



Introduction
The octopus stocks that occupy the western and northern coasts of Yucatan Peninsula are considered by far one of the most important resources for small-scale fishers from Mexico due to their high productivity, economic value and international demand (Cabrera-Vázquez et al., Reference Cabrera-Vázquez, Ramos-Miranda and Salas2012). Records show that two species are exploited, namely red octopus, Octopus maya, Voss & Solís-Ramírez, 1966 and the common octopus, O. ‘vulgaris’ type I (Cuvier, 1797; Jereb et al., Reference Jereb, Roper, Norman and Finn2014). However, new studies suggest that the latter corresponds to O. insularis (Lima et al., Reference Lima, Berbel-Filho, Leite, Rosas and Lima2017).
Octopus maya contributes more than 60% to the fishing production of octopus in the region (Velázquez-Abunader et al., Reference Velázquez-Abunader, Salas and Cabrera2013). It is an endemic species of the continental shelf of the Yucatan Peninsula. It has been observed to be abundant both in shallow (<30 m) and deeper waters (up to 60 m), but more abundant in shallow waters (DOF, 2016). The species displays a heterogeneous distribution, having the greatest abundance in the coasts alongside the State of Campeche, predominantly composed of small individuals, while the largest individuals are found alongside the State of Yucatan (Cabrera-Vázquez et al., Reference Cabrera-Vázquez, Ramos-Miranda and Salas2012; Gamboa-Álvarez et al., Reference Gamboa-Álvarez, López-Rocha and Poot-López2015). A more recent study suggests that perhaps two closely related sub-stocks of O. maya exist in the region: the first occupies the western coast of the Yucatan Peninsula, where reproduction exhibits a clear seasonality with a peak during the winter, and a second stock is located at the north of the Yucatan peninsula, where spawners can be found all year round (Ángeles-González et al., Reference Ángeles-González, Calva, Santos-Valencia, Avila-Poveda, Olivares, Diaz and Rosas2017).
The most recent stock assessment indicates that O. maya is exploited at the ‘maximum level’ (i.e. close to the maximum sustainable yield) with annual landings of more than 10,000 tons (Jurado-Molina, Reference Jurado-Molina2010). In order to maintain production levels, the authority established a minimum legal size of 11 cm mantle length, a closed fishing season (from January to July), and total allowable catch (TAC). The TAC is obtained from biomass estimations using surplus production models based on the catch landings reports (DOF, 2016).
Octopus maya is captured by two fleets: a small-scale fleet (boats of 5–12 m length) that operates in shallow waters (up to 20 m depth) and a medium-scale fleet (boats of 15–25 m length) that operates in areas deeper than 20 m. Both fleets use small boats 4 m in length (locally known as alijos) which are drifted by the currents to catch octopus (Salas et al., Reference Salas, Cabrera, Palomo, Bobadilla, Ortega and Torres2008). These fleets use the same fishing gears and operate in different fishing grounds but sometimes overlap due to the accessibility and high abundance of the resource in those areas (Salas et al., Reference Salas, Cabrera, Palomo, Bobadilla, Ortega and Torres2008; Gamboa-Álvarez et al., Reference Gamboa-Álvarez, López-Rocha and Poot-López2015). Likewise, as a result of easy access and low monitoring costs, most of the studies on biology and stock assessment for O. maya refers to animals found in the shallow waters of those fishing grounds (<30 m) (Cabrera-Vázquez et al., Reference Cabrera-Vázquez, Ramos-Miranda and Salas2012; Velázquez-Abunader et al., Reference Velázquez-Abunader, Salas and Cabrera2013; Avila-Poveda et al., Reference Avila-Poveda, Koueta, Benítez-Villalobos, Santos-Valencia and Rosas2016; Ángeles-González et al., Reference Ángeles-González, Calva, Santos-Valencia, Avila-Poveda, Olivares, Diaz and Rosas2017; Duarte et al., Reference Duarte, Hernández-Flores, Salas and Seijo2018), however, there is a dearth of information on the fraction of the population that occupies areas from 30 m to 60 m depth. The private sector of Mexico has expressed its intention to expand the fishing grounds for the medium-scale fleet to deeper waters in view of its economic importance (DOF, 2016). It is for that reason that the objective of this study is to evaluate the available biomass of O. maya and learn more about its distribution in coastal areas in the Campeche Bank where the depth is between 30 m and 60 m, to provide basic information for its management.
Materials and methods
Study area
The study area, known as the Campeche Bank, is located in the coastal zone at the north-east of the Yucatan Peninsula, between 30 m and 60 m depth (Figure 1). The area is strongly influenced by the Yucatan current, which produces a stationary upwelling, from May to September, but there is vertical mixing during winter due to strong north winds from 70 km h−1 to more than 100 km h−1 (from October to January) (Enriquez et al., Reference Enriquez, Mariño-Tapia and Herrera-Silveira2010; Salas-Pérez et al., Reference Salas-Pérez, Salas-Monreal, Monreal-Gómez, Riveron-Enzastiga and Llasat2012). The average temperature is 20°C with a range of 17–30°C. The upwelling enhances the concentration of nutrients resulting in a high biological productivity.

Fig. 1. Study area for fishing of the red octopus (Octopus maya) to the east of Campeche Bank, Mexico.
Fieldwork
Four research cruises independent of the fishery were conducted from May 2016 to January 2017. Each cruise was made on board a vessel of the medium-scale fleet with landing port in Progreso, Yucatan. An average of 29 (±2) sampling sites were surveyed per ship; the distance between sampling sites was 28 km in May–June, and 14 km in the other cruises (Figure 1). Sampling sites were systematically aligned in the study area, using spsample function of sp package (Pebesma & Bivand, Reference Pebesma and Bivand2005) of the programming language R (R Core Team, 2017). During the season closed for fishing, two cruises were carried out, May–June 2016 and July 2016, just when the fishing season started. Two additional cruises were made in December 2016 and January 2017, to represent the end of the fishing season.
The survey and collection of organisms were done through regular fishing operations. The vessel was a mother ship of five alijos (4 m length); each carrying two rustic poles made of bamboo of ~8 m length, one in the bow and the other in the stern of the boat. Each pole had two nylon lines tied with fishes (Diplectrum sp. and Haemulon sp.) as bait, which were dragged along the sea floor as the boat drifted at sea (Jurado-Molina, Reference Jurado-Molina2010; Velázquez-Abunader et al., Reference Velázquez-Abunader, Salas and Cabrera2013; Gamboa-Álvarez et al., Reference Gamboa-Álvarez, López-Rocha and Poot-López2015; Markaida et al., Reference Markaida, Méndez-Loeza and Rosales-Raya2017). Each alijo had a global positioning system (GPS) to track the course and thus measure the swept area. The initial and final times were recorded to standardize the effective fishing effort in three hours and the sampling effort in five alijos per sampling site per day. In each sampling site, the total number of octopuses captured (N t), the total weight of the catch (TW) and the individual weight of octopuses (W i) were recorded.
Area of influence of sampling sites
In order to have a better approach to the potential area of influence of each sampling site, Thiessen (or Voroni) polygons were deployed (Brassel & Reif, Reference Brassel and Reif1979), to calculate the area of each polygon and, finally, obtain the representative area of each sampling site in relation to the total sample area. Thiessen polygons and the area of each polygon were calculated with ArcMap 9.2 software (Sawatzky et al., Reference Sawatzky, Raines, Bonham-Carter and Looney2009).
Biomass assessment
Four methods were used to calculate the O. maya biomass per research cruise: stratified random method (Cochran, Reference Cochran1980; Scheaffer et al., Reference Scheaffer, Mendenhall and Ott1987), swept area method (Pierce & Guerra, Reference Pierce and Guerra1994), geostatistical biomass model (Rivoirard et al., Reference Rivoirard, Simmonds, Foote, Fernandes and Bez2008), and an unpublished method of weighted swept area, the advantage of which is that it does not assume a priori homogeneous distribution of the resource in the whole area, as the traditional swept area method does (Pierce & Guerra, Reference Pierce and Guerra1994).
The stratified random method uses the frequencies distribution of total weight of the catch, which is classified by strata (Cochran, Reference Cochran1980). This method requires to calculate the number of strata (expressed in kg) by means of the Sturges rule (Nevárez-Martínez et al., Reference Nevárez-Martínez, Hernández-Herrera, Morales-Bojórquez, Balmori-Ramírez, Cisneros-Mata and Morales-Azpeitia2000) which calculates the number of intervals of the catch, starting from the minimum and maximum catches recorded in each cruise. Equations to calculate biomass were the following. The average counting (expressed in kg) in the i th stratum (ȳ i) was:
 $$\bar{y}_i = \displaystyle{1 \over N}\mathop \sum \limits_\; ^i y_{\,ji}$$
$$\bar{y}_i = \displaystyle{1 \over N}\mathop \sum \limits_\; ^i y_{\,ji}$$The variance estimator for ȳ i:
 $${\hat V}\lpar {{\bar{y}}_i} \rpar = s_i^2 = \displaystyle{1 \over {N_i}}\mathop \sum \limits_{\,j = 1}^L (y_{\,ji}-\bar{y}_i)^2$$
$${\hat V}\lpar {{\bar{y}}_i} \rpar = s_i^2 = \displaystyle{1 \over {N_i}}\mathop \sum \limits_{\,j = 1}^L (y_{\,ji}-\bar{y}_i)^2$$The estimator of the total size of the population expressed in kg:
 $$N\bar{y}_{{\rm st}} = \mathop \sum \limits_{i = 1}^L N_i\bar{y}_i$$
$$N\bar{y}_{{\rm st}} = \mathop \sum \limits_{i = 1}^L N_i\bar{y}_i$$The variance estimator for the total population size 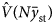 ${\hat V}\lpar {N{\bar{y}}_{{\rm st}}} \rpar $:
${\hat V}\lpar {N{\bar{y}}_{{\rm st}}} \rpar $:
 $${\hat V}\lpar {N{\bar{y}}_{{\rm st}}} \rpar = \mathop \sum \limits_{i = 1}^L N_i^2 \left( {\displaystyle{{N_i-n_i} \over {N_i}}} \right)\left( {\displaystyle{{S_i^2} \over {n_i}}} \right)$$
$${\hat V}\lpar {N{\bar{y}}_{{\rm st}}} \rpar = \mathop \sum \limits_{i = 1}^L N_i^2 \left( {\displaystyle{{N_i-n_i} \over {N_i}}} \right)\left( {\displaystyle{{S_i^2} \over {n_i}}} \right)$$The confidence interval (P = 0.95) for the population size:
 $$N\bar{y}_{{\rm st}} \pm 2\sqrt {\mathop \sum \limits_{i = 1}^L N_i^2 \left( {\displaystyle{{N_i-n_i} \over {N_i}}} \right)\left( {\displaystyle{{S_i^2} \over {n_i}}} \right)} $$
$$N\bar{y}_{{\rm st}} \pm 2\sqrt {\mathop \sum \limits_{i = 1}^L N_i^2 \left( {\displaystyle{{N_i-n_i} \over {N_i}}} \right)\left( {\displaystyle{{S_i^2} \over {n_i}}} \right)} $$where N i is the total number of sampled units (km2) in the ith stratum, L is the number of strata, n i is the number of sampling units (km2) in the ith stratum, y i is the average weight in the ith stratum, and S i2 is the variance of the counting in the ith stratum.
The swept area method considers the catch in weight (biomass) obtained from the area swept by the alijos, assuming a homogeneous distribution of the resource in the study zone, with a single estimate for the whole area sampled.
Total biomass (B T) was calculated with the next equation (Pierce & Guerra, Reference Pierce and Guerra1994):
 $$B_{\rm T}{\rm \;} = \mathop \sum \limits_{i = 1}^n \left( {Y_t\displaystyle{{A_t} \over {a_t}}} \right)$$
$$B_{\rm T}{\rm \;} = \mathop \sum \limits_{i = 1}^n \left( {Y_t\displaystyle{{A_t} \over {a_t}}} \right)$$with variance:
 $${\hat V}\lpar {B_{\rm T}} \rpar = \mathop \sum \limits_{i = 1}^n \left( {\displaystyle{{A_t^2 m_tS_t^2} \over {a_t^2}}} \right)$$
$${\hat V}\lpar {B_{\rm T}} \rpar = \mathop \sum \limits_{i = 1}^n \left( {\displaystyle{{A_t^2 m_tS_t^2} \over {a_t^2}}} \right)$$where Y t is the total catch in the study area, A t is the total area of study, a t is the cumulated area swept by the five alijos, S t2 is the variance of the total catch in the study area, m t is the number of fishing trials and 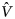 ${\hat V}$(B T) is the variance of the total biomass. In this case, a i represented the area swept by the ith alijo. Therefore, the total swept area a t (expressed in km2) for each fishing trial was calculated as:
${\hat V}$(B T) is the variance of the total biomass. In this case, a i represented the area swept by the ith alijo. Therefore, the total swept area a t (expressed in km2) for each fishing trial was calculated as:
 $$a_t{\rm \;} = \mathop \sum \limits_{i = 1}^5 a_i$$
$$a_t{\rm \;} = \mathop \sum \limits_{i = 1}^5 a_i$$a i was calculated with the following equation:
 $$a_i = D_i\; \times \; LJ_i$$
$$a_i = D_i\; \times \; LJ_i$$where D i is the distance travelled by the ith alijo, obtained from the track recorded by the GPS and LJ i is the length between the extreme tips of the ith alijo's bamboo poles (LJ i = 8 m). Finally, total abundance (N T) for each cruise ship was calculated with the equation:
 $$N_{\rm T} = \displaystyle{{B_{\rm T}} \over {\overline {{\rm TW}}}} $$
$$N_{\rm T} = \displaystyle{{B_{\rm T}} \over {\overline {{\rm TW}}}} $$where 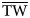 $\overline {{\rm TW}} $ is the average weight of the octopus as obtained from the biological sampling. For the estimation of B T the assumptions were the same as for the swept area method (details of the method are contained in Csirke, Reference Csirke1989).
$\overline {{\rm TW}} $ is the average weight of the octopus as obtained from the biological sampling. For the estimation of B T the assumptions were the same as for the swept area method (details of the method are contained in Csirke, Reference Csirke1989).
In order to estimate the biomass using the geostatistical biomass model, we proceeded to calculate the catch per unit of area (CPUA, expressed in number of octopuses per km2), obtained by dividing the number of octopuses captured by the corresponding area at each sampling site. The spatial correlation of CPUA was calculated by means of omnidirectional empirical variograms, which measure the correlation between the variance generated by all the differences of the data pairs separated by a distance previously established, with that distance (h) (Hernández-Flores et al., Reference Hernández-Flores, Condal, Poot-Salazar and Espinoza-Mendez2015). Thereafter, a kriging interpolation technique was applied to obtain the densities throughout the interpolation nodes between the neighbouring values (Cressie, Reference Cressie1992) and produce a spatial structure that depends on the spatial arrangement of the population (Webster & Oliver, Reference Webster and Oliver2007).
The empirical variograms were obtained with the equation:
 $$\lpar h \rpar = \displaystyle{1 \over {2N\lpar h \rpar }}\mathop \sum \limits_{i = 1}^{N\lpar h \rpar } \lsqb {C(x_i)-C(x_i + h)} \rsqb ^2\; $$
$$\lpar h \rpar = \displaystyle{1 \over {2N\lpar h \rpar }}\mathop \sum \limits_{i = 1}^{N\lpar h \rpar } \lsqb {C(x_i)-C(x_i + h)} \rsqb ^2\; $$where γ(h) is the variance for h distance, N(h) is the number of paired observations separated by distance h, C(x i) is the CPUA observed at site x i and C(x i + h) is the CPUA observed at any another site separated h distance from site x i. The obtained interpolations were divided into CPUA intervals, obtaining an average value for the ith interval (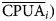 $\overline {{\rm CPUA}} _i)$. The total abundance of the ith interval (Ni) was obtained from multiplying the (
$\overline {{\rm CPUA}} _i)$. The total abundance of the ith interval (Ni) was obtained from multiplying the (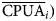 $\overline {{\rm CPUA}} _i)$ by total area covered by the ith interval, so the total abundance (N T) was obtained with the equation:
$\overline {{\rm CPUA}} _i)$ by total area covered by the ith interval, so the total abundance (N T) was obtained with the equation:
 $$N_{\rm T} = \mathop \sum \limits_{i = 1}^n \overline {{\rm CPUA}} _i\; \times A_i$$
$$N_{\rm T} = \mathop \sum \limits_{i = 1}^n \overline {{\rm CPUA}} _i\; \times A_i$$and the biomass was obtained with the equation:
 $$B_{\rm T} = N_{\rm T}\overline {{\rm TW}} $$
$$B_{\rm T} = N_{\rm T}\overline {{\rm TW}} $$The weighted swept area method, proposed in this study, consisted in analysing the catches registered by the five alijos that operated at every ith sampling site (a t) as the only data for that site. The total biomass was obtained by adding the individual biomass estimated in each sampling site. Thus, the biomass was obtained with the next equation:
 $$B_{\rm T}{\rm \;} = \mathop \sum \limits_{i = 1}^n Y_i\left( {\displaystyle{{A_i} \over {a_i}}} \right)$$
$$B_{\rm T}{\rm \;} = \mathop \sum \limits_{i = 1}^n Y_i\left( {\displaystyle{{A_i} \over {a_i}}} \right)$$With standard deviation:
 $${{\widehat {\rm SD}}\left( {B_{\rm T}} \right)} = \; {\sqrt {\mathop \sum \nolimits {\left( {Y_i-\bar{Y}} \right)}^2}} \; \; \; \left( {\displaystyle{{A_i} \over {a_i}}} \right)$$
$${{\widehat {\rm SD}}\left( {B_{\rm T}} \right)} = \; {\sqrt {\mathop \sum \nolimits {\left( {Y_i-\bar{Y}} \right)}^2}} \; \; \; \left( {\displaystyle{{A_i} \over {a_i}}} \right)$$where Y i is the total catch in the ith stratum, Y is the average catch in the study area, A i is the total area in the ith stratum, a i is the swept area in that stratum and 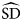 $\widehat{{{\rm SD}}}$(B T) is the standard deviation of total biomass. Abundance was again calculated with equation (10).
$\widehat{{{\rm SD}}}$(B T) is the standard deviation of total biomass. Abundance was again calculated with equation (10).
For the interpretation of the weighted swept area method, it was necessary to modify the assumption of densities homogeneity, so the total catch Y i of the distribution area A i was specific for every sampling site. Another assumption was that each alijo had the same probability of catching the octopus at a fixed radius of action such that the sampling effort could be extrapolated to a constant area a. The swept area is considered as the area covered by each alijo drifting at each sampling site. Finally, within the area, each unit of sampling effort has the efficiency at every moment to catch only a fraction of the octopus population.
Spatial distribution pattern
To describe the type of pattern distribution of O. maya, the equation proposed by Guerra (Reference Guerra1981) was modified. The average probability of octopus presence per sampling site was estimated, as well as the type of distribution. Then, the parameters p and k of the negative binomial distribution were estimated.
 $$P\lpar {x/k} \rpar = \; \left( {\displaystyle{{k\lpar {k + 1} \rpar \lpar {k + 2} \rpar \ldots \lpar {k + x-1} \rpar } \over {x!}}} \right)p^xq^k$$
$$P\lpar {x/k} \rpar = \; \left( {\displaystyle{{k\lpar {k + 1} \rpar \lpar {k + 2} \rpar \ldots \lpar {k + x-1} \rpar } \over {x!}}} \right)p^xq^k$$To demonstrate if octopus's distribution was random (i.e. homogeneous in the study area) or if it formed patches (i.e. aggregate in some places), a simple random distribution was created assuming the negative binomial distribution. According to this method, the estimation of the parameter of the negative binomial distribution (k) could be: K 1 = 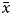 $\bar{x}$2/S2–
$\bar{x}$2/S2–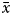 $\bar{x}$, testing some of the following conditions: if
$\bar{x}$, testing some of the following conditions: if 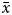 $\bar{x}$ value was low, then K/
$\bar{x}$ value was low, then K/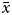 $\bar{x}$>6, if
$\bar{x}$>6, if 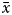 $\bar{x}$ was high then K > 13, and if
$\bar{x}$ was high then K > 13, and if 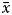 $\bar{x}$ value was moderate then
$\bar{x}$ value was moderate then 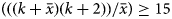 $\lpar {((k + \bar{x})(k + 2))/\bar{x}} \rpar \ge 15$.
$\lpar {((k + \bar{x})(k + 2))/\bar{x}} \rpar \ge 15$.
If none of these conditions occurs, K 1 is inadequate; then, it is calculated with:
 $$K_2{\rm lo}{\rm g}_{10}\left( {1 + \displaystyle{{\bar{x}} \over {K_2}}} \right) = {\rm lo}{\rm g}_{10\;} \left( {\displaystyle{N \over {\,f_0}}} \right)$$
$$K_2{\rm lo}{\rm g}_{10}\left( {1 + \displaystyle{{\bar{x}} \over {K_2}}} \right) = {\rm lo}{\rm g}_{10\;} \left( {\displaystyle{N \over {\,f_0}}} \right)$$in any case, p =  $\; \bar{x}$/K (18)
$\; \bar{x}$/K (18)
Once the parameters were calculated, to verify if the distribution was in patch, a goodness-of-fit test was applied between the distribution function of the total sample and the theoretical negative binomial distribution (Zar, Reference Zar1999).
Results
Biomass
The coefficient of variation (CV) for the biomass obtained with the four methods was lower for the cruise of May–June (CV = 0.12) and higher for January (CV = 0.26). The areas of influence for each sampling site determined by the Thiessen polygons ranged from 60 to 940 km2 with an average of 242 km2. The lower biomass was calculated for the cruise of May–June (47.3 ± 6.8 t), while the highest was estimated for December (141.22 ± 12.7 t) (Table 1). Of the four models, the geostatistical biomass model consistently resulted in the lowest values in the four cruise ships, while the other three methods produced results more alike. This is because geostatistical analysis assumes a heterogeneous distribution pattern generated by the parameters of the semivariogram, through which the minimum size of each pixel is calculated. On the other hand, the other methods extrapolate the average values of biomass to units of areas wider than those of the geostatistical model. The precision of the geostatistical method will depend on how well it represents the real spatial distribution of the abundance within a reduced coverage relative to the other methods. The geostatistical biomass model estimations were between 22% and 47% lower than those of the other models (Table 1). A similar pattern was observed for densities; however, the increases from one month to the next were not as marked as in biomass. The highest densities were recorded in the cruises of May–June and July 2016 (13.4 and 20.5 octopus km2, respectively), while the lowest densities were observed in the cruises of May-June 2016 and January 2017 (7.6 and 10.3 octopus km2, respectively). Similarly, the geostatistical biomass model resulted in the lowest values of density in the four cruises and the weighted swept area method produced the highest values (Table 1).
Table 1. Estimated values of the biomass per cruise ± standard error (SE) and the density ± standard error (SE) of the red octopus (Octopus maya) in the north-eastern Campeche Bank

The biomasses were standardized to a total area of 5000 km2. CV: coefficient of variation of the estimates by cruise of the four methods.
Distribution
The value of the parameters p and k (k 2 = 2, P = 0.5) of the negative binomial distribution showed that O. maya presented a patchy distribution (Figure 2), suggesting that the abundance increases according to distance in a specific area and then begins to decrease at higher distances. This is plausible if we consider that the study area deepens as the latitude increases. So, in the shallower water the abundance increases.

Fig. 2. Semivariogram analysis of the abundances (CPUA; Org. km−2) of the red octopus (Octopus maya) in the north-eastern Campeche Bank, Mexico. The behaviour indicated a grouped type distribution.
The cruises made before the fishing season (May–June and July 2016) recorded the highest densities and abundances in the south and south-west of the study area (Figure 3). The octopuses displayed a heterogeneous distribution throughout the study area with lower CPUE overall in the cruises carried out at the end of the fishing season (December 2016 and January 2017); nevertheless, areas of aggregation continued appearing in the analysis, although with lower densities than in May–June and July of 2016 (Figure 3).

Fig. 3. Spatiotemporal distribution of the Catch Per Unit Area (CPUA: Org. km−2) of the red octopus (Octopus maya) in the north-eastern Campeche Bank, Mexico.
Discussion
Many cephalopod fisheries are managed through total allowable catches, which are usually based on the evaluation of the biomass before the start of each fishing season (Nevárez-Martínez et al., Reference Nevárez-Martínez, Hernández-Herrera, Morales-Bojórquez, Balmori-Ramírez, Cisneros-Mata and Morales-Azpeitia2000). This is the case of O. maya, although frequently the TAC is exceeded in some seasons (Jurado-Molina, Reference Jurado-Molina2010). This is mainly due to their reproductive strategies that in many cases are semelparous, as well as their short longevity and rapid growth. These biological characteristics mean that the populations consist of intra-annual cohorts that are replaced year after year (Hernández-Herrera et al., Reference Hernández-Herrera, Morales-Bojórquez, Cisneros-Mata, Nevárez-Martínez and Rivera-Parra1998; Arreguín-Sánchez et al., Reference Arreguín-Sánchez, Solís-Ramírez and González de la Rosa2000). That is why it is important to calculate the biomass of exploited cephalopods at different times during the fishing season, since this will reveal the stock size, recruitment periods and the time when the biomass increases (i.e. stock reduction analysis and proportional escapement analysis) (Rosenberg et al., Reference Rosenberg, Kirkwood, Crombie and Beddington1990).
This was the first study to determine the biomass and distribution of O. maya carried out in the north-eastern zone of the Campeche Bank between 30 m and 60 m depth. Most techniques to calculate biomass use catch and fishing effort data, which are not always available as in the case of the O. maya fishery. However, this study used a systematic sampling design, independent of the fishery, which has the advantage of covering a larger distribution area, controlling the sampling effort (Pierce & Guerra, Reference Pierce and Guerra1994; Hernández-Flores et al., Reference Hernández-Flores, Condal, Poot-Salazar and Espinoza-Mendez2015).
Given that there are no previous studies on the biomass of octopus for the fraction of the population that occurs more than 30 m deep in the study area, this study used four methods to analyse the data, with particular characteristics and assumptions. Our results show that differences in the biomass estimates from each of the four methods (CV <26.5% per cruise) could be biologically relevant and an important consideration for managers (Pierce & Guerra, Reference Pierce and Guerra1994). These differences in the results could be related to factors such as the distribution pattern of the resource and the sampling design; for example, in the swept area method, the weighted swept area method and the geostatistical biomass method, the distance between sampling sites is key so as not to exceed the area of extrapolation per sampling site, while in the stratified method the number of intervals is key in the estimate. It is instructive to apply the Sturges rule from the start of the analysis (Nevárez-Martínez et al., Reference Nevárez-Martínez, Hernández-Herrera, Morales-Bojórquez, Balmori-Ramírez, Cisneros-Mata and Morales-Azpeitia2000). The assumption of heterogeneous distribution of the resource is perfectly applicable to the benthic organisms that remain in the same habitat as long as the conditions are favourable, and that present a patchy distribution, such as was the case of O. maya.
In resources such as the jumbo squid from the Gulf of California, biomass has been calculated through the stratified random method and the swept area method, showing significant differences in the results of both (Nevárez-Martínez et al., Reference Nevárez-Martínez, Hernández-Herrera, Morales-Bojórquez, Balmori-Ramírez, Cisneros-Mata and Morales-Azpeitia2000). These discrepancies were attributed to the type of stratification used in each method, since the randomized method stratified the catch data, while the swept area method stratified the data spatially (Nevárez-Martínez et al., Reference Nevárez-Martínez, Hernández-Herrera, Morales-Bojórquez, Balmori-Ramírez, Cisneros-Mata and Morales-Azpeitia2000). Therefore, in addition to the method, it is important that fisheries managed with total allowable catches apply the precautionary approach considering the most conservative result (Nevárez-Martínez et al., Reference Nevárez-Martínez, Hernández-Herrera, Morales-Bojórquez, Balmori-Ramírez, Cisneros-Mata and Morales-Azpeitia2000), which in the case of O. maya should be applied when estimating in the fishing grounds. This precautionary approach should be applied in the areas with the greatest fishing effort.
The distribution of O. maya has not been thoroughly studied; most studies have covered the immediately coastal zone with the highest concentration of octopuses between 0 and 30 m depth. Some studies have suggested that the O. maya has a heterogeneous distribution in the shallow waters of the Campeche Bank (<30 m depth) (Solís-Ramírez & Chávez, Reference Solís-Ramírez and Chávez1986; Gamboa-Álvarez et al., Reference Gamboa-Álvarez, López-Rocha and Poot-López2015) as a response to changes in the environment such as the effect of the wind during winter or the type of substratum. Cephalopods are organisms highly sensitive to environmental changes, so they will carry out active migrations in search of favourable conditions to continue their life cycle (Pierce et al., Reference Pierce, Valavanis, Guerra, Jereb, Orsi-Relini, Bellido, Katara, Piatkowski, Pereira, Balguerias, Sobrino, Lefkaditou, Wang Jianjun, Santurtun, Boyle, Hastie, MacLeod, Smith, Viana, González and Zuur2008). In this study, although in general, O. maya showed a patchy distribution, during the December and January cruises it was more random, with few aggregations of low CPUA values. This type of distribution has been reported by Gamboa-Álvarez et al. (Reference Gamboa-Álvarez, López-Rocha and Poot-López2015) in the shallow waters of the Yucatan Peninsula, probably due to the dynamics of the ocean in the region that includes significant changes in temperature (Enriquez et al., Reference Enriquez, Mariño-Tapia and Herrera-Silveira2010), which is a key factor for the biological processes of the species (Ángeles-González et al., Reference Ángeles-González, Calva, Santos-Valencia, Avila-Poveda, Olivares, Diaz and Rosas2017). In this sense, it has been reported that O. maya has a low capacity to adapt to high variations of temperature, producing a significant negative impact on its survival rate and abundance (Noyola et al., Reference Noyola, Caamal-Monsreal, Díaz, Re, Sanchez and Rosas2013). As shown by Hermosilla et al. (Reference Hermosilla, Rocha and Valavanis2011), there is a negative correlation between sea bottom temperature and abundance of O. vulgaris in the Mediterranean Sea. In consequence, temperature changes limit octopus distribution in deeper waters, which seems to be the origin of the distribution observed in this study.
As occurs in other cephalopods such as inshore squids and some octopod species including the common octopus (O. vulgaris), O. maya shows a great plasticity in its life cycle, which gives it a great ability to adapt to the prevailing conditions where it lives (Pecl & Jackson, Reference Pecl and Jackson2008; Otero et al., Reference Otero, González, Guerra and Álvarez-Salgado2009; Ramos et al., Reference Ramos, Pecl, Moltschaniwskyj, Strugnell, León and Semmens2014), but there are no studies that correlate environmental variables with the biomass and distribution of O. maya. Therefore, it seems that the home range of this species should be well specified, which could be a priority for future research. However, spatial differences in population structure of this species have been evaluated. Authors such as Velázquez-Abunader et al. (Reference Velázquez-Abunader, Salas and Cabrera2013) indicated that the landings of the medium-scale fleet (which fishes in deeper waters than the small-scale fleet) were mainly composed of large organisms, so the stratum of the population that occurs in deeper waters could be composed mostly of mature individuals of the spawning stock. Thus, this fraction of the stock could contribute significantly to the recruitment of the entire population, even to the fraction that is currently exploited (<30 m depth), so it is suggested to avoid the exploitation of this resource in deeper areas. In addition, the methods used in this study could be applied to calculate the biomass in the most intense fishing areas, as long as a stratified sampling design is applied. Therefore, future work should make an assessment of the biomass and distribution of O. maya in shallower fishing areas.
Author ORCID
Iván Velázquez–Abunader, 0000-0003-3216-2007
Acknowledgements
The authors thank the Federación de Sociedades Cooperativas de la Industria Pesquera del Centro y Oeste del estado de Yucatán, SC de RL for the logistical support and with the boat to carry out the cruises. To Jesús Miguel Soto Vázquez for his support in the fieldwork. Thanks are also due to Mr Babatunde O. Musa for proofreading.
Financial support
The results of this study were obtained as part of the project Distribución, reproducción, biomasa y patrones de movimiento del pulpo común Octopus vulgaris Cuvier, 1797 en la costa de Yucatán with financial support of the Consejo Nacional de Ciencia y Tecnología (grant number 237057).




