



Introduction
The events at the Permian-Triassic boundary profoundly changed the evolutionary history of Earth's biota (Sepkoski, Reference Sepkoski1981; Erwin, Reference Erwin1994). Foraminifera were among the major faunal groups severely affected by these events, and several distinct Permian foraminiferal assemblages became extinct at the end of the Permian Period. Following this end-Permian mass extinction, the largest biotic and ecological crisis of the Phanerozoic (Raup and Sepkoski, Reference Raup and Sepkoski1982; Bambach, Reference Bambach2006; Payne and Clapham, Reference Payne and Clapham2012), recovery of foraminifera and other benthic marine organisms was delayed and interrupted by ongoing adverse environmental conditions as well as several distinct environmental crises (Payne et al., Reference Payne, Lehrmann, Wei, Orchard, Schrag and Knoll2004; Lau et al., Reference Lau, Maher, Altıner, Kelley, Kump, Lehrmann, Silva-Tamayo, Weaver, Yu and Payne2016; Zhang et al., Reference Zhang, Romaniello, Algeo, Lau, Clapham, Richoz, Hermann, Smith, Horacek and Anbar2018). Due to the complexity of environmental change and biological response during the extended recovery interval, which lasted into the Middle Triassic (Hallam, Reference Hallam1991; Payne et al., Reference Payne, Summers, Rego, Altıner, Wei, Yu and Lehrmann2011; Chen and Benton, Reference Chen and Benton2012), detailed taxonomic study and fine-scale time resolution are required to establish the pattern of recovery and its relationship to environmental change.
Foraminifera are well suited to the study of recovery dynamics due to their diversity and abundance in Lower and Middle Triassic carbonates. The Triassic foraminiferal fauna of the Great Bank of Guizhou, an isolated carbonate platform of latest Permian to Late Triassic age within the Nanpanjiang Basin of south China, are particularly advantageous for this purpose. Foraminifera are abundant in many samples, distributed throughout stratigraphic sections exceeding 2000 m in thickness, and preserved in depositional environments from platform interior to basin margin (Payne et al., Reference Payne, Lehrmann, Wei and Knoll2006, Reference Payne, Summers, Rego, Altıner, Wei, Yu and Lehrmann2011; Song et al., Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan, Niu, Chen, Yang and Wang2011a).
The Great Bank of Guizhou, studied in five different measured sections, namely Dawen (PDW), Dajiang (PDJ), Middle Triassic Dajiang (MDJ), Guandao (PDG), and Upper Guandao (PUG), contains well-diversified Early to early Late Triassic benthic foraminifera assemblages. Under the Phylum Foraminifera, the studied foraminifera assemblages belong to classes Miliolata, Textulariata, Fusulinata, Nodosariata, and to an uncertain class housing the involutinid- and robertinid-type foraminifera.
During the second half of the last century, several authors described and documented Early to Middle Triassic foraminiferal taxa (Ho, Reference Ho1959; Luperto, Reference Luperto1965; Pantić, Reference Pantić1965; Kochansky-Devidé and Pantić, Reference Kochansky-Devidé and Pantić1966; Koehn-Zaninetti, Reference Koehn-Zaninetti1969; Baud et al., Reference Baud, Zaninetti and Brönnimann1971, Reference Baud, Brönnimann and Zaninetti1974; Premoli Silva, Reference Premoli Silva1971; Brönnimann and Zaninetti, Reference Brönnimann and Zaninetti1972; Brönnimann et al., Reference Brönnimann, Zaninetti and Bozorgnia1972a, Reference Brönnimann, Zaninetti, Bozorgnia and Huberb, Reference Brönnimann, Cadet and Zaninetti1973a, Reference Brönnimann, Cadet and Zaninettib, Reference Brönnimann, Zaninetti, Moshtaghian and Huberc, Reference Brönnimann, Zaninetti, Moshtaghian and Huber1974; Zaninetti et al., Reference Zaninetti, Brönnimann, Bozorgnia and Huber1972a, Reference Zaninetti, Brönnimann and Baudb, Reference Zaninetti, Brönnimann and Baudc, Reference Zaninetti, Brönnimann, Huber and Moshtaghian1978, Reference Zaninetti, Rettori and Martini1994; Efimova, Reference Efimova1974; Gazdzicki et al., Reference Gazdzicki, Trammer and Zwidzka1975; Stampfli et al., Reference Stampfli, Zaninetti, Brönnimann, Jenny-Deshusses and Stampfli-Vuille1976; Zaninetti, Reference Zaninetti1976; Gazdzicki and Smit, Reference Gazdzicki and Smit1977; Dağer, Reference Dağer1978a; Trifonova, Reference Trifonova1978a, Reference Trifonovab, Reference Trifonovac, Reference Trifonova1992, Reference Trifonova1993, Reference Trifonova1994; Čatalov and Trifonova, Reference Čatalov and Trifonova1979; Altıner and Zaninetti, Reference Altıner and Zaninetti1981; Salaj et al., Reference Salaj, Borza and Samuel1983; He, Reference He1984, Reference He1988, Reference He1993; Orovecz-Scheffer, Reference Orovecz-Scheffer1987; Benjamini, Reference Benjamini1988; He and Wang, Reference He and Wang1990; He and Cai, Reference He and Cai1991; Altıner and Koçyiğit, Reference Altıner and Koçyiğit1993; Rettori et al., Reference Rettori, Angiolini and Muttoni1994; Rettori, Reference Rettori1995). Some selected genera directly from the late Paleozoic in the definition of their new taxa, such as Rectocornuspira (now Postcladella), Cornuspira (previously described as Cyclogyra), Permodiscus, Endothyra, and Earlandia, therefore studies relating Early Triassic foraminiferal taxa to the Permian-Triassic mass extinction event were carried out only rarely prior to the end of the last century (Nakazawa et al., Reference Nakazawa, Kapoor, Ishii, Bando, Okimura and Tokuoka1975; Altıner et al., Reference Altıner, Baud, Guex and Stampfli1980; Taraz et al., Reference Taraz, Golshani, Nakazawa, Shimizu, Bando, Ishii, Murata, Okimura, Sakagami, Nakamura and Tokuoka1981; Sheng et al., Reference Sheng, Chen, Wang, Rui, Liao, Bando, Ishii, Nakazawa and Nakamura1984; Neri and Pasini, Reference Neri and Pasini1985; Pasini, Reference Pasini1985; Broglio Loriga et al., Reference Broglio Loriga, Neri, Pasini and Posenato1988; Cirilli et al., Reference Cirilli, Pirini Radrizzani, Ponton and Radrizzani1998).
As the importance of the Permian-Triassic boundary mass extinction within the evolution of Phanerozoic marine ecosystems became clear (Sepkoski, Reference Sepkoski1981; Raup and Sepkoski, Reference Raup and Sepkoski1982; Wignall and Hallam, Reference Wignall and Hallam1992; Erwin, Reference Erwin1993), research into the causes and mechanisms of the mass extinction and controls on subsequent biotic recovery accelerated (Erwin, Reference Erwin1994, Reference Erwin2007; Rampino and Adler, Reference Rampino and Adler1998; Jin et al., Reference Jin, Wang, Wang, Shang, Cao and Erwin2000; Tong and Shi, Reference Tong and Shi2000; Leven and Korchagin, Reference Leven and Korchagin2001; Wignall and Newton, Reference Wignall and Newton2003; Altıner et al., Reference Altıner, Groves and Özkan-Altıner2005; Groves and Altıner, Reference Groves and Altıner2005; Kaiho et al., Reference Kaiho, Kajiwara, Chen and Gorjan2006; Song et al., Reference Song, Tong, Zhang, Wang and Chen2007, Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan, Niu, Chen, Yang and Wang2011a, Reference Song, Wignall, Tong and Yin2013; Yin et al., Reference Yin, Feng, Lai, Baud and Tong2007; Algeo et al., Reference Algeo, Chen, Fraiser and Twitchet2011; Payne et al., Reference Payne, Summers, Rego, Altıner, Wei, Yu and Lehrmann2011; Rego et al., Reference Rego, Wang, Altıner and Payne2012). As part of this previous research, the evolution of foraminifera in the Lower to Middle Triassic stratigraphy has been studied to understand extinction and recovery dynamics. Foraminiferal paleontologists and stratigraphers started to trace and document foraminifera along measured sections encompassing the Permian-Triassic boundary and to include data from other disciplines in order to improve their foraminifera-based chronology (Broglio Loriga and Cassinis, Reference Broglio Loriga, Cassinis, Sweet, Zunyi, Dickins and Hongfu2003; Ünal et al., Reference Ünal, Altıner, Yilmaz and Özkan-Altıner2003; Altıner et al., Reference Altıner, Groves and Özkan-Altıner2005; Groves et al., Reference Groves, Altıner and Rettori2005, Reference Groves, Rettori, Payne, Boyce and Altıner2007; Mohtat-Aghai and Vachard, Reference Mohtat-Aghai and Vachard2005; Angiolini et al., Reference Angiolini, Carabelli, Nicora, Crasquin-Soleau, Marcoux and Rettori2007; Théry et al., Reference Théry, Vachard, Dansart, Álvaro, Aretz, Boulvain, Munnecke, Vachard and Vennin2007; Vuks, Reference Vuks2007; Galfetti et al., Reference Galfetti, Bucher, Martini, Hochuli, Weissert, Crasquin-Soleau, Brayard, Goudmand, Brühwiler and Goudun2008; Maurer et al., Reference Maurer, Rettori and Martini2008; Korchagin, Reference Korchagin2011; Krainer and Vachard, Reference Krainer and Vachard2011; Nestell et al., Reference Nestell, Kolar-Jurkovšek, Jurkovšek and Aljinović2011; Song et al., Reference Song, Tong and Chen2011b, Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015, Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016, Reference Song, Wignall and Dunhill2018; Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015).
The purpose of this study is to give a comprehensive taxonomic account of the Early to Middle Triassic foraminifera from the Great Bank of Guizhou, including descriptions of one new genus and three new species recovered. In addition, the occurrence patterns of the taxa along measured stratigraphic sections are used to develop a foraminiferal biostratigraphic framework for the Lower Triassic through Anisian interval. This biostratigraphic study excludes the Ladinian and Carnian stages of the measured sections because the foraminiferal data recovered from these stages are more fragmentary due to a combination of unfavorable facies and poor preservation in collected samples. The occurrence patterns are further used to assess the recovery of foraminifera following the end-Permian mass extinction within an Early to Middle Triassic timescale calibrated by conodonts.
Geological setting and studied stratigraphic sections
The foraminiferal study is based on samples from five stratigraphic sections measured on outcrops of the Great Bank of Guizhou (GBG), an isolated carbonate platform of latest Permian to earliest Late Triassic age located in the Nanpanjiang Basin of the Yangtze Block, southern China (Fig. 1). The Nanpanjiang Basin was a deep-marine embayment in the southern margin of the south China tectonic block and is bordered by the Yangtze Platform, a vast shallow-marine carbonate platform that stretched across south China. The GBG initiated in the latest Permian during a relative rise in sea level that drowned much of the Yangtze Platform (Fig. 1.1; Lehrmann, Reference Lehrmann1993; Lehrmann et al., Reference Lehrmann, Wei and Enos1998, Reference Lehrmann, Payne, Enos, Montgomery, Wei, Yu, Xiao and Orchard2005, Reference Lehrmann, Donghong, Enos, Minzoni, Ellwood, Orchard, Zhang, Wei, Dillett, Koenig, Steffen, Druke, Druke, Kessel and Newkirk2007, Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015; Payne et al., Reference Payne, Lehrmann, Wei, Orchard, Schrag and Knoll2004, Reference Payne, Lehrmann, Wei and Knoll2006). Uppermost Permian skeletal carbonates of the Wujiaping Formation underlie the platform interior, whereas fine-grained siliciclastics of the uppermost Permian Dalong Formation underlie the Permian/Triassic transition beneath Lower Triassic slope facies. During the Early Triassic, the depositional setting of the GBG was a low-relief bank with oolite shoals developed at the margin, a shallow subtidal to peritidal interior, and slopes dominated by hemipelagic carbonate mud intercalated with thin carbonate turbidites and debris flows shed from the margin (Lehrmann et al., Reference Lehrmann, Wei and Enos1998, Reference Lehrmann, Payne, Enos, Montgomery, Wei, Yu, Xiao and Orchard2005). Recently, reconstruction of the GBG in the latest Permian to Early Triassic interval has been partly revised by Kelley et al. (Reference Kelley, Lehrmann, Yu, Jost, Meyer, Lau, Altıner, Li, Minzoni, Schaal and Payne2020), who defined three stages of development. These authors considered the latest Permian–Smithian interval as the initiation and low-relief bank stages, and the Smithian to late Spathian interval as the aggrading and steepening stage. The Lower Triassic platform interior consists of shallow-marine carbonates of the Daye Formation overlain by dolomitized shallow-marine carbonates of the Anshun Formation. The Lower Triassic shales, micritic carbonates, carbonate turbidites, and allodapic breccias of the slope belong to the Luolou Formation. In the Middle Triassic, Anisian Tubiphytes reefs rimmed the outer margin and slope of the GBG while peritidal conditions continued in the interior (Fig. 1.2). The platform subsequently developed a high-relief escarpment during Ladinian. The escarpment profile continued until the early Carnian, when the GBG drowned due to accelerated tectonic subsidence and was subsequently buried by siliciclastic turbidites (Lehrmann et al., Reference Lehrmann, Wei and Enos1998, Reference Lehrmann, Payne, Enos, Montgomery, Wei, Yu, Xiao and Orchard2005). The Middle Triassic shallow-marine carbonates of the platform interior belong to the Yangliujing Formation, and the Middle Triassic slope carbonates belong to the Xinyang Formation. The Upper Triassic siliciclastic turbidites of the Bianyang Formation filled the remaining accommodation in the basin and, ultimately, buried the platform top.

Figure 1. (1) Early Triassic paleogeographic map (Lehrmann et al., Reference Lehrmann, Wei and Enos1998; Payne et al., Reference Payne, Lehrmann, Wei, Orchard, Schrag and Knoll2004). Hash pattern indicates the Nanpanjiang Basin and brick pattern indicates the Yangtze Platform and GBG. (2) Schematic cross section of the Great Bank of the Guizhou, illustrating the locations of Dawen, Dajiang, Middle Triassic Dajiang, Guandao, and Upper Guandao sections within the platform architecture.
The studied stratigraphic sections span a range of depositional environments. Three of the studied sections—the Dawen, Dajiang, and Middle Triassic Dajiang sections—are located in the platform interior (Fig. 1.2). Both the Dawen and Dajiang sections cover the stratigraphic interval from Griesbachian to Smithian. The Middle Triassic Dajiang section consists of four short sections (~20 m) measured at intervals through a part of the thick carbonate platform spanning from the Pelsonian (mid Anisian) to Ladinian. The Guandao and Upper Guandao sections, representing the slope and basin-margin environments, cover the entire Griesbachian–lower Carnian interval (Fig. 2).

Figure 2. Stratigraphic columns of measured sections. Timescale is constrained by conodont occurrence data and physical stratigraphic and carbon isotope correlations between the basin margin and the platform interior (Payne et al., Reference Payne, Lehrmann, Wei, Orchard, Schrag and Knoll2004; Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015; Kelley et al., Reference Kelley, Lehrmann, Yu, Jost, Meyer, Lau, Altıner, Li, Minzoni, Schaal and Payne2020). 1–12 = Biozones of foraminifera (for biozone names see Fig. 21). Gray vertical bar indicates intervals devoid of foraminifera.
Materials
From the stratigraphic sections measured in the Great Bank of Guizhou, we collected a total of 1106 samples for lithostratigraphy, facies analysis, isotope stratigraphy, conodont and foraminiferal taxonomy, and biostratigraphy. From the samples displaying fossil fragments and suitable carbonate facies, 598 thin sections were prepared for the foraminifera, 246 of which contained at least one foraminifer. The total number of foraminifera specimens is >2500, of which 351 were illustrated in the figures of this paper.
Foraminifera recovered from the samples were first used in a genus-level study by Payne et al. (Reference Payne, Summers, Rego, Altıner, Wei, Yu and Lehrmann2011). These authors carried out a study on the Early–Middle Triassic trends in diversity, evenness, and size of the foraminifers to investigate the tempo and mode of biotic recovery. The current study concentrates, with a different scope and resolution, on species-level taxonomic description with many illustrations, construction of a new biostratigraphic scheme calibrated by conodonts, and a discussion of species-level recovery of Early–Middle Triassic foraminifera, with attention to phyletic relationships.
Repositories and institutional abbreviations
Types, figures, and other specimens examined in this study are deposited in the thin section laboratory of the Department of Geological Sciences, Stanford University, with the following catalog abbreviations: Dawen (PDW), Dajiang (PDJ), Middle Triassic Dajiang (MDJ), Guandao (PGD) and upper Guandao (PUG) sections.
Systematic paleontology
Demir Altıner and Jonathan L. Payne
As indicated by Vachard (Reference Vachard2016, Reference Vachard, Lucas and Shen2018) the current classification of foraminifera based on wall microstructure disagrees with the results of molecular phylogenetic studies of extant species (e.g., Pawlowski et al., Reference Pawlowski, Holzmann and Tyszka2013). However, molecular results remain difficult to reconcile with the Paleozoic–Mesozoic fossil record. In this study, the Triassic foraminifera of the Great Bank of Guizhou have been classified at first in distinct populations at species rank, and then placed into reasonable categories at the genus level based on wall structure and composition and other morphological characters. Despite uncertainties in higher taxonomic ranks, we consider most of the families used in the Triassic foraminiferal paleontology to be adequate for recognized genera and species. However, intermediate taxonomic ranks, including superfamily, suborder, and order, are less stable in their use across studies and have not been used in this study. For the largest groups of foraminifera, classified based on wall microstructure and assigned class rank, we have largely followed Vachard (Reference Vachard2016, Reference Vachard, Lucas and Shen2018) and Cavalier-Smith (Reference Cavalier-Smith2002, Reference Cavalier-Smith2003), the latter of whom demonstrated that foraminifera constitute a phylum or subphylum.
Class Miliolata Saidova, Reference Saidova1981
Remarks
One of the dominant groups of Early–Middle Triassic foraminifera on the Great Bank of Guizhou is the bilocular Miliolata, consisting of Cornuspiridae, Arenovidalinidae, Meandrospiridae, Hoyenellidae, and Agathamminidae and the primitive multilocular miliolid families Ophthalmiidae, Quinquelloculinidae, and Galeanellidae.
Family Cornuspiridae Schultze, Reference Schultze1854
Subfamily Cornuspirinae Schultze, Reference Schultze1854
Genus Postcladella Krainer and Vachard, Reference Krainer and Vachard2011
Postcladella kalhori (Brönnimann, Zaninetti, and Bozorgnia, Reference Brönnimann, Zaninetti and Bozorgnia1972a)
Figure 3.1–3.26

Figure 3. All specimens are from Dawen (PDW) and Dajiang (PDJ) sections. (1–26) Postcladella kalhori (Brönnimann, Zaninetti, and Bozorgnia, Reference Brönnimann, Zaninetti and Bozorgnia1972a); (27–38) Postcladella grandis (Altıner and Zaninetti, Reference Altıner and Zaninetti1981). (1, 2, 22) PDW-011; (3) PDJ-164; (4, 7, 18–21, 25, 27, 30–32, 34) PDJ-174; (5) PDJ-178; (6, 17, 23, 24, 29) PDW-108; (8, 9) PDJ-157; (10, 11) PDW-099; (12, 28, 33, 36) PDW-118; (13) PDJ-162; (14) PDJ-200; (15) PDW-087; (16) PDJ-161; (26) PDW-123; (35) PDJ-170; (37) PDJ-158; (38) PDW-126. Scale bars = 100 μm.
Remarks
There is currently disagreement regarding the taxonomy of Early Triassic cornuspirin taxa, which is one of the most common foraminiferal groups in basal Triassic strata. Two of the frequently cited taxa, Rectocornuspira kalhori and Cyclogyra? mahajeri, were described by Brönnimann et al. (Reference Brönnimann, Zaninetti and Bozorgnia1972a). The primary difference between these two taxa, commonly encountered in axial sections, is that the lumen of the final whorl of C.? mahajeri overlaps laterally onto the previous coil. Brönnimann et al. (Reference Brönnimann, Zaninetti and Bozorgnia1972a) added that this character distinguishes this species from the axially slender and strongly biumbilicate planispiral stage of R. kalhori. This approach was altered by Gaillot and Vachard (Reference Gaillot and Vachard2007, p. 84), who considered Early Triassic Rectocornuspira as an uncoiled Cornuspira and stated that ‘Rectocornuspira would correspond to Cornuspira with a morphological adaptation, more or less developed, depending on the local or regional ecological parameters.’ This interpretation did not take into account the morphological difference between the kalhori and mahajeri populations.
Although the reasons given by Krainer and Vachard (Reference Krainer and Vachard2011) were correct for the creation of their new genus Postcladella for the kalhori population, these authors did not consider the morphological difference between kalhori and mahajeri populations as a valid criterion for distinguishing between the species, and therefore synonymized these two taxa under Postcladella. More recently, Nestell et al. (Reference Nestell, Kolar-Jurkovšek, Jurkovšek and Aljinović2011), describing the foraminifera from the Permian–Triassic transition in Slovenia, rejected Postcladella of Krainer and Vachard (Reference Krainer and Vachard2011), illustrated all P. kalhori populations as ‘Cornuspira’ mahajeri, and considered Postcladella (given as ‘Rectocornuspira’) to be a teratologic form of Cornuspira, following Gaillot and Vachard (Reference Gaillot and Vachard2007). In Nestell et al. (Reference Nestell, Nestell, Ellwood, Wardlaw, Basu, Ghosh, Lan, Rowe, Hunt, Tomkin and Ratcliffe2015), the kalhori population took priority over the mahajeri population, and was illustrated and described under the agglutinated genus Ammodiscus. The basis of this taxonomic reassignment was explained as the absence of calcium carbonate and the presence of a significant amount of elemental carbon, along with oxygen and silica in the walls of kalhori specimens. In our view, however, these authors did not sufficiently discuss the diagenetic processes that could have affected the tests of the kalhori specimens. Specifically, it is possible that the specimens illustrated from the washed conodont residues do not display the originally preserved wall. Therefore, in this study, we consider Postcladella as a valid genus and Postcladella kalhori as a porcelaneous form belonging to the Class Miliolata.
Cornuspirins recognized in the Chinese samples consist of three species within the studied interval. Among these taxa, the individuals assigned to the Postcladella kalhori population (Fig. 3.1–3.26) are nearly identical to types described under the genus Rectocornuspira by Brönnimann et al. (Reference Brönnimann, Zaninetti and Bozorgnia1972a); however, the size variation in the Chinese forms is much greater. The diameter of the coiled portion reaches up to 160 μm, as noted previously in Altıner and Zaninetti (Reference Altıner and Zaninetti1981). The number of whorls, reaching to 2.5 or 3 in some specimens, is also greater than in the type specimens, which display 1.5–2 whorls. Previously, P. kalhori was generally reported from the microbialite-bearing successions from the base of the Triassic (Zaninetti, Reference Zaninetti1976; Rettori, Reference Rettori1995 and the references therein). In the more recent literature, P. kalhori has been reported as Cornuspira mahajeri from the Griesbachian of Antalya Nappes, Turkey (Angiolini et al., Reference Angiolini, Carabelli, Nicora, Crasquin-Soleau, Marcoux and Rettori2007); Alborz Mountains, Iran (Angiolini et al., Reference Angiolini, Checconi, Gaetani and Rettori2010); western Slovenia (Nestell et al., Reference Nestell, Kolar-Jurkovšek, Jurkovšek and Aljinović2011); the Dead Sea region, Jordan (Powell et al., Reference Powell, Stephenson, Nicora, Rettori, Borlengi and Perri2016); and from the Nanpanjiang basin, south China (Bagherpour et al., Reference Bagherpour, Bucher, Baud, Brosse, Vennemann, Martini and Goudun2017). Hips (Reference Hips1996) reported P. kalhori from the Spathian of northern Hungary, but did not satisfactorily illustrate these forms. The associated forms illustrated as Cyclogyra mahajeri were misidentified and belong to Pseudoammodiscus (formerly known as Ammodiscus). The same problem has also occurred in the description of material from Israel. Korngreen et al. (Reference Korngreen, Orlov-Labkovsky, Bialik and Benjamini2013) mentioned the presence of P. kalhori in strata of later Early Triassic age, but these forms were not illustrated in their study. Postcladella also has been partly misidentified in Rossignol et al. (Reference Rossignol, Bourquin, Hallot, Poujol, Debard, Martini, Villeneuve, Cornée, Brayard and Roger2018) from northern Vietnam. Their figure 11C and 11E illustrated specimens that are Glomospirella vulgaris Ho, Reference Ho1959, not Postcladella (Rossignol et al., Reference Rossignol, Bourquin, Hallot, Poujol, Debard, Martini, Villeneuve, Cornée, Brayard and Roger2018, fig. 11E is associated with Arenovidalina sp.).
In both the Dawen and Daijiang sections, P. kalhori ranges from the base of Griesbachian to the lowermost Dienerian (Figs. 5, 6). The stratigraphic range in the Chinese sections is indistinguishable from the range given in Rettori (Reference Rettori1995). In several recent studies, including Crasquin-Soleau et al. (Reference Crasquin-Soleau, Richoz, Marcoux, Angiolini, Nicora and Baud2002, Reference Crasquin-Soleau, Marcoux, Angiolini, Richoz, Nicora, Baud and Bertho2004) from the Antalya Nappes, Turkey, Song et al. (Reference Song, Tong, Chen, Yang and Wang2009, Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016), Yang et al. (Reference Yang, Chen, Wang, Tong, Song and Chen2011), and Dai et al. (Reference Dai, Song, Wignall, Jia, Bai, Wang, Chen and Tian2018) from south China, and Kolar-Jurkovšek et al. (Reference Kolar-Jurkovšek, Jurkovšek, Nestell and Aljinović2018) from western Slovenia, the stratigraphic range of P. kalhori has been partially studied in stratigraphic sections covering only a few meters of the base of Triassic and reported as Griesbachian based on conodont zones. Galfetti et al. (Reference Galfetti, Bucher, Martini, Hochuli, Weissert, Crasquin-Soleau, Brayard, Goudmand, Brühwiler and Goudun2008) reported the full range as Griesbachian to Dienerian from the Nanpanjiang Basin, similar to the range given in this study. Some authors reported this interval simply as Induan (Insalaco et al., Reference Insalaco, Virgone, Courme, Gaillot, Kamali, Moallemi, Loftpour and Monibi2006 from southern Iran; Powell et al., Reference Powell, Nicora, Perri, Rettori, Posenato, Stephenson, Masri, Borlenghi and Gennari2019 from Jordan). Krainer and Vachard (Reference Krainer and Vachard2011) reported the stratigraphic distribution of P. kalhori from the Werfen Formation (southern Austria) also as Induan, based on Broglio Loriga et al. (Reference Broglio Loriga, Góczán, Haas, Lenner, Neri, Orovecz-Scheffer, Posenato, Szabó and Makk1990), and added that this species also could be present in the Olenekian. However, neither in Krainer and Vachard (Reference Krainer and Vachard2011) nor other studies have properly illustrated the kalhori population from well-dated Olenekian strata.
Postcladella grandis (Altıner and Zaninetti, Reference Altıner and Zaninetti1981)
Figures 3.27–3.38, 4.1–4.16

Figure 4. All specimens are from Dawen (PDW) and Dajiang (PDJ) sections. (1–16) Postcladella grandis (Altıner and Zaninetti, Reference Altıner and Zaninetti1981); (17) Cornuspira mahajeri? (Brönnimann, Zaninetti, and Bozorgnia, Reference Brönnimann, Zaninetti and Bozorgnia1972a). (1, 17) PDW-108; (2) PDJ-178; (3) PDW-102; (4, 6) PDW-096; (5) PDJ-164; (7, 10, 11, 13–15) PDW-118; (8, 16) PDW-123; (9) PDJ-158; (12) PDJ-157. Scale bars = 100 μm.
Remarks
The other species that we recognize in the Chinese material also belongs to the genus Postcladella. Introduced by Altıner and Zaninetti (Reference Altıner and Zaninetti1981) from Turkey, R. kalhori f. grandis was raised by Lehrmann et al. (Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015) to a species rank under the genus Postcladella. Cyclogyra nov. sp.? of Resch (Reference Resch1979) and Rectocornuspira reschi of Orovecz-Scheffer (Reference Orovecz-Scheffer1983) were considered as synonyms of Postcladella grandis (Figs. 3.27–3.38, 4.1–4.16). Although Rettori (Reference Rettori1995) considered Postcladella grandis to be a synonym of P. kalhori, the former species is easily distinguished from the latter by the larger size of the tubular chamber, which is nearly twice that of P. kalhori in every step of its ontogeny, and the larger diameter of the coiled portion of the test. In addition, the first appearance of P. grandis always postdates that of P. kalhori in south China, Turkey, and the Transdanubian Range of Hungary (Altıner and Zaninetti, Reference Altıner and Zaninetti1981; Orovecz-Scheffer, Reference Orovecz-Scheffer1987; Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). Postcladella grandis was also illustrated or described from other studies in the Tethyan belt. For example, the specimens illustrated as R. kalhori in Brönnimann et al. (Reference Brönnimann, Zaninetti and Bozorgnia1972a, pl. 4, fig. 15) and Cyclogyra? mahajeri in Brönnimann et al. (Reference Brönnimann, Zaninetti and Bozorgnia1972a, pl. 4, fig. 18) should be assigned to P. grandis. Forms from Austria illustrated by Krainer and Vachard (Reference Krainer and Vachard2011, pl. 5, figs. 9, 10, 14) are referable to P. grandis. Cyclogyra? mahajeri illustrated from the Alborz Mountains, Iran by Stampfli et al. (Reference Stampfli, Zaninetti, Brönnimann, Jenny-Deshusses and Stampfli-Vuille1976) is P. grandis. In addition, P. grandis has been illustrated as both C.? mahajeri and R. kalhori in Orovecz-Scheffer (Reference Orovecz-Scheffer1987).
The stratigraphic range of P. grandis is much shorter than the range of P. kalhori (Figs. 5, 6). The first occurrence of P. grandis is higher in the Griesbachian in China than in Turkey (Altıner and Zaninetti, Reference Altıner and Zaninetti1981), and its last occurrence is just below the Griesbachian-Dienerian boundary in the Dawen and Daijiang sections. The stratigraphic range of C. mahajeri? is similar to that of P. kalhori, as stated by Rettori (Reference Rettori1995). Forms close to this species are present both in Dawen and Dajiang sections, within the interval corresponding to the stratigraphic range of P. grandis.
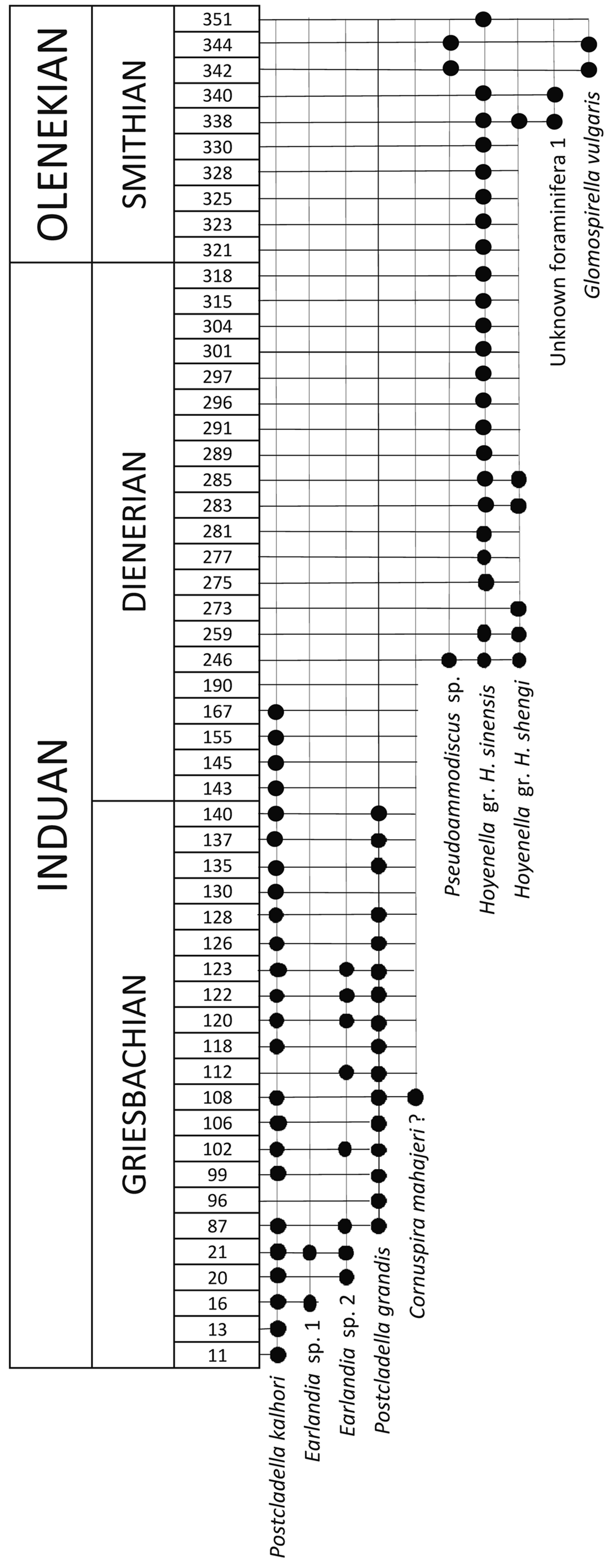
Figure 5. Stratigraphic occurrences of foraminiferal taxa at Dawen measured section (barren samples not shown). Because the goal of the figure is to illustrate the occurrences and co-occurrences of species for the purpose of chronostratigraphy, only fossiliferous samples are included along with sample numbers. For stratigraphic positions of illustrated samples, we refer readers to Figure 2.

Figure 6. Stratigraphic occurrences of foraminiferal taxa at Dajiang measured section (barren samples not shown). In this section, samples numbered less than 44 were collected from the Permian strata and are not considered in this study.
Genus Cornuspira Schultz, Reference Schultze1854
Cornuspira mahajeri? (Brönnimann, Zaninetti, and Bozorgnia, Reference Brönnimann, Zaninetti and Bozorgnia1972a)
Figure 4.15
Remarks
Rarely recorded forms assigned to Cornuspira mahajeri? (Fig. 4.15) in this study differ from the types (given as Cyclogyra? mahajeri by Brönnimann et al., Reference Brönnimann, Zaninetti and Bozorgnia1972a) in having slightly larger dimensions of the test and the lumen of the final whorl covering the previous whorl in a less-pronounced way.
Subfamily Calcivertellinae Loeblich and Tappan, Reference Loeblich, Tappan and Moore1964
Genus Planiinvoluta Leischner, Reference Leischner1961
Planiinvoluta? mesotriasica Baud, Zaninetti, and Brönnimann, Reference Baud, Zaninetti and Brönnimann1971
Figure 7.1–7.7

Figure 7. All specimens are from Guandao (PGD), Upper Guandao (PUG) and Middle Triassic Dajiang (MDJ) sections. (1–7) Planiinvoluta? mesotriasica Baud, Zaninetti, and Brönnimann, Reference Baud, Zaninetti and Brönnimann1971; (8–13) Arenovidalina weii n. sp.; (14, 15) Arenovidalina abriolense (Luperto, Reference Luperto1965); (16–20) Meandrospira pusilla (Ho, Reference Ho1959); (21–24, 26) Meandrospira cheni (Ho, Reference Ho1959); (25, 27–32) Meandrospira dinarica Kochansky-Devidé and Pantić, Reference Kochansky-Devidé and Pantić1966. (1) MDJ-025; (2) PUG-065; (3, 5) PUG-085; (4) MDJ-039; (6, 32) PUG-054; (7) MDJ-035; (8) PGD-131; (9) PGD-112 (holotype); (10) PGD-111; (11) PGD-109; (12, 13) PGD-096; (14) PUG-051; (15) PUG-089; (16) MDJ-007; (17, 19) PGD-174; (18) PUG-012; (20, 31) PUG-009; (21) PGD-151; (22) PGD-168; (23) PGD-154; (24, 26) PGD-157; (25) PGD-226; (27) MDJ-019; (28) PUG-027; (29) PGD-215; (30) PUG-023. Scale bars = 100 μm.
Remarks
The subfamily Calcivertellinae, a group of tubular, porcelaneous foraminifera, is represented only by Planiinvoluta? mesotriasica (Fig. 7.1–7.7) in the studied material. The size of the proloculus, diameter of the tubular chamber, and the test are similar to those of the types originally described by Baud et al. (Reference Baud, Zaninetti and Brönnimann1971). However, the genus attribution remains unclear because the individuals belonging to this species never display a proper planispirally enrolled second chamber as described by Leischner (Reference Leischner1961). In the recent literature, P.? mesotriasica has been clearly illustrated in Altıner and Koçyiğit (Reference Altıner and Koçyiğit1993) and Okay et al. (Reference Okay, Altıner and Kiliç2015) from the Anisian of central and northern Anatolia (Turkey), and in Fugagnoli and Posenato (Reference Fugagnoli and Posenato2004) from northern Italy. The specimens illustrated as Meandrospira? sp. from western Kyushu, Japan, by Kobayashi et al. (Reference Kobayashi, Martini and Zaninetti2005) are most probably sections of P.? mesotriasica. The stratigraphic range of P.? mesotriasica is from Bithynian to Illyrian in the Upper Guandao section and Illyrian in the Middle Triassic Dajiang section (Figs. 8–10).

Figure 8. Stratigraphic occurrences of foraminiferal taxa in the lower part of Upper Guandao section (barren samples not shown). OL.: Olenekian; Spat.: Spathian.

Figure 9. Stratigraphic occurrences of foraminiferal taxa in the upper part of Upper Guandao section (barren samples not shown).
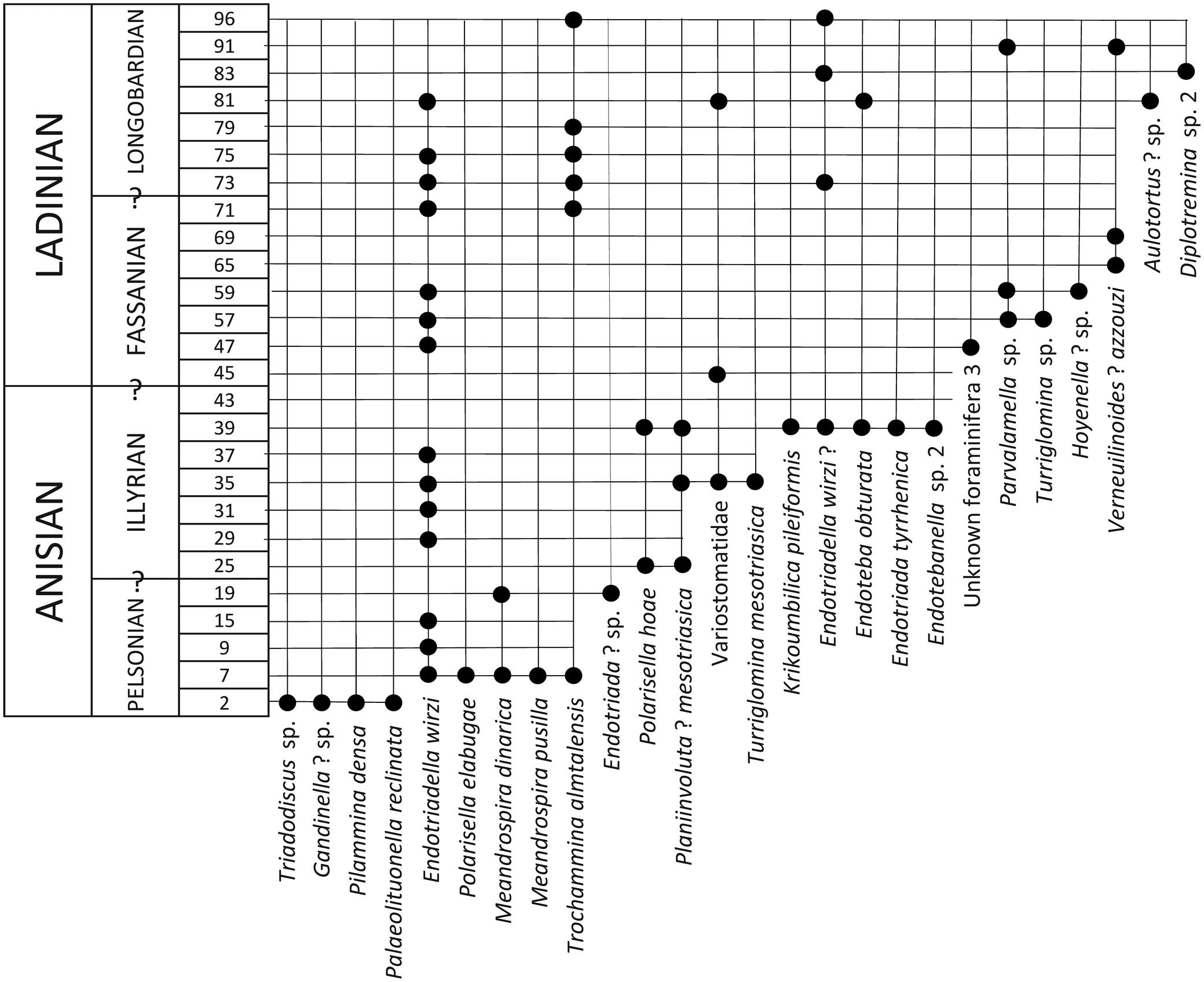
Figure 10. Stratigraphic occurrences of foraminiferal taxa at Middle Triassic Dajiang measured section (barren samples not shown).
Family Arenovidalinidae Zaninetti and Rettori in Zaninetti et al., Reference Zaninetti, Rettori, He and Martini1991
Subfamily Arenovidalininae Zaninetti and Rettori in Zaninetti et al., Reference Zaninetti, Rettori, He and Martini1991
Genus Arenovidalina Ho, 1959
Type species
Arenovidalina chialingchiangensis Ho, Reference Ho1959, from the Triassic Chialingkiang Limestone of South Szechuan, China.
Arenovidalina abriolense (Luperto, Reference Luperto1965) Figure 7.14, 7.15
Remarks
Two species from the subfamily Arenovidalininae, Arenovidalina abriolense (Luperto, Reference Luperto1965) (Fig. 7.14, 7.15) and Arenovidalina weii n. sp. (Fig. 7.8–7.13), are present in the Chinese sections. Arenovidalina abriolense, previously defined as a Permian form and thus placed under the genus Hemigordius by Luperto (Reference Luperto1965), was later placed in the genus Ophthalmidum by Ciarapica et al. (Reference Ciarapica, Cirilli, Martini, Rettori, Salvini-Bonnard and Zaninetti1990). Consisting of a proloculus followed by an undivided second chamber, the abriolense population does not carry the primitively developed multilocular character of the genus Ophthalmidium, and is therefore placed under the genus Arenovidalina in this study. Arenovidalina abriolense differs from the two other, similar-looking species of Arenovidalina, A. chialingchiangensis Ho, Reference Ho1959 and A. amylovoluta Ho, Reference Ho1959 (Ho, Reference Ho1959; Zaninetti, Reference Zaninetti1976; Okay et al., Reference Okay, Altıner and Kiliç2015) in having a more tightly enrolled and slowly growing second chamber. In Senowbari-Daryan et al. (Reference Senowbari-Daryan, Zühlke, Bachtädt and Flügel1993), Ophthalmidium (=Arenovidalina) chialingchiangensis illustrated from the Anisian of the northern Dolomites, Italy, belongs to A. abriolense. Some specimens illustrated as Ophthalmidium spp. from the Anisian of western Kyushu, Japan, are referable to A. abriolense (Kobayashi et al., Reference Kobayashi, Martini and Zaninetti2005). Ophthalmidium abriolense illustrated in Velledits et al. (Reference Velledits, Péró, Blau, Senowbari-Daryan, Kovács, Piros, Pocsai, Szúgyi-Simon, Dumitrica and Palfy2011) from NE Hungary is Eoophthalmidium tricki Langer, Reference Langer1968. The stratigraphic range of A. abriolense is Anisian (Aegean) to Carnian (Cordevolian) in the Upper Guandao section (Figs. 8, 9). The description of the other Arenovidalina species, A. weii n. sp., is given below.
Arenovidalina weii new species
Figure 7.8–7.13
- ?1990
Arenovidalina amylovoluta Ho; Baroz et al., pl. 4, figs. 8, 9.
Holotype
PGD-112 (Fig. 7.9).
Diagnosis
A laterally compressed Arenovidalina species with a maximum of four whorls, becoming evolute in the late stage of the ontogeny.
Occurrence
Smithian, Guandao section of the Great Bank of Guizhou, Nanpanjiang Basin, south China, Sample PGD-112 (Figs. 2, 11).

Figure 11. Stratigraphic occurrences of foraminiferal taxa at Guandao measured section (barren samples not shown).
Description
Test is laterally compressed and consists of a relatively large proloculus and a maximum number of four planispiral whorls rapidly increasing in height. The coiling, involute in the earlier 2.5 whorls, becomes evolute in the last 1.5 whorls (Fig. 7.9). A possible dimorphism is present in the population (Fig. 7.10). In microspheric forms, the number of whorls increases to five and the test displays a slight deviation in the axis of coiling in the initial 1–2 whorls. The wall is originally porcelaneous and appears slightly granular due to recrystallization. It is relatively thicker when compared with the volume of the test.
Etymology
This new species is dedicated to Dr. Jiayong Wei of the Guizhou Geological Survey, China, who made great contributions to the understanding of the geology of the Nanpanjiang Basin.
Materials
Samples PGD-96, 109, 111, 112, 131 (Smithian). More than 20 specimens. Six of these specimens, including the holotype, are illustrated in Figure 7.8–7.13.
Microfossil association
In the samples collected from the Smithian, Arenovidalina weii n. sp. is associated with Hoyenella gr. H. sinensis, H. gr. H. shengi, Glomospirella vulgaris, Pseudoammodiscus sp., Praetriadodiscus zaninettiae Altıner and Payne, Reference Altıner and Payne2017, and P. tappanae Altıner and Payne, Reference Altıner and Payne2017.
Dimensions
Diameter of proloculus: 50–55 μm. Diameter of test: 360–420 μm. Width of test: 100–115 μm. W/D ratio: 0.24–0.32. Height of last whorl (only lumen): 55–60 μm. Thickness of wall: 10–14 μm.
Remarks
Arenovidalina weii n. sp. is probably the oldest Arenovidalina species known in the Triassic stratigraphy. It differs from the two other well-known species, A. chialingchiangensis and A. amylovoluta, in having a nearly parallel-sided, compressed test and few whorls, which increase rapidly in height and become evolute toward the end of the ontogeny. The specimens illustrated as Arenovidalina amylovoluta by Baroz et al. (Reference Baroz, Martini and Zaninetti1990) from Greece probably belong to our A. weii n. sp. population. These forms are similar to A. weii n. sp. in having highly compressed axial profiles and evolute last whorls, but they contain more whorls and a smaller proloculus. Arenovidalina sp. illustrated by Kobayashi (Reference Kobayashi1996) from the Anisian of the Kanto Mountains, Japan, is also characterized by involute earlier whorls and evolute later whorls. However, the D/W ratio is greater in the Japanese forms. Finally, the microfacies photographs illustrated by Song et al. (Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016, fig. 9c) from the Smithian of south China contain sections of A. weii n. sp.
Family Meandrospiridae Saidova, Reference Saidova1981 emend. Zaninetti et al., Reference Zaninetti, Ciarapica, Martini, Salvini-Bonnard and Rettori1987a
Subfamily Meandrospirinae Saidova, Reference Saidova1981 emend. Zaninetti et al., Reference Zaninetti, Ciarapica, Martini, Salvini-Bonnard and Rettori1987a
Remarks
Meandrospirin foraminifera of the Great Bank of Guizhou are classified under two genera, Meandrospira and Meandrospiranella. The validity of the former genus in the Triassic stratigraphy has been questioned recently by Ueno et al. (Reference Ueno, Miyahigashi and Martini2018), who suggested the replacement of Meandrospira with Citaella, which was originally described by Premoli Silva (Reference Premoli Silva1964). These authors concluded that Citaella, Meandrospira, and Streblospira are meandrospiral homeomorphs that appeared independently at different times in the evolutionary history of the class Miliolata. In contrast, we argue that Triassic Meandrospira was derived as a Lazarus taxon from Permian Streblospira, as argued by Groves and Altıner (Reference Groves and Altıner2005) and Altıner et al. (Reference Altıner, Groves and Özkan-Altıner2005). Recently, Streblospira (S. minima) Kotljar et al., Reference Kotljar, Zakharov, Koczyrkevica, Chedija, Vuks and Guseva1984 has been illustrated from the Changhsingian of western Slovenia by Kolar-Jurkovšek et al. (Reference Kolar-Jurkovšek, Jurkovšek, Nestell and Aljinović2018). Previous findings of Streblospira within middle and upper Permian strata (Kotljar et al., Reference Kotljar, Zakharov, Koczyrkevica, Chedija, Vuks and Guseva1984; Şahin et al., Reference Şahin, Altıner and Ercengiz2012) demonstrate that the genus was not confined to lower Permian strata, as was supposed by Ueno et al. (Reference Ueno, Miyahigashi and Martini2018). Furthermore, we consider that the ‘absence’ of Meandrospira in post-Anisian to Jurassic rocks may not indicate the ‘phylogenetical isolation’ of ‘Citaella’ in the Early to Middle Triassic. Tubular foraminifera might survive as rare members of communities, confined to narrow ecological and/or environmental niches before becoming more common again in a much younger time interval, as in Meandrospira (Charollais et al., Reference Charollais, Brönnimann and Zaninetti1966; Altıner, Reference Altıner1991; Ivanova and Kołodziej, Reference Ivanova and Kołodziej2010). Because the ancestor of Cretaceous Meandrospira is not known from the Cretaceous, we take the more conservative route and continue to use Meandrospira in Triassic foraminiferal taxonomy.
Genus Meandrospira Loeblich and Tappan, Reference Loeblich and Tappan1946
Type species
Meandrospira washitensis Loeblich and Tappan, Reference Loeblich and Tappan1946, from the Lower Cretaceous Washita Group of southern Oklahoma and northern Texas, USA.
Meandrospira pusilla (Ho, Reference Ho1959)
Figure 7.16–7.20
Remarks
From the Meandrospira population in the Chinese material, the well-known species M. pusilla (Ho, Reference Ho1959) (Fig. 7.16–7.20), characterized by 1.5–2 whorls and a tightly coiled spire consisting of 8–9 sections of the zig-zag bends of the meandering second tubular chamber in the last whorl, is nearly identical to the holotype described by Ho (Reference Ho1959). Meandrospira pusilla was extensively reported from the whole Tethyan Belt (Zaninetti, Reference Zaninetti1976; Rettori, Reference Rettori1995). More recently, it has been reported from southern Austria by Krainer and Vachard (Reference Krainer and Vachard2011) and from Jordan by Powell et al. (Reference Powell, Stephenson, Nicora, Rettori, Borlengi and Perri2016). Some of the specimens illustrated as M. pusilla from the Qingyan section of south China by Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015) belong to M. dinarica Kochansky-Devidé and Pantić, Reference Kochansky-Devidé and Pantić1966.
Meandrospira pusilla ranges from the Spathian to the Bithynian in the Guandao and Upper Guandao sections (Figs. 8, 11). It is also present in the Pelsonian of the Middle Triassic Dajiang section (Fig. 10). In the Smithian of the Daijiang section (Fig. 6), forms identified as ‘transitional to Meandrospira’ could also be included in the population of M. pusilla. Meandrospira cheni is strictly confined to the Spathian in the Guandao section (Fig. 11). Meandrospira? deformata has been recognized from the Pelsonian to lower Illyrian of the Upper Guandao section (Fig. 8).
Meandrospira cheni (Ho, Reference Ho1959)
Figure 7.21–7.24, 7.26
Remarks
Meandrospira cheni (Ho, Reference Ho1959) (Fig. 7.21–7.24, 7.26), a similarly coiled but larger form than M. pusilla, differs from M. pusilla in having fewer (6–8) zig-zag bends of the meandering second tubular chamber. Our specimens are nearly identical to forms illustrated as the cheni population in Ho (Reference Ho1959) and He (Reference He1993) from China. Some of the M. pusilla sections illustrated from Julfa, northwestern Iran, by Baud et al. (Reference Baud, Brönnimann and Zaninetti1974) belong to M. cheni. The specimen illustrated from Hydra (Greece) by Rettori et al. (Reference Rettori, Angiolini and Muttoni1994) is probably an Endoteba section. Meandrospira cheni also has been reported from the Spathian of Israel (Korngreen et al., Reference Korngreen, Orlov-Labkovsky, Bialik and Benjamini2013) and the northern Arab Emirates (Maurer et al., Reference Maurer, Rettori and Martini2008).
Meandrospira dinarica Kochansky-Devidé and Pantić, Reference Kochansky-Devidé and Pantić1966
Figures 7.25, 7.27–7.32, 12.1–12.4
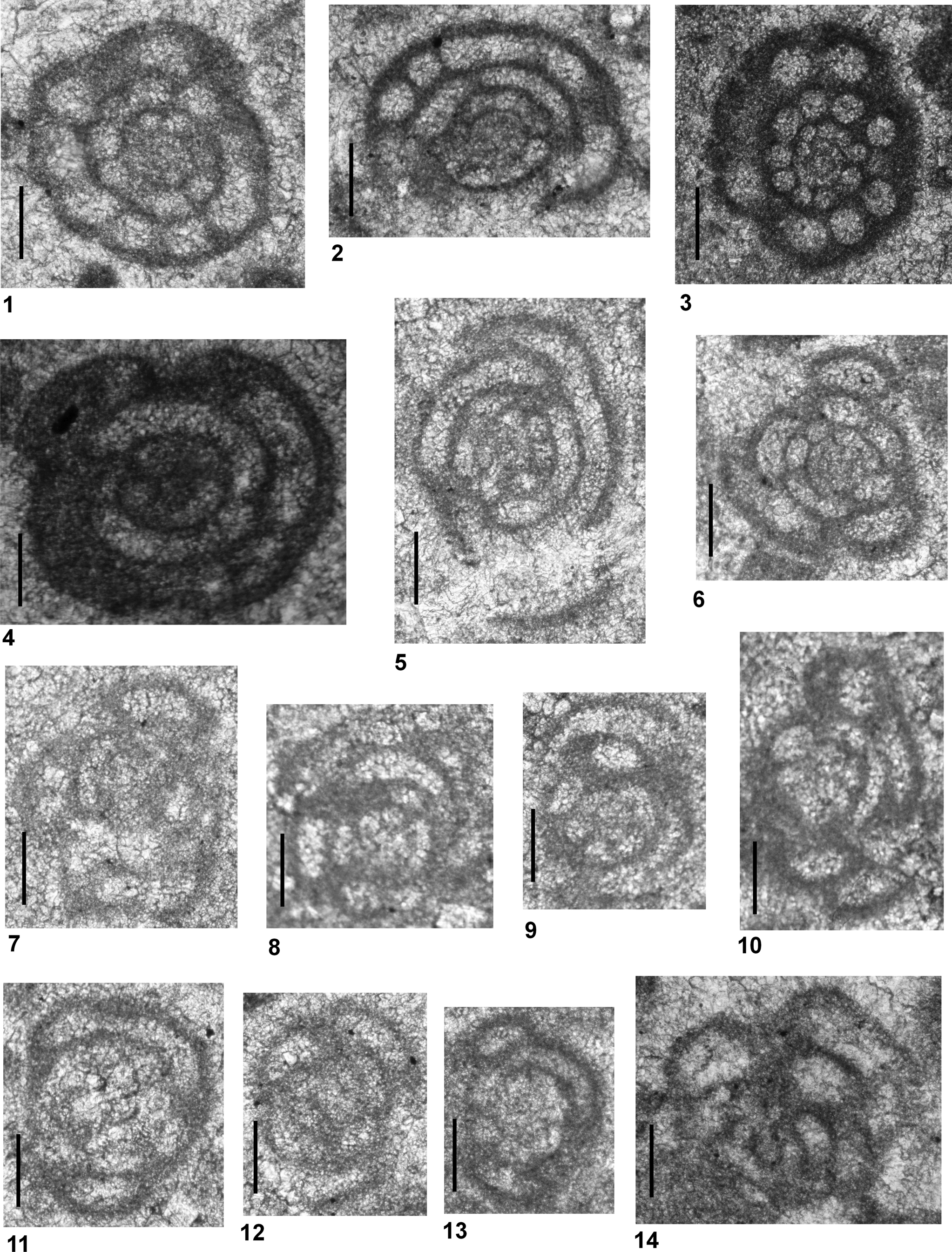
Figure 12. All specimens are from Guandao (PGD) and Upper Guandao (PUG) sections. (1–4) Meandrospira dinarica Kochansky-Devidé and Pantić, Reference Kochansky-Devidé and Pantić1966 (5–13) Meandrospira? enosi n. sp.; (14) Meandrospira? deformata Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967. (1) PUG-037; (2) PUG-031; (3) PGD-212; (4) PUG-019; (5–12) PGD-176; (13) PGD-174; (14) PUG-054. Scale bars = 100 μm.
Remarks
The other well-established species of Meandrospira, M. dinarica (Figs. 7.25, 7.27–7.32, 12.1–12.4), consisting of 8–10 sections of the second tubular chamber in the last whorl, is characterized by rectangular outlines of the sections of the tubular chamber in the equatorial plane. Meandrospira dinarica is one of the most widely cited Anisian species in the Tethyan Belt (Zaninetti, Reference Zaninetti1976; Rettori, Reference Rettori1995). In more recent literature, it has been reported from the Anisian of Japan (Kobayashi, Reference Kobayashi1996; Kobayashi et al., Reference Kobayashi, Martini and Zaninetti2005), Thailand (Kobayashi et al., Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006), Laos (Miyahigashi et al., Reference Miyahigashi, Hara, Hisada, Nakano, Charoentitirat, Charusiri, Khamphoveng, Martini and Ueno2017), Vietnam (Martini et al., Reference Martini, Zaninetti, Cornée, Villeneuve, Tran and Ta1998), northern Italy (Fugagnoli and Posenato, Reference Fugagnoli and Posenato2004; Berra et al., Reference Berra, Rettori and Bassi2005), and Hungary (Velledits et al., Reference Velledits, Péró, Blau, Senowbari-Daryan, Kovács, Piros, Pocsai, Szúgyi-Simon, Dumitrica and Palfy2011). Korchagin (Reference Korchagin2008) identified M. dinarica as M. cheni from Pamirs. Tian et al. (Reference Tian, Tong, Algeo, Song, Song, Chu, Shi and Bottjer2014), based on Song et al. (Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan, Niu, Chen, Yang and Wang2011a), reported earliest M. dinarica from the Smithian–Spathian interval of south China. This unusual report is probably the result of a taxonomic misidentification. Both M. pusilla and M. cheni might have been misidentified as M. dinarica (see also Song et al., Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015).
Meandrospira dinarica, whose first appearance nearly coincides with the Olenekian-Anisian boundary, extends from the Aegean to the Pelsonian in the Guandao and Upper Guandao sections (Figs. 8, 11). It also occurs in the Pelsonian of the Middle Triassic Dajiang section (Fig. 10). From the Meandrospiranella side, M. cf. M. samueli is found in the uppermost Aegean, whereas M. irregularis? occurs in the Pelsonian to lowermost Illyrian of the Upper Guandao section (Fig. 8).
Meandrospira? deformata Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967
Figures 12.14, 13.1–13.4

Figure 13. All specimens are from Dawen (PDW), Dajiang (PDJ), Middle Triassic Dajiang (MDJ), Guandao (PGD), and Upper Guandao (PUG) sections. (1–4) Meandrospira? deformata Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967; (5) Meandrospiranella cf. M. samueli Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967; (6) Meandrospiranella irregularis? Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967; (7–9) Turriglomina mesotriasica (Koehn-Zaninetti, Reference Koehn-Zaninetti1968); (10–13) Turriglomina cf. T. magna (Urošević, Reference Urošević1977); (14) Turriglomina carnica Dağer, Reference Dağer1978b; (15–33) Hoyenella gr. H. sinensis (Ho, Reference Ho1959). (1, 4) PUG-067; (2, 3) PUG-069; (5) PUG-014; (6) PUG-057; (7) PUG-089; (8) PUG-091; (9) MDJ-35; (10) PUG-055; (11) PUG-077; (12) PUG-075; (13) PUG-085; (14) PUG-145; (15, 33) PDW-285; (16) PGD-101; (17) PGD-087; (18) PDW-281; (19) PDW-275; (20, 30) PGD-090; (21) PDW-259; (22) PGD-151; (23) PGD-176; (24) PDJ-302; (25) PDJ-313; (26–28) PDJ-303; (29) PDJ-281; (31) PDW-315; (32) PGD-168. Scale bars = 100μm.
Remarks
Meandrospira? deformata Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967 (Figs. 12.14, 13.1–13.4), a doubtful member of the genus Meandrospira, is characterized by an irregularly coiled meandering second chamber throughout its ontogeny. After the type description of Salaj et al. (Reference Salaj, Biely and Bistricky1967), M.? deformata has been reported from the Anisian of Slovenia (Flügel et al., Reference Flügel, Ramovš and Bucur1994), Japan (Kobayashi, Reference Kobayashi1996), Pamirs (Korchagin, Reference Korchagin2008), and more recently from Laos (Miyahigashi et al., Reference Miyahigashi, Hara, Hisada, Nakano, Charoentitirat, Charusiri, Khamphoveng, Martini and Ueno2017) as Citaella? deformata.
Meandrospira? enosi new species
Figure 12.5–12.13
Holotype
PGD-176 (Fig. 12.5).
Diagnosis
A meandrospirin species characterized by an initial Meandrospira stage followed by oscillating whorls, but not zig-zag bends, of the tubular chamber.
Occurrence
Spathian, Guandao section of the Great Bank of Guizhou, Nanpanjiang Basin, south China, PGD-176 (Figs. 2, 11).
Description
Following a globular and small proloculus, the initial stage of the new species is typically coiled like a Meandrospira with three whorls. The height of the tubular chamber increases rather rapidly in the final whorl of this stage and, generally, seven sections of zig-zag bends of the second tubular chamber are present in the last whorl. Later in the ontogeny, coiling changes its style and direction. This stage is characterized by 1.5 oscillating whorls, generally arranged perpendicular to the axis of coiling of the Meandrospira stage. The wall is porcelaneous, similar to the other Meandrospira species in this study.
Etymology
This new species is dedicated to Prof. Dr. Paul Enos from the University of Kansas, USA, for his contributions to the Permian–Triassic carbonates in China.
Materials
Samples PGD-171, 176, 178, 181 (Spathian). More than 20 specimens. Nine of these specimens are illustrated in Figure 12.5–12.13.
Microfossil association
In the Spathian samples, the new species co-occurs with Hoyenella gr. H. sinensis, Meandrospira pusilla, Meandrospira cheni, Trochammina sp. 1, Verneuilinoides? azzouzi (Salaj, Reference Salaj1978), Endoteba bithynica Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994, Endotebanella kocaeliensis (Dağer, Reference Dağer1978b), Krikoumbilica pileiformis He, Reference He1984, Variostoma sp. 2, and Diplotremina sp. 2.
Dimensions
Diameter of proloculus: 36–50 μm. Diameter of test: 280–340 μm. Height of the last whorl (only lumen): 36–40 μm. Thickness of wall: 15–18 μm.
Remarks
Meandrospira? enosi n. sp., probably derived from M. cheni, is easily distinguished from the other Meandrospira species of the Lower–Middle Triassic by the coiling in the second stage of its ontogeny, characterized by oscillating whorls of the second tubular chamber arranged perpendicular to the coiling axis of the Meandrospira stage. The genus Meandrovoluta, described by Fugagnoli et al. (Reference Fugagnoli, Giannetti and Rettori2003) from the Liassic of the southern Alps, Italy, is morphologically close to M.? enosi n. sp. The coiling style of our new species differs from Meandrovoluta in two ways: (1) Meandrovoluta is characterized by a more irregularly coiled, widely meandering tubular chamber in the second stage of its morphology, whereas M.? enosi n. sp. is characterized by a tubular chamber coiled simply in oscillating whorls; and (2) the Meandrospira-like stage is irregular and complicated in the holotype of Meandrovoluta, whereas this stage is more Meandrospira-like in the M.? enosi n. sp. population, even morphologically close to M. cheni. We remain uncertain whether a new genus should be created to house forms such as the enosi population within meandrospirin foraminifera.
Genus Meandrospiranella Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967
Meandrospiranella cf. M. samueli Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967
Figure 13.5
Remarks
Species doubtfully attributed to Meandrospiranella are rare and sporadic in the Chinese material. Meandrospiranella cf. M. samueli Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967 (Fig. 13.5) is an incomplete specimen lacking the uncoiled portion. Flügel et al. (Reference Flügel, Ramovš and Bucur1994) illustrated a form similar to ours from the Pelsonian of Slovenia. Zaninetti et al. (Reference Zaninetti, Brönnimann and Baud1972c), from Switzerland, and Baroz et al. (Reference Baroz, Martini and Zaninetti1990), from Greece, illustrated specimens very similar to the holotype. Meandrospiranella samueli illustrated by Velledits et al. (Reference Velledits, Péró, Blau, Senowbari-Daryan, Kovács, Piros, Pocsai, Szúgyi-Simon, Dumitrica and Palfy2011) from the Anisian of Hungary probably belongs neither to the genus nor to the species of this form. The illustrated specimen is characterized by a granular wall, biserial chambers in the uncoiled portion, and an unrecognizable coiled portion. Meandrospiranella irregularis? Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967 (Fig. 13.6) is characterized by a well-defined, broadly meandering, uncoiled portion that has not been illustrated in the holotype (Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967).
Subfamily Turriglomininae Zaninetti in Limogni et al., Reference Limogni, Panzanelli-Fratoni, Ciarapica, Cirilli, Martini, Salvini-Bonnard and Zaninetti1987
Genus Turriglomina Zaninetti in Limogni et al., Reference Limogni, Panzanelli-Fratoni, Ciarapica, Cirilli, Martini, Salvini-Bonnard and Zaninetti1987
Remarks
Characterized by an initial meandrospiroid or glomospiroid stage followed by a tightly coiled helicoidal stage consisting of numerous whorls, the genus Turriglomina is composed of three species in the Triassic of the Great Bank of Guizhou. Turriglomina mesotriasica (Koehn-Zaninetti, Reference Koehn-Zaninetti1968) (Fig. 13.7–13.9), as described by Koehn-Zaninetti (Reference Koehn-Zaninetti1969), consists of a slender test with up to 14 tightly coiled whorls. Turriglomina mesotriasica has been extensively reported from the Triassic deposits of the Tethyan Belt (Zaninetti, Reference Zaninetti1976; Rettori, Reference Rettori1995). In more recent literature, Emmerich et al. (Reference Emmerich, Zamparelli, Bechstädt and Zühlke2005) reported this form from the Illyrian to Ladinian of the Latemar (Dolomites, Italy). Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015) illustrated this species (given as Turritellella mesotriasica) from the Qingyan section of south China. However, the form illustrated by Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015) does not display the slender aspect of T. mesotriasica and should be corrected as T. conica He, Reference He1984 from Guizhou, south China. Turriglomina cf. T. magna (Urošević, Reference Urošević1977) (Fig. 13.10–13.13), a relatively larger form, is always found as incomplete oblique sections. Here we note that T. guangxiensis described by He and Cai (Reference He and Cai1991) from the Middle Triassic of Guangxi, China, is a synonym of T. magna. Turriglomina carnica Dağer, Reference Dağer1978b is very rare and recognized in oblique sections with spiny extensions from the periphery of the test (Fig. 13.14).
Turriglomina mesotriasica ranges from the upper Pelsonian to the base of the Carnian (Cordevolian) in the Upper Guandao section (Figs. 8, 9). It occurs also in the Illyrian of the Middle Triassic Dajiang section (Fig. 10). Turriglomina cf. T. magna has been recognized in the upper Pelsonian–Illyrian, whereas T. carnica occurs rarely in the Carnian of the Upper Guandao section (Figs. 8, 9)
Turriglomina mesotriasica (Koehn-Zaninetti, Reference Koehn-Zaninetti1968)
Figure 13.7–13.9
Remarks
Koehn-Zaninetti (Reference Koehn-Zaninetti1968) published a condensed version of Koehn-Zaninetti (Reference Koehn-Zaninetti1969) with the same title in which she reported the new taxa that she discovered in this paper. Although these taxa were also described as new in 1969 (her main work) we give priority to the 1968 publication. See additional remarks under Genus Turriglomina.
Turriglomina cf. T. magna (Urošević, Reference Urošević1977)
Figure 13.10–13.13
Remarks
See Remarks under Genus Turriglomina.
Remarks
See Remarks under Genus Turriglomina.
Remarks
A variety of sections belonging to the genus Hoyenella, characterized by an ovoid to discoidal test with a large globular proloculus, an early stage coiled in several different planes, and a planispirally coiled later stage, have been grouped under two distinct and two questionable populations.
Hoyenella gr. H. sinensis (Ho, Reference Ho1959)
Figures 13.15–13.33, 14.1
Remarks
Although nearly all early hoyenellid forms of Early to Middle Triassic age (Glomospirella sinensis var. rara Ho, Reference Ho1959, Glomospirella facilis Ho, Reference Ho1959, and Glomospirella shengi Ho, Reference Ho1959; Glomospirella elbursorum Brönnimann et al., Reference Brönnimann, Zaninetti, Bozorgnia and Huber1972b and Palaeonubecularia minuta Brönnimann et al., Reference Brönnimann, Zaninetti, Bozorgnia and Huber1972b; and Calcitornella gebzensis Dağer, Reference Dağer1978b) have been synonymized under Hoyenella gr. H. sinensis by Rettori (Reference Rettori1994, Reference Rettori1995), we distinguish two main hoyenellid groups in the material of the Great Bank of Guizhou. Hoyenella gr. H. sinensis (Figs. 13.15–13.33, 14.1) differs from H. gr. H. shengi (Fig. 14.2–14.9) by its more voluminous irregularly coiled early stage, whereas in the H. gr. H. shengi population, planispiral coiling in the later stage is more prominent. In addition to taxa synonymized under H. gr. H. sinensis by Rettori (Reference Rettori1995), we also consider Glomospira roesingi Blau et al., Reference Blau, Wenzel, Senff and Lukas1995, described from the Scythian–Anisian of Germany, as a synonym of H. gr. H. sinensis based on the similarity of the wall composition and the coiling of the tubular chamber.
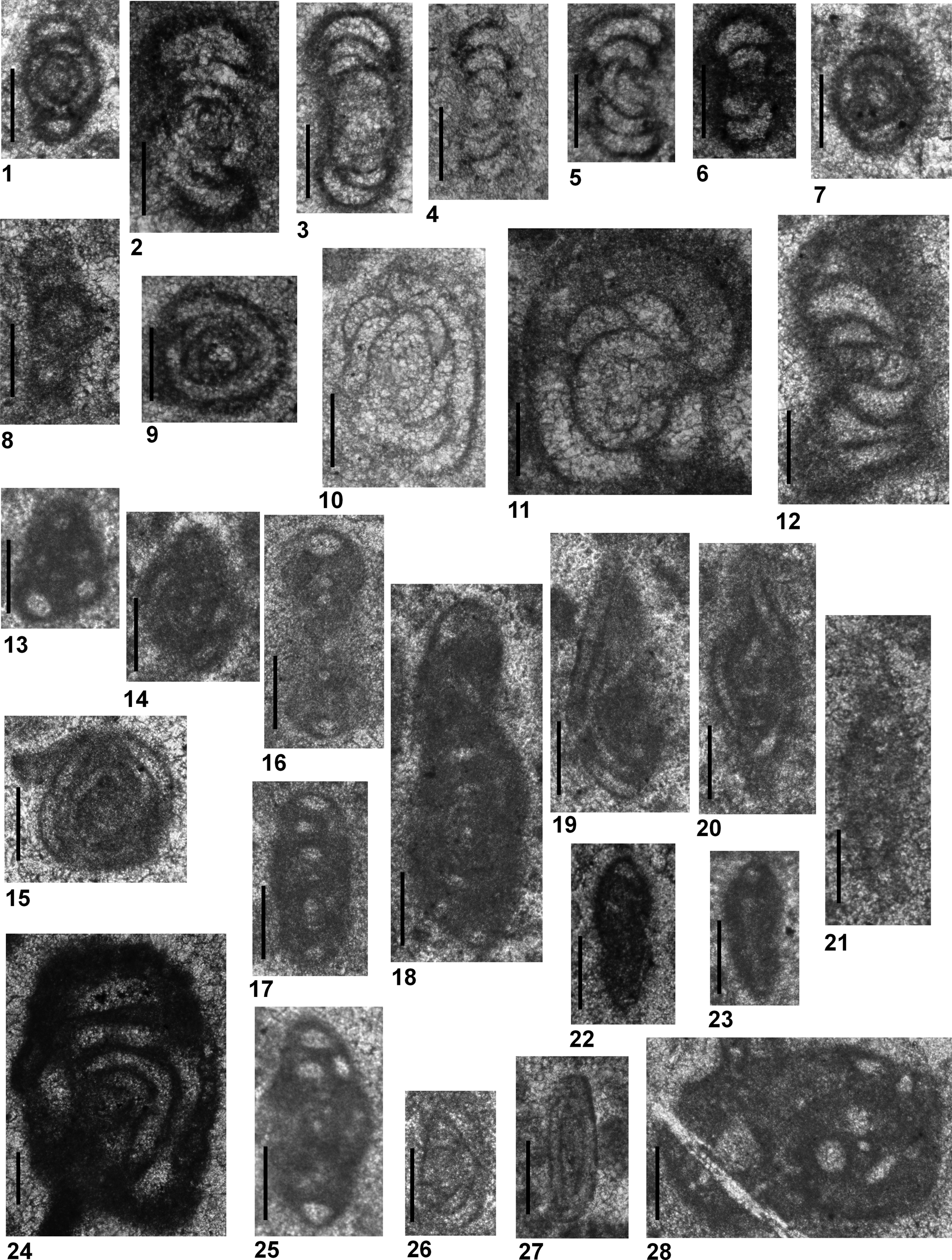
Figure 14. All specimens are from Dawen (PDW), Dajiang (PDJ), Guandao (PGD), and Upper Guandao (PUG) sections. (1) Hoyenella gr. H. sinensis (Ho, Reference Ho1959); (2–9) Hoyenella gr. H. shengi (Ho, Reference Ho1959); (10) Hoyenella? sp. 1; (11, 12) Hoyenella? sp. 2; (13, 14) Agathammina? sp.; (15) Ophthalmidium exiguum Koehn-Zaninetti, Reference Koehn-Zaninetti1969; (16–18) Ophthalmidium sp. 1; (19–21) Ophthalmidium sp. 2; (22, 23) Ophthalmidium sp. 3; (24) Ophthalmidium? sp. 5; (25) Ophthalmidium sp. 4; (26) Gsollbergella? sp.1; (27) Gsollbergella? sp. 2; (28) Galeanella sp. (1, 7) PGD-090; (2, 6) PDW-273; (3) PUG-009; (4, 5) PDW-259; (8) PGD-096; (9) PDJ-302; (10) PUG-027; (11) PGD-226; (12) PUG-101; (13, 14, 17–20, 23) PUG-143; (15) PUG-109; (16, 25) PUG-137; (21) PUG-131; (22) PUG-047; (24) PGD-217; (26) PUG-097; (27) PUG-087; (28) PUG-139. Scale bars = 100 μm.
In the Dajiang and Dawen sections (Figs. 5, 6), H. gr. H. sinensis and H. gr. H. shengi make their first appearances in the upper Dienerian strata. In the Guandao section these species range up to the Bithynian (Fig. 11), whereas in the Upper Guandao section they range into the Illyrian (Fig. 8). In Israel (Korngreen et al., Reference Korngreen, Orlov-Labkovsky, Bialik and Benjamini2013), Kashmir Valley, India (Baud and Bhat, Reference Baud and Bhat2014), and south China (Galfetti et al., Reference Galfetti, Bucher, Martini, Hochuli, Weissert, Crasquin-Soleau, Brayard, Goudmand, Brühwiler and Goudun2008) the first appearance of H. gr. H. sinensis also has been reported as Dienerian in successions that are well dated by conodonts.
Hoyenella gr. H. shengi (Ho, Reference Ho1959)
Figure 14.2–14.9
Remarks
Two other, questionable hoyenellid species, H.? sp. 1 (Fig. 14.10) and H.? sp. 2 (Fig. 14.11, 14.12), characterized by more complicated coiling of the tubular chamber, are rare. Hoyenella? sp. 1 has been recorded from the uppermost Bithynian in the Upper Guandao section (Fig. 8).
Hoyenella? sp. 2
Figure 14.11, 14.12
Remarks
See Remarks under Hoyenella? sp. 1. Hoyenella? sp. 2 ranges from the Bithynian to lower Ladinian (Fassanian) in the Guandao and Upper Guandao sections (Figs. 9, 11).
Family Agathamminidae Ciarapica, Cirilli, and Zaninetti in Ciarapica et al., Reference Ciarapica, Cirilli, Passeri, Trincianti and Zaninetti1987
Genus Agathammina Neumayr, Reference Neumayr1887
Agathammina? sp.
Figure 14.13, 14.14
Remarks
Oblique sections of some coiled bilocular forms with a probable Quinqueloculina-like arrangement of the tubular chamber are reported as Agathammina? sp. (Fig. 14.13, 14.14) from the Upper Guandao section. Such forms range from the Ladinian (Fassanian) to the Carnian (Cordevolian) (Fig. 9).
Family Ophthalmidiidae Wiesner, Reference Wiesner1920
Genus Ophthalmidium Kübler and Zwingli, Reference Kübler and Zwingli1870
Ophthalmidium exiguum Koehn-Zaninetti, Reference Koehn-Zaninetti1969
Figure 14.15
Remarks
Identical to forms described by Koehn-Zaninetti (Reference Koehn-Zaninetti1969), Ophthalmidium exiguum is characterized by tests of small dimensions, thin wall, and chambers half-whorl long in the adult. It has been recorded from the Longobardian (Upper Ladinian) of the Upper Guandao section (Fig. 9). Typical sections of O. exiguum have been illustrated from the Ladinian to Carnian of Turkey (Altıner and Zaninetti, Reference Altıner and Zaninetti1981) and China (He and Wang, Reference He and Wang1990). Forms reported more recently from the Anisian of the southern Alps (Italy) by Faletti and Ivanova (Reference Faletti and Ivanova2003) and the Qingyan section of south China by Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015) do not belong to the population of O. exiguum. They should be attributed to the lineage of Eoophthalmidium tricki, described by Langer (Reference Langer1968; see also Zaninetti, Reference Zaninetti1976; Okay et al., Reference Okay, Altıner and Kiliç2015). The rest of the ophthalmidiid fauna consists of five different populations.
Ophthalmidium sp. 1
Figure 14.16–14.18
Remarks
Ophthalmidium sp. 1 is recognized in robust axial sections, tends to uncoil toward the end of its ontogeny, and ranges from Longobardian (Ladinian) to Cordevolian (Carnian) in the Upper Guandao section (Fig. 9).
Ophthalmidium sp. 2
Figure 14.19–14.21
Remarks
Ophthalmidium sp. 2, characterized by an elongate ellipsoidal test with narrow tubular chambers and a slightly sigmoidal coiling, also ranges from Longobardian to Cordevolian in the Upper Guandao section (Fig. 9).
Ophthalmidium sp. 3
Figure 14.22, 14.23
Remarks
Small and slightly biumbilicate tests of Ophthalmidium sp. 3 have been recorded from the Pelsonian to Cordevolian interval of the Upper Guandao section (Figs. 8, 9).
Ophthalmidium sp. 4
Figure 14.25
Remarks
Ophthalmidium sp. 4, with a losangic outline, is also found in the Cordevolian of the Upper Guandao section (Fig. 9).
Ophthalmidium? sp. 5
Figure 14.24
Remarks
Ophthalmidium? sp. 5 (Fig. 14.24) is quite close to Eoophthalmidium of Langer (Reference Langer1968), with a cornuspirine tubular second chamber in early whorls and gradually shortened chambers in the adult similar to the illustrations given in Okay et al. (Reference Okay, Altıner and Kiliç2015; see also Zaninetti and Brönnimann, Reference Zaninetti and Brönnimann1969). This form occurs in the Bithynian of the Guandao section (Fig. 11).
Family Quinqueloculinidae Cushman, Reference Cushman1917
Genus Gsollbergella Zaninetti, Reference Zaninetti1979
Gsollbergella? sp. 1
Figure 14.26
Remarks
Questionable forms assigned to Gsollbergella? sp. 1 (Fig. 14.26) and G.? sp. 2 (Fig. 14.27), characterized by incipient divisions in the tubular chamber, have been recognized in the Fassanian (lower Ladinian) and Pelsonian to Illyrian (Anisian), respectively, of the Upper Guandao section (Figs. 8, 9). However, transverse sections of such forms are not known, and only a quinqueloculine-type coiling would enable confident assignment to the genus Gsollbergella.
Gsollbergella? sp. 2
Figure 14.27
Remarks
See Remarks under Gsollbergella? sp. 1.
Family Galeanellidae Zaninetti et al., Reference Zaninetti, Altıner, Dağer and Ducret1982
Genus Galeanella Kristan, Reference Kristan1958
Galeanella sp.
Figure 14.28
Remarks
A primitive form of the genus Galeanella, G. sp., characterized by a thick perforated wall and probably with a biloculine chamber arrangement, has been recorded in the Cordevolian (lower Carnian) strata of the Upper Guandao section (Fig. 9). According to Zaninetti, Martini, and Altıner (Reference Zaninetti, Martini and Altıner1992) and Zaninetti and Martini (Reference Zaninetti and Martini1993), the earliest representatives of the genus Galeanella appeared early in the Carnian. This unusual Galeanella has also been illustrated by He and Wang (Reference He and Wang1990) from the Carnian of south China. The Chinese specimen could be considered as one of the earliest Galeanella populations.
Class Textulariata Mikhalevich, Reference Mikhalevich1980
Remarks
In the Great Bank of Guizhou, Textulariata is quite diverse and composed of the bilocular family Ammodiscidae and several multilocular families, namely Trochamminidae, Reophacidae, Spiroplectamminidae, Placopsilinidae, Verneulinidae, Cuneolinidae, Piallinidae, and Textulariidae.
Family Ammodiscidae Reuss, Reference Reuss1862
Subfamily Glomospirellinae Ciarapica and Zaninetti, Reference Ciarapica and Zaninetti1985
Genus Gandinella Ciarapica and Zaninetti, Reference Ciarapica and Zaninetti1985
Gandinella? sp.
Figure 15.12

Figure 15. All specimens are from Dawen (PDW), Dajiang (PDJ), Middle Triassic Dajiang (MDJ), Guandao (PGD), and Upper Guandao (PUG) sections. (1–5) Glomospirella vulgaris Ho, Reference Ho1959; (6, 7) Glomospirella sp. 1; (8, 9) Glomospirella sp. 2 (Glomospirella lampangensis? Kobayashi et al., Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006); (10, 11) Glomospira sp. (Pilammina praedensa? Urošević, Reference Urošević1988); (12) Gandinella? sp.; (13–20) Pilammina densa Pantić, Reference Pantić1965; (21–24) Pilammina densa? Pantić, Reference Pantić1965; (25–27) Pilamminella grandis Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967. (1, 2) PDJ-319; (3) PGD-131; (4) PDW-342; (5) PGD-096; (6, 19–22) PUG-029; (7) PUG-054; (8, 9) PGD-236; (10) PDJ-302; (11) PDJ-303; (12) MDJ-02; (13) PUG-063; (14) PUG-059; (15) PUG-054; (16) PUG-045; (17) PUG-037; (18) PGD-217; (23) PUG-015; (24) PUG-051; (25, 26) PUG-027; (27) PUG-033. Scale bars = 100 μm.
Remarks
In the subfamily Glomospirellinae, forms tending to display a sigmoidal coiling following a streptospiral early stage are reported as Gandinella? sp. Such forms have been discovered in the Pelsonian substage of the Middle Triassic Dajiang section (Fig. 10).
Genus Glomospira Rzehak, Reference Rzehak1885
Glomospira sp.
Figure 15.10, 15.11
Remarks
Glomospira sp., recorded from the Smithian of the Dajiang section (Fig. 6), is characterized by small, entirely streptospiral tests, nearly identical to forms illustrated as G. regularis by Ho (Reference Ho1959). Rettori (Reference Rettori1995) placed such forms in synonymy under Pilammina praedensa Urošević, Reference Urošević1988.
Genus Glomospirella Plummer, Reference Plummer1945
Glomospirella vulgaris Ho, Reference Ho1959
Figure 15.1–15.5
Remarks
The genus Glomospirella is represented by three different populations. Among these, G. vulgaris consists of a deuteroloculus describing an initial streptospiral coiling with 3–5 whorls and then a planispiral coiling tending to become slightly sigmoidal in the adult. It has been recorded in the Smithian of the Dawen (Fig. 5), Dajiang (Fig. 6), and Guandao (Fig. 11) sections. In the recent literature, Glomospirella illustrated by Song et al. (Reference Song, Tong and Chen2011b) from the Olenekian of the Yangtze Block is probably G. vulgaris. In addition, most of the forms illustrated as Hoyenella spp. by Song et al. (Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016) in the facies photographs of the Olenekian of south China should also be referred to G. vulgaris. From the European side, G. vulgaris described from the Anisian of Poland and Slovakia by Rychliński et al. (Reference Rychliński, Ivanova, Zaglarz and Bucur2013) should be referred either to Gandinella or Glomospirella.
Glomospirella sp. 1
Figure 15.6, 15.7
Remarks
Glomospirella sp. 1, recorded from the uppermost Bithynian to Longobardian interval of the Upper Guandao section (Figs. 8, 9), is characterized by a tightly coiled deuteroloculus with a smaller chamber height, similar to the morphology of Glomospira tenuifistula described by Ho (Reference Ho1959). However, this form has a Glomospirella-like planispiral stage and more compressed axial sections.
Glomospirella sp. 2
Figure 15.8, 15.9
Remarks
The other Glomospirella population, G. sp. 2, is also distinct, with its pronounced streptospiral stage occupying more than half of the volume of the test and tightly coiled, oscillating to planispiral two or three whorls. Glomospirella sp. 2 is morphologically very close to Glomospirella lampangensis Kobayashi et al., Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006, which was described from northern Thailand. However, more material is required in order to decide on the degree of similarity in coiling between our G. sp. 2 and G. lampangensis. This form appears in the Aegean close to the Olenekian-Anisian boundary in the Guandao section and ranges into the Bithynian (Fig. 11).
Subfamily Pilammininae Martini, Vachard, and Zaninetti, Reference Martini, Vachard and Zaninetti1995
Genus Pilammina Pantić, Reference Pantić1965
Pilammina densa Pantić, Reference Pantić1965
Figure 15.13–15.20
Remarks
In the ammodiscid foraminiferal fauna, the main biostratigraphic markers in the Triassic stratigraphy belong to genera such as Pilammina and Pilamminella. Pilammina densa Pantić, Reference Pantić1965 has been reported from the Anisian by many authors (e.g., Baud et al., Reference Baud, Zaninetti and Brönnimann1971; Premoli Silva, Reference Premoli Silva1971; Brönnimann et al., Reference Brönnimann, Cadet and Zaninetti1973a, Reference Brönnimann, Cadet and Zaninettib; Gazdzicki et al., Reference Gazdzicki, Trammer and Zwidzka1975; Zaninetti, Reference Zaninetti1976; Dağer, Reference Dağer1978a; Trifonova, Reference Trifonova1978a; Salaj et al., Reference Salaj, Borza and Samuel1983; He, Reference He1984; He and Wang, Reference He and Wang1990; Altıner and Koçyiğit, Reference Altıner and Koçyiğit1993; Rettori, Reference Rettori1995; Bucur et al., Reference Bucur, Strutinski and Paica1997; de Bono et al., Reference de Bono, Martini, Zaninetti, Hirsch, Stampfli and Vavassis2001; Fugagnoli and Posenato, Reference Fugagnoli and Posenato2004; Berra et al., Reference Berra, Rettori and Bassi2005; Kobayashi et al., Reference Kobayashi, Martini and Zaninetti2005, Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006; Marquez, Reference Marquez2005; Payne et al., Reference Payne, Summers, Rego, Altıner, Wei, Yu and Lehrmann2011; Velledits et al., Reference Velledits, Péró, Blau, Senowbari-Daryan, Kovács, Piros, Pocsai, Szúgyi-Simon, Dumitrica and Palfy2011; Song et al., Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015; Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015; Miyahigashi et al., Reference Miyahigashi, Hara, Hisada, Nakano, Charoentitirat, Charusiri, Khamphoveng, Martini and Ueno2017). It is mainly characterized by numerous streptospirally and tightly coiled whorls with planes of coiling changing slowly (Fig. 15.13–15.20).
In the Guandao and Upper Guandao sections (Figs. 8, 11), P. densa ranges from the Bithynian to the Pelsonian, as calibrated with conodonts (Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). It has been recorded in the Pelsonian of the Middle Triassic Dajiang section (Fig. 10). The first appearance of P. densa nearly coincides with the Aegean/Bithynian boundary, consistent with reporting by Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015). Although the P. densa Zone has been assigned to the late Anisian in western Tethyan domains (See Rettori, Reference Martini, Rettori, Urošević and Zaninetti1995 and the references therein), this form has never been recorded from the upper Anisian (Illyrian) in the Chinese sections.
Pilammina densa? Pantić, Reference Pantić1965
Figure 15.21–15.24
Remarks
Smaller forms with numerous whorls describing partly a sigmoidal coiling (Fig. 15.21, 15.22) and displaying a quadrangular profile (Fig. 15.23, 15.24) have been questionably assigned to P. densa. Similar forms were reported by Flügel et al. (Reference Flügel, Ramovš and Bucur1994) as Glomospira? sp. cf. G.? micas He and Yue, Reference He and Yue1987 from the Pelsonian of Slovenia. Pilammina densa? of this study is also close to some specimens illustrated as P. densa from the Anisian of Malaysia by Gazdzicki and Smit (Reference Gazdzicki and Smit1977) and Greece by Rettori et al. (Reference Rettori, Angiolini and Muttoni1994).
Genus Pilamminella Salaj, Reference Salaj1978
Pilamminella grandis Salaj in Salaj et al., Reference Salaj, Biely and Bistricky1967
Figure 15.25–15.27
Remarks
The genus Pilamminella is represented by P. grandis in the Chinese material. This form is characterized by tests consisting of oscillating to planispiral whorls in the adult following a tightly coiled Pilammina-like stage. Rettori (Reference Rettori1995) considered this form as a synonym of P. semiplana; however, P. grandis differs from this form in the absence of sigmoidal coiling in the initial stage and rather less tightly coiled whorls in the planispiral stage. Some sections of foraminifera illustrated as Glomospira densa or Glomospira sp. by Kobayashi (Reference Kobayashi1996) from the Anisian of the Kanto Mountains, Japan, belong to P. grandis. Similarly, P. grandis has been reported from Japan as Glomospirella irregularis in Kobayashi et al. (Reference Kobayashi, Martini and Zaninetti2005). Bucur et al. (Reference Bucur, Strutinski and Paica1997) extensively illustrated P. grandis from the Anisian of the southern Carpathians in Romania. In relatively recent literature, P. grandis has been reported as P. semiplana, following Rettori (Reference Rettori1995), from the Anisian of northern Italy by Fugagnoli and Posenato (Reference Fugagnoli and Posenato2004) and Berra et al. (Reference Berra, Rettori and Bassi2005). Pilamminella grandis ranges from Aegean to Pelsonian in the Upper Guandao section (Fig. 8) and Aegean to Bithynian in the Guandao section (Fig. 11).
Subfamily Tolypammininae Cushman, Reference Cushman1928
Genus Tolypammina Rhumbler, Reference Rhumbler1895
Tolypammina gregaria Wendt, Reference Wendt1969
Figure 16.1

Figure 16. All specimens are from Middle Triassic Dajiang (MDJ), Guandao (PGD), and Upper Guandao (PUG) sections. (1) Tolypammina gregaria Wendt, Reference Wendt1969; (2, 3) Reophax sp. 1; (4) Reophax sp. 2; (5, 6) Reophax sp. 3; (7, 8) Reophax sp. 4; (9, 10) Reophax sp. 5; (11) Reophax sp. 6; (12) Reophax sp. 7; (13, 14) Malayspirina sp.; (15–17) Placopsilina sp.; (18–23) Trochammina almtalensis Koehn-Zaninetti, Reference Koehn-Zaninetti1968; (24, 25) Trochammina? sp. 1; (26) Trochammina? sp. 2; (27) Trochammina? sp. 3; (28) Piallina bronnimanni Martini, Rettori et al., Reference Martini, Vachard and Zaninetti1995; (29) Piallina? sp.; (30–32) Verneuilinoides? azzouzi (Salaj, Reference Salaj1978); (33) Textularia sp.1; (34, 35) Gaudryina sp. (Gaudryina triadica? Kristan-Tollmann, Reference Kristan-Tollmann1964); (36–39) Palaeolituonella reclinata He in He and Cai (Reference He and Cai1991); (40) Palaeolituonella sp. [Palaeolituonella meridionalis? (Luperto, Reference Luperto1965)]. (1) PUG-131; (2, 23) PUG-059; (3, 28) PUG-145; (4, 9, 18) PUG-081; (5, 32) PUG-077; (6, 33) PUG-143; (7) PUG-037; (8, 37) PUG-093; (10) PUG-137; (11, 20, 34, 39) PUG-054; (12) PUG-045; (13, 14) PUG-101; (15) PGD-126; (16) PGD-228; (17) PGD-210; (19) PUG-091; (21) MDJ-96; (22) PUG-097; (24) PGD-168; (25) PGD-167; (26) PUG-113; (27) PUG-099; (29) PUG-087; (30) MDJ-65; (31) PGD-181; (35) PUG-067; (36) PUG-123; (38) PUG-031; (40) PUG-093. Scale bars = 100 μm.
Remarks
Attached tests composed of an elongate, undivided, tubular chamber winding irregularly over the attachment surface are recognized as Tolypammina gregaria Wendt, Reference Wendt1969 in this study. This form has been reported from the Anisian to Carnian interval from several localities in the Triassic of Europe (Zaninetti, Reference Zaninetti1976). The specimens described and illustrated by Brönnimann and Zaninetti (Reference Brönnimann and Zaninetti1972) from the Muschelkalk of southern France as Tolypammina? aff. gregaria (Brönnimann and Zaninetti, Reference Brönnimann and Zaninetti1972, pl. 9, Figs. 2, 5) are nearly identical to forms discovered from the Great Bank of Guizhou.
Family Reophacidae Cushman, Reference Cushman1927
Genus Reophax de Montfort, Reference de Montfort1808
Reophax sp. 1
Figure 16.2, 16.3
Remarks
In the studied Chinese stratigraphic sections, rectilinear uniserial tests of the genus Reophax, with a coarsely agglutinated wall and single, terminal and rounded aperture, appear at the base of the Anisian Stage. Seven different populations, reported with open nomenclature, have been recognized in the studied material. Reophax sp. 1, relatively smaller in size among the Chinese Reophax populations, consists of three or four chambers gradually increasing in width and height (Fig. 16.2, 16.3). In the Upper Guandao section it ranges from Pelsonian to Cordevolian (Carnian) (Figs. 8, 9).
Reophax sp. 2
Figure 16.4
Remarks
Reophax sp. 2, again a small form, is characterized by a very narrow and tubular lumen in the two early chambers and the last chamber increasing rapidly in height (Fig. 16.4). It occurs from the Aegean to the Bithynian in the Guandao section (Fig. 11) and from the Aegean to the Illyrian in the Upper Guandao section (Fig. 8).
Reophax sp. 3
Figure 16.5, 16.6
Remarks
Reophax sp. 3 consists of comparatively wider chambers than Reophax sp. 1 (Fig. 16.5, 16.6) and is present in the Bithynian of the Guandao section (Fig. 11) and the Bithynian to Cordevolian (Carnian) of the Upper Guandao section (Figs. 8, 9).
Reophax sp. 4
Figure 16.7, 16.8
Remarks
Reophax sp. 4 is distinguished by a last chamber rapidly increasing in height (Fig. 16.7, 16.8). It occurs from the Pelsonian to the Illyrian in the Upper Guandao section (Fig. 8).
Reophax sp. 5
Figure 16.9, 16.10
Remarks
Reophax sp. 5, a larger form, consists of a large proloculus and six succeeding chambers increasing very slowly in height and width (Fig. 16.9, 16.10). It occurs in Anisian to Carnian strata of the Guandao and Upper Guandao sections (Figs. 8, 9, 11). Flügel et al. (Reference Flügel, Ramovš and Bucur1994) reported this form as Earlandinita elongata Salaj et al., Reference Salaj, Biely and Bistricky1967 from the Pelsonian of Slovenia.
Reophax sp. 6
Figure 16.11
Remarks
Reophax sp. 6, again a larger sized reophacid, is composed of three pyriform chambers rapidly growing in height with depressed sutures (Fig. 16.11). It has been recorded in the Pelsonian to Longobardian (Ladinian) interval of the Upper Guandao section (Figs. 8, 9).
Reophax sp. 7
Figure 16.12
Remarks
Reophax sp. 7, a long and arcuate reophacid, consists of eight chambers increasing slowly in size and becoming pyriform toward the end of ontogeny (Fig. 16.12). It has been recorded in the Pelsonian of the Upper Guandao section (Fig. 8).
Family Spiroplectamminidae Cushman, Reference Cushman1927
Subfamily Spiroplectammininae Cushman, Reference Cushman1927
Genus Malayspirina Vachard in Fontaine, Khoo, and Vachard, Reference Fontaine, Khoo and Vachard1988
Malayspirina sp.
Figure 16.13, 16.14
Remarks
The genus Malayspirina was placed in the Carboniferous family Forschiidae Dain in Dain and Grozdilova (Reference Dain and Grozdilova1953) when it was created by Vachard (in Fontaine et al., Reference Fontaine, Khoo and Vachard1988). Despite the morphological similarity with Spiroplectammina, Malayspirina differs from it by the irregularity of the coiled and uncoiled parts of the test (Fontaine et al., Reference Fontaine, Khoo and Vachard1988) and the highly reduced coiled stage. Forms illustrated from the Kanto Mountains, Japan, by Kobayashi (Reference Kobayashi1996) probably are not Malayspirina. Rather, they are sections of the uncoiled part of Endotebanella. In Malayspirina specimens of the Great Bank of Guizhou (Fig. 16.13, 16.14), the reduced coiled stage probably lies in a plane oblique to the axis of the biserial arrangement of chambers characterized by a highly thickened wall. Malayspirina sp. is found in the Illyrian (upper Anisian) to Fassanian (lower Ladinian) interval of the Upper Guandao section (Figs. 8, 9) and this range is definitely older than the upper Ladinian–Carnian range assigned to the type species, Malayspirina fontainei Vachard in Fontaine et al., Reference Fontaine, Khoo and Vachard1988.
Family Placopsilinidae Rhumbler, Reference Rhumbler1913
Genus Placopsilina d'Orbigny, Reference d'Orbigny1850
Placopsilina sp.
Figure 16.15–16.17
Remarks
Attached to a substratum, coarsely agglutinated Placopsilina sp. specimens from China are characterized by a highly reduced coiled stage, 6–8 chambers in the uncoiled stage, and a single terminal aperture. In the uncoiled stage, the axis characteristically deviates 20–25° from the surface of attachment and chambers progressively grow in height. Placopsilina sp. differs from P.? florae and P.? lacera of Trifonova (Reference Trifonova1967) and P.? hyerensis of Brönnimann and Zaninetti (Reference Brönnimann and Zaninetti1972) by a much smaller coiled stage and a slender test containing more chambers in the uncoiled stage. Placopsilina sp. has been found in the Aegean–Bithynian of the Guandao section (Fig. 11).
Family Trochamminidae Schwager, Reference Schwager1877
Genus Trochammina Parker and Jones, Reference Parker and Jones1859
Trochammina almtalensis Koehn-Zaninetti, Reference Koehn-Zaninetti1968
Figure 16.18–16.23
Remarks
Koehn-Zaninetti (Reference Koehn-Zaninetti1968) published a condensed version of Koehn-Zaninetti (Reference Koehn-Zaninetti1969) with the same title in which she reported the new taxa that she discovered in this paper. Although these taxa were also described as new in 1969 (her main work) we give priority to the 1968 publication. Trochammina almtalensis Koehn-Zaninetti, Reference Koehn-Zaninetti1968 specimens of the Great Bank of Guizhou from south China are characterized by the small size and trochospirally coiled three or four whorls consisting of four chambers per whorl (Fig. 16.18–16.23). They are nearly identical to forms illustrated in the original description by Koehn-Zaninetti (Reference Koehn-Zaninetti1969). Typical specimens of T. almtalensis were illustrated from the Anisian of China by He and Wang (Reference He and Wang1990) and He and Cai (Reference He and Cai1991). Recently, Haig et al. (Reference Haig, McCartain, Barber and Backhouse2007) considered the almtalensis population to be close to the genus Siphovalvulina Septfontaine, Reference Septfontaine1988. We disagree with this suggestion because T. almtalensis does not possess a central siphonal canal, as is characteristic of the type species of Siphovalvulina. According to Tian et al. (Reference Tian, Tong, Algeo, Song, Song, Chu, Shi and Bottjer2014), T. almtalensis first appears in the Smithian of the Bianyang section of the Nanpanjiang Basin, south China. This report, not based on illustrated specimens, is probably an erroneous identification.
Trochammina? sp. 1
Figure 16.24, 16.25
Remarks
In the absence of transverse sections, three other populations have been questionably assigned to the genus Trochammina. The small tests of T.? sp. 1 (Fig. 16.24, 16.25) are characteristically found in the Spathian of the Guandao section (Fig. 11).
Trochammina? sp. 2
Figure 16.26
Remarks
Populations of Trochammina? sp. 2, characterized by very high trochospiral tests, are found in the Ladinian of the Upper Guandao section (Fig. 9).
Trochammina? sp. 3
Figure 16.27
Remarks
Populations of Trochammina? sp. 3, characterized by very low trochospiral tests, are found in the Ladinian of the Upper Guandao section (Fig. 9).
Family Verneuilinidae Cushman, Reference Cushman1911
Subfamily Verneuilinoidinae Suleymanov, Reference Suleymanov1973
Genus Verneuilinoides Loeblich and Tappan, Reference Loeblich and Tappan1949
Verneuilinoides? azzouzi (Salaj, Reference Salaj1978)
Figure 16.30–16.32
Remarks
Following the original description from the Anisian to Carnian of the western Carpathians by Salaj (Reference Salaj1978), Verneuilinoides? azzouzi has been reported several times as a species belonging to the genus Valvulina (Salaj et al., Reference Salaj, Borza and Samuel1983; Michalík et al., Reference Michalík, Masaryk, Lintnerová, Soták, Jendrejáková, Papšová and Buček1993; Flügel et al., Reference Flügel, Ramovš and Bucur1994; Korchagin, Reference Korchagin2008; Ueno et al., Reference Ueno, Miyahigashi, Kamata, Kato, Charoentitirat and Limruk2012). However, this triserial form (Fig. 16.30–16.32) does not possess the canaliculate wall structure and valvular tooth covering the aperture, as given in the type description of the genus Valvulina. Following He and Wang (Reference He and Wang1990), we place this population questionably under the genus Verneuilinoides because the aperture is not clearly visible in our material. Verneuilinoides? azzouzi has been misidentified as Gaudryina triadica Kristan-Tollmann, Reference Kristan-Tollmann1964 from the Qingyan section of south China (Song et al., Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015; Figs. 5, 9). In the GBG material, this species is present in the Fassanian to Longobardian (Ladinian) of the Middle Triassic Dajiang section and in the Spathian and Illyrian of the Guandao and Upper Guandao sections (Figs. 8, 10, 11).
Subfamily Verneuilininae Cushman, Reference Cushman1911
Genus Gaudryina d'Orbigny, Reference d'Orbigny1839
Gaudryina sp.
Figure 16.34, 16.35
Remarks
Gaudryina sp., with a reduced triserial stage and a following biserial stage consisting of three or four rows, is found in the Pelsonian to Illyrian interval of the Upper Guandao section and in the Aegean of the Guandao section (Figs. 8, 11). Chinese specimens closely resemble Gaudryina triadica Kristan-Tollmann, Reference Kristan-Tollmann1964 from the Rhaetian Zlambach Marls of Austria and G. triadica specimens illustrated from the Anisian of Japan (Kobayashi et al., Reference Kobayashi, Martini and Zaninetti2005) and Thailand (Kobayashi et al., Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006). It differs from two other Triassic species, G. racema Trifonova, Reference Trifonova1961 and G. triassica Trifonova, Reference Trifonova1961, in having less globular chambers and a wider frontal profile of the test, respectively.
Family Piallinidae Rettori and Zaninetti in Rettori et al., Reference Rettori, Zaninetti, Martini and Vachard1993
Genus Piallina Rettori and Zaninetti in Rettori et al., Reference Rettori, Zaninetti, Martini and Vachard1993
Piallina bronnimanni Martini, Rettori et al., Reference Martini, Vachard and Zaninetti1995
Figure 16.28
Remarks
Two specimens from the Chinese material have been assigned to the family Piallinidae (Rettori and Zaninetti in Rettori et al., Reference Rettori, Zaninetti, Martini and Vachard1993; Martini, Rettori et al., Reference Martini, Vachard and Zaninetti1995). Piallina bronnimanni Martini, Rettori et al., Reference Martini, Vachard and Zaninetti1995 (Fig. 16.28) is small, probably starting with a low trochospiral initial portion followed by a triserial stage composed of globular chambers. Very similar specimens of P. bronnimanni have been described by Rettori et al. (Reference Rettori, Loriga and Neri1998) from the lower Carnian of northeastern Italy. Both Piallina bronnimanni and Piallina? sp. occur in the Upper Guandao section (Figs. 8, 9). Piallina bronnimanni is from the Cordevolian (Carnian).
Piallina? sp.
Figure 16.29
Remarks
The other form, assigned questionably to Piallina? sp., is again low trochospiral in the early portion; however, the chamber organization in the later stage of the ontogeny is not clearly visible. Both Piallina bronnimanni and Piallina? sp. occur in the Upper Guandao section (Figs. 8, 9). Piallina? sp. is older than Piallina bronnimanni and found in the Illyrian (Anisian).
Family Cuneolinidae Saidova, Reference Saidova1981
Subfamily Cuneolininae Saidova, Reference Saidova1981
Genus Palaeolituonella Bérczi-Makk, Reference Bérczi-Makk1981
Palaeolituonella reclinata He in He and Cai, Reference He and Cai1991
Figure 16.36–16.39
Remarks
Two distinct populations of Palaeolituonella are present in the Chinese material. Palaeolituonella reclinata He in He and Cai, Reference He and Cai1991 (Fig. 16.36–16.39) differs from the type species, P. meridionalis (Luperto, Reference Luperto1965), by its smaller and elongate bell-shaped test and thinner wall, and should not be synonymized under P. meridionalis, as was considered by Rettori (Reference Rettori1995). Although poorly described, the other Chinese species, P. minima He and Wang, Reference He and Wang1990, is a form close to P. reclinata, but differs from it by a less-pronounced uniserial stage and rapidly enlarging chambers in the initial stage. Haplophragmium sp., illustrated from the Lower Triassic to middle Anisian Karatash Group of Pamirs by Korchagin (Reference Korchagin2008), is probably an oblique section of P. reclinata. Gale et al. (Reference Gale, Peybernes, Celarc, Hoćevar, Šelih and Martini2018) reported P. reclinata under the name of P. meridionalis from the lower Carnian of Slovenia.
Palaeolituonella reclinata is present in the Upper Guandao section from the uppermost Aegean (Anisian) to the Longobardian (Ladinian) (Figs. 8, 9). It has been also recorded from the Pelsonian of the Middle Triassic Dajiang section (Fig. 10).
Palaeolituonella sp.
Figure 16.40
Remarks
Our second population in the Chinese material is characterized by a larger test and thicker wall. Despite the similarity to P. meridionalis, we classify such forms Palaeolituonella sp. (Fig. 16.40) in the absence of well-oriented sections. Palaeolituonella sp. has been found in the upper Illyrian (Anisian) of the Upper Guandao section (Fig. 8).
Family Textulariidae Ehrenberg, Reference Ehrenberg1838
Genus Textularia de Blainville, Reference de Blainville1824
Textularia sp.
Figure 16.33
Remarks
Very rare, small, entirely biserial tests discovered in the Cordevolian (Carnian) of the Upper Guandao section (Fig. 9) have been classified as Textularia sp. (Fig. 16.33) in the Chinese material.
Class Fusulinata Gaillot and Vachard, Reference Gaillot and Vachard2007
Remarks
The Class Fusulinata comprises four families in the material of the Great Bank of Guizhou. These are Earlandiidae, Pseudoammodiscidae, Endotebidae, and Endotriadidae.
Family Earlandiidae Cummings, Reference Cummings1955
Remarks
See Remarks under Class Fusulinata Gaillot and Vachard, Reference Gaillot and Vachard2007.
Genus Earlandia Plummer, Reference Plummer1930
Remarks
The genus Earlandia, well known from the Carboniferous and characterized by rectilinear, bilocular tests with microgranular walls, is also present in the uppermost Permian (Changhsingian) deposits of the Southern Biofacies Belt in Turkey (Altıner et al., Reference Altıner, Özkan-Altıner, Koçyiğit, Bozkurt, Winchester and Piper2000), as well as in several localities of the Middle East (Gaillot and Vachard, Reference Gaillot and Vachard2007). It is a Permian-Triassic boundary survivor (Groves and Altıner, Reference Groves and Altıner2005; Groves et al., Reference Groves, Altıner and Rettori2005, Reference Groves, Rettori, Payne, Boyce and Altıner2007; Vachard et al., Reference Vachard, Pille and Gaillot2010; Krainer and Vachard, Reference Krainer and Vachard2011), also appearing in the lowermost Triassic deposits of the Great Bank of Guizhou. Recently, Nestell et al. (Reference Nestell, Nestell, Ellwood, Wardlaw, Basu, Ghosh, Lan, Rowe, Hunt, Tomkin and Ratcliffe2015) opposed the use of Earlandia in the Early Triassic and suggested classifying all Earlandia-like forms within the agglutinated genus Hyperammina. The walls of Earlandia specimens in our Chinese material are composed of microgranular calcite and are directly comparable with those of the other Earlandia populations from the Paleozoic (e.g., Earlandia gr. E. elegans, E. gr. E. moderata, E. gr. E. minor, etc.). We consider the specimens examined by Nestell et al. (Reference Nestell, Nestell, Ellwood, Wardlaw, Basu, Ghosh, Lan, Rowe, Hunt, Tomkin and Ratcliffe2015) to be most probably diagenetically altered or totally replaced forms. Thus, we argue that the use of the genus Hyperammina is inadequate for the Early Triassic specimens.
The genus Earlandia consists of three distinct populations in the Chinese foraminiferal fauna: Earlandia tintinniformis (Misik, Reference Misik1971), E. sp. 1, and E. sp. 2.
Earlandia tintinniformis (Misik, Reference Misik1971)
Figure 17.5–17.7

Figure 17. All specimens are from Dawen (PDW), Dajiang (PDJ), Guandao (PGD), and Upper Guandao (PUG) sections. (1, 2) Earlandia sp. 1; (3, 4) Earlandia sp. 2; (5–7) Earlandia tintinniformis (Misik, Reference Misik1971); (8–14) Pseudoammodiscus sp.; (15–19) Endoteba controversa Vachard and Razgallah, Reference Vachard and Razgallah1988; (20–23) Endoteba bithynica Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994; (24–27) Endoteba obturata (Brönnimann and Zaninetti, Reference Brönnimann and Zaninetti1972); (28, 29) Endoteba badouxi (Zaninetti and Brönnimann in Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972c). (1, 2) PDJ-055; (3, 4) PDJ-172; (5) PUG-091; (6, 18) PUG-054; (7) PUG-045; (8–10, 12, 14) PDJ-319; (11) PDW-344; (13) PDG-111; (15) PUG-037; (16) PUG-075; (17, 20) PGD-219; (19) PUG-069; (21) PGD-207; (22) PGD-220; (23) PGD-236; (24) PUG-085; (25) PUG-063; (26) PUG-029; (27) PUG-109; (28) PUG-123; (29) PUG-107. Scale bars = 100 μm.
Remarks
Earlandia tintinniformis (Fig. 17.5–17.7), a conical tubular form nearly identical to specimens illustrated in Zaninetti et al. (Reference Zaninetti, Brönnimann and Baud1972c), differs from Earlandia sp. 1 and E. sp. 2 by the progressive increase of the diameter of the deuteroloculus during ontogeny. This form is found only in the Anisian Stage of the GBG. It is rare in the Aegean of the Guandao section (Fig. 11), but more common in the Bithynian to Illyrian deposits of the Upper Guandao section (Fig. 8). The specimen illustrated from the Aegean of the Qingyan section from south China by Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015) is not E. tintinniformis. This form should be classified as an undeterminable Earlandia with a thick wall.
Earlandia sp. 1
Figure 17.1, 17.2
Remarks
Earlandia sp. 1, characterized by a much smaller test (Fig. 17.1, 17.2) than Earlandia sp. 2 (Fig. 17.3, 17.4), is found only in the lowermost Griesbachian beds of the Dawen and Dajiang sections (Figs. 5, 6). Earlandia tintinniformis differs from Earlandia sp. 1 by the progressive increase of the diameter of the deuteroloculus during ontogeny. Nestell et al. (Reference Nestell, Kolar-Jurkovšek, Jurkovšek and Aljinović2011) illustrated this form as ‘E.’ gracilis from the uppermost Permian of western Slovenia. Earlandia sp. described by Okuyucu et al. (Reference Okuyucu, Ivanova, Bedi and Ergen2014) from the Induan of northwestern Turkey is also referable to our E. sp. 1.
Earlandia sp. 2
Figure 17.3, 17.4
Remarks
Earlandia sp. 2 has a longer stratigraphic range, Griesbachian in the Dawen section (Fig. 5) and Griesbachian–lower Dienerian in the Dajiang section (Fig. 6). It has been reported as Earlandia sp. from the Griesbachian of China (Song et al., Reference Song, Tong, Chen, Yang and Wang2009, Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016), Slovenia (Nestell et al., Reference Nestell, Kolar-Jurkovšek, Jurkovšek and Aljinović2011), and Turkey (Ünal et al., Reference Ünal, Altıner, Yilmaz and Özkan-Altıner2003). Angiolini et al. (Reference Angiolini, Carabelli, Nicora, Crasquin-Soleau, Marcoux and Rettori2007) reported this form as E. amplimuralis from the Antalya Nappes, Turkey, whereas Krainer and Vachard (Reference Krainer and Vachard2011) preferred to name it as E. dunningtoni in the material of Austria. In a more recent study, Kolar-Jurkovšek et al. (Reference Kolar-Jurkovšek, Jurkovšek, Nestell and Aljinović2018) classified E. sp. 2 as Hyperammina deformis Bérczi-Makk, Reference Bérczi-Makk1987 from the base of the Triassic, described as Permian-Triassic Boundary (PTB) transitional beds in Slovenia. Earlandia tintinniformis differs from Earlandia sp. 2 by the progressive increase of the diameter of the deuteroloculus during ontogeny.
Family Pseudoammodiscidae Conil and Lys in Conil and Pirlet, Reference Conil and Pirlet1970
Remarks
In his latest analysis on the macroevolutionary patterns of Paleozoic foraminifera, Vachard (Reference Vachard2016, Reference Vachard, Lucas and Shen2018) limited the stratigraphic range of Pseudoammodiscidae to the Serpukhovian Stage. However, as indicated by Altıner and Payne (Reference Altıner and Payne2017), the pseudoammodiscid clade appears to have survived through the Permian as a sporadically occurring, ‘hidden’ taxon among the rapidly evolving fusulinoideans and other members of Fusulinata, Nodosariata, and Miliolata. Pseudoammodiscidae apparently survived the Permian-Triassic boundary crisis and appeared as a Lazarus taxon in the late Dienerian both in China and Turkey.
Genus Pseudoammodiscus Conil and Lys in Conil and Pirlet, Reference Conil and Pirlet1970
Pseudoammodiscus sp.
Figures 17.8–17.14, 19.19
Remarks
Specimens of Pseudoammodiscus sp. (Figs. 17.8–17.14, 19.19; Fig. 17.10 has been re-illustrated on Fig. 19.19 for comparison with Praetriadodiscus) discovered in the Lower Triassic deposits of the Great Bank of Guizhou from the Nanpanjiang Basin were previously reported and partly illustrated as an undetermined pseudoammodiscid ancestor by Altıner and Payne (Reference Altıner and Payne2017). These authors considered the specimens that they illustrated to be identical to forms reported as Ammodiscus parapriscus Ho, Reference Ho1959 from Turkey by Altıner and Zaninetti (Reference Altıner and Zaninetti1981). They stated that the true parapriscus population, as described by Ho (Reference Ho1959), is a taxon related to the evolution of porcelaneous hoyenellid foraminifera, and the specimens discovered both in China and Turkey belong to an undetermined pseudoammodiscid population, reported now as Pseudoammodiscus sp. in this study. Pseudoammodiscus sp. is present from the Dienerian to Smithian in the Dawen section (Fig. 5). It has been recorded in the Smithian of both Dajiang and Guandao sections (Figs. 6, 11).
Family Endotebidae Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994
Remarks
The family Endotebidae is diverse and common in the upper Spathian and Middle Triassic deposits of the Great Bank of Guizhou. It is represented by three genera: Endoteba, Endotebanella, and Spinoendotebanella n. gen. The description of the new genus Spinoendotebanella and its type species, S. lehrmanni n. gen., n. sp., is given below.
Genus Endoteba Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994
Endoteba controversa Vachard and Razgallah, Reference Vachard and Razgallah1988
Figure 17.15–17.19
Remarks
The type of the family, Endoteba, comprises five distinct populations. Endoteba controversa, the type species of the genus, is characterized by five chambers in the last whorl, a wall progressively increasing in thickness, and curved and cuneiform septa. The specimens illustrated as Endoteba ex gr. controversa by Chablais (Reference Chablais2010) from the Upper Triassic atoll-like carbonates of Japan probably are not correct because these forms do not possess cuneiform septa and are characterized by a thinner wall. The holotype of E. controversa has been described from the Permian of Tunisia (Vachard and Razgallah, Reference Vachard and Razgallah1988), and the species survived the Permian-Triassic boundary. According to Rettori (Reference Rettori1995; see also Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994), the stratigraphic range of this form extends from the base of the Triassic through the top of the Rhaetian; however, this range has not been confirmed by solid stratigraphic data. Endoteba controversa has been recorded from the Bithynian of the Guandao section (Fig. 11) and the Bithynian to lower Longobardian (Ladinian) of the Upper Guandao section (Figs. 8, 9).
Endoteba bithynica Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994
Figure 17.20–17.23
Remarks
Characterized by an irregular coiling, thick wall, and probably a longer spire, as indicated in the diagnosis by Vachard et al. (Reference Vachard, Martini, Rettori and Zaninetti1994), E. bithynica has a stratigraphic range from the upper Spathian to Bithynian in the Guandao section (Fig. 11). Recently, specimens from northern Thailand that are similar to the GBG material were illustrated in Kobayashi et al. (Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006).
Endoteba obturata (Brönnimann and Zaninetti, Reference Brönnimann and Zaninetti1972)
Figure 17.24–17.26
Remarks
Specimens belonging to Endoteba obturata, with its nautiloid profile, 4–5 chambers in the last whorl, and a thick wall, are nearly identical to forms illustrated in Brönnimann and Zaninetti (Reference Brönnimann and Zaninetti1972), Brönnimann et al. (Reference Brönnimann, Cadet and Zaninetti1973a, Reference Brönnimann, Cadet and Zaninettib), Vachard et al. (Reference Vachard, Martini, Rettori and Zaninetti1994), and Kobayashi et al. (Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006). Haig et al. (Reference Haig, McCartain, Barber and Backhouse2007) reported this form from the Carnian of East Timor. Endoteba obturata is present from the uppermost Bithynian to Illyrian in the Upper Guandao section (Fig. 8) and the Illyrian to Longobardian in the Middle Triassic Dajiang section (Fig. 10).
Endoteba badouxi (Zaninetti and Brönnimann in Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972c) Figure 17.28, 17.29
Remarks
Larger Endoteba specimens with seven chambers in the last whorl are similar to forms described in Zaninetti et al. (Reference Zaninetti, Brönnimann and Baud1972c) from Switzerland, and reported in this study as Endoteba badouxi. The stratigraphic range of E. badouxi is reported as Anisian to Carnian in Rettori (Reference Rettori1995). In the Chinese material, this form has been recorded from the Illyrian to the Longobardian in the Upper Guandao section (Figs. 8, 9). Endoteba badouxi was reported from the Middle Triassic of China (He and Wang, Reference He and Wang1990), Illyrian–Ladinian of Italy (Emmerich et al., Reference Emmerich, Zamparelli, Bechstädt and Zühlke2005), and Pelsonian–Carnian of Hungary (Velledits et al., Reference Velledits, Péró, Blau, Senowbari-Daryan, Kovács, Piros, Pocsai, Szúgyi-Simon, Dumitrica and Palfy2011). The specimen illustrated from Pamirs by Korchagin (Reference Korchagin2008) is characterized by fewer chambers and is probably not E. badouxi.
Endoteba sp.
Figure 18.1, 18.2
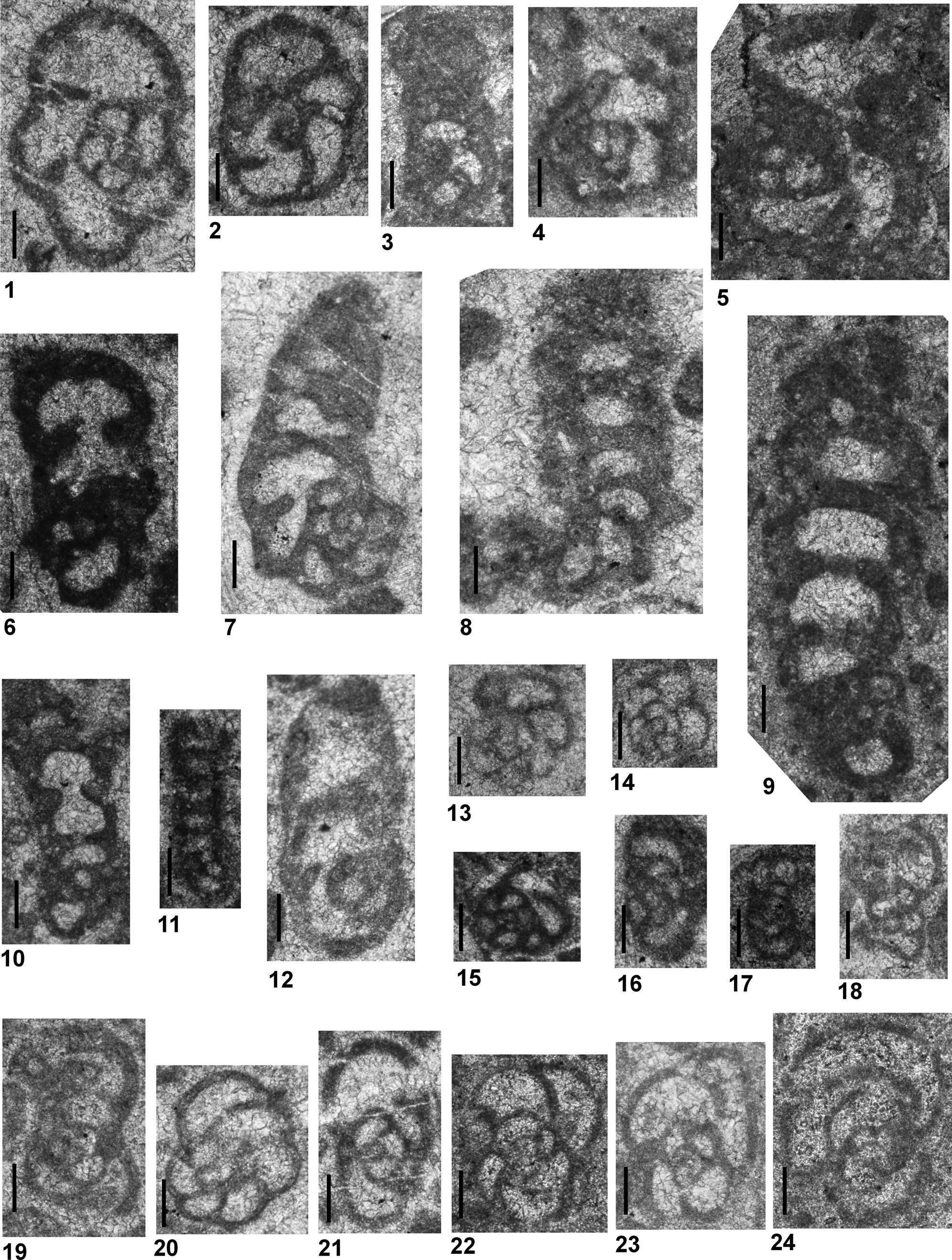
Figure 18. All specimens are from Middle Triassic Dajiang (MDJ), Guandao (PGD), and Upper Guandao (PUG) sections. (1, 2) Endoteba sp.; (3–6) Endotebanella kocaeliensis (Dağer, Reference Dağer1978b); (7–10) Spinoendotebanella lehrmanni n. gen., n. sp.; (11) Endotebanella sp. 2; (12) Endotebanella sp. 1; (13–17) Endotriada tyrrhenica Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994; (18, 19) Endotriada sp. 3; (20, 21) Endotriada sp. 1; (22–24) Endotriada sp. 2. (1, 2) PUG-081; (3) PUG-101; (4) PUG-009; (5) PUG-087; (6) PGD-217; (7) PUG-027; (8, 20) PUG-029; (9) PUG-012; (10) MDJ-39; (11) PGD-238; (12) PUG-037; (13) PUG-043; (14) PUG-054; (15) PUG-071; (16) PUG-063; (17) PUG-089; (18, 19, 23) PUG-077; (21) PUG-031; (22, 24) PUG-099. Scale bars = 100 μm.
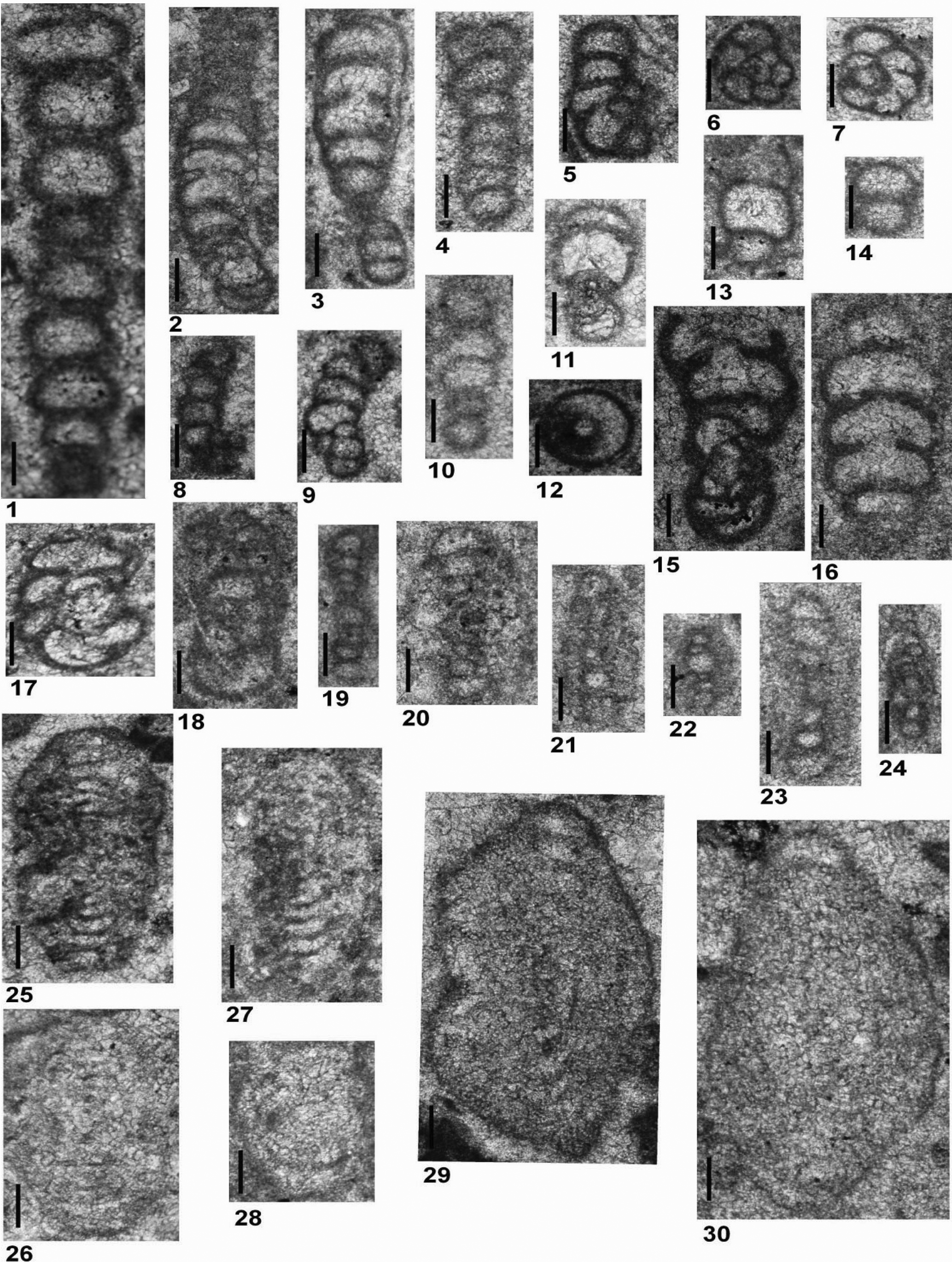
Figure 19. All specimens are from Dajiang (PDJ), Middle Triassic Dajiang (MDJ), Guandao (PGD), and Upper Guandao (PUG) sections. (1–14) Endotriadella wirzi (Koehn-Zaninetti, Reference Koehn-Zaninetti1968); (15, 16) Endotriadella wirzi? (Koehn-Zaninetti, Reference Koehn-Zaninetti1968); (17, 18) Endotriadella lombardi (Zaninetti and Brönnimann in Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972c); (19) Pseudoammodiscus sp.; (20, 22) Praetriadodiscus zaninettiae Altıner and Payne, Reference Altıner and Payne2017; (21, 23) Praetriadodiscus tappanae Altıner and Payne, Reference Altıner and Payne2017; (24) Praetriadodiscus ? sp.; (25–27) Triadodiscus sp.; (28) Parvalamella sp.; (29, 30) Aulotortus? eotriasicus Zaninetti, Rettori, and Martini, Reference Zaninetti, Rettori and Martini1994. (1, 15) PUG-043; (2) MDJ-09; (3) PUG-016; (4) PUG-037; (5) PUG-045; (6) PUG-067; (7, 17) PUG-029; (8) MDJ-39; (9) PUG-017; (10) PUG-054; (11) PUG-014; (12) PUG-089; (13) PUG-091; (14, 28) MDJ-57; (16) MDJ-15; (18) PUG-083; (19) PDJ-319; (20–24) PGD-131; (25–27) PUG-023; (29) PUG-045; (30) PUG-023. Scale bars = 100 μm.
Remarks
Specimens with 4.5–5 globular chambers in the last whorl and a thin wall, from the Pelsonian–Illyrian in the Upper Guandao section (Fig. 8), are considered to be a distinct Endoteba population and named as Endoteba sp. (Fig. 18.1, 18.2).
Genus Endotebanella Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994
Remarks
The genus Endotebanella is composed of three morphologically distinct populations: Endotebanella kocaeliensis, E. sp. 1, and E. sp. 2.
Endotebanella kocaeliensis (Dağer, Reference Dağer1978b)
Figure 18.3–18.6
Remarks
Endotebanella kocaeliensis consists of four or five chambers progressively growing in volume and a short, uncoiled stage with two or three chambers (Fig. 18.3–18.6). Chinese specimens are close to the holotype described by Dağer (Reference Dağer1978a, b) and its emended definition given by Vachard et al. (Reference Vachard, Martini, Rettori and Zaninetti1994). In Beccaletto et al. (Reference Beccaletto, Bartolini, Martini, Hochuli and Kozur2005), the specimen illustrated from an Upper Triassic block of northwestern Turkey does not belong to the E. kocaeliensis population. The coiled portion of this form differs from the kocaeliensis type by the presence of more chambers. The illustrated specimen might not even belong to the genus Endotebanella because the uncoiled portion appears to be biserial. Endotebanella kocaeliensis specimens illustrated from Upper Triassic atoll-like carbonates of Japan by Chablais (Reference Chablais2010) are similarly not convincing. Coiled portions are not clearly visible in the illustrated specimens. Finally, in a more recent study, the specimen illustrated as E. kocaeliensis from the Qingyan section of south China by Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015) is close to our Endotebanella sp. 2 (Fig. 18.11). Endotebanella kocaeliensis ranges from the Spathian to the Bithynian in the Guandao section (Fig. 11). It has been recorded from Aegean to lower Longobardian strata in the Upper Guandao section (Figs. 8, 9).
Endotebanella sp. 1
Figure 18.12
Remarks
The other two populations of Endotebanella, E. sp. 1 (Fig. 18.12) and E. sp. 2 (Fig. 18.11), are characterized by a maximum of three chambers in the coiled stage and a slender test, respectively. Both forms are rare in the Bithynian and Pelsonian of the Guandao and Upper Guandao sections (Figs. 8, 11).
Endotebanella sp. 2
Figure 18.11
Remarks
See Remarks under Endotebanella sp. 1.
Genus Spinoendotebanella new genus
Type species
Spinoendotebanella lehrmanni new species.
Diagnosis
A coiled–uncoiled endotebid genus with spinose projections at chamber corners in the coiled stage.
Etymology
The prefix spino- in the name of the new genus has been used to refer the presence of spinose projections at chamber corners.
Remarks
Spinoendotebanella n. gen. is the only representative of the Family Endotebidae exhibiting spinose projections at chamber corners of the coiled stage. It was most probably derived from the genus Endotebanella.
Spinoendotebanella lehrmanni new species
Figure 18.7–18.10
Holotype
The specimen comes from sample PUG-027 (Fig. 18.7), Upper Guandao section.
Diagnosis
A species of Spinoendotebanella n. gen. with nearly two complete planispiral whorls, five or six chambers in the last whorl, and a test attaining 1 mm in length.
Occurrence
Bithynian, Upper Guandao section of the Great Bank of Guizhou, Nanpanjiang Basin, south China, PUG-027.
Description
Following a proloculus of medium size, the coiled portion of the test consists of two nearly planispiral whorls with 4–5 chambers in the last whorl. In the second whorl and uncoiled portion, chambers gradually increase in height. Chamber corners of the second whorl are generally ornamented with triangular spinose projections. When spinose projections are captured in the plane of equatorial section, chamber cavities also appear to be triangular in section. The uncoiled portion contains as many as five chambers. Wall is typically endotebid-type and the aperture system is as described in the definition of the genus.
Etymology
The type species is dedicated to Prof. Dr. Daniel J. Lehrmann (Trinity University, USA) for his extensive and valuable contributions to the stratigraphy, sedimentology, and paleontology of the Nanpanjiang Basin, south China.
Materials
Samples PUG-12, 23, 27, 29, 43 (Aegean to Pelsonian). More than 10 specimens, four of which are illustrated in Figure 18.7–18.10.
Microfossil association
In the samples, the new species is accompanied by Planiinvoluta? mesotriasica, Arenovidalina abriolense, Meandrospira pusilla, M. dinarica, Hoyenella gr. H. sinensis, Hoyenella sp. 1, Glomospirella sp. 1, Pilammina densa, Pilamminella grandis, Reophax sp. 5, Earlandia tintinniformis, Endoteba obturata, Endotriada tyrrhenica Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994, Endotriada sp. 1, Endotriadella wirzi (Koehn-Zaninetti, Reference Koehn-Zaninetti1968), Endotriadella wirzi?, E. lombardi (Zaninetti and Brönnimann in Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972c), Triadodiscus sp., Aulotortus? eotriasicus Zaninetti, Rettori, and Martini, Reference Zaninetti, Rettori and Martini1994, Lamelliconinae?, Krikoumbilica pileiformis, Diplotremina sp. 2, and Dentalina? sp. 2.
Dimensions
Diameter of proloculus: 52–60 μm (holotype: 52 μm). Diameter of coiled part: 360–400 μm (holotype: 380 μm). Maximum width of test: 380 μm. Maximum length of test: 955 μm (holotype: 720 μm). Thickness of wall 35–45 μm (holotype: 35 μm).
Remarks
Spinoendotebanella lehrmanni n. gen. n. sp. is the only endotebid population ornamented with spines at chamber corners of the second whorl of its spire. It is very probable that the morphologically close Endotebanella kocaeliensis is the ancestor of this form.
Family Endotriadidae Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994
Remarks
The other Endothyra-like group in the Triassic of the GBG is the family Endotriadidae, which differs from Endotebidae primarily in having a thin and uniform, microgranular wall. The type-genus, Endotriada, consists of four different populations.
Genus Endotriada Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994
Endotriada tyrrhenica Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994
Figure 18.13–18.17
Remarks
The type-species, E. tyrrhenica, is present in the GBG and characterized by a small test with depressed sutures, 6–7 chambers in the last whorl, and a highly compressed profile in the axial section. The forms illustrated (Fig. 18.13–18.17) are nearly identical to forms described by Vachard et al. (Reference Vachard, Martini, Rettori and Zaninetti1994). Although E. tyrrhenica is a well-established species, several sections attributed to the tyrrhenica population from the Upper Triassic of Slovenia (Gale et al., Reference Gale, Kolar-Jurkovšek, Šmuc and Rožič2012), Turkey (Beccaletto et al., Reference Beccaletto, Bartolini, Martini, Hochuli and Kozur2005), Thailand (Ueno et al., Reference Ueno, Miyahigashi, Kamata, Kato, Charoentitirat and Limruk2012), and Japan (Chablais, Reference Chablais2010) and the Middle Triassic of Indonesia (Rossignol et al., Reference Rossignol, Bourquin, Hallot, Poujol, Debard, Martini, Villeneuve, Cornée, Brayard and Roger2018) are problematic either because of incomplete illustrations or incorrect interpretation of the wall structure and the test morphology. Endotriada tyrrhenica has been found in the Pelsonian to Illyrian strata of the Upper Guandao section (Fig. 8).
Endotriada sp. 1
Figure 18.20, 18.21
Remarks
Three populations of Endotriada are left in open nomenclature. Endotriada sp. 1 is characterized by lenticular tests, always with an initial skewed coiling. Endotriada sp. 1 has been recorded from the Bithynian to Pelsonian (Figs. 8, 9).
Endotriada sp. 2
Figure 18.22–18.24
Remarks
Endotriada sp. 2 is characterized by nautiloid tests, always with an initial skewed coiling. Endotriada sp. 2 has been recorded from the Pelsonian to Fassanian interval (Figs. 8, 9).
Endotriada sp. 3
Figure 18.18, 18.19
Remarks
Endotriada sp. 3 is characterized by biumbilicate tests, always with an initial skewed coiling. Endotriada sp. 3 has been recorded very rarely from the Illyrian of the Upper Guandao section (Figs. 8, 9).
Genus Endotriadella Vachard et al., Reference Vachard, Martini, Rettori and Zaninetti1994
Endotriadella wirzi (Koehn-Zaninetti, Reference Koehn-Zaninetti1968)
Figure 19.1–19.14
Remarks
The other member of the family Endotriadidae, Endotriadella, contains two distinct species. The E. wirzi population, quite frequent in the Triassic of the GBG, is nearly identical to specimens described as Ammobaculites wirzi by Koehn-Zaninetti (Reference Koehn-Zaninetti1968, Reference Koehn-Zaninetti1969) and Zaninetti et al. (Reference Zaninetti, Brönnimann and Baud1972c), with a spiral portion of the test composed of 1.5–2 whorls and an uncoiled, rectilinear part consisting of 6–8 wide and globular chambers. Koehn-Zaninetti (Reference Koehn-Zaninetti1968) published a condensed version of Koehn-Zaninetti (Reference Koehn-Zaninetti1969) with the same title in which she reported the new taxa that she discovered in this paper. Although these taxa were also described as new in 1969 (her main work) we give priority to the 1968 publication.
Endotriadella wirzi has been found in the uppermost Spathian–Bithynian in the Guandao section (Fig. 11), the Aegean–Longobardian of the Upper Guandao section (Figs. 8, 9), and the Pelsonian–Longobardian of the Middle Triassic Dajiang section (Fig. 10). Endotriadella wirzi has been reported extensively from the Middle Triassic of Europe (Zaninetti, Reference Zaninetti1976; Rettori, Reference Rettori1995). In more recent literature, Emmerich et al. (Reference Emmerich, Zamparelli, Bechstädt and Zühlke2005) reported E. wirzi from the Illyrian to Ladinian of the Latemar platform (Dolomites, Italy). Apthorpe (Reference Apthorpe2003) described a terminal opening with possible radiating slits in the free specimens of E. wirzi extracted from the Locker Shale, Western Australia. In Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015), the first appearance of E. wirzi was reported from the uppermost Smithian of south China. Sections of Diplosphaerina inaequalis Derville, Reference Derville1931, reported from the Anisian by these authors, are probably the oblique sections cutting through the uncoiled portion of E. wirzi.
Endotriadella wirzi?
Figure 19.15, 19.16
Remarks
Endotriadella wirzi?, with a much larger test and wider chambers in the uncoiled part, has a similar stratigraphic range to that of E. wirzi. Flügel et al. (Reference Flügel, Ramovš and Bucur1994) reported forms similar to our E. wirzi? as Earlandinita grandis Salaj, Reference Salaj1978 from the Pelsonian of Slovenia. Specimens in Rettori et al. (Reference Rettori, Angiolini and Muttoni1994) from Hydra (Greece), illustrated as E. wirzi, are nearly identical to the uncoiled portion of our E. wirzi?.
Endotriadella lombardi (Zaninetti and Brönnimann in Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972c)
Figure 19.17, 19.18
Remarks
Another distinct species, E. lombardi (Fig. 19.17, 19.18), is characterized by tests with a shorter uncoiled portion and a more prominent initial spiral part when compared with E. wirzi (Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972c). This form is present in the Pelsonian to Longobardian strata of the Upper Guandao section (Figs. 8, 9).
Class uncertain
Remarks
This uncertain class includes all Triassic aragonite-walled foraminifera belonging to orders Involutinida and Robertinida. In the latest classification based on molecular phylogenetic studies, Pawlowski et al. (Reference Pawlowski, Holzmann and Tyszka2013) suggested that Order Involutinida could be assigned to their Class Tubothalamea, which contains both agglutinated and calcareous (calcitic and aragonitic) taxa. They suggested that Order Robertinida falls within their Class Globothalamea, consisting of several multi-chambered groups (except the orders Lagenida and Fusulinida) with agglutinated and calcareous walls. In this study, our taxonomic classification requires wall composition and structure to be used in our taxonomic hierarchy, including the class rank. The study of Pawlowski et al. (Reference Pawlowski, Holzmann and Tyszka2013) could be a meaningful agglomeration of different taxonomic groups. However, the taxonomic rank that they suggest should not be considered in class rank; instead, it may be a higher position in the hierarchical classification of foraminifera.
Order Involutinida Hohenegger and Piller, Reference Hohenegger and Piller1977
Family Triadodiscidae Zaninetti, Reference Zaninetti, Rettori and Martini1984 emend. Altıner and Payne, Reference Altıner and Payne2017
Subfamily Triadodiscinae Zaninetti, Reference Zaninetti, Rettori and Martini1984
Genus Praetriadodiscus Altıner and Payne, Reference Altıner and Payne2017
Remarks
The family Triadodiscidae was introduced by Zaninetti (Reference Zaninetti1984) for the planispirally coiled, lenticular, and trochospirally coiled conic forms, with aragonitic walls and a coiling mode consisting of one lamella for one complete whorl of the deuteroloculus. It recently has been emended by Altıner and Payne (Reference Altıner and Payne2017) and a new genus, Praetriadodiscus, was created in the subfamily Triadodiscinae for lenticular and planispiral forms characterized by a double-layered, inner microgranular and outer hyaline and aragonitic wall structure.
Praetriadodiscus zaninettiae Altıner and Payne, Reference Altıner and Payne2017
Figure 19.20, 19.22
Remarks
Praetriadodiscus zaninettiae differs from its congener, P. tappanae, in having a smaller form ratio (D/W), and thus a more inflated test in axial section. Both of these species evolved from a Pseudoammodiscus ancestor (Fig. 19.19) and the transition from the ancestor to Praetriadodiscus has been clearly illustrated in Altıner and Payne (Reference Altıner and Payne2017). Two distinct populations, P. zaninettiae and P. tappanae, come from the Smithian of the Guandao section (Fig. 11) and are partly illustrated in this study, in addition to the specimens illustrated in Altıner and Payne (Reference Altıner and Payne2017).
Praetriadodiscus tappanae Altıner and Payne, Reference Altıner and Payne2017
Figure 19.20, 19.22
Remarks
Praetriadodiscus? sp. is a form characterized by deviations of the axis of coiling as seen in the axial profile and a less-developed outer hyaline layer.
Genus Triadodiscus Piller, Reference Piller1983
Triadodiscus sp.
Figure 19.25–19.27
Remarks
Forms illustrated as Triadodiscus sp. differ from the type of Triadodiscus, T. eomesozoicus, in having more robust tests with a less angular periphery and more densely coiled deuteroloculus with a low chamber height. A very similar population to our Triadodiscus sp. has been illustrated as Arenovidalina amylovoluta by Kobayashi (Reference Kobayashi1996) from the Kanto Mountains, Japan. In Kobayashi et al. (Reference Kobayashi, Martini and Zaninetti2005), similar forms have been reported as T. eomesozoicus from the Anisian of western Kyushu, Japan. Triadodiscus sp. has been recovered from the Bithynian of the Guandao and Upper Guandao sections (Figs. 8, 11) and the Pelsonian of the Middle Triassic Dajiang section (Fig. 10). Involutinid forms reported from south China as ‘Triadodiscus eomesozoicus’ (Oberhauser, Reference Oberhauser1957) and ‘Aulotortus? bakonyensis’ Blau, Reference Blau1989 by Song et al. (Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016) may have been misclassified. These specimens were first reported by Song et al. (Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan, Niu, Chen, Yang and Wang2011a) from the upper Smithian in association with species such as Meandrospira dinarica and Pilammina densa, which are typical markers of the Anisian.
Subfamily Lamelliconinae Zaninetti et al., Reference Zaninetti, Ciarapica, Decrouez and Martini1987b?
Lamelliconinae?
Figure 20.1–20.5
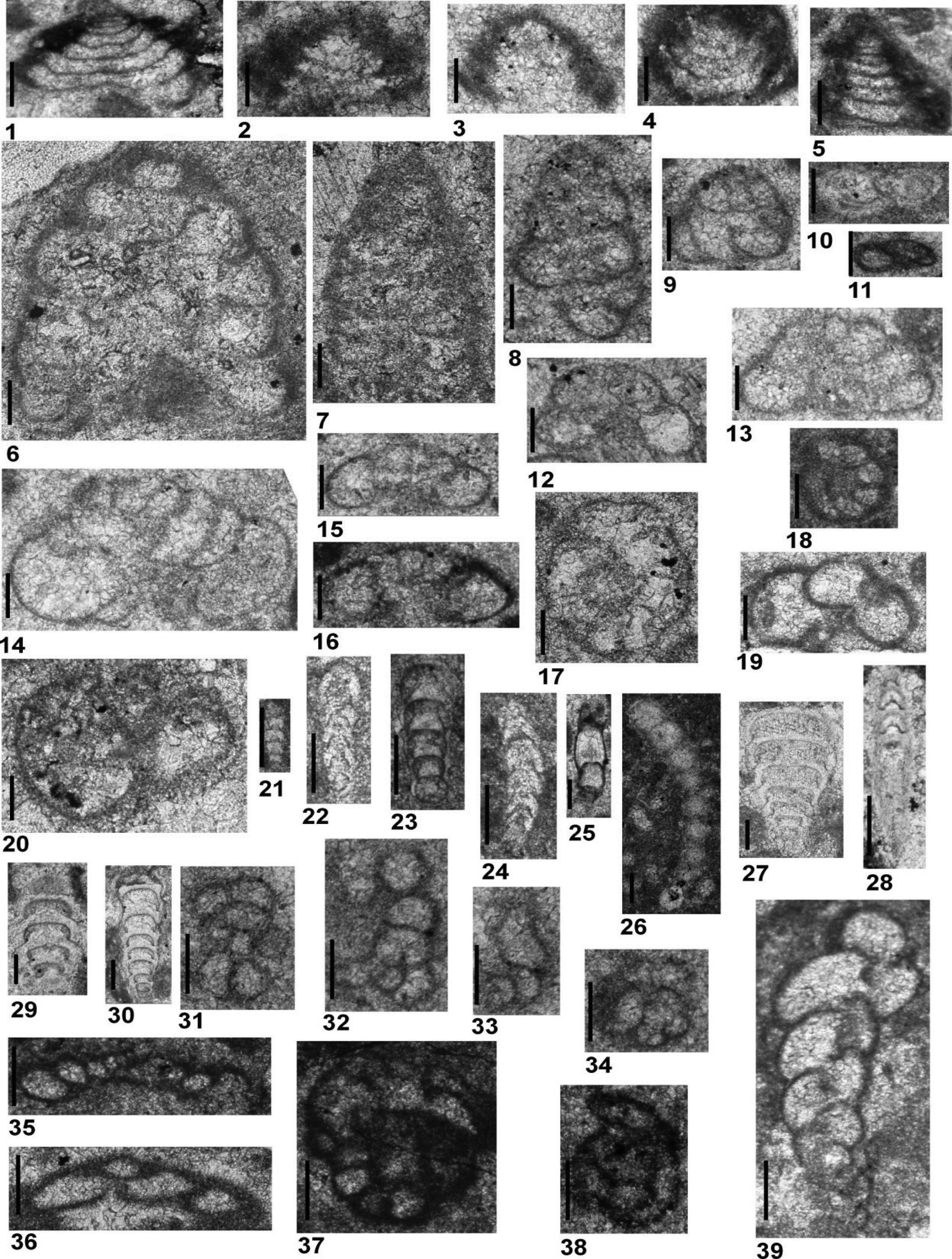
Figure 20. All specimens are from Dawen (PDW), Dajiang (PDJ), Middle Triassic Dajiang (MDJ), Guandao (PGD), and Upper Guandao (PUG) sections. (1–5) Lamelliconinae?; (6) Variostoma? sp. 1; (7) Variostoma? sp. 2; (8) Variostoma sp. 1; (9) Variostoma sp. 2; (10) Plagiostomella? sp. (11) Oberhauserella? sp.; (12, 13, 17, 18) Krikoumbilica pileiformis He, 1984; (14) Diplotremina sp. 1; (15, 16) Diplotremina sp. 2; (19) Abriolina? sp.; (20) Abriolina mediterranea Luperto, 1963; (21) Polarisella elabugae (Cherdyntsev, 1914); (22, 24) Polarisella hoae (Trifonova, 1967); (23) Protonodosaria exploita (Trifonova, 1978c); (25) Dentalina? sp. 1; (26) Dentalina? sp. 2; (27, 28) Geinitzinita sp.; (29) Nodosinelloides? sp.; (30) Austrocolomia sp. (31–34) Unknown foraminifera 1; (35–37) Unknown foraminifera 2; (38) Unknown foraminifera 4; (39) Unknown foraminifera 3. (1, 3–5) PUG-029; (2) PUG-043; (6) PUG-137; (7) PUG-111; (8) PUG-063; (9, 15) PUG-081; (10) PUG-065; (11) PUG-037; (12) MDJ-39; (13) PUG-027; (14) PUG-031; (16) PGD-217; (17) PGD-185; (18) PUG-039; (19, 24) PUG-091; (20) PUG-093; (21) MDJ-07; (22) PUG-097; (23) PGD-225; (25) PUG-019; (26) PUG-041; (27) PUG-109; (28) PUG-017; (29) PUG-085; (30) PUG-145; (31–34) PDW-340; (35, 37) PGD-211; (36) PGD-212; (38) PDJ-058; (39) MDJ-47. Scale bars = 100 μm.
Remarks
Trochospirally coiled, conical forms with five or six whorls and aragonitic walls from the Aegean–Pelsonian of the Upper Guandao section and the Bithynian of the Guandao section (Figs. 8, 11) are reported as Lamelliconinae? In this study. These forms are probably the oldest records of the trochospiral involutinid foraminifera. Some Anisian forms similar to our Chinese specimens have been described in previous studies (e.g., Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972c; Zaninetti, Reference Zaninetti1976) as populations belonging to the genus Turrispirillina (Spirillinidae). We strongly question the validity of this taxonomy and argue that such forms, lacking a proper calcitic wall, should be classified under a taxon housing bilocular and trochospiral forms with aragonitic walls.
Family Aulotortidae Zaninetti, Reference Zaninetti1984
Subfamily Parvalamellinae Rigaud, Martini, and Rettori, Reference Rigaud, Martini and Rettori2012
Genus Parvalamella Rigaud, Martini, and Rettori, Reference Rigaud, Martini and Rettori2012
Parvalamella sp.
Figure 19.28
Remarks
Following Rigaud et al. (Reference Rigaud, Martini and Rettori2012), we assign our streptospirally coiled, recrystallized involutinid forms to Parvalamella sp. The Chinese forms are most likely closely related to the Parvalamella praegaschei-friedli group (Parvalamella praegaschei Koehn-Zaninetti, Reference Koehn-Zaninetti1968 and Parvalamella friedli [Kristan-Tollmann, Reference Kristan-Tollmann1962]). However, we disagree with Rigaud et al. (Reference Rigaud, Martini and Rettori2012) in the assignment of their newly created subfamily Parvalamellininae to the family Involutinidae. In our view, the presence of laterally more restricted L2 lamellae in the subfamily Parvalamellininae does not justify this distinction. The subfamily Aulotortinae is not composed only of planispiral forms, as Rigaud et al. (Reference Rigaud, Martini and Rettori2012) have stated in comparison with Parvalamellininae. On the contrary, the presence of populations displaying an irregular coiling in Rigaud et al.'s (Reference Rigaud, Martini and Rettori2012) ontogeny in Aulotortinae (ex., Aulotortus gr. A. sinuosus Weynschenk, Reference Weynschenk1956) is evidence for grouping Parvalamellininae and Aulotortinae under the family Aulotortidae. Parvalamella sp. has been recorded from Ladinian (Fassanian to Longobardian) of the Middle Triassic Dajiang section (Fig. 10).
Subfamily Aulotortinae Zaninetti, Reference Zaninetti1984
Genus Aulotortus Weynschenk, Reference Weynschenk1956
Aulotortus? eotraisicus Zaninetti, Rettori, and Martini, Reference Zaninetti, Rettori and Martini1994
Figure 19.29, 19.30
Remarks
We assign lenticular involutinid forms with a subangular periphery and a nearly planispiral coils whose diameters approach 1 mm to Aulotortus? eotriasicus, described by Zaninetti et al. (Reference Zaninetti, Rettori and Martini1994). This form was previously reported by Brönnimann et al. (Reference Brönnimann, Cadet and Zaninetti1973a) as Involutina sinuosa pragsoides Oberhauser, Reference Oberhauser1964 and, as noted in Zaninetti et al. (Reference Zaninetti, Rettori and Martini1994), A.? eotriasicus differs from this latter form in having a narrower tubular chamber, a larger number of whorls, and a subangular periphery of the test. In more recent literature, A.? eotriasicus has been reported as A.? sp. from the Anisian of the Kanto Mountains, Japan (Kobayashi, Reference Kobayashi1996). Emmerich et al. (Reference Emmerich, Zamparelli, Bechstädt and Zühlke2005) reported typical specimens of A.? eotriasicus from the Anisian of the Latemar platform (Dolomites, Italy). Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015) illustrated A.? eotriasicus as A. sinuosus from the Anisian of the Qingyan section of south China. Aulotortus? eotriasicus has been recorded from the uppermost Bithynian to Pelsonian in the Upper Guandao section (Fig. 8).
Order Robertinida Loeblich and Tappan, Reference Loeblich and Tappan1984
Family Variostomatidae Kristan-Tollmann, Reference Kristan-Tollmann1963
Remarks
Variostomatidae is the oldest family in the Order Robertinida, which originated in the Early Triassic (Rigaud et al., Reference Rigaud, Martini and Vachard2015; Rigaud and Blau, Reference Rigaud and Blau2016). As stated in Zaninetti (Reference Stampfli, Zaninetti, Brönnimann, Jenny-Deshusses and Stampfli-Vuille1976), it is difficult to study variostomatid taxa in thin section, even if sections of foraminifera are oriented. This difficulty arises because of the complexity of the apertural system of the group, which is a crucial morphological feature used in the distinction of genera. For this reason, with the exception of Krikoumbilica pileiformis, we have assigned different morphogroups recognized in thin section to different variostomatid taxa with open nomenclature.
Genus Krikoumbilica He, Reference He1984
Krikoumbilica pileiformis He, Reference He1984
Figure 20.12, 20.13, 20.17, 20.18
Remarks
Krikoumbilica pileiformis has been recognized in the Chinese material in the Spathian of the Guandao section (Fig. 11), the upper Bithynian to Longobardian of the Upper Guandao section (Figs. 8, 9), and the Illyrian of the Middle Triassic Dajiang section (Fig. 10). It is characterized by a large umbilicus and plano-convex test. Following the description of K. pileiformis from the Anisian–Ladinian of Guizhou, south China, by He (Reference He1984), this species has been partly or fully illustrated as Diplotremina or D. astrofimbriata Kristan-Tollmann, Reference Kristan-Tollmann1960 in some studies from Italy (Senowbari-Daryan et al., Reference Senowbari-Daryan, Zühlke, Bachtädt and Flügel1993), Japan (Kobayashi et al., Reference Kobayashi, Martini and Zaninetti2005), Thailand (Kobayashi et al., Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006), and Laos (Miyahigashi et al., Reference Miyahigashi, Hara, Hisada, Nakano, Charoentitirat, Charusiri, Khamphoveng, Martini and Ueno2017). In Song et al. (Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015), the axial sections of K. pileiformis have been reported as Duotaxis spp. from the Anisian of Qingyan section, south China.
Genus Variostoma Kristan-Tollmann, Reference Kristan-Tollmann1960
Remarks
Four different trochospiral forms of varying degree have been assigned to Variostoma. Two of these populations (Variostoma? sp. 1 and V.? sp. 2), characterized by a rather wider umbilicus, have been doubtfully assigned to the genus.
Variostoma? sp. 1
Figure 20.6
Remarks
Variostoma? sp. 1 has been found in the Cordevolian of the Upper Guandao section (Fig. 9).
Variostoma? sp. 2
Figure 20.7
Remarks
Variostoma? sp. 2 has been found in the Fassanian–Longobardian of the Upper Guandao section (Fig. 9).
Variostoma sp. 1
Figure 20.8
Remarks
Variostoma sp. 1 has been recognized in the Spathian of the Guandao section (Fig. 11) and the Longobardian of the Upper Guandao section (Fig. 9).
Variostoma sp. 2
Figure 20.9
Remarks
Variostoma sp. 2 has been recorded in the Spathian to Aegean of the Guandao section (Fig. 11) and the Illyrian of the Upper Guandao section (Fig. 8).
Genus Diplotremina Kristan-Tollmann, Reference Kristan-Tollmann1960
Remarks
Low trochospiral morphogroups have been assigned to the genus Diplotremina.
Diplotremina sp. 1
Figure 20.14
Remarks
Diplotremina sp. 1 has been recorded from the Pelsonian of the Upper Guandao section (Fig. 8).
Diplotremina sp. 2
Figure 20.15, 20.16
Remarks
Diplotremina sp. 2 is found in the Spathian to Bithynian of the Guandao section (Fig. 11), the Aegean to Pelsonian of the Upper Guandao section (Fig. 8), and the Longobardian of the Middle Triassic Dajiang section (Fig. 10).
Genus Plagiostomella Kristan-Tollmann, Reference Kristan-Tollmann1960
Plagiostomella? sp.
Figure 20.10
Remarks
A very low trochospiral form with nearly pseudoplanispiral coiling assigned to Plagiostomella? sp. has been recognized from the uppermost Pelsonian in the Upper Guandao section (Fig. 9), earlier than the known stratigraphic range of the genus, which is Carnian (Zaninetti, Reference Zaninetti1976).
Family Oberhauserellidae? Fuchs, Reference Fuchs1970
Genus Oberhauserella Fuchs, Reference Fuchs1967
Oberhauserella? sp.
Figure 20.11
Remarks
The family Oberhauserellidae, as an aragonitic taxon, has been classified in the Order Robertinida in recent literature (Rigaud et al., Reference Rigaud, Martini and Vachard2015). Very rare, small specimens, doubtfully assigned to Oberhauserella? sp., have been recorded in the Pelsonian of the Upper Guandao section (Fig. 8). These forms are characterized by very low trochospiral axial sections with a convex spiral side and a depressed umbilicus probably covered by an apertural flap.
Class Nodosariata Mikhalevich, Reference Mikhalevich1993
Family Protonodosariidae Mamet and Pinard, Reference Mamet and Pinard1992
Remarks
This family, which originated in the latest Carboniferous and occurs frequently in the Permian, is a Permian-Triassic boundary survivor and its stratigraphic range extends up to the end of the Triassic (Vachard, Reference Vachard2016, Reference Vachard, Lucas and Shen2018). Two well-known genera from the Paleozoic, Protonodosaria and Polarisella, are present in the Triassic of the GBG. The genus Nodosinelloides is questionably present. Austrocolomia has been included under the family Protonodosariidae in this study.
Genus Polarisella Mamet and Pinard, Reference Mamet and Pinard1992
Polarisella elabugae (Cherdyntsev, Reference Cherdyntsev1914)
Figure 20.21
Remarks
Characterized by thick-walled spherical chambers and a small, igloo-shaped, reduced lumen, the elabugae population is described as Polarisella elabugae (Cherdyntsev, Reference Cherdyntsev1914) in this study. This form was described for the first time by Cherdyntsev (Reference Cherdyntsev1914) as Nodosaria elabugae. Two distinct forms, N. abriolinae described from the Abriola Limestone in Italy by Luperto (Reference Luperto1963) and ‘N’. armeniensis described from the Caucasus by Efimova (Reference Efimova1974), are synonyms of the elabugae population. It is common in the Changhsingian limestone of the Taurus Mountains (Lys and Marcoux, Reference Lys and Marcoux1978; Altıner, Reference Altıner1981, Reference Altıner, Tekeli and Göncüoğlu1984; Groves et al., Reference Groves, Altıner and Rettori2005) and is thus a Permian-Triassic boundary survivor. In our material, this form has been recorded from the Pelsonian of the Upper Guandao and Middle Triassic Dajiang sections (Figs. 8, 10). Recently, it also has been reported from south China as ‘N.’ elabugae from the Hindeodus parvus Zone of the Griesbachian Substage (Dai et al., Reference Dai, Song, Wignall, Jia, Bai, Wang, Chen and Tian2018) and Permian-Triassic boundary beds of Slovenia (Kolar-Jurkovšek et al., Reference Kolar-Jurkovšek, Jurkovšek, Nestell and Aljinović2018).
Polarisella hoae (Trifonova, Reference Trifonova1967)
Figure 20.22, 20.24
Remarks
Polarisella hoae was originally described as Dentalina hoi and is characterized by nearly conical, thick-walled chambers circumscribing small conical lumina. It is a Permian-Triassic boundary survivor because it is present in the Changhsingian and in Lower Triassic strata of southern Turkey (Groves et al., Reference Groves, Altıner and Rettori2005). In China, it is present in the entire Triassic (Ho, Reference Ho1959; Lin, Reference Lin, Meng and Zhang1987; He and Cai, Reference He and Cai1991; He, Reference He1993). In the GBG material, it has been recorded from the Pelsonian to Fassanian of the Upper Guandao section (Figs. 8, 9) and the Illyrian of the Middle Triassic Dajiang section (Fig. 10). In recent literature, it has been reported from the Lower Triassic Khuff Formation of Saudi Arabia (Vachard et al., Reference Vachard, Gaillot, Vaslet and Le Nindre2005) and from the PTB transitional beds of western Slovenia (Kolar-Jurkovšek et al., Reference Kolar-Jurkovšek, Jurkovšek, Nestell and Aljinović2018).
Genus Protonodosaria Gerke, Reference Gerke1959
Protonodosaria exploita (Trifonova, Reference Trifonova1978c)
Figure 20.23
Remarks
Characterized by spherical chambers, each perched high atop the previous one, and a rather large proloculus, Protonodosaria exploita has been found in the Aegean to Bithynian deposits of the Guandao section (Fig. 11) and the Bithynian to Pelsonian of the Upper Guandao section (Fig. 8). This species is known mostly from the Anisian of Bulgaria (Trifonova, Reference Trifonova1978b), China (Kristan-Tollmann, Reference Kristan-Tollmann1983), Japan (Kobayashi, Reference Kobayashi1996), and Turkey (Groves et al., Reference Groves, Altıner and Rettori2005). Recently, it has been reported from the Griesbachian of Italy (Groves et al., Reference Groves, Rettori, Payne, Boyce and Altıner2007), south China (Song et al., Reference Song, Tong, Chen, Yang and Wang2009), and western Slovenia (Nestell et al., Reference Nestell, Kolar-Jurkovšek, Jurkovšek and Aljinović2011).
Genus Nodosinelloides Mamet and Pinard, Reference Mamet and Pinard1992
Nodosinelloides? sp.
Figure 20.29
Remarks
Sections of probable protonodosariid foraminifera reported as Nodosinelloides? sp. in this study are characterized by uniserial, hemispherical to subquadratic chambers. However, it is not certain whether the sections in the GBG material are characterized by a simple terminal opening, as is the case in Nodosinelloides, which is a late Carboniferous–Permian taxon. If the forms that we recognize are true Nodosinelloides, then this taxon should also be a Permian/Triassic boundary survivor. Nodosinelloides? sp. has been recorded from the Spathian to Bithynian in the Guandao section (Fig. 11) and the Bithynian to Cordevolian in the Upper Guandao section (Figs. 8, 9). According to Song et al. (Reference Song, Tong, Zhang, Wang and Chen2007, Reference Song, Tong and Chen2011b), Nodosinelloides (given as N. aequiampla [Zolotova and Baryshnikov, Reference Zolotova and Baryshnikov1980]) is present in the Griesbachian of south China.
Genus Austrocolomia Oberhauser, Reference Oberhauser1960
Austrocolomia sp.
Figure 20.30
Remarks
Austrocolomia sp. is possibly an intermediate form between A. cordevolica Oberhauser, Reference Oberhauser1967 and A. marschalli Oberhauser, Reference Oberhauser1967, and is characterized by the gradual increase in the width of uniserial hemispherical to subquadratic chambers with marked shoulders at chamber corners and a simple terminal aperture. This form has been found in the Cordevolian of the Upper Guandao section (Fig. 9).
Family Geinitzinidae Bozorgnia, Reference Bozorgnia1973
Genus Geinitzinita Sellier de Civrieux and Dessauvagie, Reference Sellier de Civrieux and Dessauvagie1965
Geinitzinita sp.
Figure 20.27, 20.28
Remarks
Geinitzinita, a member of the Family Geinitzinidae that appeared and flourished in the Permian (Vachard, Reference Vachard2016, Reference Vachard, Lucas and Shen2018), occurs rarely in the Chinese material. Characterized by low and wide chambers in the longitudinal frontal sections, the diagnostic feature of the genus is visible in oblique longitudinal lateral sections with centrally arched chambers. Geinitzinita sp. has been recorded from the Bithynian to Longobardian of the Upper Guandao section (Figs. 8, 9).
Family Nodosariidae? Ehrenberg, Reference Ehrenberg1838
Genus Dentalina Risso, Reference Risso1826
Remarks
Two arcuate nodosariid forms with an unknown type of terminal aperture have been questionably assigned to the genus Dentalina. These questionable forms are found in the Anisian of the Guandao and Upper Guandao sections (Fig. 8, 11).
Dentalina? sp. 1
Figure 20.25
Remarks
See Remarks under Genus Dentalina. In the literature, specimens close to our D.? sp. 2 were illustrated as Dentalina vadaszi Oberhauser, Reference Oberhauser1960 by Song et al. (Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan, Niu, Chen, Yang and Wang2011a, Reference Song, Wang, Tong, Chen, Tian, Song and Chu2015) from the Olenekian to Anisian of south China.
Family Abriolinidae Zaninetti and Rettori in Zaninetti et al., Reference Zaninetti, Martini and Altıner1992b
Genus Abriolina Luperto, Reference Luperto1963
Abriolina sp.
Figure 20.19
Remarks
When initially created by Zaninetti and Rettori in Zaninetti et al. (Reference Zaninetti, Rettori, Martini, Cirilli and Ciarapica1992), Abriolinidae was placed in the superfamily Geinitzinacea, which is considered one of the main taxonomic units of the Class Nodosariata (Vachard, Reference Vachard2016, Reference Vachard, Lucas and Shen2018). Although it is reasonable on morphological grounds to place this double-wall-bearing group in the superfamily Geinitzinacea, which originated in the Paleozoic, our observations of the Chinese material suggest that the family Abriolinidae might be a morphologically convergent form that evolved during the Triassic. Trochospiral forms morphologically similar to trochamminid-like forms in the Illyrian of the Upper Guandao section (Fig. 8) could represent a population ancestral to Abriolinidae. We classify such forms as Abriolina? sp. (Fig. 20.19) due to the presence of a rudimentary and discontinuous outer hyaline layer covering the main microgranular to finely agglutinated inner layer.
Abriolina mediterranea Luperto, Reference Luperto1963
Figure 20.20
Remarks
The true abriolinid taxon, Abriolina mediterranea, is found in the late Illyrian–Fassanian interval in the Upper Guandao section (Figs. 8, 9). This taxon was created based on specimens from the Abriola Limestone in southern Italy by Luperto (Reference Luperto1963), who erroneously considered this form as a Late Permian foraminiferan. Emmerich et al. (Reference Emmerich, Zamparelli, Bechstädt and Zühlke2005) illustrated the typical specimens of this species from the Illyrian to Ladinian of the Latemar platform (Dolomites, Italy). These low trochospiral forms with a large proloculus and subglobular to subangular chambers are also characterized by a double wall, an inner microgranular to granular layer, and an outer with consistently occurring hyaline fibrous layer.
Unknown foraminifera
Remarks
Four foraminiferal populations could not be named and are left with an open nomenclature in the Chinese material.
Unknown foraminifera 1
Figure 20.31–20.34
Remarks
Unknown foraminifera 1 is probably a calcivertellid and characterized by tests with a coiled stage followed by an uncoiled stage with a folded tubular chamber making zigzag bends. This population has been recorded from the Smithian of the Dawen and the Spathian of the Guandao sections (Fig. 5, 11).
Unknown foraminifera 2
Figure 20.35–20.37
Remarks
Unknown foraminifera 2 is a multilocular form, probably a trochamminid, with at least four whorls and a broad depression toward the umbilicus. It has been found in the Aegean of the Guandao section (Fig. 11).
Unknown foraminifera 3
Figure 20.39
Remarks
Unknown foraminifera 3 is characterized by a reduced coiled stage and an uncoiled portion comprising biserial chambers with a hook-shaped valvular tooth extending from septal edges to protect the aperture. This interesting form is found in the Fassanian of the Middle Triassic Dajiang section (Fig. 10).
Unknown foraminifera 4
Figure 20.38
Remarks
Unknown foraminifera 4 is possibly a porcelaneous tubular form coiled in various planes. It could be a Permian survivor and referred to one of the populations of Hemigordiellina, as emended by Vachard in Vachard and Beckary (Reference Vachard and Beckary1991). However, this form is rare in the Griesbachian of Dajiang section (Fig. 6) and not well enough preserved to decide on the correct taxonomic position.
Biostratigraphy
Several authors emphasized the biostratigraphic importance of Lower–Middle Triassic foraminifera (Zaninetti, Reference Zaninetti1976; Vachard and Fontaine, Reference Vachard and Fontaine1988; Altıner and Koçyiğit, Reference Altıner and Koçyiğit1993; Rettori et al., Reference Rettori, Angiolini and Muttoni1994; Rettori, Reference Rettori1995; Muttoni et al., Reference Muttoni, Kent, Meço, Nicora, Gaetani, Balini, Germani and Rettori1996; Marquez, Reference Marquez2005; Kobayashi et al., Reference Kobayashi, Martini, Rettori, Zaninetti, Ratanasthien, Saegusa and Nakaya2006; Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). There have been numerous studies of the Lower–Middle Triassic foraminiferan biostratigraphy, but the resulting biostratigraphic zones have rarely been used in geological studies or for long-distance correlations.
The first attempts were focused on the European Triassic, particularly in the western Carpathians (Salaj, Reference Salaj1969, Reference Salaj1978; Salaj et al., Reference Salaj, Borza and Samuel1983), Carpatho-Balkans (Salaj et al., Reference Salaj, Trifonova and Gheorghian1988), Bulgaria (Trifonova, Reference Trifonova1978a, Reference Trifonovab, Reference Trifonova1992, Reference Trifonova1993), southern Poland (Gazdzicki et al., Reference Gazdzicki, Trammer and Zwidzka1975), the Transdanubian Range of Hungary (Orovecz-Scheffer, Reference Orovecz-Scheffer1987), and the Prealps in Switzerland (Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972b). Further attempts addressed western Asia, including the Kocaeli Peninsula of Turkey (Zaninetti and Dağer, Reference Zaninetti and Dağer1978) and the Caucasus of the Russian territory (Efimova, Reference Efimova, Azbell and Grigelis1991). None of these studies developed a complete zonal scheme for the entire Lower Triassic–Anisian interval. Apart from taxonomic problems in some of these studies, poor calibration of the ranges of taxa used in the zonation schemes and the strong facies control on the stratigraphic distribution of foraminiferal taxa hindered the use of these zones as standard in the Geologic Time Scale.
Toward the end of the 20th century, two important studies helped to clarify the overall biostratigraphic subdivision of the Lower–Middle Triassic. The first of these studies focused on northwest Sichuan and southern Shaanxi, China (He, Reference He1993). Although incomplete, three main zones were used, namely Arenovidalina chialingchiangensis and Meandrospira pusilla zones for the Scythian and a Glomospira densa-Meandrospira dinarica Zone for the Anisian. These zones were further divided into subzones using critical species such as Glomospirella vulgaris and Glomospira sinensis. A second, more important study, illustrating the zones of foraminifera and biochronohorizons for the Triassic of the Tethyan Realm, has come out in the ‘Mesozoic–Cenozoic of Western European Basins’ volume of de Graciansky et al. (Reference de Graciansky, Hardenbol, Jacquin and Vail1998). In the Griesbachian (Induan)–Illyrian (Anisian) interval, Hardenbol et al. (Reference Hardenbol, Thierry, Farley, Jacquin, de Graciansky, Vail, de Graciansky, Hardenbol, Jacquin and Vail1998; see also Kamoun et al., Reference Kamoun, Peybernès, Martini, Zaninetti, Vila, Trigui and Rigane1998) described the Rectocornuspira kalhori Zone from the Griesbachian, a non-characterized interval spanning from Dienerian to Smithian, the Meandrospira pusilla Zone from the Spathian, the Meandrospira dinarica Zone from the Aegean to Bithynian, the Paulbronnimannia judicariensis Zone from the Pelsonian, and the Pilammina densa Zone from the Illyrian.
There have been no meaningful improvements to the Lower Triassic–Anisian foraminiferal zonation since the study of Hardenbol et al. (Reference Hardenbol, Thierry, Farley, Jacquin, de Graciansky, Vail, de Graciansky, Hardenbol, Jacquin and Vail1998). Among different versions of foraminiferal biostratigraphy proposed for limited intervals of Lower–Middle Triassic, the studies of Muttoni et al. (Reference Muttoni, Kent, Meço, Balini, Nicora, Rettori, Gaetani and Krystyn1998), Faletti and Ivanova (Reference Faletti and Ivanova2003), Vachard et al. (Reference Vachard, Gaillot, Vaslet and Le Nindre2005), Korngreen and Benjamini (Reference Korngreen and Benjamini2006), Velić (Reference Velić2007), Vuks (Reference Vuks2007), Korchagin (Reference Korchagin2008), and Maurer et al. (Reference Maurer, Rettori and Martini2008) can be given as examples.
Griesbachian (Induan)–Illyrian (Anisian) foraminiferal biostratigraphy in the Great Bank of Guizhou
In the Great Bank of Guizhou, 12 biozones and two unnamed intervals have been defined from the base of the Griesbachian (Induan) to the top of the Illyrian (Anisian).
Postcladella kalhori-Postcladella grandis Zone
The type section of this biozone is in the Dawen section and defined as a 107.5 m thick limestone succession between 9.0–116.5 meters of the section. The zone is fixed by the successive first occurrences of Postcladella kalhori and P. grandis (Figs. 2, 21; samples 11–87 in Fig. 5). Postcladella kalhori makes its first appearance <2 m above the end-Permian extinction horizon, marked by an abrupt contact between fossiliferous pre-extinction limestone and post-extinction thrombolitic microbialite, thus the lower boundary of this zone approximates the Permian-Triassic boundary. The limestone lithology of the zone consists of thrombolites, micritic to high-energy limestones, oolitic dolomites, and thrombolite-bearing cyclic limestone deposits. Postcladella kalhori is usually associated with Earlandia sp. 1 and E. sp. 2 in the samples. The zone is also present in the Dajiang section, but absent in the Guandao section, probably due to unfavorable facies consisting of shales, allodapic breccias, and unfossiliferous carbonate mudstones.

Figure 21. Biozones of foraminifera (1–12) and stratigraphic ranges of important foraminifera and conodont species. P. kalhori: Postcladella kalhori; P. grandis: Postcladella grandis; H. gr. H. sinensis: Hoyenella gr. H. sinensis; A. weii: Arenovidalina weii; G. vulgaris: Glomospirella vulgaris; M. pusilla: Meandrospira pusilla; M. dinarica: Meandrospira dinarica; P. densa: Pilammina densa; A.? eotriasicus: Aulotortus? eotriasicus; T. mesotriasica: Turriglomina mesotriasica; A. mediterranea: Abriolina mediterranea.
The Postcladella kalhori-Postcladella grandis Zone is defined for the first time in the Triassic literature. It has the potential to be recognized also in the western Tethys because the first appearance of P. grandis always postdates the first appearance of P. kalhori, both in Turkey (Altıner and Zaninetti, Reference Altıner and Zaninetti1981) and in the Transdanubian Range, Hungary (Orovecz-Scheffer, Reference Orovecz-Scheffer1987).
The Postcladella kalhori-Postcladella grandis Zone corresponds to an important part of the Griesbachian Substage. In Song et al. (Reference Song, Tong, Chen, Yang and Wang2009, Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016), P. kalhori was reported from the H. parvus, I. staeschei, and I. isarsica conodont zones (Griesbachian) of the Dajiang section. No conodonts have been recorded from the levels corresponding to the P. kalhori-P. grandis Zone in the Dawen section. However, Chen et al. (Reference Chen, Beatty, Henderson and Rowe2009) reported some Griesbachian conodont zones from the Dawen section, which is at least partially equivalent to the lower part of our zone. Finally, P. kalhori has been reported from the uppermost Permian in some recent literature (Firi et al., Reference Firi, Sremac and Vlahović2016; Tian et al., Reference Tian, Tong, Xiao, Benton, Song, Liang, Wu, Chu and Algeo2018). However, mentioned or poorly illustrated specimens do not, in our view, belong to the kalhori population in these studies, and there is no need to extend this zone into the uppermost Permian.
Postcladella grandis Zone
Between 116.5 and 160 meters of the Dawen section, a 43.5 m thick, thrombolite-bearing cyclic limestone interval is the type section of the Postcladella grandis Zone. It is a taxon-range zone defined by the first and last occurrences of P. grandis (Figs. 2, 21; samples 87–140 in Fig. 5). The zone also has been recognized in the Dajiang section. The last appearance of P. grandis is very close to the Griesbachian-Dienerian boundary, which is correlated with the Guandao section on the basis of carbon-isotope and conodont data (Payne et al., Reference Payne, Lehrmann, Wei, Orchard, Schrag and Knoll2004; Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015).
In the zone, P. grandis is associated with P. kalhori, Earlandia sp. 2, Cornuspira mahajeri?, and unknown foraminifera 4. Although not properly defined, this zone is surely present in the Taurides (Turkey) (Altıner and Zaninetti, Reference Altıner and Zaninetti1981) and the Transdanubian Range in Hungary (Orovecz-Scheffer, Reference Orovecz-Scheffer1987). The marker of the zone is also present in the Werfen Formation (Austria) (Krainer and Vachard, Reference Krainer and Vachard2011, pl. 5, figs. 9, 10, 14). The upper part of the Griesbachian Substage could be characterized in the future after the definition of this zone in this part of Europe.
Postcladella grandis-Postcladella kalhori Zone
The type section of this zone measures 24 m, between 160 and 184 meters of the Dawen section, and is fixed on the basis of successive last occurrences of P. grandis and P. kalhori (Figs. 2, 21; samples 143–167 in Fig. 5). The Postcladella grandis-Postcladella kalhori Zone, consisting of a thrombolite-bearing cyclic limestone succession, is assigned to the lower part of the Dienerian Substage. This interpretation is based on the carbon isotope correlation and conodonts obtained from the Guandao section, where the Griesbachian-Dienerian boundary is placed on the basis of the first occurrence of Neospathodus dieneri following the last occurrence of Hindeodus parvus in the underlying Griesbachian (Payne et al., Reference Payne, Lehrmann, Wei, Orchard, Schrag and Knoll2004; Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). Consistent with our study, Galfetti et al. (Reference Galfetti, Bucher, Martini, Hochuli, Weissert, Crasquin-Soleau, Brayard, Goudmand, Brühwiler and Goudun2008) properly identified the stratigraphic distribution of P. kalhori based on conodont data from the Nanpanjiang Basin. This study confirms the chronostratigraphic value of our zone, covering at least the lower part of the Dienerian Substage.
Unnamed interval in the Dienerian
The interval consisting of thrombolite-bearing cyclic limestones, between 184 and 253 meters of the Dawen section and 250 and 326 meters of the Dajiang section, does not contain any foraminifera. Therefore, this interval could not be zoned within the biostratigraphic framework of this study. This interval in the platform interior sections probably corresponds in age to breccia deposits in the Guandao slope section in the GBG, an unfavorable setting for the occurrence of foraminifera.
Hoyenella gr. H. sinensis Zone
The type section of this zone is defined in the Dawen section. It consists of a 68 m thick, thrombolite-bearing cyclic limestone between 253 and 321 meters of the section (Figs. 2, 21; samples 246–340 in Fig. 5). Successive first occurrences of Hoyenella gr. H. sinensis and Glomospirella vulgaris define the lower and upper boundaries. The zone is slightly thicker in the Dajiang section, and only the upper part is recognized in the Guandao section. The marker of the zone is associated with H. gr. H. shengi, transitional forms to Meandrospira, Pseudoammodiscus sp., Glomospira sp. (=Pilammina praedensa?), and unknown foraminifera 1 in the Dawen, Dajiang, and Guandao sections. The zone corresponds to the upper part of the Dienerian Substage and the lower part of the Smithian, thus encompassing the Induan-Olenekian boundary (Figs. 2, 21). This boundary is recognized in the Dawen section on the basis of carbon isotope chemostratigraphy and conodont biostratigraphy of the Guandao section, where it is delineated by the first occurrences of Eurygnathodus, Novispathodus waageni, and Nv. posterolongatus following the last occurrence of Dienerian marker Neospathodus dieneri. In recent literature, the first occurrence of Hoyenella gr. H. sinensis in south China (Galfetti et al., Reference Galfetti, Bucher, Martini, Hochuli, Weissert, Crasquin-Soleau, Brayard, Goudmand, Brühwiler and Goudun2008), Israel (Korngreen et al., Reference Korngreen, Orlov-Labkovsky, Bialik and Benjamini2013), and India (Baud and Bhat, Reference Baud and Bhat2014) has been reported in the Dienerian, based on conodonts.
Glomospirella vulgaris-Arenovidalina weii Zone
The zone is 34 m thick and corresponds to the interval between 160 and 194 meters of the Guandao section. It is made up of limestone intercalated with distinct allodapic breccia levels (Figs. 2, 21; samples 96–132 in Fig. 11). The lower and upper boundaries are defined based on the first and last occurrences of Glomospirella vulgaris and Arenovidalina weii n. sp., respectively. In addition to these taxa, the zone contains Hoyenella gr. H. sinensis, H. gr. H. shengi, Pseudoammodiscus sp., and, in the upper part, the oldest involutinid taxa, Praetriadodiscus zaninettiae and P. tappanae (see Altıner and Payne, Reference Altıner and Payne2017). In China, as previously reported by He (Reference He1993), the Glomospirella vulgaris Subzone of the Arenovidalina chialingchiangensis Zone occurs below the Spathian Meandrospira pusilla Zone, and probably corresponds to our zone.
In our biostratigraphic framework, the Glomospirella vulgaris-Arenovidalina weii Zone, assigned a middle to upper Smithian age, corresponds to the upper part of the Discretella discreta and Guangxidella bransoni conodont zones, defined in the Guandao section by Lehrmann et al. (Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015) (see also Fig. 21).
Unnamed interval in the Spathian
In the Guandao section between 194 and 238 meters, an unfavorable lithology consisting of dolomite and allodapic breccia, did not yield any foraminifera (Fig. 2, 21). This interval, ~44 m thick, does contain conodonts and, specifically, the first occurrence of Novispathodus? crassatus constrains the position of the Smithian-Spathian boundary in the Guandao section (Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). The unnamed interval is located below the first occurrence of conodonts Novispathodus? triangularis and Triassospathodus homeri, and corresponds to the lower Spathian.
Meandrospira pusilla Zone
This zone, ~39 m thick and entirely composed of limestone, measures between 238 and 277 meters of the Guandao section. It is defined on the basis of successive first occurrences of Meandrospira pusilla and M. dinarica, and corresponds to an important part of the Spathian Substage (Fig. 2, 21; samples 149–195 in Fig. 11). In the type section, the first occurrence of M. pusilla is above the unnamed interval in the Spathian and is nearly coeval with the first occurrence of Novispathodus? triangularis and Triassospathodus homeri (Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). At the upper boundary, the first occurrence of M. dinarica coincides with the turnover in conodont assemblages represented by Chiosella timorensis, Neogondolella regalis, and Gladiogondolella tethydis. As defined in this study, the zone is incomplete because the lower boundary, marked by the first occurrence of M. pusilla, is facies-controlled. If facies had been suitable, M. pusilla may well have appeared much lower in the succession.
In the zone, in addition to Hoyenella gr. H. sinensis and unnamed foraminifera 1, which range into the underlying zones, several important Triassic species make their first appearances, including Meandrospira cheni, M.? enosi n. sp., M.? sp., Endoteba bithynica, Endotebanella kocaeliensis, Endotebidae, Endotriadella wirzi, Trochammina? sp.1, Verneuilinoides? azzouzi, Krikoumbilica pileiformis, Variostoma sp. 1, Variostoma sp. 2, Diplotremina sp. 2, and Nodosinelloides? sp.
Although the marker of the zone has been used sometimes under different invalid species names, the Meandrospira pusilla Zone is one of the most frequently cited biostratigraphic units from the Lower Triassic. It has been reported from the western Carpathians or Carpatho-Balkan Range (Salaj, Reference Salaj1969, Reference Salaj1978; Trifanova, Reference Trifonova1978a; Salaj et al., Reference Salaj, Borza and Samuel1983, Reference Salaj, Trifonova and Gheorghian1988), the Transdanubian Range of Hungary (Orovecz-Scheffer, Reference Orovecz-Scheffer1987), the Kocaeli Peninsula from Turkey (Zaninetti and Dağer, Reference Zaninetti and Dağer1978), southern China (He, Reference He1993), Tunisia (Kamoun et al., Reference Kamoun, Peybernès, Martini, Zaninetti, Vila, Trigui and Rigane1998), and as a standard zone of Spathian age in the European basins volume (de Graciansky et al., Reference de Graciansky, Hardenbol, Jacquin and Vail1998) by Hardenbol et al. (Reference Hardenbol, Thierry, Farley, Jacquin, de Graciansky, Vail, de Graciansky, Hardenbol, Jacquin and Vail1998).
Meandrospira dinarica-Pilammina densa Zone
This zone, corresponding to the Aegean Substage, is 14 m thick and measures from 18–32 meters in the Upper Guandao section. The lower and upper boundaries of the zone are defined on the basis of successive first occurrences of Meandrospira dinarica and Pilammina densa (Figs. 2, 21; samples 7–14 in Fig. 8). The first occurrence of M. dinarica is in the thin limestone layer overlying the first prominent ash layer of latest Spathian age (Fig. 2) and nearly coincides with the first occurrence of Chiosella timorensis, the index conodont used to draw the Olenekian-Anisian boundary (Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). At the upper boundary, the first occurrence of P. densa is nearly coeval with the first occurrence of the conodont Nicoraella germanica and approximates the Aegean-Bithynian boundary.
In addition to taxa with ranges from the Lower Triassic into the Aegean, such as Hoyenella gr. H. sinensis, H. gr. H. shengi, Meandrospira pusilla, Endoteba bithynica, Endotebanella kocaeliensis, Endotriadella wirzi, Diplotremina sp. 2, and Variostoma sp. 2, the zone contains several other taxa that first occur in the Aegean. These are Arenovidalina abriolense, Meandrospiranella cf. M. samueli, Glomospirella sp. 2, Tolypammina gregaria, Pilamminella grandis, Reophax sp. 2, Reophax sp. 5, Palaeolituonella reclinata, Gaudryina sp., Placopsilina sp., Earlandia tintinniformis, Spinoendotebanella lehrmanni n. gen., n. sp., Lamelliconinae?, and unknown foraminifera 2.
Although M. dinarica has been recognized as a very important marker for Anisian biostratigraphy by several authors (Salaj, Reference Salaj1969; Salaj et al., Reference Salaj, Trifonova and Gheorghian1988; Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972b; He, Reference He1993), it was Rettori et al. (Reference Rettori, Angiolini and Muttoni1994) who noticed the first occurrence of M. dinarica lower in the Anisian and close to the Olenekian-Anisian boundary, based on conodont data from the Eros Limestone in Greece. In the standard zonation of Hardenbol et al. (Reference Hardenbol, Thierry, Farley, Jacquin, de Graciansky, Vail, de Graciansky, Hardenbol, Jacquin and Vail1998; see also Kamoun et al., Reference Kamoun, Peybernès, Martini, Zaninetti, Vila, Trigui and Rigane1998), the M. dinarica-P. densa Zone has been assigned to the Aegean–Bithynian based on the successive occurrences of M. dinarica, Paulbronnimannia judicariensis (Premoli Silva, Reference Premoli Silva1971), and Pilammina densa. In this study, this interval has been shortened to the Aegean because the first occurrence of P. densa is very close to the Aegean-Bithynian boundary, based on reliable conodont data.
Pilammina densa Zone
This zone is defined on the basis of the successive first appearances of Pilammina densa and Aulotortus? eotriasicus. It comprises 23 m of limestone between 32–55 meters in the Upper Guandao section (Figs. 2, 21; samples 15–23 in Fig. 8). The zone corresponds to the Bithynian Substage, except the topmost part where A.? eotriasicus makes its first appearance. The first occurrence of P. densa coincides nearly with the first occurrences of Nicoraella germanica and Ni. kockeli, index conodonts used to draw the Aegean-Bithynian boundary (Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). The upper boundary of the zone in the Upper Guandao section has been fixed slightly below the Bithynian-Pelsonian boundary. According to the conodont study, the Bithynian-Pelsonian boundary is loosely constrained because of the late occurrence of the Pelsonian index Paragondolella bulgarica and some taxonomic problems regarding Neogondolella ex gr. N. regalis. We have decided on the lowest level as the Bithynian-Pelsonian boundary, among the possible positions of the boundary suggested by Lehrmann et al. (Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015), considering also the stratigraphic position of characteristic Pelsonian foraminiferal associations.
Several major foraminiferal taxa with first occurrences in older zones occur also in the Bithynian: Hoyenella gr. H. sinensis, H. gr. H. shengi, Meandrospira pusilla, M. dinarica, Arenovidalina abriolense, Pilamminella grandis, Palaeolituonella reclinata, Earlandia tintinniformis, Endoteba bithynica, Spinoendotebanella lehrmanni n. gen., n. sp., Endotebanella kocaeliensis, Endotriadella wirzi, E. wirzi?, and Lamelliconinae?. The following taxa occur for the first time in the P. densa zone: Planiinvoluta? mesotriasica, Hoyenella? sp. 2, Ophthalmidium? Sp. 5, Reophax sp. 3, Endoteba controversa, Endotebanella sp. 2, Endotriada sp. 1, Protonodosaria exploita, Geinitzinita sp., Dentalina? sp. 1, and Dentalina? sp. 3.
The P. densa Zone has been used in previous studies to distinguish chronostratigraphic intervals within the Anisian Stage (Salaj, Reference Salaj1969, 1976; Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972b; Gazdzicki et al., Reference Gazdzicki, Trammer and Zwidzka1975; Zaninetti and Dağer, Reference Zaninetti and Dağer1978; Salaj et al., Reference Salaj, Borza and Samuel1983, Reference Salaj, Trifonova and Gheorghian1988; Orovecz-Scheffer, Reference Orovecz-Scheffer1987; Budurov et al., Reference Budurov, Calvet, Goy, Marquez-Aliaga, Marquez, Trifonova, Arche, Hagdorm and Seilacher1993; He, Reference He1993; Hardenbol et al., Reference Hardenbol, Thierry, Farley, Jacquin, de Graciansky, Vail, de Graciansky, Hardenbol, Jacquin and Vail1998; Kamoun et al., Reference Kamoun, Peybernès, Martini, Zaninetti, Vila, Trigui and Rigane1998; Faletti and Ivanova, Reference Faletti and Ivanova2003). In this study, the lowest occurrence of P. densa coincides with the Aegean-Bithynian boundary, which is consistent with the interpretation of Muttoni et al. (Reference Muttoni, Kent, Meço, Nicora, Gaetani, Balini, Germani and Rettori1996) based on conodont data. Therefore, this zone is a reliable index for the Bithynian substage, if its upper boundary is fixed with a reliable taxon such as A.? eotriasicus.
Aulotortus? eotriasicus Zone
Corresponding to the interval between the lowest and highest occurrences of Aulotortus? eotriasicus, the zone is ~51 m thick and measures from 55–106 meters in the Upper Guandao section (Figs. 2, 21; samples 23–45 in Fig. 8). Chronostratigraphically, it corresponds to the uppermost Bithynian and lower Pelsonian, and is composed primarily of limestone interrupted by one prominent allodapic breccia level. In the Pelsonian, A.? eotriasicus occurs above the last true Neogondolella regalis and is associated with the conodonts Nicoraella germanica and Ni. kockeli, extending from the Bithynian into the Pelsonian (Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). The last individuals of A.? eotriasicus occur below the first occurrence of the Pelsonian index conodont Paragondolella bulgarica, which is partially present in the Pelsonian of the Upper Guandao section (Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015).
Major foraminiferal taxa that occur in lower zones of the Anisian and range into the A.? eotriasicus Zone include Planiinvoluta? mesotriasica, Meandrospira dinarica, Pilammina densa, Pilamminella grandis, Palaeolituonella reclinata, Spinoendotebanella lehrmanni n. gen., n. sp., and Endotriadella wirzi. In addition, the following foraminiferal taxa appear for the first time within the A.? eotriasicus zone: Hoyenella? sp. 1, Glomospirella sp. 1, Reophax sp. 4, Reophax sp. 7, Endoteba obturata, Endotebanella sp. 1, Endotriada tyrrhenica, Endotriadella lombardi, Triadodiscus sp., Oberhauserella? sp.1, and Polarisella elabugae.
This zone, largely corresponding to the Pelsonian Substage, is surely present in the Dinarids (Bosnia-Herzegovina) and Greece (Brönnimann et al., Reference Brönnimann, Cadet and Zaninetti1973a; Rettori et al., Reference Rettori, Angiolini and Muttoni1994; Zaninetti et al., Reference Zaninetti, Rettori and Martini1994); however, no biozonation has been suggested in these studies. Muttoni et al. (Reference Muttoni, Kent, Meço, Balini, Nicora, Rettori, Gaetani and Krystyn1998) reported A.? eotriasicus from the base of the Illyrian of Albania, higher than its previously known range within the Anisian, and assigned this interval to the ‘pragsoides’ Zone, adopted from the A. pragsoides Zone of Salaj et al. (Reference Salaj, Trifonova and Gheorghian1988). However, the conodonts reported in their study are highly fragmentary and true Illyrian markers seem to be absent to calibrate this zone.
Aulotortus? eotriasicus-Meandrospira dinarica Zone
The zone is defined on the basis of successive last occurrences of Aulotortus? eotriasicus and Meandrospira dinarica. It is composed of a 95 m thick limestone interval interrupted by some important carbonate breccia levels between 106 and 201 meters of the Upper Guandao section (Figs. 2, 21; samples 47–65 in Fig. 8). The zone corresponds to the middle and upper part of the Pelsonian Substage, and the upper boundary nearly coincides with the last occurrence of the index conodont Paragondolella bulgarica, approximating the Pelsonian-Illyrian boundary.
Several major taxa, including M. dinarica, Pilammina densa, and Pilamminella grandis, range into this zone from underlying zones of the Anisian Stage. Among these taxa, only Spinoendotebanella lehrmanni n. gen., n. sp. makes its last appearance close to the lower boundary of the zone. In addition, the following foraminiferan species occur for the first time within the zone: Meandrospira? deformata, Meandrospiranella irregularis?, Turriglomina cf. T. magna, T. mesotriasica, Ophthalmidium sp. 3, Gsollbergella? sp. 2, Reophax sp. 1, Reophax sp. 6, Trochammina almtalensis, Malayspirina sp., Endoteba sp., Endotriada sp. 2, Plagiostomella sp., Diplotremina sp. 1, and Polarisella hoae.
Across the Tethys, the last occurrence of M. dinarica has been reported at or very close to the Pelsonian-Illyrian boundary (Zaninetti, Reference Zaninetti1976; Salaj et al., Reference Salaj, Trifonova and Gheorghian1988; Rettori, Reference Rettori1995; Hardenbol et al., Reference Hardenbol, Thierry, Farley, Jacquin, de Graciansky, Vail, de Graciansky, Hardenbol, Jacquin and Vail1998; Kamoun et al., Reference Kamoun, Peybernès, Martini, Zaninetti, Vila, Trigui and Rigane1998; Muttoni et al., Reference Muttoni, Kent, Meço, Balini, Nicora, Rettori, Gaetani and Krystyn1998). Therefore, using this last appearance as a boundary in the definition of the upper M. dinarica zone is very useful for the characterization of the Pelsonian substage in the Middle Triassic biostratigraphy.
Turriglomina mesotriasica Zone
Corresponding to an important part of the Illyrian Substage and composed of limestone intercalated with carbonate breccia levels, this zone measures 75 m between 201–276 meters of the Upper Guandao section. The zone is defined based on the last occurrence of Meandrospira dinarica and the first occurrence of Abriolina mediterranea. A third taxon, Turriglomina mesotriasica, has been used to name this zone. Its first occurrence is in the uppermost part of the underlying upper M. dinarica Zone, very close to the Pelsonian-Illyrian boundary (Figs. 2, 21; samples 67–89 in Fig. 8). The lower boundary of this zone coincides with the last occurrence of Pelsonian index conodont Paragondolella bulgarica and the first occurrence of P. excelsa, index conodont of the Illyrian (Lehrmann et al., Reference Lehrmann, Stepchinski, Altıner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani, Wang, Wei, Yu, Griffiths, Minzoni, Schall, Li, Meyer and Payne2015). The upper boundary lies at a datum lower than the Anisian-Ladinian boundary, which is delineated by the first occurrence of Budurovignathus truempyi, the index conodont for the base of Fassanian.
Some biostratigraphically important taxa, including Meandrospira pusilla, M. dinarica, Pilammina densa, and Pilamminella grandis, which first appear in the Olenekian and older Anisian zones, make their last appearances close to the Pelsonian-Illyrian boundary and do not extend into this zone. However, several taxonomically well-established taxa, including Planiinvoluta? mesotriasica, Arenovidalina abriolense, M.? deformata, Turriglomina cf. T. magna, Malayspirina sp., Verneuilinoides? azzouzi, Endoteba controversa, E. obturata, Endotebanella kocaeliensis, Endotriada tyrrhenica, Endotriadella wirzi, E. lombardi, and Krikoumbilica pileiformis, extend into this zone. The following species occur for the first time within the zone: Turriglomina sp., Endoteba badouxi, Endotriada sp. 3, and Piallina? sp.
Although Pilammina densa is the preferred foraminiferal marker of the upper Anisian in the European basins (Zaninetti et al., Reference Zaninetti, Brönnimann and Baud1972b, Salaj et al., Reference Salaj, Trifonova and Gheorghian1988; Rettori, Reference Rettori1995; Hardenbol et al., Reference Hardenbol, Thierry, Farley, Jacquin, de Graciansky, Vail, de Graciansky, Hardenbol, Jacquin and Vail1998), its stratigraphic range does not extend higher than Pelsonian in the Great Bank of Guizhou. Here we use the Turriglomina mesotriasica Zone to recognize the Illyrian Substage because it is well calibrated with conodont data.
Abriolina mediterranea Zone
In the upper Illyrian, this zone is defined on the basis of the successive first occurrences of Abriolina mediterranea and Agathammina-like forms (Agathammina? sp.) displaying coiling in five planes, as in Quinqueloculina, but without septation. The zone is represented by 18 m of limestone with the intercalation of one prominent breccia and measures between 276–294 meters of the Upper Guandao section (Figs. 2, 21; samples 91–95 in Fig. 8). The appearance of coiling in five planes in Agathammina? sp., never observed in the Anisian stratigraphy, is coeval with the first occurrence of the index conodont of the Fassanian, Budurovignathus truempyi.
Nearly all foraminiferal assemblages recognized in the underlying Turriglomina mesotriasica Zone are recorded in this zone. Abriolina? sp. and Palaeolituonella sp. occur for the first time.
Abriolina mediterranea is used for the first time as a zonal marker in this study. Although not formally defined, the zone is probably present in the Latemar platform (Dolomites, Italy), where A. mediterranea has been reported from Illyrian to Ladinian strata by Emmerich et al. (Reference Emmerich, Zamparelli, Bechstädt and Zühlke2005).
Early Triassic–early Middle Triassic diversity and recovery
Following one of the most severe mass extinctions in Earth's history, at the Permian-Triassic boundary of ~252 my ago, the recovery of marine ecosystems was both protracted and non-monotonic, with variation in recovery dynamics among higher taxa as well as interruptions of recovery due to later environmental disturbances (Hallam, Reference Hallam1991; Kidder and Worsley, Reference Kidder and Worsley2004; Payne et al., Reference Payne, Lehrmann, Wei, Orchard, Schrag and Knoll2004, Reference Payne, Lehrmann, Wei and Knoll2006; Lehrmann et al., Reference Lehrmann, Ramezani, Bowring, Martin, Montgomery, Enos, Payne, Orchard, Hongmei and Wei2006; Payne and Kump, Reference Payne and Kump2007; Fraiser and Bottjer, Reference Fraiser and Bottjer2007a; Galfetti et al., Reference Galfetti, Hochuli, Brayard, Bucher, Weissert and Vigran2007, Reference Galfetti, Bucher, Martini, Hochuli, Weissert, Crasquin-Soleau, Brayard, Goudmand, Brühwiler and Goudun2008; Knoll et al., Reference Knoll, Bambach, Payne, Pruss and Fischer2007; Algeo et al., Reference Algeo, Chen, Fraiser and Twitchet2011; Chen and Benton, Reference Chen and Benton2012; Sun et al., Reference Sun, Joachimski, Wignall, Yan, Chen, Jiang, Wang and Lai2012, Reference Sun, Wignall, Joachimski, Bond, Grasby, Sun, Yan, Wang, Chen and Lai2015; Song et al., Reference Song, Wignall, Chu, Tong, Sun, Song, Weihong and Tian2014; Tian et al., Reference Tian, Tong, Algeo, Song, Song, Chu, Shi and Bottjer2014; Wei et al., Reference Wei, Shen, Scheopfer, Krystyn, Richoz and Algeo2015; Lau et al., Reference Lau, Maher, Altıner, Kelley, Kump, Lehrmann, Silva-Tamayo, Weaver, Yu and Payne2016; Penn et al., Reference Penn, Deutsch, Payne and Sperling2018; Zhang et al., Reference Zhang, Romaniello, Algeo, Lau, Clapham, Richoz, Hermann, Smith, Horacek and Anbar2018; Goudemand et al., Reference Goudemand, Romano, Leu, Bucher, Trotter and Williams2019). The combined effects of extinction and recovery led to replacement of the Paleozoic Fauna by the Modern Fauna (Sepkoski, Reference Sepkoski1984; Fraiser and Bottjer, Reference Fraiser and Bottjer2007b). In addition to foraminifera, recovery dynamics have been studied in microfossil groups such as radiolarians (Feng et al., Reference Feng, He, Gu, Meng, Jin and Zhang2007; Feng and Algeo, Reference Feng and Algeo2014) and conodonts (Orchard, Reference Orchard2007; Stanley, Reference Stanley2009), as well as invertebrate clades such as corals (Senowbari-Daryan et al., Reference Senowbari-Daryan, Zühlke, Bachtädt and Flügel1993; Flügel, Reference Flügel2002), brachiopods (Rodland and Bottjer, Reference Rodland and Bottjer2001; Z.Q. Chen et al., Reference Chen, Kailho and George2005; J. Chen et al., Reference Chen, Tong, Song, Luo, Huang and Xiang2015), bryozoans (Powers and Pachut, Reference Powers and Pachut2008), gastropods (Erwin and Pan, Reference Erwin, Pan and Hart1996; Fraiser and Bottjer, Reference Fraiser and Bottjer2004; Nützel, Reference Nützel2005; Payne, Reference Payne2005; Brayard et al., Reference Brayard, Nützel, Stephen, Bylund, Jenks and Bucher2010), ammonoids (McGowan, Reference McGowan2004; Brayard et al., Reference Brayard, Bucher, Escarguel, Fluteau, Bourquin and Galfetti2006, Reference Brayard, Escarguel, Bucher, Monnet, Brüchwiler, Goudemand, Galfetti and Guex2009; Stanley, Reference Stanley2009), and echinoderms and vertebrates (Twitchett and Oji, Reference Twitchett and Oji2005; Chen and McNamara, Reference Chen and McNamara2006), yielding valuable information on recovery patterns during the Early Triassic and their correlation with ongoing environmental disturbances.
The foraminiferal fauna of the Great Bank of Guizhou was studied recently by Payne et al. (Reference Payne, Summers, Rego, Altıner, Wei, Yu and Lehrmann2011) to investigate the tempo and mode of biotic recovery from the end-Permian mass extinction. They reported gradual increases in the diversity, evenness, and sizes of foraminifera through the Early Triassic and earliest Middle Triassic, with stable values reached early in the Anisian. Payne et al. (Reference Payne, Summers, Rego, Altıner, Wei, Yu and Lehrmann2011) stated that the recovery patterns on the GBG correspond well with available global data and appear to parallel those of many benthic invertebrate clades. Based on the metrics of standing diversity, evenness, and size, the recovery trajectory in foraminifera appears to have been more gradual than those of pelagic taxa, such as ammonoids and conodonts (Brayard et al., Reference Brayard, Escarguel, Bucher, Monnet, Brüchwiler, Goudemand, Galfetti and Guex2009; Stanley Reference Stanley2009), suggesting a decoupling of recovery processes between benthic and pelagic environments. However, these metrics do not capture species-level changes in the composition of the foraminiferal biota and their implications for recovery dynamics. Consequently, some similarities between the foraminiferan and pelagic recovery dynamics may have been overlooked.
In this study, a more detailed, species-level taxonomy has been carried out on the material of the GBG used by Payne et al. (Reference Payne, Summers, Rego, Altıner, Wei, Yu and Lehrmann2011), which has revealed further details on the recovery pattern and diversity in the Early Triassic. The species-level data shed new light on recovery dynamics and, furthermore, highlight the ways in which analyses at higher taxonomic levels may capture some aspects of recovery dynamics, but also obscure or distort other aspects.
Ninety-seven foraminifera populations recognized at species rank have been plotted in a range chart from the base of Griesbachian (Permian-Triassic boundary) to the upper limit of the Illyrian (Anisian-Ladinian boundary) (Fig. 22). In actuality, the number of distinct, species-level taxa is probably >100 because four of these taxa given in the list of Figure 22 (Variostomatidae, Endotebidae, unknown Nodosariata, and Lamelliconinae?) probably contain more than one taxon. Within the foraminiferal biostratigraphic framework, calibrated by conodont biostratigraphy, we recognize three groups of taxa whose stratigraphic ranges and phyletic relationships to one another demarcate three major phases in the Early–Middle Triassic recovery of foraminiferan communities.

Figure 22. Range chart showing the delayed and progressive recovery phases of foraminifera. For the abbreviations of biozone markers of foraminifera, see Figure 21. 1 = Postcladella kalhori; 2 = Earlandia sp. 1; 3 = Unknown foraminifera 4; 4 = Earlandia sp. 2; 5 = Postcladella grandis; 6 = Cornuspira mahajeri?; 7 = Hoyenella gr. H. sinensis; 8 = Hoyenella gr. H. shengi; 9 = Pseudoammodiscus sp.; 10 = Glomospira sp.; 11 = Unknown foraminifera 1; 12 = Glomospirella vulgaris; 13 = Arenovidalina weii; 14 = Transitional forms to Meandrospira; 15 = Praetriadodiscus zaninettiae; 16 = Praetriadodiscus tappanae; 17 = Meandrospira pusilla; 18 = Endotebanella kocaeliensis; 19 = Meandrospira cheni; 20 = Krikoumbilica pileiformis; 21 = Trochammina? sp. 1; 22 = Nodosinelloides? sp.; 23 = Meandrospira? enosi n. sp.; 24 = Variostoma sp. 1; 25 = Variostoma sp. 2; 26 = Verneuilinoides? azzouzi; 27 = Endoteba bithynica; 28 = Diplotremina sp. 2; 29 = Endotriadella wirzi; 30 = Hoyenella ? sp.; 31 = Variostomatidae; 32 = Endotebidae; 33 = Meandrospira? sp.; 34 = Meandrospira dinarica; 35 = Endotriadella wirzi?; 36 = Pilamminella grandis; 37 = Unknown Nodosariata; 38 = Reophax sp. 2; 39 = Glomospirella sp. 2; 40 = Earlandia tintinniformis; 41 = Gaudryina sp.; 42 = Placopsilina sp.; 43 = Tolypammina gregaria; 44 = Unknown foraminifera 2; 45 = Reophax sp. 5; 46 = Spinoendotebanella lehrmanni n. gen., n. sp.; 47 = Arenovidalina abriolense; 48 = Lamelliconinae ?; 49 = Protonodosaria exploita; 50 = Meandrospiranella samueli; 51 = Palaeolituonella reclinata; 52 = Pilammina densa; 53 = Triadodiscus sp.; 54 = Endoteba controversa; 55 = Ophthalmidium? sp. 5; 56 = Planiinvoluta? mesotriasica; 57 = Geinitzinita sp.; 58 = Reophax sp. 3; 59 = Dentalina? sp. 1; 60 = Hoyenella? sp. 2; 61 = Dentalina? sp. 2; 62 = Endotebanella sp. 2; 63 = Endotriada sp. 1; 64 = Aulotortus? eotriasicus; 65 = Hoyenella ? sp. 1; 66 = Glomospirella sp. 1; 67 = Endotriadella lombardi; 68 = Endoteba obturata; 69 = Endotebanella sp. 1; 70 = Reophax sp. 4; 71 = Oberhauserella? sp.; 72 = Polarisella elabugae; 73 = Endotriada tyrrhenica; 74 = Reophax sp. 7; 75 = Ophthalmidium sp. 3; 76 = Gsollbergella? sp. 2; 77 = Endoteba sp.; 78 = Endotriada sp. 2; 79 = Meandrospira? deformata; 80 = Reophax sp. 6; 81 = Trochammina almtalensis; 82 = Turriglomina cf. T. magna; 83 = Polarisella hoae; 84 = Gandinella sp. 1; 85 = Meandrospiranella irregularis?; 86 = Reophax sp. 1; 87 = Turriglomina mesotriasica; 88 = Malayspirina sp.; 89 = Plagiostomella? sp.; 90 = Diplotremina sp. 1; 91 = Endoteba badouxi; 92 = Turriglomina sp.; 93 = Endotriada sp. 3; 94 = Piallina? sp.; 95 = Abriolina? sp.; 96 = Abriolina mediterranea; 97 = Palaeolituonella sp.
Step1: Griesbachian–early Dienerian: disaster taxa and their evolutionary derivatives
The foraminiferan community that characterizes the interval from the end-Permian extinction horizon through the Griesbachian and into the mid-Dienerian has low diversity and a distinctive taxonomic composition (Fig. 22). The community largely belongs to three genera: Postcladella (previously known as Rectocornuspira), Cornuspira, and Earlandia. The later appearance of P. grandis, which most likely evolved from P. kalhori, illustrates that the community was not entirely static; rather, it contains evidence of evolutionary trends that may reflect recovery from the mass extinction event or some amelioration of environmental constraints on recovery.
Defined by Fischer and Arthur (Reference Fischer, Arthur, Cook and Enos1977) as opportunistic species that proliferated at times of biotic crises, ‘disaster taxa’ have been further described by Kauffman and Harries (Reference Kauffman, Harries and Hart1996) as species that undergo very short-term, large-scale population blooms immediately following biotic crises, early in the survival phase. The studied samples of the Great Bank of Guizhou reveal a distinct pattern of low-diversity, high-dominance foraminiferal assemblages that persist through the Griesbachian before disappearing by the end of the early Dienerian (Fig. 23). This long interval of dominance by a few species may explain why it is also associated with internal evolutionary trends, such as the increase in size from P. kalhori to P. grandis. However, the lineages that dominated this early post-extinction interval do not appear to have played a role in the later Early Triassic recovery of foraminifera.
Figure 23. Disaster taxa from Dawen (PDW) and Dajiang (PDJ) sections. (1) Postcladella (Pk: P. kalhori; Pg: P. grandis) from the Postcladella grandis Zone (Griesbachian) (PDJ-174); (2) Postcladella (Pk: P. kalhori) from the Postcladella kalhori-Postcladella grandis Zone (Griesbachian) (PDW-011); (3) Earlandia (E: E. sp. 2) associated with Postcladella (Pk: P. kalhori; Pg: P. grandis) from the Postcladella grandis Zone (Griesbachian) (PDW-120); (4) Earlandia (E: E. sp. 2) associated with Postcladella (Pk: P. kalhori) from the Postcladella grandis Zone (Griesbachian) (PDW-120). Horizontal bars = 200 μm.
Similar Griesbachian to lowermost Dienerian foraminiferan assemblages have been reported across the Tethyan belt. Hallam and Wignall (Reference Hallam and Wignall1997), based mainly on data from south China, were the first to characterize an Early Triassic foraminiferan genus, Earlandia spp., explicitly as a disaster taxon. Later, Altıner et al. (Reference Altıner, Groves and Özkan-Altıner2005), Groves and Altıner (Reference Groves and Altıner2005), and Groves et al. (Reference Groves, Altıner and Rettori2005, Reference Groves, Rettori, Payne, Boyce and Altıner2007) characterized a group of species, consisting of P. kalhori, C. mahajeri, and Earlandia spp., as disaster taxa that make their first appearances in the thrombolitic facies directly overlying the Changhsingian carbonates, both in Turkey and Italy. Later, Song et al. (Reference Song, Tong, Chen, Yang and Wang2009, Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016), from south China, and Krainer and Vachard (Reference Krainer and Vachard2011), from southern Austria, described similar taxa from similar facies in lowermost Triassic strata. According to Altıner et al. (Reference Altıner, Groves and Özkan-Altıner2005), this assemblage of disaster taxa is usually associated in the early Griesbachian with several well-known Permian forms that survived the end-Permian mass extinction. These surviving taxa, including Globivalvulina, Kamurana, Geinitzina, Syzrania, and maybe several others, survived the immediate extinction interval, but did not leave any further descendants in the Triassic as part of the long-term recovery process (‘failed crisis progenitors’ of Kauffman and Harries, Reference Kauffman, Harries and Hart1996). In a more recent paper by Song et al. (Reference Song, Tong, Wignall, Luo, Tian, Song, Huang and Chu2016), a distinct population of Globivalvulina (G. lukachiensis Nestell et al., Reference Nestell, Kolar-Jurkovšek, Jurkovšek and Aljinović2011), known from the Capitanian–Changhsingian stages of the Permian, has been erroneously classified as a disaster taxon from the lower Griesbachian of south China. This taxon is more likely a failed survivor (Altıner et al., Reference Altıner, Groves and Özkan-Altıner2005).
Assessing the evolutionary paleoecological dynamics of foraminifera during the immediate survival interval of the Griesbachian is further complicated by highly unusual foraminiferal taxa reported without any illustration in the study of Sun et al. (Reference Sun, Wignall, Joachimski, Bond, Grasby, Sun, Yan, Wang, Chen and Lai2015) from the Smithian–Spathian sections of south China. These authors reported the typical Griesbachian–early Dienerian disaster taxa Postcladella (given as Rectocornuspira) and Earlandia, as well as some Permian forms (Paleotextularia, Cribrogenerina, Globivalvulina, Nodosinella), from Smithian–Spathian strata in association with Meandrospira pusilla and Hoyenella. If this report does not reflect mistakes in taxonomic identifications, the mixed fauna most probably could be explained by reworking of Permian and Griesbachian–Dienerian specimens into Smithian–Spathian strata of the Jiarong and Mingtang sections of the Nanpanjiang Basin. These sections contain platform material that was transported and redeposited in slope sections, providing a potential mechanism for this scenario. The co-occurrences of taxa in these sections that have not been reported to co-occur elsewhere argue against an interpretation of the local survival of the Permian and Griesbachian forms into the Smithian and Spathian.
The stratigraphic range of the high-dominance, low-diversity foraminiferan assemblage extending from the base of Griesbachian to mid-Dienerian coincides in time with the strongest geochemical proxy evidence for persistent and expanded ocean anoxia (cf., Lau et al., Reference Lau, Maher, Altıner, Kelley, Kump, Lehrmann, Silva-Tamayo, Weaver, Yu and Payne2016). According to Zhang et al. (Reference Zhang, Romaniello, Algeo, Lau, Clapham, Richoz, Hermann, Smith, Horacek and Anbar2018), two successive expansions of anoxia occurred on the continental shelves within this time interval—the earlier event lasted from latest Changhsingian to earliest Griesbachian (C1), and the later event lasted from latest Griesbachian to earliest Dienerian (C2). The earlier event coincides with the range and extinction of failed survivors and with the first appearances of disaster taxa immediately following the end-Permian extinction. The second pulse corresponds to the continuation of the survival of disaster taxa from latest Griesbachian to earliest Dienerian, and finally to their extinction.
Step 2: Late Dienerian–Smithian: Lazarus taxa and their evolutionary derivatives
Following a mid-Dienerian gap in the occurrence of foraminifera on the GBG in the studied sections, there is a substantial shift in the composition of foraminiferan communities and recovery dynamics during the late Dienerian to Smithian (Fig. 22). Most of the taxa that occur in this step are Lazarus taxa with Permian ancestors, but no obvious representatives in Griesbachian to lower Dienerian strata (e.g., Hoyenella spp., Pseudoammodiscus sp., Glomospira sp. [=Pilammina praedensa?], Glomospirella vulgaris, Arenovidalina weii n. sp., transitional forms to Meandrospira). Hoyenella and Arenovidalina originated in the late Paleozoic (Vachard, Reference Vachard, Lucas and Shen2018), but they reappeared in the late Dienerian and in the Smithian, respectively. Glomospira and Glomospirella also originated in the late Paleozoic (Permian) (Vachard, Reference Vachard, Lucas and Shen2018), and then reappeared in the late Dienerian–Smithian interval. The forms illustrated as Glomospirella? from the uppermost Changhsingian of southwestern Guangxi, south China by Gu et al. (Reference Gu, Feng and He2007) are certainly related to Early Triassic Glomospirella faunas. Glomospirella? mamilla Gu et al., Reference Gu, Feng and He2007, which is similar or identical to Glomospirella vulgaris, is recognized as a Lazarus taxon in this study in the Smithian of the GBG. Apparently, such Glomospirella species of Guangxi adapted themselves to deeper-water environments (as a refugium?) close to the Permian-Triassic boundary and later, in the aftermath of the crisis, spread over the carbonate platform.
The pseudoammodiscid clade, which appeared in the Devonian and survived through the Permian, reappeared in the Early Triassic (Altıner and Payne, Reference Altıner and Payne2017). Pseudoammodiscus sp. first occurs in the upper Dienerian in China, as well as in Turkey (Altıner and Zaninetti, Reference Altıner and Zaninetti1981). This form apparently survived into the Smithian and finally gave rise to Praetriadodiscus close to the Smithian-Spathian boundary. Praetriadodiscus is an ancestor or close relative of the last common ancestor of all involutinid taxa (Altıner and Payne, Reference Altıner and Payne2017). The genus Meandrospira, which occurs in the Spathian, has been interpreted as a Lazarus taxon representing the phyletic continuation of the Permian form assigned to Streblospira (Altıner et al., Reference Altıner, Groves and Özkan-Altıner2005). The specimens that we found in the Smithian of the Dajiang section probably belong to this Lazarus lineage.
Within the detailed biostratigraphic framework based on both conodonts and foraminifera, it appears that the recovery of Lazarus taxa in the late Dienerian–Smithian interval was interrupted by an extinction event. Several foraminiferal Lazarus taxa (Pseudoammodiscus sp., Arenovidalina weii n. sp., Glomospirella vulgaris, Praetriadodiscus zaninettiae, and P. tappanae) disappeared close to the Smithian-Spathian boundary (Figs. 21, 22). However, from these Lazarus taxa, the main species of the genus Hoyenella (H. gr. H. sinensis and H. gr. H. shengi) and the unknown foraminifera 1 survived and occur in the Spathian and Middle Triassic Series of the Great Bank of Guizhou.
The data from this study are not consistent with the data and interpretations given in Song et al. (Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan, Niu, Chen, Yang and Wang2011a), in which it was suggested that benthic marine diversity began to recover in the early Smithian, little more than 1 Myr after the mass extinction. We interpret the coincidence of the first occurrences of several foraminiferal taxa in the Smithian in the dataset of Song et al. (Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan, Niu, Chen, Yang and Wang2011a) to result either from taxonomic misassignments, which are impossible to assess due to the lack of illustrated material, or due to miscorrelation of studied sections. In Song et al. (Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan, Niu, Chen, Yang and Wang2011a), several taxa that are well known from younger levels of the Lower Triassic, Anisian, or even from the Late Triassic (e.g., Meandrospira dinarica, Pilammina densa, Trochammina almtalensis, Endotriadella wirzi, and Gsollbergella spiroloculiformis [Oravecz-Scheffer, Reference Oravecz-Scheffer1971]) are reported in the early Smithian. Because we did not observe these forms in our material, and they have not been reported from correlative stratigraphic horizons elsewhere, we cannot place strong weight on these reports.
An episode of expanded oceanic anoxia during the mid-late Smithian (the C3 event of Zhang et al., Reference Zhang, Romaniello, Algeo, Lau, Clapham, Richoz, Hermann, Smith, Horacek and Anbar2018) is correlated broadly with the extinction of several important Lazarus taxa. However, further assessment is required to determine how closely spaced in time these extinctions were and how closely they coincide to intervals of rapid environmental change, such as the Smithian-Spathian boundary crisis (Galfetti et al., Reference Galfetti, Hochuli, Brayard, Bucher, Weissert and Vigran2007).
Step 3: Spathian–Anisian: progressive recovery
The interval spanning from the base of the Spathian to the top of the Illyrian is characterized by the marked diversification of foraminifera (Fig. 22). Among the 80 taxa identified within this interval in this study, short-ranging taxa are generally populations defined with open nomenclature (Reophax spp. 1–7; Ophthalmidium spp. 1–5; etc.), based on our latest taxonomic interpretations (see taxonomy section). In the Spathian–Anisian interval, these taxa appeared in association with several well-established and wider ranging forms, such as Meandrospira pusilla, M. dinarica, Pilammina densa, and Turriglomina mesotriasica.
Several clear trends have been noted in the evolution of foraminifera during the Spathian. Three distinct populations, M. pusilla, M. cheni, and M.? enosi n. sp., were successively derived from the Lazarus taxon Meandrospira (Altıner et al., Reference Altıner, Groves and Özkan-Altıner2005). The Permian genus Endoteba reappeared in the Spathian and gave way to the first occurrences of E. bithynica, Endotebanella kocaeliensis, and Endotriadella wirzi in this interval. Species belonging to the well-known Triassic families Variostomatidae and Trochamminidae also made their first appearances in the Spathian.
The rest of the Anisian diversity increase continued at a fairly constant rate, within the constraints of available data, although there may have been an acceleration close to the Spathian-Aegean boundary (Fig. 22). Within this context of progressive recovery, the C4 event of Zhang et al. (Reference Zhang, Romaniello, Algeo, Lau, Clapham, Richoz, Hermann, Smith, Horacek and Anbar2018), which corresponds to expansion of sea-floor anoxia at the Early-Middle Triassic transition, does not seem to have been associated with any obviously biological response in foraminiferan communities on the GBG. Although Zhang et al. (Reference Zhang, Romaniello, Algeo, Lau, Clapham, Richoz, Hermann, Smith, Horacek and Anbar2018) recognized this end-Spathian event as a crisis of global extent, there is relatively little evidence at present for a large biological response in benthic marine communities.
Many of the foraminiferal populations recognized in the Anisian derived from ancestors that originated in the Olenekian (Meandrospira dinarica from M. cheni, species of Turriglomina and other meandrospirid forms from M. pusilla, derivation of Ophthalmidium spp. from Arenovidalina originated in the Smithian, Spinoendotebanella lehrmanni n. gen., n. sp. from E. kocaeliensis, Endotriadella wirzi? [large form] from E. wirzi, Trochammina almtalensis from the lineage originated from Trochammina sp. 1). Other species have obvious evolutionary precursors in the Anisian (progressively derived Endoteba, Endotebanella, Endotriada, and Endotriadella species). However, some other taxa also occur during this phase of recovery whose phylogenetic origins cannot be demonstrated with the available material from China. For example, Pilammina praedensa, a possible ancestor of P. densa and Pilamminella grandis, has been recorded only questionably in the Smithian (as Glomospira sp.). For the origin of Reophax, which first appears at the base of Anisian, no specimens have been recovered in the Lower Triassic. In addition to these taxa, the origins of some taxa first occurring in the Anisian (Palaeolituonella reclinata, Aulotortus? eotriasicus, Glomospirella sp. 1, G. sp. 2 [Glomospirella lampangensis?], Gaudryina sp. [Gaudryina triadica?]) remain unclear and are not well constrained by the studied material from the GBG.
In summary, foraminiferal standing diversity increased progressively through the late Early Triassic (Spathian) and into the Middle Triassic (Anisian). The maximum number of species encountered is nearly 20 in the Spathian, ~40 in the Aegean/Bithynian, and 60 in the Pelsonian, whereas in the Griesbachian–Smithian, the diversity remained always <10. Following the extinction of far more than 100 calcareous foraminiferan species in the Changhsingian (Groves and Altıner, Reference Groves and Altıner2005), the Griesbachian to Smithian interval is characterized by two successive stages of recovery. The first is characterized by the high abundance of a few disaster taxa, with many failed survivors (Altıner et al., Reference Altıner, Groves and Özkan-Altıner2005; Groves et al., Reference Groves, Altıner and Rettori2005; Song et al., Reference Song, Tong, Chen, Yang and Wang2009) during the Griesbachian to early Dienerian. The second consists of a fauna dominated by Lazarus taxa in the late Dienerian–Smithian. The taxa that dominated these two intervals produced few lineages that participated in the long-term recovery process. The diversification of taxa that contributed to the Middle Triassic diversification of foraminifera started in the Spathian with several new originations, and continued with these taxa becoming ancestors of many species that made their first appearances in the Anisian.
Acknowledgments
We thank A. Bush, R. Kodner, J. Wei, J. Xiao, and H. Yao for assistance in the field. We thank R. Martini, J. Jin, B. Huber, and two anonymous reviewers for comments that greatly improved the quality and clarity of the manuscript. This study was partially supported by a grant from the U.S. National Science Foundation (EAR-0807377-007 to J.L.P.).























