino2008) and Lymnaea stagnalis (Wright et al., Reference Wright, Lacchini, Davies and Walker2006; Russo & Madec, Reference Russo and Madec2007; Russo et al., Reference Russo, Madec and Brehélin2008), and were also reported in the abalone species Haliotis diversicolor (Chen et al., Reference Chen, Yang, Peng, Chen and Tsai1996; Gopalakrishnan et al., Reference Gopalakrishnan, Thilagam, Huang and Wang2009), Haliotis asinine (Sahaphong et al., Reference Sahaphong, Linthong, Wanichanon, Riengrojpitak, Kangwanrangsan, Viyanant, Upatham E., Pumthong, Chansue and Sobhon2001), Haliotis discus discus (Donaghy et al., Reference Donaghy, Hong, Lambert, Park, Shim and Choi2010), Haliotis rufescens and Haliotis cracherodii (Armstrong et al., Reference Armstrong, Armstrong, Krassner and Pauley1971; Martello et al., Reference Martello, Friedman and Tjeedema2000; Martello & Tjeerdema, Reference Martello and Tjeerdema2001), and Haliotis tuberculata (Serpentini et al., Reference Serpentini, Ghayor, Poncet, Hebert, Galera, Pujol, Boucaud-Camou and Lebel2000; Malham et al., Reference Malham, Lacoste Gélébart, Cueff and Poulet2003; Poncet & Lebel, Reference Poncet and Lebel2003; Travers et al., Reference Travers, Mirella Da Silva, Le Goïc, Marie, Donval, Huchette, Koken and Paillard2008) and in only a few other gastropods (Pauley et al., Reference Pauley, Krassner and Chapman1971; Kumazawa et al., Reference Kumazawa, Tanigawa, Tanaka, Osatake and Tanaka1990, Reference Kumazawa, Tanigawa, Kasagi and Tanaka1991; Adamowicz & Bolaczek, Reference Adamowicz and Bolaczek2003; Gorbushin & Iakovleva, Reference Gorbushin and Iakovleva2006; Martin et al., Reference Martin, Oakes, Tousignant, Crabtree and Yamakawa2007; Mahilini & Rajendran, Reference Mahilini and Rajendran2008; Donaghy et al., Reference Donaghy, Hong, Lambert, Park, Shim and Choi2010).
Classification of gastropod haemocytes has been based on light and electron microscopy (Adema et al., Reference Adema, Harris and Van Deutekom-Mulder1992; Chen et al., Reference Chen, Yang, Peng, Chen and Tsai1996; Adamowicz & Bolaczek, Reference Adamowicz and Bolaczek2003; Gorbushin & Iakovleva, Reference Gorbushin and Iakovleva2006; Martin et al., 2007; Mahilini & Rajendran, Reference Mahilini and Rajendran2008), differential centrifugation (Adema et al., Reference Adema, Mohandas, Van Der Knaap and Sminia1994), flow cytometry (Russo & Lagadic, Reference Russo and Lagadic2004; Cossarizza et al., Reference Cossarizza, Pinti, Troiano and Cooper2005; Russo & Madec, Reference Russo and Madec2007; Russo et al., Reference Russo, Madec and Brehélin2008; Travers et al., Reference Travers, Mirella Da Silva, Le Goïc, Marie, Donval, Huchette, Koken and Paillard2008; Donaghy et al., Reference Donaghy, Hong, Lambert, Park, Shim and Choi2010), enzyme content (Granath & Yoshino, Reference Granath and Yoshino1983), lectin and antibody binding (Yoshino & Granath Jr, Reference Yoshino and Granath1985; Dikkeboom et al., Reference Dikkeboom, Tijnagel and Van Der Knaap1988) and functional studies (Cheng, Reference Cheng, Bullo and Cheng1984).
One or two types of haemocytes are commonly described (Voltzow, Reference Voltzow, Harrison and Kohn1994). Sminia & Barendsen (Reference Sminia and Barendsen1980) suggest that only one category of haemocyte, the amoebocyte, exists in the freshwater snails, but many researchers argue that granular and agranular haemocytes could be readily recognized in other molluscs. It is now commonly accepted that two types of haemocytes exist, namely granulocytes and hyalinocytes (agranulocytes) (Cheng, Reference Cheng, Ratcliffe and Rowley1981; Yonow & Renwrantz, Reference Yonow and Renwrantz1986). Hyalinocytes contain few or no granules, and granulocytes contain granules and an eccentric, round to ovoid nucleus. While granulocytes may appear to be homogeneous, various hyalinocyte subpopulations were reported (Chen et al., Reference Chen, Yang, Peng, Chen and Tsai1996; Matricon-Gondran & Letocart, Reference Matricon-Gondran and Letocart1999a; Adamowicz & Bolaczek; Reference Adamowicz and Bolaczek2003; Gorbushin & Iakovleva, Reference Gorbushin and Iakovleva2006) and also juvenile or blast-like cells (Barracco et al., Reference Barracco, Steil and Gargioni1993; Chen et al., Reference Chen, Yang, Peng, Chen and Tsai1996; Matricon-Gondran & Letocart, Reference Matricon-Gondran and Letocart1999a; Gorbushin & Iakovleva, Reference Gorbushin and Iakovleva2006; Travers et al., Reference Travers, Mirella Da Silva, Le Goïc, Marie, Donval, Huchette, Koken and Paillard2008, Donaghy et al., Reference Donaghy, Hong, Lambert, Park, Shim and Choi2010). It is not clear whether such diversity in haemocyte subpopulations represents distinct cell lineages, variations in physiological state, or differences in methodology being applied.
Babylonia areolata is classified in the Gastropoda, Prosobranchia, Neogastropoda, Buccinidae. Neogastropoda represents a broad class of Gastropoda. Until now, there has been little research about morphology of blood cells in Neogastropoda.
Babylonia areolata is a commercially important aquaculture species distributed along the south-east coast of mainland China. Annual output is more than 1000 tons, valued at more than 100 million Renminbi (RMB). The increasing bacterial diseases such as vibriosis, proboscis intumescence disease and shell cast disease (Feng et al., Reference Feng, Zhou, Xie, Pu, Lu, Lin, Zhou and Fu2008) have threatened the sustainable development of natural and cultured stocks of Babylonia areolata. In the context of infectious diseases in the molluscan aquaculture, research must be focused not only on the diagnosis of diseases but also on producing disease-resistant animals. This latter strategy depends heavily on the development of the knowledge concerning marine invertebrate immunology. Investigation into the Babylonia areolata immune system is very important because little is known about the cytoimmunity of marine gastropods compared to that of bivalve molluscs. Characterization of the haemocytes is the first step for understanding the immune function and its potential failure during disease outbreaks. The immune response of Babylonia areolata, especially their haemocyte composition, has not been studied. The aims of this work are to offer a definition of blood cells of Babylonia areolata and enrich the research on gastropod immunology.
MATERIALS AND METHODS
Animals
The adults of Babylonia areolata (2–3.5 cm shell length) were collected from Dongshan Haitian Aquaculture Co., Ltd, Fujian Province. The specimens were checked for parasites or pathogens, and parasites and pathogens were not found. Snails were maintained in flow-through water (26–29%, 25°C and pH 7.8–8.5). A layer of fine calcareous sand was added to allow burrowing. They were fed daily with oyster and chopped fresh fish.
Sampling of haemolymph
Snails (2–3.5 cm shell length) were sampled. Surface water adhering to the snail was removed and the foot was cleaned with absorbent paper. By touching the foot with the point of a micropipette tip, the snail was forced to retract deeply into its shell and extruded haemolymph (cf Sminia, Reference Sminia1972). In this way about 100 µl of haemolymph could be obtained from each snail. The blood was collected with an Eppendorf pipette, to avoid haemocyte aggregation, and the hemolymph was immediately transferred into 1.5 ml Eppendorf tubes containing the same quantity of anticoagulants (Anticoagulants ZA: the solution consist of glucose 2.05 g, sodium citrate (2H2O) 0.80 g, NaCl 0.42 g, HEPES 10 Mm in 100 ml distilled water; 10% citric acid adjusted to pH 6.1 (112°C sterilization )) and the mixture was agitated to avoid likely clumping of haemocytes.
Haemocyte morphology—light microscopy
Differential staining was carried out using improved Wright's stain and safranin dye. Haemolymph from 9 snails (2–3.5 cm shell length) was pooled. To the hemolymph/anticoagulant mixture (1:1 by volume) was added the same volume of 100% methanol, fixed in methanol for 6 minutes. An 8 µl suspension was placed on a glass slide, smeared evenly, and blow-dried with electric blower, stained for 12 minutes with Wright's stain, washed with double distilled water, then air-dried. We have also tried using safranin staining, stained for 5 minutes with safranin dye.
Haemocyte morphology—electron microscopy (EM)
Haemolymph from 9 snails (2–3.5 cm shell length) was pooled. A 0.5 ml haemolymph was sampled and 0.5 ml 5% glutaraldehyde was added in Eppendorf tubes and fixed for 1 hour at 4°C, then centrifuged at 700 rpm/min for 60 seconds. The supernatant was removed; the pellet was added in 0.3 ml 4% agarose solution which maintained at 50°C. Agar blocks were added to the EM fixative, 2.5% glutaraldehyde. After fixation for 2 hours at 4°C, the suspension was centrifuged (800 g, 10 minutes). The pellet was washed in Pipes buffer with sucrose for 2 hours at 4°C, and then incubated in 1% osmium tetroxide in Pipes buffer for 75 minutes at 4°C. After being washed in Pipes buffer, the cells were put into 1.5% agar at 40°C and centrifuged (1400 g, 5 minutes). The haemocytes were then dehydrated through an ethanol series and finally embedded via propylene oxide in Taab epoxy resin (Taab Ltd, Aldermaston, UK). Ultrathin sections were cut using an ultramicrotome, ultrathin sections with the thickness in 90 nm, double-stained with uranyl acetate followed by lead citrate, and then examined using a JEM2100 electron microscope.
Cell counts and size measurement
An 8 µl suspension of the haemolymph/anticoagulant mixture was placed on a glass slide and stained with Wright's stain, and each type of haemocyte was counted. Cells and nucleus diameters of the haemocytes were measured using a light microscope with an eye-piece graticule. To obtain cell and nucleus diameter of granulocytes and hyalinocytes, 100 cells per snail were measured; there were 16 snails (2–3.5 cm shell length) for cell counts and size measurement. In total, 1600 cell and nucleus diameters were measured and then the N/C ratio (N indicates nucleus diameters, C indicates cell diameters) was calculated.
As type I granulocytes and type II granulocytes cannot be distinguished in the light microscope, for type II granulocytes, cells and nucleus diameters of type II granulocytes were measured using transmission electron microscopy.
Histological study
Nine snails (2–3.5 cm shell length) were sampled. In order to explore the role of these tissues in haematopoeisis, the alimentary tract and the digestive gland were removed from their shells, fixed in Bouin's fluid for histological studies. Further procedures included dehydration through an ascending series of ethanol concentrations (LeicaTP1020), clearing in xylol and paraffin embedding were followed. Five μm sections were stained with haematoxylin and eosin. Stained slides were examined under light microscope.
The relationships between the concentration of haemocytes and the snail shell length and shell weight
Twenty-seven other snails were equally divided into three size-groups: small, (2.76 ± 0.17 cm), medium (3.06± 0.05 cm) and large (3.31 ± 0.12 cm). Nine snails for each size-group, 100 µl haemocyte samples from each snail, and haemolymph samples were pooled for each size-group. To these were added the same volume of anticoagulant. We measured 8 µl of the mixture using a blood cell haemocytometer, and we counted the number of haemocytes and the haemocyte concentration. Cells were counted 5 times for each size-group and the mean value was calculated using one-way analysis of variance (ANOVA).
RESULTS
Haemocyte morphology—light microscopy
Comparing Wright's staining and safranin staining, Wright's differential staining was the most successful in characterizing the haemocytes. Wright's staining can distinguish haemocyte populations better and make the demarcation line between nucleus and cytoplasm clear. For safranin staining, the cytoplasm and nucleus were stained red; the demarcation line between nucleus and cytoplasm lacked definition and colour difference was not obvious. Two haemocyte types were distinguished by light microscopy: granulocytes and hyalinocytes, based on the presence or the absence of cytoplasmic granules, respectively. Cytoplasmic granules were present in the granulocyte endoplasm, whereas hyalinocytes had few or none.
GRANULOCYTES
With the differential staining, the nucleus appeared blue and the cytoplasm purplish-red. The granulocytes were oval and contained a very high density of large deep-carmine stained granules throughout their entire cytoplasm. They had an oval nucleus, with a diameter of 3.62± 0.71 µm, and the granulocytes themselves had a diameter of 8.01± 0.94 µm. The granules were approximately 0.5 µm in diameter and there was a low karyoplasmic ratio (Figure 1 A–E).

Fig. 1. Light microscopy of haemocytes in Babylonia areolata. (A–E) Light microscopy of granulocytes; (A–D) granulocyte stained with Wright's stain, showing a blue oval nuclear area and the cytoplasm packed with large carmine pigment granules, about 0.5 µm in diameter, and characterized by their spherical shape; (E) granulocyte stained with safranin dye, granulocyte (gh); hyalinocytes (hh); nucleus (n); granule (g); (F–U) light microscopy of larger hyalinocytes; (G, H, I & O) cell shape is in turn thread-like, spindly, kidney-shaped and spherical; (G–I) cell diameter is between 5.7 and 8.2 µm; (G, I & J) cell has a kidney-shaped nucleus; (H) cell has a strip-shaped nucleus; (K–P) nucleus is bifoliate, heart-shaped, horse hoof-shaped, oval, spherical, or binucleate in turn; (Q, S, T & U) cells have pseudopodia (p); (U) cell stained with safranin, showing pseudopodia and nucleus; (V–Y) light microscopy of small hyalinocytes; (V–X) cell diameter is <6.2 µm, with a large nucleus, tiny cytoplasm, and an oval or rotund nucleus; (Y) arrow points at the small hyalinocytes. Scale bar: A–Y = 5 µm.
HYALINOCYTES
With staining, the nucleus appeared blue and the cytoplasm light blue or violet due to metachromasia. The hyalinocytes were also recognizable as to their small size, high karyoplasmic ratio, and the cytoplasm contained few or no granules. Hyalinocytes consist of two classes—large and small hyalinocytes.
LARGE HYALINOCYTES
These cells were various shapes, oval, round, thread-like, spindly, or kidney-shaped; had one or two nuclei; the nuclei varied in shape (kidney-shaped, like two leaves, heart-shaped, horse hoof-shaped, or peanut-shaped), and they did not contain any appreciable number of granules under the light microscope. These hyalinocytes showed great ability to produce pseudopodia (Figure 1 F–U).
SMALL HYALINOCYTES
The two most abundant cell types were granulocytes and large hyalinocytes, and small hyalinocytes were very rare. These cells (approximately 3–5 µm in diameter) were spherical or ovoid in shape and their cytoplasm formed a thin layer around the nucleus (Figure 1 V–Y).
Non-adherent haemocyte morphology—electron microscopy
The morphological features of Babylonia areolata haemocytes using a transmission electron microscope were previously described for light microscopy, and again the two haemocyte types could be seen.
GRANULOCYTES
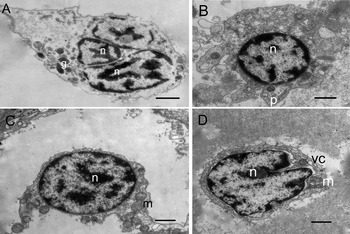
The granulocytes had abundant electron-dense cytoplasmic particles surrounded by membranes, that is, cytoplasmic granules, with diameters between 0.2 and 1.0 µm. The cytoplasm contained a variable number of mitochondria, the Golgi complex, endoplasmic reticulum, and small electron-lucid vesicles of different sizes, some of them probably originating in the Golgi complex or the smooth endoplasmic reticulum. Based on the number of granules and the granule shape, there were two types of granulocytes: type I granulocytes (Figure 2 A–C) and type II granulocytes (Figure 2 D, E). Type I granulocytes had large numbers of granules in the cytoplasm, each about 0.5 µm in diameter and oval. Type II granulocytes contained a few granules, of various shapes.

Fig. 2. Electronic microscopy of granulocytes in Babylonia areolata. (A–C) Electron microscopy of type I granulocytes in B. areolata, spherical or oval cells containing many large oval granules, 0.3–0.6 µm in diameter with protuberances from their external surface that form filopodia; (A) granulocytes with asymmetrical shape; (B, C) portion of granulocytes with organelles gathered around the nucleus and a wide cortical region. Vacuole (vc); rotund or oval granule (gv); nucleus (n); mitochondria (m); pseudopodia (p); rough endoplasmic reticulum (rer); smooth endoplasmic reticulum (ser), the letters represent the same meaning in following figure; (D, E) electronic microscopy of type II granulocytes in B. areolata; (D) type II granulocytes, oval and small nucleus; (E) portion of granulocytes showing peripheral zone of cytoplasm filled with dense various types of granules. Golgi complex (ga); bacilliform granule (gb); tubules (t). Scale bar: A–E 1 µm.
HYALINOCYTES
The hyalinocytes had no cytoplasmic granules, and the nucleus was either in a central or an eccentric position. The cytoplasm contained a variable number of mitochondria and small electron-lucid vesicles of different sizes (Figure 3 A, B). The haemocytes with a large nucleus, a small amount of cytoplasm containing a large number of mitochondria, belonged to the small hyalinocytes (Figure 3 C, D).

Fig. 3. Electron microscopy of hyalinocytes in Babylonia areolata. (A–D) Electron transmission microscopy of hyalinocytes in B. areolata. Hyalinocytes with asymmetrical shape, pseudopodia can be observed in some hyalinocytes, they have one or several nucleus, and a cytoplasm containing few or no granules, the nucleus was either in a central or an eccentric position; (A, B) large hyalinocytes, haemocyte with large nucleus, a small amount of cytoplasm, a small number of vacuoles and mitochondria in the cytoplasm; (B) hyalinocytes showing pseudopodia; (C, D) cells with a large nucleus, containing a great number of mitochondria, are small hyalinocytes; granule (g). Scale bar: A–D =1 µm.
Cell counts and size measurement
The diameter of 1550 haemocytes was measured (we planned to measure the size of 1600 cells, 16 snails and 100 cells per snail; each of the five snails was just measuring 90 cells/individual so the result was 1550 cells), and the distribution of haemocyte diameters of Babylonia areolata was divided into three ranges: <6.2 µm, 6.2 µm–7.4 µm and >7.4 µm; the respective numbers of haemocytes were 192, 629, and 726 respectively. The mean cell diameter and N/C ratio of haemocytes in the three different ranges are shown in Table 1.
Table 1. Number and size of Babylonia areolata haemocytes.

Small hyalinocytes accounted for about 3.15% of circulating haemocytes and displayed a high N/C size-ratio (0.69 ± 0.13). Large hyalinocytes were intermediate sized cells with intermediate N/C ratio (0.59 ± 0.10) and large hyalinocytes accounted for about 37.39% of circulating haemocytes. Granulocytes had large cells and a low N/C ratio (Table 2).
Table 2. Microscopic characterization of the haemocyte populations, mean values ± standard error, and ranges of cell and nucleus diameter and N/C ratio of Babylonia areolata haemocytes.

C, cell diameter; N, nucleus diameter. Different letters in same row show extremely significant difference (P < 0.01) among haemocyte population. Cells and nucleus diameters of type II granulocytes were measured using transmission electron microscopy.
The results of the ANOVA demonstrate a significant difference in cell size, nucleus size and N/C ratio (P < 0.01) between the haemocyte types (Table 2). Granulocytes had larger cell diameters, smaller nucleus diameters and a smaller N/C ratio than hyalinocytes. Nucleus diameter and N/C ratio of type I granulocytes and type II granulocytes were statistically (ANOVA) not different.
Histological study
In B. areolata, we examined a tissue slice of the digestive gland and the alimentary tract. The tissues were stained with Ehrlich's haematoxylin and eosin (HE) by routine protocol to study the general tissue (Figure 4 A–E). Haemocytes of B. areolata occur in the connective tissue (tissue haemocytes) as single cells (Figure 4 B, C), in small groups or in large accumulations. The small groups were seen to be randomly scattered in the connective tissue throughout the visceral mass (e.g. the connective tissue between the hepatopancreas and the alimentary tract; Figure 4D). Large accumulations of haemocytes are present in the connective tissue around the hepatopancreas (Figure 4E).

Fig. 4. Paraffin sections of the digestive gland and the alimentary tract in Babylonia areolata. (A–E) Paraffin sections of the digestive gland and the alimentary tract in B. areolata; (A) transverse section of the alimentary tract: ×50; (B, C) single cells in the connective tissue (arrows): ×400; (D) the small groups in the connective tissue between hepatopancreas and the alimentary tract (arrows): ×400; (E) large accumulations of haemocytes are present in the connective tissue around the hepatopancreas (arrows): ×400; connective tissue (ct); the inner epithelia of the alimentary tract (ie); hepatopancreas (l). Scale bar: A = 240 µm; B–E = 30 µm.
The relationships between the concentration of haemocytes and the snail shell length and shell weight
There were significant differences in shell length (P < 0.05) and weight (P < 0.05) among the three size-groups of the snails, but there was no significant difference in the haemocyte concentration among the three groups. The relationship between concentration of the haemocytes and the shell length and weight is summarized in Table 3. The concentration of haemocytes in the medium sized snails was similar to that in the small sized snails, and it did not increase as the shell length increased.
Table 3. Statistical analysis of shell length, weight and concentration of haemocytes.

L shows shell length; G shows weight; C shows concentration of haemocytes. Different letters in same row or same column show significant difference.
DISCUSSION
No single taxonomic system has been widely accepted for gastropod haemocyte classification, probably due to the absence of specific definitions for the gastropod haemocytes and the different morphological features used to designate cell types. Although haemocyte nomenclature has not yet been standardized, two main schemes are broadly followed for gastropod haemocyte classification. The first was contributed by Cuénot (Reference Cuénot1891), who characterized three types of gastropod haemocytes, namely finely granular, coarsely granular and lymphocyte-like haemocytes. The second scheme simply separates gastropod haemocytes into granulocytes and hyalinocytes (Takatsuki, Reference Takatsuki1934).
Hyalinocytes are agranulocytes, which have a large nucleo-cytoplasmic ratio. They have prominent clear zones in the cytoplasm under light microscopy, and are generally surrounded by a thin rim of scanty cytoplasm with none or a few cytoplasmic granules (Cheng, Reference Cheng1975, Reference Cheng, Ratcliffe and Rowley1981; Hine, Reference Hine1999). Similar findings are observed in other gastropods, viz. Biomphalaria glabrata, Lymnaea stagnalis, Bulinus natalensis, Achatina fulica, Achatina achatina and Planorbarius corneus (Ottaviani, Reference Ottaviani1992); Helix aspersa (Adema et al., Reference Adema, Harris and Van Deutekom-Mulder1992); Clithon retropictus (Kumazawa et al., Reference Kumazawa, Tanigawa, Tanaka, Osatake and Tanaka1990); Trachea vittata, Pila globosa and Indoplanorbis exustus (Mahilini & Rajendran, Reference Mahilini and Rajendran2008); and Haliotis discus discus and Turbo cornutus (Donaghy et al., Reference Donaghy, Hong, Lambert, Park, Shim and Choi2010).
In this study, two types of hyalinocytes can be distinguished by cell size and N/C ratio: large hyalinocytes and small hyalinocytes. Small hyalinocytes have similar characteristics with the blast-like cells in abalone Haliotis tuberculata (Travers et al., Reference Travers, Mirella Da Silva, Le Goïc, Marie, Donval, Huchette, Koken and Paillard2008; Donaghy et al., Reference Donaghy, Hong, Lambert, Park, Shim and Choi2010). Small hyalinocytes should be blast-like cells. Blast-like cells are already reported in snails, Biomphalaria tenagophila (Barracco et al., Reference Barracco, Steil and Gargioni1993) and Lymnaea truncatula (Monteil & Matricon-Gondran, Reference Monteil and Matricon-Gondran1993) and in periwinkle, Littorina littorea (Gorbushin & Iakovleva, Reference Gorbushin and Iakovleva2006). In Tapes philippinarum, small hyalinocytes are suggested as stem cells (blastocytes) because of their morphology and immunocrossreactivity with an anti-human CD34 antibody that identified haematopoietic cells in mammals (Cima et al., Reference Cima, Matozzo, Marin and Ballarin2000).
The nuclei of amoebocytes differ obviously in shape from oval and round to kidney-shape and lobulated (Sminia, Reference Sminia1972). Since a variation in nucleus shape may be an indication of the age of the cell in vertebrate blood cells, this might also be the case in B. areolata, i.e. young cells have round nuclei and older ones have kidney-shape or lobulated nuclei (Sminia, Reference Sminia1974). In this study, we investigated cells that have round nuclei (Figure 1V, W) and cells that have kidney-shape (Figure 1I) or lobulated nuclei (Figure 1K). The small hyalinocytes N/C ratio is quite similar to that reported for blast-cells by other authors (Travers et al., Reference Travers, Mirella Da Silva, Le Goïc, Marie, Donval, Huchette, Koken and Paillard2008; Donaghy et al., Reference Donaghy, Hong, Lambert, Park, Shim and Choi2010). The results suggest that small hyalinocytes might be blast-like cells.
The granulocyte cytoplasm has a peripheral zone filled with dense granules of various types; granulocytes were reported in some species including the terrestrial snail Helix aspersa maxima (Adamowicz & Bolaczek, Reference Adamowicz and Bolaczek2003), the abalone H. asinina (Sahaphong et al., Reference Sahaphong, Linthong, Wanichanon, Riengrojpitak, Kangwanrangsan, Viyanant, Upatham E., Pumthong, Chansue and Sobhon2001), the freshwater snails B. glabrata (Matricon-Gondran & Letocart, Reference Matricon-Gondran and Letocart1999a, Reference Matricon-Gondran and Letocartb), Biomphalaria tenagophila (Barracco et al., Reference Barracco, Steil and Gargioni1993), and P. globosa and I. exustus (Mahilini & Rajendran, Reference Mahilini and Rajendran2008). In the present study, granulocytes were divided into type I granulocytes and type II granulocytes. Type II granulocytes were similar to those found in a number of invertebrates including bivalves or to the numerous peroxidase granules in the haemocytes of Lymnaea stagnalis (Sminia et al., Reference Sminia, Van Der Knaap and Boerrigter-Barendsen1982) or Lymnaea truncatula (Monteil & Matricon-Gondran, Reference Monteil and Matricon-Gondran1993).
No granular haemocytes were described in some gastropod species (Travers et al., Reference Travers, Mirella Da Silva, Le Goïc, Marie, Donval, Huchette, Koken and Paillard2008) using flow cytometry and electron microscopy, suggesting that granulocytes did not exist in the abalone Haliotis tuberculata. In other gastropods, no granular haemocytes were found including marine gastropods such as the abalone H. diversicolor (Chen et al., Reference Chen, Yang, Peng, Chen and Tsai1996), the common periwinkle L. littorea (Gorbushin & Iakovleva, Reference Gorbushin and Iakovleva2006), the sea hare Aplysia californica and the giant keyhole limpet Megathura crenulata (Martin et al., Reference Martin, Oakes, Tousignant, Crabtree and Yamakawa2007), and the disc abalone Haliotis discus discus (Donaghy et al., Reference Donaghy, Hong, Lambert, Park, Shim and Choi2010). A classification scheme by cellular activities might represent an alternative.
Haemocyte subpopulations can also be defined based on surface determinants recognized either by lectins (Schoenberg & Cheng, Reference Schoenberg and Cheng1980), or by monoclonal antibodies (Yoshino & Granath, Reference Yoshino and Granath1985). Therefore, the classification of gastropod haemocytes might consider comprehensive factors, not only morphological and behavioural criteria. In molluscs, making use of specific antibodies and gene probes for the confirmation of haemocyte subpopulations and locations are essential steps for reliable analysis of immunological systems in the future (Jing & Wenbin, Reference Jing and Wenbin2005).
The haemocyte concentration of the B. aveolate has low correlations with the shell length and weight. The haemocyte concentration is similar among the snails of different size-classes. Perhaps the concentration is determined by other factors, such as the activity of the snail, the degree of the food abundance and environmental factors. Environmental factors are known to affect the number of molluscan haemocytes in circulation. For example, exposure to higher temperatures rapidly increases haemocyte numbers (Davies & Partridge, Reference Davies and Partridge1972). This requires further study.
In conclusion, this paper presents analysis of B. areolata haemocytes using cell measurements and light and electron microscopy. Cell size and cells stained by Wright's stain were observed showing two types of haemocytes (hyalinocytes and granulocytes) with different size, colour and relative abundance. These results were consistently tested by electron microscopy, calculation of the N/C ratio. Two subtypes were distinguishable amongst hyalinocytes: small hyalinocytes and large hyalinocytes. Small hyalinocytes shoud be blast-like cells. Two subtypes were distinguishable amongst granulocytes: type I granulocytes and type II granulocytes. Snails with a shell length from 2.7 to 3.3 cm showed no differences in the abundance of haemocytes.
ACKNOWLEDGEMENTS
We are grateful to Professor John Hodgkiss for his help with English. This work was supported in part by the Earmarked Fund for Modern Agro-industry Technology Research System (No. nycytx-47) and Research Project of Technical Exploitation of Fujian Province (No. 98-Z-8).