


INTRODUCTION
Many species of skates and rays have similar life history characteristics to large sharks, i.e. slow growth, late maturity, low reproductive rates and a direct relationship between stock size and recruitment (Ebert & Sulikowski, Reference Ebert and Sulikowski2007). An increase in skate catches may necessitate the implementation of management plans for the sustainable use of these resources (Dulvy & Reynolds, Reference Dulvy and Reynolds2002). However, the development of such fishery management plans is difficult because there are few incentives to collect detailed biological information for skates, particularly those of low value and non-target species.
Recent stock assessments of skates suggest that several species have been overexploited or have even collapsed. Dulvy & Reynolds (Reference Dulvy and Reynolds2002) reported the common skate, Dipturus batis (Linnaeus, 1758) has disappeared in the Irish Sea, while the longnosed skate, D. oxyrinchus (Linnaeus, 1758) and the spear-nose skate, Rostroraja alba (Lacepède, 1803) had smaller distributions than where they were previously recorded. The decline of large skates coupled with an increase in the abundance of small skates have resulted in a structural change in the marine ecosystem (Dulvy et al., Reference Dulvy, Metcalfe, Glanville, Pawson and Reynolds2000). Such major changes clearly indicate the urgent need for species-specific fisheries-related biological information, so as to ensure that skate stocks are exploited in a sustainable way.
The Kwangtung skate, Dipturus kwangtungensis (Chu, 1960) is a medium-sized, demersal species that inhabits tropical and subtropical coastal waters of the western North Pacific Ocean. It is broadly distributed off southern Japan, East China Sea, South China Sea and Taiwanese waters (Ishiyama, Reference Ishiyama1967; Hou, Reference Hou2002). In Taiwan, this species is found in the coastal waters of south-western and northern Taiwan and is one of the most abundant by-catch species of skates in the trawl fishery in those areas, based on our port surveys (Chen C. C., personal observation). According to the catch statistics, the annual yield of batoids in Yilan County, northern Taiwan, increased remarkably from 1993 to 2008 (Fisheries Year Book – Taiwan, 2010) indicating an increase in exploitation or abundance. Unfortunately, species-specific catch statistics for batoids were not available because, with the exception of the large individuals, most of the catch was considered to be of low economic value and regarded as trash fish.
There have been a number of studies of batoids in Taiwanese waters, including a taxonomic study of the genus Raja (Chen & Joung, Reference Chen and Joung1989), reproductive biology of the sepia stingray, Urolophus aurantiacus (Yu, Reference Yu2007) and estimates of life history parameters of the sharpspine skate, Okameiei acutispina (Joung et al., Reference Joung, Lee, Liu and Liao2011). However, biological information on D. kwangtungensis is sparse and limited to a preliminary description of its fishery biology (Hou, Reference Hou2002). Detailed estimates of age and growth which are important for stock assessment and fisheries management, are, however, lacking for this species.
Therefore, the objective of this study was to provide the first detailed information on age and growth of D. kwangtungensis in waters off northern Taiwan. It is anticipated that the growth parameters derived from this study can be used as input parameters for further assessment of the stock in this region.
MATERIALS AND METHODS
Specimen collection
The specimens of D. kwangtungensis were opportunistically collected mainly from the trash fish piles at Tashi fish market, Yilan County, north-eastern Taiwan, on a monthly basis (Table 1). The specimens were caught by commercial trawl fishing in the waters off northern Taiwan between July 2006 and July 2008 (Figure 1). All specimens were taken back to the laboratory for further analysis.

Fig. 1. Sampling area (shaded) of Dipturus kwangtungensis in this study. Dashed lines indicate the isobaths.
Table 1. Size ranges and sex ratios of Dipturus kwangtungensis used in this study. Sex ratio is (the number of males)/(the number of females).

*Significant at 5% level using χ2 test.
Body metrics and sex ratio
Measurements of the specimens were taken of total length (TL), disc width (DW) and body weight (W in g) and the sex was identified. Inner clasper length (CL) was measured to the nearest 0.1 cm for males. A simple linear regression was used for describing the relationship between DW and TL and a general linear model (GLM) (Neter et al., Reference Neter, Wasserman and Kutner2005) was used to compare these relationships between sexes. The sex ratio was expressed as the ratio of the number of males to the number of females. A Chi-square test (χ2, Zar, Reference Zar2010) was used to examine the homogeneity of the sex ratio.
Age determination
The subsample arbitrarily selected from each length interval (5 cm TL), based on its proportion to the whole sample, was used for ageing analysis. Prior to this study, a non-invasive structure – caudal thorns – was examined, but failed to provide clear band patterns and could not therefore be used. Therefore, the vertebral centra, which provided clear band patterns, were used for age determination. To select the location of the vertebral column with most consistent band pair reading and minimum vertebral centrum diameter variation, six specimens (three females, 41, 38 and 38.3 cm TL; three males, 44.4, 45.5 and 52.1 cm TL) were used to compare variations in banding patterns on the vertebral centra from different locations along the vertebral column of the specimens. Vertebrae taken from each specimen were soaked in 5% KOH for 60 min to remove connective tissue, washed in running water for 24 h, and then rinsed in 95% alcohol for 24 h and air dried. The diameter of each vertebral centrum (D) was measured to the nearest 0.1 mm. The coefficient of variation (V) on the diameter of the vertebral centrum was calculated every three consecutive vertebrae as a group using the formula:
 $V = (S/\overline X ) \times 100\% $
, where S is the standard deviation of the diameters of three consecutive vertebral centra,
$V = (S/\overline X ) \times 100\% $
, where S is the standard deviation of the diameters of three consecutive vertebral centra,
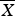 $\overline X $
is the mean diameter of three consecutive vertebral centra. The analysis revealed that the 19th to the 21th vertebrae (beginning of the abdominal cavity) had the smallest variance and exhibited the same band counts as those vertebrae in other locations for both sexes, thus these vertebrae of each specimen were used for age analysis in this study.
$\overline X $
is the mean diameter of three consecutive vertebral centra. The analysis revealed that the 19th to the 21th vertebrae (beginning of the abdominal cavity) had the smallest variance and exhibited the same band counts as those vertebrae in other locations for both sexes, thus these vertebrae of each specimen were used for age analysis in this study.
Cleaned vertebral centra were epoxy-impregnated for 12 h and then sectioned along the longitudinal plane to 0.5 mm thicknesses with a low speed saw (Buehler, Düsseldorf, Germany) (Davis et al., Reference Davis, Cailliet and Ebert2007), and polished to 0.1–0.3 mm with 2000-grain sandpaper (Joung et al., Reference Joung, Lee, Liu and Liao2011). Several staining methods were tried but could not enhance the band pairs in the sectioned vertebrae. Hence, vertebral sections were rinsed in 70% alcohol, and then mounted on slides with cytoseal. These vertebral centra were examined using a microscope (Olympus CX21, Tokyo, Japan) with reflected light and images were captured by an attached digital camera (Nikon E5000, Tokyo, Japan). These images were processed by the image process system (D3C) (Joung et al., Reference Joung, Chen, Lee and Liu2008, Reference Joung, Lee, Liu and Liao2011).
Growth band pairs (comprised of one opaque and one translucent band per pair, interpreted under conditions of reflected light) (Figure 2) were counted without prior knowledge of the sex or length of the specimens. Counts were accepted only if both counts by two readers were in agreement. If the estimated numbers of bands differed, the centrum was recounted by the first reader and the final count was accepted as the agreed number. If the third count did not match one of the previous two counts, the sample was discarded (Joung et al., Reference Joung, Liao, Liu, Chen and Leu2005, Reference Joung, Chen, Lee and Liu2008, Reference Joung, Lee, Liu and Liao2011).

Fig. 2. Growth band pairs formed on the sectioned vertebral centrum of a male Dipturus kwangtungensis with 43.2 cm TL. The dots indicate opaque bands.
As the ovulation season of D. kwangtungensis could not be clearly defined (Hou, Reference Hou2002), the birth date was assumed to be 1 January based on the gestation period and the time of mating (Hou, Reference Hou2002). The age at the first opaque band formation (1 June) was assumed to be 0.5 year. The age of every skate was estimated based on the number of band pairs being counted plus the time lag between sampling date and band formation accordingly. The index of the average percentage error (IAPE) (Beamish & Fournier, Reference Beamish and Fournier1981) and the coefficient of variation (CV) (Chang, Reference Chang1982) were used to compare reproducibility of the age determination between two readings. The relationship between TL and D can be expressed as TL = a − Db , where a, b are parameters. The analysis of residual sum of squares (ARSS, Chen et al., Reference Chen, Jackson and Harvey1992) was used to compare the TL-D relationships between sexes.
The marginal increment ratio (MIR) is commonly used in determining the periodicity of band pair formation. However, high uncertainty occurs on the measurement of the radii of the ultimate and penultimate bands for specimens with old ages. To reduce this uncertainty, the centrum edge analysis (Cailliet & Goldman, Reference Cailliet, Goldman, Carrier, Musick and Heithaus2004) was used to determine the time of band formation using the monthly frequency changes in band edge type. The periodicity obtained from centrum edge analysis was further verified by a statistical model (Okamura & Semba, Reference Okamura and Semba2009), which combined a statistical model for binary data with a statistical model for circular data. The most possible periodicity of band pair formation from annual, biannual, and no cycle was selected based on an Akaike's information criterion corrected (AICc, Akaike, Reference Akaike, Petrov and Csaki1973).
Growth functions
Four commonly used growth functions for fish were used to model the observed length at age data of D. kwangtungensis and were described as follows:
-
1. von Bertalanffy growth function (VBGF, von Bertalanffy, Reference von Bertalanffy1938):
where L t is the length at age t, L ∞ is the asymptotic length, k is the growth coefficient of von Bertalanffy function, t is the age (year from birth), and t 0 is the theoretical age at length 0. $${L_t}\, = \,{L_\infty} (1 - {e^{ - k(t - {t_0})}});$$
$${L_t}\, = \,{L_\infty} (1 - {e^{ - k(t - {t_0})}});$$
-
2. Two-parameter VBGF (Fabens, Reference Fabens1965)
where L 0 is the length at birth and is set as 5.9 cm based on the smallest free-swimming individual observed in this study.
$${L_t} = {L_\infty} - ({L_\infty} - {L_0}){e^{ - kt}}$$
-
3. Robertson (Logistic) growth function (Robertson, Reference Robertson1923):
where k R and t 0 are the growth coefficient and constant of Robertson function, respectively.
$${L_t} = \displaystyle{{{L_\infty}} \over {1 + {e^{ - {k_R}(t - {t_0})}}}}.$$
-
4. Gompertz growth function (Gompertz, Reference Gompertz1825):
where k G is the growth coefficient of Gompertz function, and t 0 is the constant to be estimated.
$${L_t} = {L_\infty} {e^{ - {e^{ - {k_G}(t - {t_0})}}}}.$$




These four functions were fitted using the Gauss–Newton algorithm in the NLIN procedure of the statistical package SAS ver. 9.0 (SAS Institute 2001, Cary, NC, USA). The goodness of fit of the four growth functions was compared based on corrected Akaike's Information Criterion (AICc). AICc is expressed as:
 $$AI{C_c}\, = \,AIC + \displaystyle{{2K(K + 1)} \over {N - K - 1}},$$
$$AI{C_c}\, = \,AIC + \displaystyle{{2K(K + 1)} \over {N - K - 1}},$$
AIC = N × ln(MSE) + 2K (Akaike, Reference Akaike, Petrov and Csaki1973), where N is the total sample size, MSE is the mean square of residuals, K is the number of parameters estimated in the growth function. The ARSS (Chen et al., Reference Chen, Jackson and Harvey1992) was used to compare the growth curves between sexes.
RESULTS
Body metrics and sex ratio
A total of 674 females ranging from 5.9 to 73.2 cm TL and 583 males ranging from 6.6 to 66.0 cm TL were collected in this study (Table 1). The sex ratio (the ratio of males to females), 0.86, of all specimens significantly differed from 1.0 (χ2 = 4.91, P < 0.05). Significant differences in sex ratio were also found in June, July and December, 2007 (Table 1). The vertebrae of 652 specimens arbitrarily chosen from each 5 cm length interval based on its proportion of the sample size were used for age determination. Consequently, 426 vertebral centra were successfully processed for age determination by vertebral band pair counts.
A GLM indicated that the interaction term of sex and TL is significant (F = 27.7, P < 0.001) suggesting that DW–TL relationships significantly differed between females and males and sex-specific equations were estimated as follows:
Female: DW = −0.538 + 0.7141 TL (N = 710, r 2 = 0.98, P < 0.05),
Male: DW = 0.056 + 0.6925 TL (N = 656, r 2 = 0.96, P < 0.05).
The ARSS indicated that there is no significant difference on the TL–D relationship between sexes (F = 3.01, P > 0.05) and the sexes-combined relationship was estimated as follows: TL = 14.11D 0.888 (N = 411, r2 = 0. 94, P < 0.001).
Age estimation
Vertebral band pairs were counted up to 14 for both sexes based on 364 (193 females and 171 males) of the 426 vertebral centra examined. In total, 62 vertebral centra (14.5%) were rejected because the third band counts differed from the previous two counts. The average IAPE was 1.83% (CV of 2.73%) for the overall sample (N = 426). Per cent agreement between two readings was 83% total agreement, 93.4% for one-band difference, and 98.8% for two-band difference.
Monthly changes in the frequency of opaque bands on the vertebral edge for both sexes combined appeared to peak in June, decreased gradually and to the lowest value in January (Figure 3). This trend indicates that the vertebral band pair is formed once a year. Okamura & Semba's (Reference Okamura and Semba2009) statistical analysis also indicated that one band pair per year had the smallest AICc and the predicted probability fit the observations well (Figure 3, Table 2).

Fig. 3. Monthly changes in the edge analysis of Dipturus kwangtungensis. Numbers indicate the sample sizes, line indicates the predicted probability of annual cycle from Okamura and Semba's model (Reference Okamura and Semba2009).
Table 2. Comparison of goodness-of-fit among different periodicities of vertebral band pair formation for Dipturus kwangtungensis.

AICc, corrected Akaike's Information Criterion.
Growth functions
The iterations of parameter estimation for VBGF did not converge and the estimated value of L∞ (503.7 cm TL) was much larger than the maximum observed lengths (73.2 cm TL) indicating that VBGF is not the best model for D. kwangtungensis. The Gompertz growth function had the smallest AICc among the four growth functions for sexes-combined length at age data (Table 3). Hence, the Gompertz function was chosen to be the best growth function for Kwangtung skates. The ARSS indicated that no significant difference on sex-specific growth equations and the sexes-combined Gompertz growth equation was estimated as follows:
 ${L_t} = 96.7{e^{ - {e^{ - 0.114(t - 5.45)}}}}$
(N = 364, P < 0.01) (Figure 4). The maximum observed size of the specimens, which was not included in the growth model building, was 73.2 cm TL corresponding to 16.7 years old.
${L_t} = 96.7{e^{ - {e^{ - 0.114(t - 5.45)}}}}$
(N = 364, P < 0.01) (Figure 4). The maximum observed size of the specimens, which was not included in the growth model building, was 73.2 cm TL corresponding to 16.7 years old.

Fig. 4. Sexes-combined VBGF, Robertson and Gompertz growth curves of Dipturus kwangtungensis in this study. ○: females, •: males.
Table 3. Estimates of growth parameters, their standard deviations (in parentheses) and goodness of fit for three growth functions fitted to observed size-at-age data for Dipturus kwangtungensis in northern waters of Taiwan.

VBGF cannot converge.
Growth coefficient: k for VBGF, k R for Robertson function, and kG for Gompertz function.
DISCUSSION
Various definitions of the birth mark of skates and rays have been made by different authors. Davis et al. (Reference Davis, Cailliet and Ebert2007) defined the birthmark as a change in angle of the corpus calcareum, and each band pair after that was considered a year of growth for B. trachura. Sulikowski et al. (Reference Sulikowski, Irvine, DeValerio and Carlson2007) described the birth mark as the first distinct mark distal to the focus that coincided with a change in the angle of the corpus calcareum for roundel skate. On the other hand, McFarlane & King (Reference McFarlane and King2006) described the first visible dark band as the birth band for Raja binoculata and R. rhina. A similar assumption was also found for Okameiei acutispina (Joung et al., Reference Joung, Lee, Liu and Liao2011). We substituted the average diameter of the first band into the TL-D relationship and obtained an estimation of the TL at the first band to be 10.7 cm which is much larger than the smallest free swimming found in this study (5.9 cm TL). Therefore, we concluded that there is no birth mark on the vertebral centrum for this species.
There has been an increase in the use of both verification and validation methodologies in chondrichthyan growth studies, such as marginal increment analysis, centrum edge analysis, size mode analysis, tag-recapture analysis, captive growth analysis, tetracycline (OTC) marking and radiocarbon analysis (Cailliet et al., Reference Cailliet, Smith, Mollet and Goldman2006). It is likely that a combination of verification and validation approaches will produce the most convincing results. For example, Natanson et al. (Reference Natanson, Mello and Campana2002) used ageing structure OTC marking, length-frequency analysis, and tag-recapture analysis to verify the ageing of the porbeagle, Lamna nasus. Cailliet et al. (Reference Cailliet, Smith, Mollet and Goldman2006) mentioned that when combined with additional techniques, such as marginal increment analysis, centrum edge analysis can provide valuable corroborative evidence to verify the periodicity of band formation. Because errors were commonly found in the measurements of radii of the ultimate and penultimate bands, instead of combining with MIA, we only used centrum edge analysis to verify the periodicity of band pair formation in this study. Although the month of the peak in centrum edge analysis has the lowest sample size, there is evidence that one band pair is deposited per year. The result was further supported by using a statistical analysis proposed by Okamura & Semba (Reference Okamura and Semba2009) indicating that one band pair was deposited per year although the no cycle model had slight support with the second highest value of AICc weight (Table 2). Most work on age and growth verification in skates indicated one growth band pair formed per year in the vertebral centra. Examples include Amblyraja radiata (Sulikowski et al., Reference Sulikowski, Kneebone and Elzey2005), Raja texana (Sulikowski et al., Reference Sulikowski, Irvine, DeValerio and Carlson2007) and Okamejei acutispina (Joung et al., Reference Joung, Lee, Liu and Liao2011). These findings further support that one band pair deposition per year is the most likely model for D. kwangtungensis in the present study.
Index of average percentage error (IAPE; Beamish & Fournier, Reference Beamish and Fournier1981) and coefficient of variation (CV; Chang, Reference Chang1982) are the two indices most commonly used in evaluating the reliability of ageing (Cailliet et al., Reference Cailliet, Smith, Mollet and Goldman2006). Campana (Reference Campana2001) suggested that a CV less than 5% is a reasonable estimate. The small value of IAPE (1.83%) and CV of 2.73% in the present study suggests that the age determination is reliable.
Two versions of the von Bertalanffy growth function (VBGF), conventional VBGF (von Bertalanffy, Reference von Bertalanffy1938), and two-parameter VBGF (Fabens, Reference Fabens1965), along with the Robertson (Robertson, Reference Robertson1923), and the Gompertz growth functions (Gompertz, Reference Gompertz1825) have all been used to describe the age and growth of skates (Neer & Thompson, Reference Neer and Thompson2005; Serra-Pereira et al., Reference Serra-Pereira, Figueiredo, Bordalo-Machado, Farias, Moura and Gordo2005; McFarlane & King, Reference McFarlane and King2006; Natanson et al., Reference Natanson, Sulikowski, Kneebone and Tsang2007). Though commonly used to model growth in elasmobranchs, researchers who have examined other models have found that VBGF often does not adequately estimate growth parameters for skate species (Neer & Cailliet, Reference Neer and Cailliet2001; McFarlane & King, Reference McFarlane and King2006; Joung et al., Reference Joung, Lee, Liu and Liao2011). Unlike the VBGF, the Robertson and Gompertz growth functions assume the maximum growth rate occurs at an intermediate age. They have proved to be the best model describing the growth of skates and rays in some cases. McFarlane & King (Reference McFarlane and King2006) and Joung et al. (Reference Joung, Lee, Liu and Liao2011) concluded that the Robertson growth function best described the growth of the big skate, Raja binoculata, and the sharpspine skate, Okamejei acutispina. While Neer & Cailliet (Reference Neer and Cailliet2001) and Mollet et al. (Reference Mollet, Ezcurra and O'Sullivan2002) found the Gompertz growth function best fit the age-length data of the Pacific electric ray, Torpedo californica, and the pelagic stingray, Dasyatis violacea.
In this study, L ∞ value estimated from the VBGF was much larger than the maximum observed total length because the parameter estimate could not converge in iterations. The biologically unrealistic results implied that VBGF is not suitable to describe the growth of D. kwangtungensis. On the other hand, L ∞ values estimated with the Gompertz growth function were about 1.32 folds of the maximum observed total length, and were believed to be more biologically realistic. Ishihara (Reference Ishihara1987) documented a maximum male of 75.7 cm TL in Japanese waters. Although this individual is much smaller than our estimate of L ∞ (96.7 cm TL), the Gompertz growth function still appears to be the best model to describe the growth of D. kwangtungensis according to the AICc criteria. The growth coefficients (kG) derived from this study were comparable to those of Rhinoptera bonasus (Neer & Thompson, Reference Neer and Thompson2005) (Table 4) which has larger asymptotic length than D. kwangtungensis.
Table 4. Comparison of growth parameters of Gompertz function for skates and rays from different studies.

Cailliet & Goldman (Reference Cailliet, Goldman, Carrier, Musick and Heithaus2004) reported that growth model estimates are greatly affected by the lack of very young or old individuals. In this study, the specimens for vertebral age analysis ranged from 5.9 to 73.2 cm TL, which may cover most of the size range of this species (Ishihara, Reference Ishihara1987). However, only few large specimens (>60 cm TL) were collected during our sampling period because some large individuals were processed at sea. The lack of large specimens may result in an overestimate of L ∞ in this study. To improve the accuracy of parameter estimation, future work should focus on the collection of large specimens.
Some elasmobranchs have size-segregation behaviour, e.g. cownose ray, Rhinoptera bonasus (Smith & Merriner, Reference Smith and Merriner1987), silky shark, Carcharhinus falciformis (Oshitani et al., Reference Oshitani, Nakano and Tanaka2003) and blue shark, Prionace glauca (Nakano & Stevens, Reference Nakano, Stevens, Camhi, Pikitch and Babcock2008). Dipturus kwangtungensis might also have such behaviour. Thus, the few large specimens collected in this study may be because the fishing ground does not cover the major habitat for these skates. McFarlane & King (Reference McFarlane and King2006) mentioned that the very large and older specimens may be difficult to sample in a heavily exploited population. A similar situation may occur in this study. The proportion of large individuals decreased during the period from 2000–2001 (Hou, Reference Hou2002) to 2006–2008 (present study), suggesting it is possible that the number of large and older individuals decreased with the increase of exploitation. However, more information on population assessment is needed to test this hypothesis.
In conclusion, this study provides the first detailed estimates of the age and growth parameters for D. kwangtungensis. This species has a life history characteristic of moderate growth coefficient (0.114 year−1); although current exploitation is unclear, to ensure the sustainable utilization of this stock, close monitoring of its catch and size composition, and periodic assessment of abundance are recommended.
FINANCIAL SUPPORT
This study was financially supported by the National Science Council, Republic of China, through grants NSC98-2313-B-019-032 and NSC99-2313-B-019-003-MY3.








