


Introduction
The conodont species Icriodus orri was erected by Klapper and Barrick (Reference Klapper and Barrick1983) for P1 elements differing from I. retrodepressus Bultynck, Reference Bultynck1970 by a wider posterior part of the basal cavity. Its large intraspecific variability was indicated by the authors by defining two morphotypes. The illustrated material was obtained from the lower part of the Spillville Formation exposed in Iowa and southern Minnesota (USA). The age of the studied sections was estimated approximately as the kockelianus Zone of the upper Eifelian.
Preliminary study of the original material included in Icriodus orri confirms its morphological inhomogeneity. The P1 elements differ not only in the presence or absence of the depression but also in the outline of the spindle, course of its axis, size, and shape of the denticles. Even the size and shape of the basal cavity, important criteria for distinction of the species from I. retrodepressus, are variable. This suggests that I. orri sensu Klapper and Barrick (Reference Klapper and Barrick1983) comprises forms that can be attributed to more than a single species.
The aim of the present study is a taxonomic revision of the material presented by Klapper and Barrick (Reference Klapper and Barrick1983), aided by the authors’ own collections from the Michigan Basin (USA) and Pripyat Graben (Belarus), together with relevant published data. In addition to morphological features, stratigraphical ranges and geographic distributions are also discussed in detail.
Studied materials and their regional background
The Belarussian material was obtained from the drill cores from the wells Zhitkovichi 2 and Pinsk 54 situated in the western part of the Pripyat Graben (Fig. 1; Golubtsov et al., Reference Golubtsov, Kruchek, Obukchovskaya and Pushkin2000, fig. 1; Narkiewicz and Kruchek, Reference Narkiewicz and Kruchek2008, fig. 2). During the Middle Devonian, this area formed a part of the intracontinental basin with a prevailing terrestrial and restricted shallow-marine sedimentation in a tropical climatic belt of the eastern part of the Euramerican continent. Studied material came from the basal parts of the dolomitic-shaly succession of the Kostyukovichi Horizon (Golubtsov et al., Reference Golubtsov, Kruchek, Obukchovskaya and Pushkin2000, fig. 2; Kruchek et al., Reference Kruchek, Pushkin and Obuchovskaya2001, fig. 5.23). This lithostratigraphic unit represents an important correlative level due to its unique open-marine aspect contrasting with enclosing terrigeneous nearshore and/or terrestrial facies. Numerous fossil remains, including conodonts, fishes, and miospores, permit correlation of this horizon with the ensensis Zone of the uppermost Eifelian (Narkiewicz and Kruchek, Reference Narkiewicz and Kruchek2008, p. 190).

Figure 1 Eifelian palaeogeographic map of Euramerica and northern Gondwana (after Scotese, Reference Scotese2002) showing locations of the studied Eifelian conodont fauna. Approximate location of Transcaucasia after Tolokonnikova and Ernst (Reference Tolokonnikova and Ernst2010).
The collection from the Michigan Basin (USA) was obtained from the United States Steel Corporation Quarry located southeast of Rogers City, Michigan, USA (Orr, Reference Orr1971, p. 105, section 15-IU10711; Bultynck, Reference Bultynck1976, p. 134, fig. 2, Locality 1). The Middle Devonian Michigan Basin was an intracontinental shallow-marine basin near the southwestern margin of Euramerica in a tropical paleoclimatic zone. The samples were taken in 1972 from the Dundee and Rogers City formations, and from the lowermost part of the Bell Shale Formation. The distribution and frequency of particular taxa was given by Bultynck (Reference Bultynck1976, table 1) without, however, any photographic documentation. Some specimens from the Dundee Formation, including two identified as Icriodus orri, were illustrated in a later paper by Bultynck (Reference Bultynck2003, pl. 2, figs. 34, 35).
The studied material, illustrated in Figures 2–4, is generally well preserved. In the Belarussian collection, small, most probably juvenile, specimens prevail. The specimens have smooth lustrous surfaces and their amber color points to a low thermal maturity (conodont CAI 1). The Michigan Basin specimens display a porous and rough surface texture that could indicate activity of subsurface fluids. The conodont CAI attains a value between 2.5 for medium-sized specimens and 3 for the large ones.
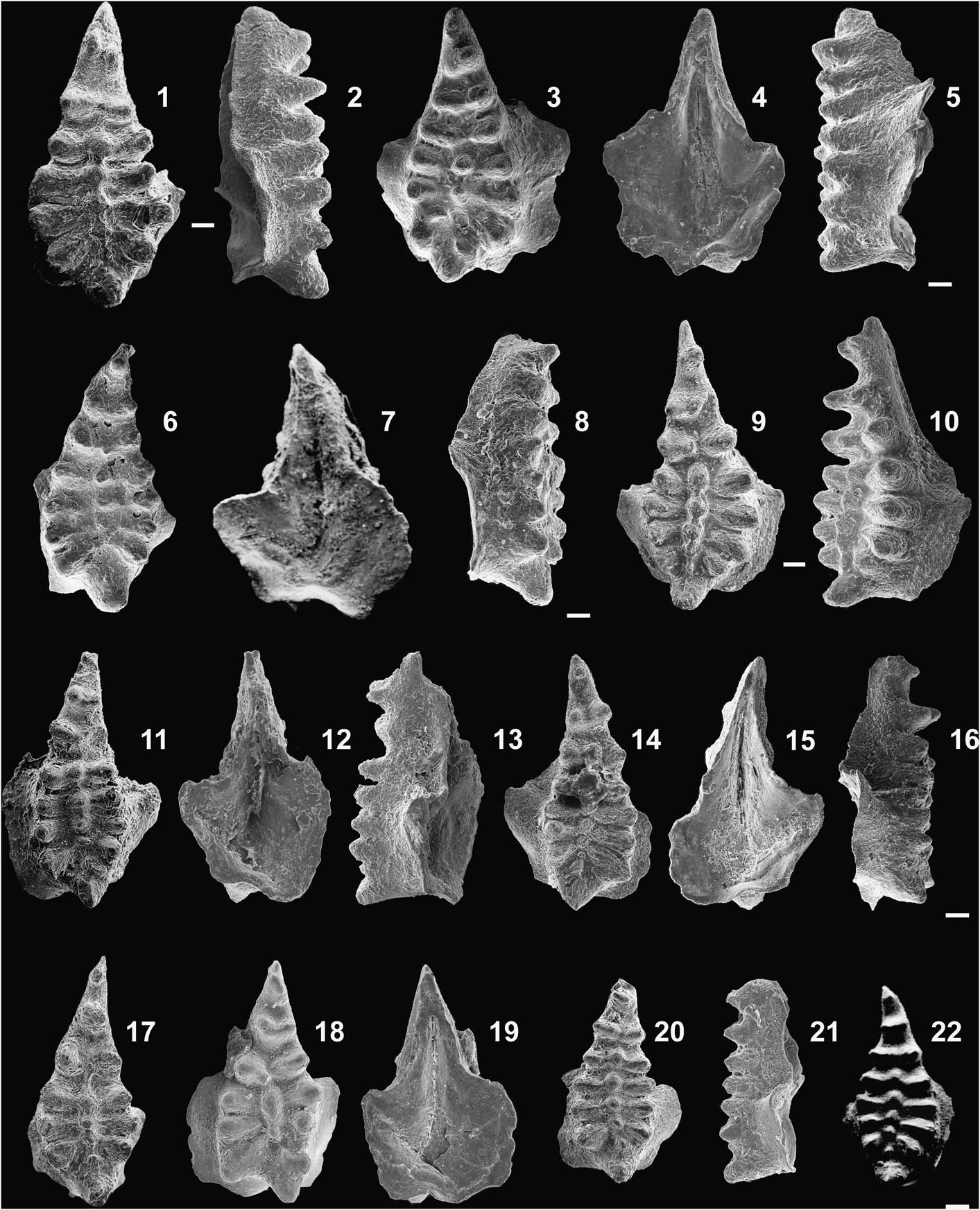
Figure 2 Conodont P1 elements of Icriodus michiganus n. sp. from the Dundee Formation, the U.S. Steel Corporation Quarry section, Michigan Basin (USA). (1, 2) Upper and lateral views, MUZ PIG 1795.II.4, sample 29. (3–5) Upper, lower, and lateral (broken anterior tip of the spindle) views of holotype, MUZ PIG 1795.II.1, sample 23. (6–8) Upper, lower, and lateral views of paratype, MUZ PIG 1795.II.2, top of the Dundee Formation, sample 01. (9, 10) Upper and lateral views, MUZ PIG 1795.II.5, sample 28. (11–13) Upper, lower, and lateral views, MUZ PIG 1795.II.6, sample 18. (14–16) Upper, lower, and lateral views of paratype, MUZ PIG 1795.II.3, sample 21. (17) Upper view, MUZ PIG 1795.II.7, sample 23. (18, 19) Upper and lower views of paratype, MUZ PIG 1795.II.8, top of the Dundee Formation, sample 01. (20, 21) Upper and lateral views, MUZ PIG 1795.II.9, sample 31. (22) Upper view, MUZ PIG 1795.II.10, sample 29. Scale bars=100 μm.

Figure 3 Conodont P1 elements of Icriodus orri Klapper and Barrick, Reference Klapper and Barrick1983. (1−3) Reillustrations of figures by Klapper and Barrick (Reference Klapper and Barrick1983, fig. 9.AF–AG,Z used with permission from SEPM): (1, 2) holotype, upper and lower views (fig. 9.AF–AG); (3) paratype, upper view (fig. 9.Z), Racine section, samples 3 and 3A, respectively, lower part of the Spillville Formation, southern Minnesota (USA). (4–11) Kostyukovichi Horizon, Pripyat Graben, Belarus; (4) upper view, MUZ PIG 1804.II.1, Pinsk 54, depth 465.0−460.0 m, sample 95w; (5) upper view, MUZ PIG 1804.II.2, Pinsk 54, depth 465.0−460.0 m, sample 95b; (6, 7) upper and lower views, MUZ PIG 1804.II.3, Zhytkovichi 2, depth 191.0−186.6 m, sample 6e; (8, 9) upper and lateral views, MUZ PIG 1804.II.4, Pinsk 54, depth 465.0−460.0 m, sample 95b; (10, 11) upper and lateral views, MUZ PIG 1804.II.5, Zhytkovichi 2, depth 191.0−186.6 m, sample 5a. (12–16) U.S. Steel Corporation Quarry section, Michigan Basin, USA., Bell Shale Formation, sample 16; (12) upper view, MUZ PIG 1795.II.11; (13, 14) upper and lateral views, MUZ PIG 1795.II.12; (15, 16) upper and lateral views, MUZ PIG 1795.II.13. Scale bars=100 μm.
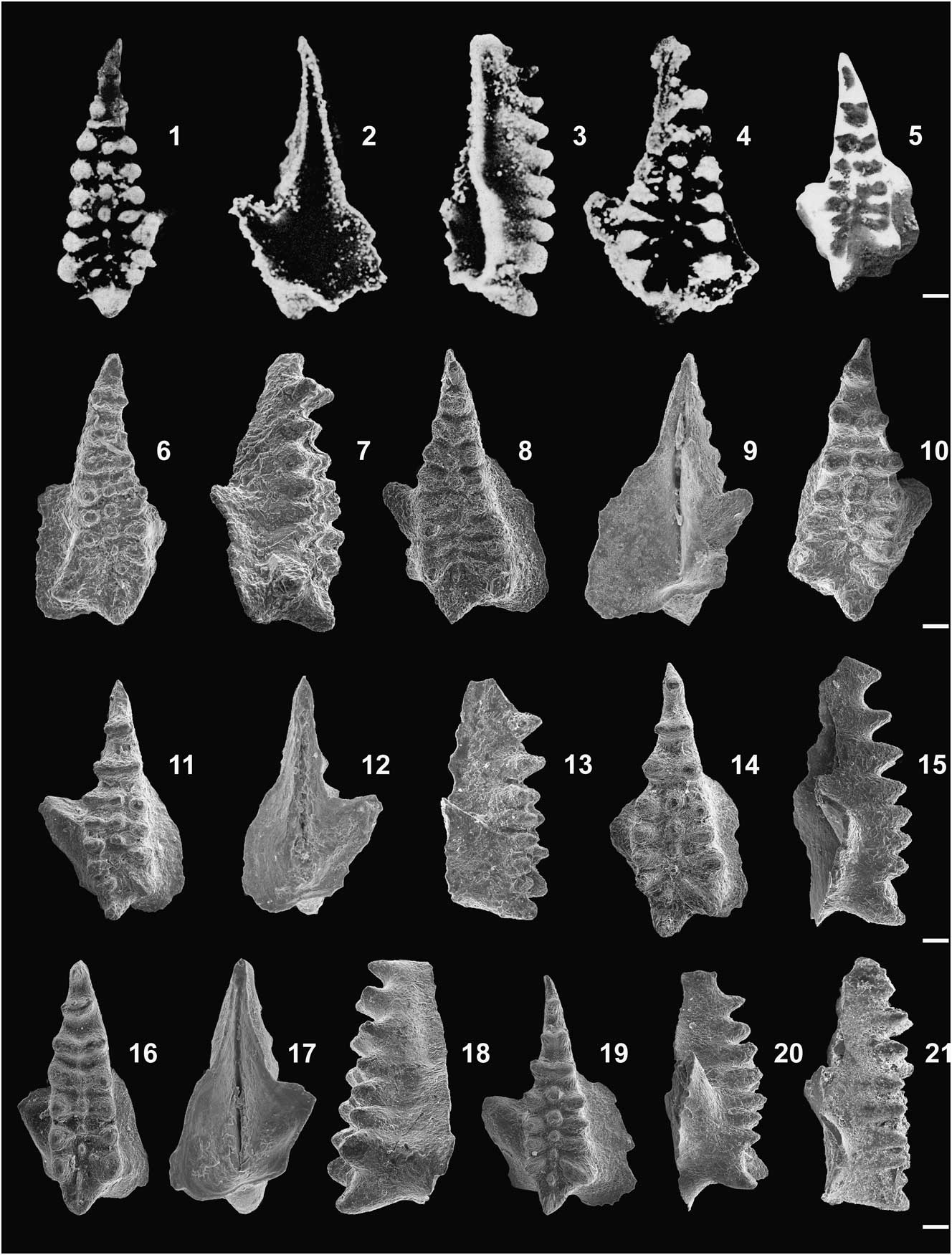
Figure 4 (1–20) Conodont P1 elements of Icriodus retrodepressus Bultynck, Reference Bultynck1970; (1–4) Reillustrations of figures by Bultynck (Reference Bultynck1970; reproduced with permission); (1–3) holotype (Bultynck, Reference Bultynck1970, pl. 30, fig. 1), upper, lower, and lateral views, Eau Noire section, Ardenne, Belgium, Couvin Formation, sample 75/6; (4) upper view (Bultynck, Reference Bultynck1970, pl. 30, fig. 3), Eau Noire section, sample 62/2. (5) Reillustration of figures by Werner and Ziegler (Reference Werner and Ziegler1982, pl. 1, fig. 27, used with permission from Senckenberg Forschngsinstitut und Naturmuseum Frankfurt), upper view, Wetteldorf Richtschnitt section, assemblage V, WP. 72, Lauch Formation southern Eifelian Hills, Germany. (6−20) Elements from U.S. Steel Corporation Quarry section, Michigan (USA), Dundee Formation; (6, 7) upper and lateral views, MUZ PIG 1795.II.14, sample 17; (8, 9) upper and lower views, MUZ PIG 1795.II.15, sample 30; (10) upper view, MUZ PIG 1795.II.16, sample 28; (11–13) upper, lower, and lateral views, MUZ PIG 1795.II.17, sample 31; (14, 15) upper and lateral views, MUZ PIG 1795.II.18, sample 23; (16–18) upper, lower, and lateral views, MUZ PIG 1795.II.19, sample 01; (19, 20) upper and lateral views, MUZ PIG 1795.II.20, sample 19. (21) Conodont P1 elements of Icriodus sp., lateral view of the specimen figured by Bultynck (Reference Bultynck2003, pl. 2, fig. 33) as I. retrodepressus, Oriskany Falls section, New York (USA), Nedrow Member, sample 2. Scale bars=100 μm.
In addition to the study of our own collections, we analyzed literature data from North America, Europe, and Asia (Fig. 1). North American data are from the USA, from Indiana (Klapper and Johnson, Reference Klapper and Johnson1980; Klug, Reference Klug1983), Ohio (Klapper and Johnson, Reference Klapper and Johnson1980; Sparling, Reference Sparling1983), Iowa (Klapper and Barrick, Reference Klapper and Barrick1983), Minnesota (Klapper and Barrick, Reference Klapper and Barrick1983), and Wisconsin (Schumacher, Reference Schumacher1971); and from Canada, from the provinces of Ontario (Orr, Reference Orr1971; Uyeno et al., Reference Uyeno, Telford and Sanford1982), and British Columbia (Chatterton, Reference Chatterton1978). The discussed European data are from the Ardennes in Belgium (Bultynck, Reference Bultynck1970; Klapper and Barrick, Reference Klapper and Barrick1983, fig. 9.V, W) and the Eifel Mountains in Germany (Weddige, Reference Weddige1977; Werner and Ziegler, Reference Werner and Ziegler1982), and those from Asia come from Transcaucasus in Azerbaijan (Khalymbadzha, Reference Khalymbadzha1990).
Rationale for the present taxonomic concepts
Analysis of the material illustrated by Klapper and Barrick (Reference Klapper and Barrick1983, fig. 9) revealed considerable morphological variability within the two morphotypes distinguished by these authors. Their Morphotype I (with a depression in the posterior part of the spindle) displays mostly oval denticles in lateral rows, and rounded and slightly smaller ones in the median row. The same form shows a fairly wide posterior part of the spindle with convex margins, contrasting with a narrow anterior part, curving inward (Klapper and Barrick, 1983, fig. 9.R, X, Y, AA, early ontogenetic stage). The single illustrated specimen shows a spindle with a triangular outline (Klappner and Barrick, 1983, fig. 9.T, intermediate ontogenetic stage). This form reveals a close similarity to the mature form identified as Icriodus retrodepressus by Klapper and Barrick (Reference Klapper and Barrick1983, fig. 9.W).
Their Morphotype II (lacking the previously mentioned depression) is characterized by transverse rows of denticles formed by a fusion of small circular denticles of a median row with lateral denticles that are transversely elongated and display sharp edges. This morphotype has a wide spindle with convex margins (Klappner and Barrick, 1983, fig. 9.AF, Z) as well as a very narrow form, the outer margin of which is convex, whereas the inner one is concave-convex (Klappner and Barrick, 1983, fig. 9.AD).
The conodont collections from the Michigan and Belarussian basins, along with the previously illustrated specimens, were evaluated to consider the morphological similarity with the four above-mentioned varieties. It appeared that forms attributed to Morphotype I considerably differed from Morphotype II forms by the presence of a depression and the characteristic lateral rows of denticles, and for this reason, were excluded from the redefined Icriodus orri (see below). The latter species is typified by the holotype designated by Klapper and Barrick (Reference Klapper and Barrick1983, fig. 9.AF−AG) and was attributed to their Morphotype II. Morphotype I encompasses specimens differing not only in spindle shape and distribution of denticles on the upper surface, but also in the length and depth of the depression, and shape of the basal cavity. The differences are so conspicuous that they justify distinction of two separate species within Morphotype I. Forms with a short shallow depression, curved longitudinal axis, a spindle composed of a wide convex posterior part and narrow anterior part, and possessing a nearly circular basal cavity, have been defined here as I. michiganus n. sp. (Fig. 2). The forms characterized by a deeper depression, triangular spindle with a nearly straight axis, and a distinct basal cavity with a spur and antispur, have been included in the amended I. retrodepressus (Fig. 4).
Diagnostic differences among the species described below, and at the same time, intraspecific similarity of the specimens from various regions, are shown in Figure 5. It should be stressed that inhomogeneity of the material analyzed by Klapper and Barrick (Reference Klapper and Barrick1983) definitely led to confusion in later taxonomic studies. Some elements included in Icriodus orri lack a depression, but at the same time, show variable shapes of the spindle denticles. There is only a slight tendency for development of sharp upper margins, and a basal cavity has a more or less distinct spur (e.g., Mawson and Talent, Reference Mawson and Talent1989; Sparling, Reference Sparling1995; Uyeno and Lespérance, Reference Uyeno and Lespérance1997; Bultynck, Reference Bultynck2003; Narkiewicz and Kruchek, Reference Narkiewicz and Kruchek2008). The concept of I. retrodepressus was also broadly understood in previous studies. It included both triangular and distinctly biconvex forms, those with a curved axis, and having a depression of variable length and depth, but also specimens lacking the depression (e.g. Weddige, Reference Weddige1977; Garcia-Lopez, Reference Garcia-Lopez1987; Khalymbadzha, Reference Khalymbadzha1990; Malec, Reference Malec2003).

Figure 5 Upper views of representative specimens of the studied icriodid species and their stratigraphic ranges established by this study. Sources of illustrations: Icriodus orri, Minnesota (herein, Fig. 3.1); Michigan (herein, Fig. 3.15; British Columbia (Chatterton, Reference Chatterton1978, pl. 6, fig. 1); Belarus (herein, Fig. 3.6, 3.8); Icriodus michiganus n. sp., Michigan (herein, Fig. 2.3); Minnesota (Klapper and Barrick, Reference Klapper and Barrick1983, fig. 9.X, used with permission from SEPM; Ohio (Sparling, Reference Sparling1983, fig. 13.AS; used with permission from SEPM); Wisconsin (Schumacher, Reference Schumacher1971, pl. 9, fig. 2, used with permission from the Wisconsin Geological and Natural History Survey); Ontario (Uyeno et al., Reference Uyeno, Telford and Sanford1982, pl. 3, fig. 16); Icriodus retrodepressus, Ardennes (herein, Fig. 4.1, reproduced with permission); Michigan (herein, Fig. 4.16); Iowa (Klapper and Barrick, Reference Klapper and Barrick1983, fig. 9.T, used with permission from SEPM; Wisconsin (Schumacher, Reference Schumacher1971, pl. 9, fig. 5, used with permission from Wisconsin Geological and Natural History Survey); Indiana (Klapper and Johnson, Reference Klapper and Johnson1980, pl. 3, fig. 20, used with permission from SEPM). Broken horizontal lines=tentative limits of stratigraphic ranges; broken vertical lines=interval lacking documented occurrence of a species; vertical arrows=probable stratigraphic range. Scale bars=100 μm.
Materials and methods
In the systematic descriptions, new synonymies for Icriodus orri and I. retrodepressus are provided, and their diagnoses have been emended with detailed descriptions and remarks highlighting differences between them and similar taxa. To better document specific diagnostic features, Figures 3 and 4 reillustrate the holotypes and specimens representing the range of intraspecific variability. The specimens are illustrated under uniform magnification to better compare different ontogenetic stages and their variability.
Repository and institutional abbreviation
The described and photographed material is housed at the Geological Museum of the Polish Geological Institute-National Research Institute in Warszawa, Poland under collection numbers MUZ PIG 1804.II (Belarus collection) and MUZ PIG 1795.II (Michigan Basin collection).
Systematic paleontology
Order Prioniodontida Dzik, Reference Dzik1976
Family Icriodontidae Müller and Müller, Reference Müller and Müller1957
Genus Icriodus Branson and Mehl, Reference Branson and Mehl1938
Type species
Icriodus expansus Branson and Mehl, Reference Branson and Mehl1938.
Icriodus michiganus new species
1971 Icriodus nodosus (Huddle); Reference OrrOrr, pl. 2, figs. 20–21.
1971 Icriodus nodosus; Reference SchumacherSchumacher, p. 93, pl. 9, figs. 1−3, 7−9.
1980 Icriodus sp. aff. I. retrodepressus Bultynck; Reference Klapper and JohnsonKlapper and Johnson, pl. 3, figs. 22, 23.
1982 Icriodus aff. I. retrodepressus ; Reference Uyeno, Telford and SanfordUyeno et al., p. 33, pl. 3, figs. 16, 18, 19, 25−27.
1983 Icriodus orri Reference Klapper and BarrickKlapper and Barrick, p. 1230, fig. 9.R, X, Y, AA.
1983 Icriodus sp. aff. I. retrodepressus; Reference SparlingSparling, p. 851, fig. 13.AS−AT.
1990 Icriodus sp. E Weddige; Reference KhalymbadzhaKhalymbadzha, table 7, figs. 6−10.
1990 Icriodus retrodepressus; Reference KhalymbadzhaKhalymbadzha, table 8, figs. 24–25.
2003 Icriodus orri; Reference BultynckBultynck, pl. 2, figs. 34, 35.
Holotype
MUZ PIG 1795.II.1, P1 element from the United States Steel Corporation Quarry, Dundee Formation, sample 23 (Fig. 2.3−2.5); paratypes, P1 elements from the United States Steel Corporation Quarry: MUZ PIG 1795.II.2, top of the Dundee Formation, sample 01 (Fig. 2.6−2.8); MUZ PIG 1795.II.3, Dundee Formation, sample 21 (Fig. 2.14−2.16); MUZ PIG 1795.II.8, top of the Dundee Formation, sample 01 (Fig. 2.18–2.19).
Diagnosis
The P1 element is stout, with a relatively short and wide spindle. The posterior part is wide with biconvex margins and a straight axis, and is characterized by a clear short depression with one, rarely two, small middle-row denticles. The anterior part of the spindle is narrow, elongated, pointed, and more or less curved inward, and is characterized by the absence of middle-row denticles. The middle row consists of four or five rounded, isolated denticles, smaller than those of the lateral rows. One or two posterior denticles from the middle row are slightly displaced anteriorly whereas the remaining are aligned with the lateral-row denticles. The lateral rows are composed of six or seven oval, isolated denticles that are very close to each other in the posterior part of the spindle, with wider spaces in its anterior part. The cusp is large, pointed upward, and inclined posteriorly. The nearly symmetric expansion of the basal cavity starts in the middle part of the spindle or more anteriorly. The more or less pronounced spur is generally rounded.
Occurrence
The new species ranges from a lower part of the costatus Zone or even from an upper part of the partitus Zone to an upper part of the kockelianus Zone. It occurs in the USA in Ohio (Dundee and Delaware formations), in Michigan (Dundee Formation), in Wisconsin (uppermost part of the Lake Church Formation), in Minnesota (lower part of the Spillville Formation), in Canada in Ontario (Dundee Formation and sandy facies of the Anderdon Member of the Lucas Formation), and in Asia in Transcaucasus (lower part of the Danzik Formation).
Description
Denticles in the lateral rows display a tendency to develop sharp edges (Fig. 2.20, 2.22). The axes of the posteriormost lateral denticles are always directed outward and backward (V-shaped), with the outer denticle usually larger than the inner one. The cusp often bears a short, transverse lateral process on the outer side in left-curved forms (Fig. 2.11, 2.18), and on the inner side in right-curved forms (Fig. 2.3, 2.14). The basal cavity consists of a narrow anterior part and a wide posterior part. The inner margin of the expanded part of the basal cavity is more or less oblique, whereas the outer margin is more or less rounded. A distinct sinus is present on the posterior margin of the basal cavity and a shallower sinus occurs in the posterior part of the inner margin of the basal cavity.
Etymology
The name of the species is derived from the Michigan Basin in the state of Michigan, USA where the taxon has been identified.
Materials
MUZ PIG 1795.II.4 (Fig. 2.1, 2.2), 1795.II.5 (Fig. 2.9, 2.10), 1795.II.6 (Fig. 2.11−2.13), 1795.II.7 (Fig. 17), 1795.II.9 (Fig. 2.20, 2.21), 1795.II.10 (Fig. 2.22).
A total of 19 specimens from the U.S. Steel Corporation Quarry, Michigan Basin, Dundee Formation, from samples 31 to 01.
Remarks
The new species differs from Icriodus orri in the presence of a depression in the posterior median part of the spindle, and in the lack of characteristic transverse denticle ridges with sharp edges. Another difference is a smaller number of the median-row denticles that are larger than those in I. orri, and a more symmetrical basal cavity lacking a sharply accentuated spur. The described species differs from I. retrodepressus in the outline and width of the spindle and in the development of the basal cavity. The spindle of I. retrodepressus is narrow and triangular, whereas the depression in its posterior part is deeper and longer. Its basal cavity is asymmetric with a distinct spur and antispur, and its inner margin is more oblique than that in I. michiganus n. sp.
Icriodus orri Klapper and Barrick, Reference Klapper and Barrick1983
1978 Icriodus aff. I. expansus Branson and Mehl; Reference ChattertonChatterton, p. 201, pl. 6, figs. 1, 2, 5.
1983 Icriodus orri Reference Klapper and BarrickKlapper and Barrick, p. 1230, only fig. 9.Z (paratype), 9.AF−AG (holotype).
non 1989 Icriodus orri; Reference Mawson and TalentMawson and Talent, pl. 1, figs. 13−15.
non 1995 Icriodus orri; Reference SparlingSparling, fig. 4.1−4.5.
non 1997 Icriodus orri; Reference Uyeno and LespéranceUyeno and Lespérance, fig. 7.21−7.25, 7.31–7.32.
non 1998 Icriodus orri; Reference UyenoUyeno, pl. 11, figs. 6, 7, 12, pl. 12, figs. 22, 23.
non 2003 Icriodus orri; Reference BultynckBultynck, pl. 2, figs. 34, 35.
non 2008 Icriodus orri; Reference Narkiewicz and KruchekNarkiewicz and Kruchek, pl. 1, figs. 11, 12, 14, 15, 19.
Holotype
Element P1 from the Racine Quarry, sample 3, Spillville Formation, southern Minnesota, USA, (Klapper and Barrick, Reference Klapper and Barrick1983, fig. 9.AF–AG).
Diagnosis
Element P1 is characterized by an elongated, biconvex to lachrymiform spindle that is widest in the middle or just behind, closer to its posterior part. The spindle axis is almost straight, particularly in its posterior part, although the anterior narrow and pointed extremity of the spindle can be more or less curved inward. The small denticles of the middle row are more or less of uniform size and have circular cross sections. They can be located just opposite or slightly anterior to those of the lateral row denticles and are connected with them, forming seven or eight transverse denticle rows. The middle-row denticles are on the same level or a little higher than the lateral-row denticles. The latter are laterally elongated, with sharp upper margins, and are markedly larger than the middle-row denticles. The cusp is mostly rounded and inclined posteriorly, and its width is equal to the lateral extent of the two posteriormost denticles. The basal cavity is asymmetrical with the outer part wider and beginning more anteriorly than the inner one; on its inner side, there is a well-pronounced spur. The expanded outer part of the basal cavity is located in the middle or posterior third of the element.
Occurrence
This species ranges from an uppermost part of the kockelianus to ensensis zones. It is present in the USA in Michigan (lower part of the Bell Shale Formation), in Minnesota (lower part of the Spillville Formation), in Canada in British Columbia (Dunedin Formation), and in Belarus in the Pripyat Graben (Kostyukovichi Horizon).
Description
The intraspecific variability comprises elongated and fairly wide forms such as the holotype (Fig. 3.1) and narrow ones similar to that illustrated from British Columbia (Fig. 5). Ornamentation can be irregular, particularly with respect to the length of lateral row denticles. In some specimens, these can be longer than usual due to a development of additional short processes (Fig. 3.4, 3.5). Denticles in the last one or two posterior transverse rows show an inverted chevron pattern, whereas those in the anterior end of the spindle are mostly straight. The space between transverse denticle rows is much larger in the anterior part of the spindle than in the posterior part. Denticles of the middle row can be connected by thin ridges, which also continue posteriorly to the tip of the cusp. These denticles occur in the posterior and middle part of the spindle, whereas in its anterior part, they can be completely fused with the denticles of lateral rows. The size of the middle-row denticles can be variable. In the most posterior transverse rows, they are always small, whereas in the next two or three rows they can be larger. The cusp can have a short transverse process on its inner side. The width of the basal cavity is variable, from considerably expanded as seen in the holotype, to medium sized in the paratype (Fig. 3.3) or in remaining specimens from the Pripyat Graben and Michigan Basin (compare Fig. 3.5, 3.6, 3.15). Usually, sinuses are present on the inner and posterior margins of the basal cavity.
Remarks
Icriodus orri differs from I. michiganus n. sp. and I. retrodepressus in lacking a depression in the posterior part of the spindle, and in its characteristic ornamentation. Icriodus norfordi Chatterton, Reference Chatterton1978 (pl. 6, fig. 12, holotype) has a nearly triangular spindle outline, whereas in I. orri, the spindle is clearly biconvex. The denticulation pattern of I. norfordi is more irregular and its basal cavity is almost symmetrical, broadly expanded close to the anterior part of the spindle and lacking a distinct spur. Icriodus orri can be distinguished from I. arkonensis arkonensis Stauffer, Reference Stauffer1938 (Klapper, Reference Klapper1975, p. 77, pl. 1, figs. 3, 4) in that the latter has thicker lateral-row denticles lacking sharp edges and almost in contact with each other. On the other hand in I. orri, the denticles are thin and have sharp upper margins, and the transverse rows are clearly separated, with spacing increasing toward the anterior end. In I. a. arkonensis, the denticles of the middle row are low and occur along the entire length of the spindle, whereas in I. orri, they are of the same height as the lateral denticles or even slightly higher, and they are missing in the anterior part. Icriodus orri has a distinct spur on the inner side of the basal cavity, which is considerably more pronounced than in I. a. arkonensis.
Icriodus retrodepressus Bultynck, Reference Bultynck1970
1970 Icriodus retrodepressus Reference BultynckBultynck, p. 110, pl. 30, figs. 1−4 (non figs. 5, 6).
1971 Icriodus nodosus (Huddle); Reference SchumacherSchumacher, pl. 9, figs. 4−6, 11−12.
1975 Icriodus retrodepressus; Reference ZieglerZiegler, p. 143, pl. 8, figs. 4, 5 (refigured from Bultynck Reference Bultynck1970, pl. 30, figs. 1, 4).
1977 Icriodus corniger retrodepressus; Reference WeddigeWeddige, p. 290, pl. 1, figs. 10, 12 (non fig. 11).
1980 Icriodus sp. aff. I. retrodepressus; Reference Klapper and JohnsonKlapper and Johnson, pl. 3, figs. 20, 21.
1982 Icriodus corniger retrodepressus; Reference Werner and ZieglerWerner and Ziegler, pl. 2, figs. 26, 27.
1982 Icriodus aff. I. retrodepressus; Reference Uyeno, Telford and SanfordUyeno et al., p. 33, pl. 3, fig. 23.
1983 Icriodus orri Reference Klapper and BarrickKlapper and Barrick; Klapper and Barrick, fig. 9.T.
1983 Icriodus retrodepressus; Reference Klapper and BarrickKlapper and Barrick, fig. 9.V, W.
1983 Icriodus retrodepressus; Reference KlugKlug, fig. 9.A−F.
non 1990 Icriodus retrodepressus; Reference KhalymbadzhaKhalymbadzha, table 8, figs. 23, 24.
non1987 Icriodus retrodepressus; Reference Garcia-LopezGarcia-Lopez, pl. 7, figs. 9−20
non 2003 Icriodus corniger retrodepressus; Reference MalecMalec , table 343, figs. 9−13.
2003 Icriodus orri; Reference BultynckBultynck, pl. 2, figs. 27, 28 (non fig. 33)
Holotype
Element P1 from the Couvin Formation, Ardenne, Belgium (Bultynck, Reference Bultynck1970, pl. 30, fig. 1a–c). The holotype has a provisional repository signature ‘b168 Bultynck, Reference Bultynck1970’ in the Paleontology Collection of the Belgian Royal Institute of Natural Sciences (Brussels).
Diagnosis
The P1 element is characterized by an elongated, triangular spindle with a pronounced depression in its posterior part, showing one to three small middle-row denticles. In adult specimens, the lateral rows consist of six to eight oval, transversely developed denticles. In the posterior part, they can be connected with the middle-row denticles. Middle-row denticles have circular cross sections and are much smaller than those of the lateral rows. The cusp, with a fine ridge on its anterior side, is large and inclined posteriorly. The basal cavity is asymmetrical and its expanded part occupies the posterior half or even two-thirds of the element. There is a well-developed oblique spur on the inner side and a rounded posterior expansion on the outer side. The spur is rectilinear and joins the posterior margin of the cavity in that way.
Occurrence
The species spans the interval from the partitus Zone to the upper part of the kockelianus Zone. It occurs in the USA in Iowa (lower part of the Spillville Formation), in Indiana (Speeds Member of the North Vernon Formation and Rocky Branch bone bed), in Michigan (Dundee Formation), in Wisconsin (uppermost part of the Lake Church Formation), in Canada in Ontario (Dundee Formation), in Europe in Belgian Ardennes (Couvin Formation), and in the Eifel Mountains of Germany (lower part of the Lauch Formation).
Description
Intraspecific variability comprises forms that are narrow and thus similar to the holotype (Fig. 4.1), wider elements (Fig. 4.4), and triangular forms with a slightly convex inner margin (Fig. 4.10, 4.14; Schumacher, Reference Schumacher1971, pl. 9, fig. 5; Klapper and Johnson, Reference Klapper and Johnson1980, pl. 3, fig. 20; Fig. 5). Anterior spindle termination can be straight or slightly curved inward. In typical representatives of Icriodus retrodepressus, the strongly accentuated spur is mostly anteriorly directed, and the outer part of the basal cavity is not much expanded. In a few specimens, however, the spur does not show a distinct anterior direction, and the outer part of the basal cavity is considerably expanded outward (e.g., Klapper and Johnson, Reference Klapper and Johnson1980, pl. 3, fig. 20; Uyeno et al., Reference Uyeno, Telford and Sanford1982, pl. 3 fig. 23; Fig. 5).
Remarks
According to Klapper and Barrick (Reference Klapper and Barrick1983, p. 1231), the size of the basal cavity was the main reason to separate Icriodus orri from I. retrodepressus. These authors mentioned that I. retrodepressus is known mainly from the Ardennes and Eifel Mountains, from a narrow interval of the partitus Zone. In fact, the basal cavity of the holotype (Fig. 4.1−4.3; Bultynck, Reference Bultynck1970, pl. 30, fig. 1) is limited mainly to the posterior part of the spindle. In the specimens illustrated from the Eifel Mountains area, however, the basal cavity can be situated either in the posterior or middle part of the spindle (Werner and Ziegler, Reference Werner and Ziegler1982, pl. 2, figs. 26, 27=Fig. 4.5). The younger representatives of I. retrodepressus from North America are characterized by a basal cavity that always expands in the anterior part of the spindle (Fig. 4.6−4.20).
Analysis of the material from the Michigan Basin and literature data demonstrate that although the size of the basal cavity can differ in European and North American representatives, its asymmetry, with its well-pronounced spur and antispur, remains a constant feature, as does morphology of the upper surface of the spindle. The changing size of the basal cavity could be related to the presumed feeding functions performed by the P1 elements. The latter functions could require, for example, stronger or weaker embedding in the soft tissues of an animal, depending on various types of food.
The differences between Icriodus retrodepressus and I. michiganus n. sp. are given above in the description of the latter species.
Conodont zonation of the Eifelian stage
Verification of published stratigraphic conodont data (Table 1, Chronostratigraphy) must take into account changes that have been introduced to the Eifelian conodont zonation since its earliest proposal. In 1966, Wittkekindt (p. 627) established Icriodus corniger, Spathognathodus bidentatus, and Polygnathus kockelianus zones for the Rhenish Slate Mountains area. The most complete Eifelian zonation proposed by Weddige (Reference Weddige1977, table 5) for the Eifel Mountains, Rhenish Slate Mountains, and neighboring areas comprised (in ascending order) upper P. patulus, P. costatus costatus, Tortodus kockelianus, and P. ensensis zones, and has been applied since then albeit with some modifications. Klapper and Ziegler (Reference Klapper and Ziegler1979, text-figs. 3, 4) introduced the T. kockelianus australis Zone below the kockelianus Zone, while attributing global significance to the zonation. At the same time, Weddige et al. (Reference Weddige, Werner and Ziegler1979) established the P. partitus Zone for the lowermost Eifelian, replacing the upper part of the former patulus Zone. Bultynck (Reference Bultynck1987) proposed to introduce the P. hemiansatus Zone as an equivalent of the upper part of the ensensis Zone in the lowermost Givetian, although the lower part of this zone was not discussed. Consequently, the ensensis Zone disappeared and ceased to be used during the next decade (as e.g., in the compilation by Clausen et al., Reference Clausen, Weddige and Ziegler1993 that is widely cited as the standard authority on conodont zonation). However, Walliser (Reference Walliser2000, fig. 2) reintroduced the ensensis Zone in the uppermost Eifelian as an equivalent of the lower part of the former ensensis Zone.
Table 1 Previous taxonomic and biostratigraphic data on Icriodus orri and related species compared with the data from this study.

The above-described biozonation is based on the first appearances of index taxa in the phylogenetic lineages of the Eifelian open and/or deeper marine genera Polygnathus Hinde, Reference Hinde1879 and Tortodus Weddige, Reference Weddige1977. For the shallower and more nearshore facies, characterized by a co-occurrence of polygnathids and pelekysgnathids, Klapper and Ziegler (Reference Klapper and Ziegler1979, text-fig. 4) distinguished the informal Pelekysgnathus pedderi, Polygnathus curtigladius and Po. pseudofoliatus faunas above the costatus Zone. The pedderi Fauna corresponds approximately to the lower part of the australis Zone, the curtigladius Fauna to the upper part of this zone and lower part of the kockelianus Zone, and the pseudofoliatus Fauna to the upper kockelianus and ensensis zones. Later, based on data from western Canada, Braun et al. (Reference Braun, Norris and Uyeno1988, fig. 2) correlated the curtigladius Fauna with the upper part of the kockelianus Zone (without the uppermost portion), and the pseudofoliatus Fauna with the ensensis Zone.
The shallow marine and/or nearshore Eifelian conodont faunas were dominated by representatives of the genus Icriodus Branson and Mehl, Reference Branson and Mehl1938. The Eifelian zonation based mostly on icriodids was first proposed by Orr (Reference Orr1971) for intracratonic North American settings. He distinguished three zones (in ascending order): the Icriodus latericrescens robustus (=Latericriodus l. robustus) Zone, the Polygnathus ‘webbi’ (=P. costatus) Zone, and I. angustus Zone. In turn, for the same time interval, Klug (Reference Klug1983, p. 82, fig. 3) defined the I. l. robustus, I. angustus angustus and Po. pseudofoliatus zones. According to Klug (Reference Klug1983), the I. l. robustus Zone is probably an equivalent of the patulus Zone and I. a. angustus Zone might correspond to the costatus Zone. The local P. pseudofoliatus Zone is correlated with the australis-ensensis zonal interval.
Changes in the conodont zonation have affected biostratigraphic correlation of the eustatic cycles in the Eifelian, particularly in its upper part. According to Johnson et al. (Reference Johnson, Klapper and Sandberg1985), the transgressive events of the cycles Ie and If correspond to the upper part of the kockelianus Zone and to the ensensis Zone, respectively. According to the recent zonation, the lower boundary of the If cycle (=otomari Event,=onset of black shale sedimentation marking the Kačak Event of House, Reference House1985) is correlated with the kockelianus/ensensis zonal boundary (Walliser, Reference Walliser2000, p. 40).
Regional biostratigraphic comparisons
In order to establish the stratigraphic ranges of the above-described Icriodus species, both new and published data on the age of the conodont-bearing strata have been reviewed and verified. Table 1 summarizes the sources of the discussed materials and compares the original interpretations with the revised results. We selected only those localities for which the taxonomic identifications could be verified based on illustrated specimens.
Spillville Formation
Icriodus orri sensu Klapper and Barrick, Reference Klapper and Barrick1983 has been encountered in the entire Spillville Formation. Klapper and Barrick (Reference Klapper and Barrick1983) tentatively attributed the lower part of the formation approximately to the kockelianus Zone, based on the presence of the curtigladius Fauna together with Ozarkodina raaschi Klapper and Barrick, Reference Klapper and Barrick1983. The upper part was dated as the ensensis Zone owing to the co-occurrence of O. raaschi and O. brevis (Bischoff and Ziegler, Reference Bischoff and Ziegler1957), the latter species first appearing in that zone (Klapper and Johnson, Reference Klapper and Johnson1980). The specimens attributed to I. orri in this report were obtained from the lower Spillville Formation. The species has been identified in the Racine Quarry in samples 3 and 3A (Klapper and Barrick, Reference Klapper and Barrick1983, fig. 4). Icriodus michiganus n. sp. has been ascertained in the Racine section in samples 2A and 3; in Spillville, in sample 1A; and in Albion, in sample 1 (Klapper and Barrick, Reference Klapper and Barrick1983, figs. 4, 5). Icriodus retrodepressus has been identified in the Spillville Quarry, sample 2.
The Racine section is the most favorable one for estimating the age of the lower Spillville Formation. In this section, Icriodus orri appears above the highest occurrence of Polygnathus curtigladius Uyeno, Reference Uyeno1979 (Klapper and Barrick, Reference Klapper and Barrick1983, fig. 4). On the other hand, I. michiganus n. sp. appears still within the range of P. curtigladius. Based on the curtigladius Fauna, Klapper and Barrick (Reference Klapper and Barrick1983, fig. 3) correlated the lower Spillville Formation with the lower part of the Elm Point Formation in Manitoba, Canada. In turn, Braun et al. (Reference Braun, Norris and Uyeno1988, fig. 2) attributed the lower Elm Point Formation to the upper (but not uppermost) part of the kockelianus Zone after taking into account all available paleontological data. This age is consistent with the earlier interpretation of Johnson et al. (Reference Johnson, Klapper and Sandberg1985, fig. 8) who considered the Elm Point Formation as transgressive strata related to their eustatic cycle Ie.
Lake Church Formation
Icriodus michiganus n. sp. and I. retrodepressus (Table 1), earlier determined as I. nodosus Huddle, Reference Huddle1934 by Schumacher (Reference Schumacher1971), have been identified in the upper part of the formation. Klapper and Barrick (Reference Klapper and Barrick1983, fig. 3) correlated these strata with the lower Spillville Formation based on a similarity of the conodont and macrofaunal assemblages. Therefore, its age is here tentatively referred to the upper but not uppermost part of the kockelianus Zone.
Delaware Formation
In the Parkertown Quarry in north-central Ohio, Sparling (Reference Sparling1983) found specimens that he determined as Icriodus sp. aff. I. retrodepressus, and that are here included in I. michiganus n. sp. (Table 1). They were encountered in the upper part of the Delaware Formation, 12.42−12.32 m above its base (sample S8OP-15). The entire formation was dated as the kockelianus Zone (Sparling, Reference Sparling1983, fig. 9).
Johnson et al. (Reference Johnson, Klapper and Sandberg1985, fig. 8) interpreted the Elm Point Formation (Canada) and Delaware Formation (USA) as co-eval transgressive units of the eustatic cycle Ie. Sparling (Reference Sparling1988, fig. 9), however, regarded only the lower part of the Delaware Formation as transgressive, and the upper as regressive. According to his data, the entire formation corresponds to the kockelianus Zone except for its lowermost and uppermost part (Sparling, Reference Sparling1988, fig. 2). Later, Sparling (Reference Sparling1999, fig. 2) attributed the Delaware Formation to the upper part of the kockelianus Zone as was earlier proposed by Johnson et al. (Reference Johnson, Klapper and Sandberg1985).
North Vernon Formation, Speeds Member
Icriodus retrodepressus first appears in this unit in the sample BMQ-4 in the Berry Materials Corporation Quarry (BMQ, locality 1) in south-central Indiana (Klug, Reference Klug1983, figs. 7, 9.A−F). The age of the sample was determined as a lower part of the Polygnathus pseudofoliatus Zone (Klug, Reference Klug1983, fig. 5). Icriodus retrodepressus co-occurs with I. angustus Stewart and Sweet, Reference Stewart and Sweet1956 (Klug, Reference Klug1983, fig. 8.D−F), among other taxa. The age can be now attributed to the kockelianus Zone, because I. retrodepressus is present in sample BMQ-5, ~0.7 m above the highest occurrence of I. latericrescens robustus Orr, Reference Orr1971 (sample BMQ-7), which disappears in the australis Zone (Klapper and Johnson, Reference Klapper and Johnson1980), whereas the highest appearance of I. angustus is in the kockelianus Zone (Klapper and Johnson, Reference Klapper and Johnson1980). Because the same age can be ascribed to sample BMQ-3, the interval between the samples BMQ-5 and BMQ-3 also belongs to the kockelianus Zone. Samples BMQ-1 and BMQ-2, localized at the top of the Speeds Member and slightly above it, respectively, contain a stratigraphically mixed assemblage indicating a gap spanning the ensensis through timorensis zones, and perhaps even a part of the kockelianus Zone. The data suggest that the analyzed BMQ-4 sample could represent an upper part of the kockelianus Zone (compare Brett et al., Reference Brett, Baird, Bartholomew, DeSantis and Ver Straeten2011, fig. 5).
Rocky Branch bone bed
The specimen that was identified as Icriodus sp. aff. I. retrodepressus by Klapper (in Klapper and Johnson, Reference Klapper and Johnson1980) in the Rocky Branch bone bed in Jennings County, southeastern Indiana, is now attributed to I. retrodepressus (Table 1). Because this unit is a lateral equivalent of the Speeds Member, its age should correspond approximately to the upper part of the kockelianus Zone (see above).
Dunedin Formation
In the Mount Jane Smith locality in northeastern British Columbia, Icriodus orri, earlier identified by Chatterton (Reference Chatterton1978) as I. aff. I. expansus (Table 1), was found 30 m below the top of the Dunedin Formation (Chatterton, Reference Chatterton1978, p. 162). These strata were dated as latest Emsian to early Eifelian based on the conodont fauna. The Mount Jane Smith section (situated in the area of the tectonosedimentary element labeled MacDonald Shelf by Morrow and Geldsetzer, Reference Morrow and Geldsetzer1988, fig. 3) is localized near the northernmost limit of the Dunedin Formation occurrence (Chatterton, Reference Chatterton1978, p. 164). Given that the formation is ~95 m thick there (Taylor and MacKenzie, Reference Taylor and MacKenzie1970, p. 12, fig. 5), it can be assumed that I. orri occurs in its upper part. According to Morrow and Geldsetzer (Reference Morrow and Geldsetzer1988, fig. 4), these strata correspond approximately to the kockelianus-lower ensensis zonal interval, which was also confirmed by Uyeno (Reference Uyeno1991, fig. 4).
Dundee Formation
This unit occurs in the Michigan Basin in southwestern Ontario Province in Canada (Orr, Reference Orr1971; Uyeno et al., Reference Uyeno, Telford and Sanford1982), northeastern Michigan State (Bultynck, Reference Bultynck1976), and northwestern Ohio (Klapper and Johnson, Reference Klapper and Johnson1980). In section 14 (Brunner Quarry) from Essex County of southwestern Ontario, the present authors identified Icriodus michiganus n. sp., earlier determined as I. nodosus by Orr (Reference Orr1971; Table 1). However, the precise locality of the relevant sample in the section is not known.
In the northern part of Michigan’s Lower Peninsula (USA), in section 15 (U.S. Steel Corporation Quarry), the age of the formation was established as Eifelian by Bultynck (Reference Bultynck1976, fig. 2) based on comparison with the Belgian Ardennes conodont succession. The present analysis and verification of the material from this section has shown that Icriodus michiganus n. sp. and I. retrodepressus occur in the entire Dundee Formation (Table 1), from sample 31 to sample 01 (sample numbers after Bultynck, Reference Bultynck1976, table 1).
From the Medusa North Quarry in Lucas County in northwestern Ohio (Klapper and Ziegler, Reference Klapper and Ziegler1967, p. 79), Klapper (in Klapper and Johnson, Reference Klapper and Johnson1980) illustrated a representative of Icriodus sp. aff. I. retrodepressus, now attributed to I. michiganus n. sp. (Table 1). It was found in the lower part of the Dundee Formation, 4.4−4.7 m below its top, in strata corresponding to the upper part of the costatus Zone.
The lower part of the Dundee Formation has been studied by Uyeno et al. (Reference Uyeno, Telford and Sanford1982, table 8) in the United Steel Company DDH No. 1 section in southwestern Ontario. Icriodus retrodepressus, earlier determined by Uyeno as I. aff. I. retrodepressus (Table 1), was found 14.94−15.54 m above the base of the formation, assigned by Uyeno et al. (Reference Uyeno, Telford and Sanford1982, fig. 5) to the upper part of the costatus Zone.
In the Michigan Basin (northern part of the Lower Peninsula of Michigan) in the lowermost sample studied by Bultynck (Reference Bultynck1976, table 1, sample 31), Icriodus angustus occurs in the basal part of the formation together with I. michiganus n. sp. (Fig. 2.20−2.21) and I. retrodepressus (Fig. 4.11−4.13). The first species has its entry in the costatus Zone (Klapper and Johnson, Reference Klapper and Johnson1980, table 7). Another characteristic species, Polygnathus angustipennatus Bischoff and Ziegler, Reference Bischoff and Ziegler1957, occurs in sample 1 (Bultynck, Reference Bultynck1976, table 1) in the lower part of the Rogers City Formation overlying the Dundee Formation. The lowest stratigraphic range of the species is in the australis Zone (Walliser and Bultynck, Reference Walliser and Bultynck2011). The above-mentioned data indicate that the entire Dundee Formation and the lowermost part of the Rogers City Formation in Michigan belong to the costatus Zone.
The lower Dundee Formation in southwestern Ontario was dated by Uyeno et al. (Reference Uyeno, Telford and Sanford1982, fig. 5) as an upper part of the costatus Zone. These authors correlated the formation with similar strata in Michigan, and, based on the data from both areas, estimated the age of the formation as the upper part of the costatus Zone. The same age is assumed by the present authors for the upper part of the Dundee Formation in northwestern Ohio.
Lucas Formation, Anderdon Member
In southwestern Ontario, 26.82−27.31 m above the base of the calcareous-sandy Anderdon Member in section 13, Uyeno et al. (Reference Uyeno, Telford and Sanford1982, fig. 3, table 4b) noted the co-occurrence of Icriodus aff. I. retrodepressus and Polygnathus costatus patulus→P. cooperi cooperi (Uyeno et al., Reference Uyeno, Telford and Sanford1982, table 4b, respectively, pl. 3, figs. 25−27, pl. 1, figs. 21−23). The first taxon is here included in I. michiganus n. sp. (Fig. 5, Table 1). The second one falls into the intraspecific variability range of P. partitus Klapper, Ziegler, and Mashkova, Reference Klapper, Ziegler and Mashkova1978. The affiliation to this species by the present authors is based on the narrow shape of the platform, the straight inner posterior margin, and the nearly straight outer margin, which converge to form a sagittate outline of the posterior platform. According to Uyeno et al. (Reference Uyeno, Telford and Sanford1982), the age of the Anderdon Member corresponds to the upper part of the undivided patulus Zone (present partitus Zone). The presence of P. partitus, ranging from the partitus Zone to the lower costatus Zone (Werner and Ziegler, Reference Werner and Ziegler1982; Berkyová, Reference Berkyová2009), suggests the same age for the sandy Anderdon strata. Nevertheless, Brett et al. (Reference Brett, Baird, Bartholomew, DeSantis and Ver Straeten2011, p. 29, fig. 5), based on the correlation of a shallow-marine fauna from various North American intracratonic basins, attributed the sandy Anderdon facies to the regressive phase of the Eif-1 cycle in the lower part of the costatus Zone.
Danzik Formation.—This unit has been recognized in the upper part of the Mount Kara-Burun section and in the lower part of the Arpachay River section near Danzik village in the Autonomous Republic of Nakhichevan (Azerbaijan, Transcaucasus) (Khalymbadzha, Reference Khalymbadzha1990, figs. 6, 7). The formation is composed predominantly of terrigenous deposits, shales, and siltstones with rare intercalations of dark bituminous limestones containing conodonts. Among these are specimens identified by Khalymbadzha (Reference Khalymbadzha1990) as Icriodus n. sp. E of Weddige (Reference Weddige1977) and I. retrodepressus (Table 1). All these elements are here attributed to I. michiganus n. sp., and the age of the strata is constrained to the costatus Zone.
European localities
Verification of Icriodus orri reported earlier from the lower part of the Kostyukovichi Horizon in the Pripyat Graben of Belarus (Narkiewicz and Kruchek, Reference Narkiewicz and Kruchek2008, pl. 1) confirmed the earlier taxonomic and biostratigraphic determinations (Table 1). Also, earlier data from the Belgian Ardennes and Eifel Mountains strata containing I. retrodepressus were confirmed, their age being attributed to the partitus Zone (Weddige, Reference Weddige1977; Werner and Ziegler, Reference Werner and Ziegler1982; Bultynck et al., Reference Bultynck, Coen-Aubert and Godefroid2000).
Biostratigraphic importance of the results
The above-discussed biostratigraphic data have been used to establish total stratigraphic ranges for the described Icriodus species (Fig. 5), and to evaluate their applicability for chronostratigraphic correlations.
Icriodus retrodepressus has the widest stratigraphic range, covering the interval from the partitus Zone to the upper but not uppermost part of the kockelianus Zone. It first appears slightly above the base of the partitus Zone in Europe. In the Lower-Middle Devonian boundary stratotype in the Wetteldorf Richtschnitt section of Germany, it occurs in the lower part of the Lauch Formation in the Bed WP 37, ~2 m above the entry of the zonal species Polygnathus partitus (Werner and Ziegler, Reference Werner and Ziegler1982; Ziegler, Reference Ziegler2000), and thus in the lower partitus Zone. In the Belgian Ardennes, the first representatives of the species were found in the Couvin Eau Noire section (Couvin Formation) in sample 62/2 and thus ~7 m above the lowest occurrence of P. partitus in sample 54, in the highest part of the Eau Noire Formation (sample numbers from Bultynck, Reference Bultynck1970, pl. 30; section from Bultynck and Godefroid, Reference Bultynck and Godefroid1974, p. 5, 6; compare with Bultynck et al., Reference Bultynck, Coen-Aubert and Godefroid2000, fig. 5). Bultynck (Reference Bultynck2003) suggested occurrence of the species in the partitus Zone in the Nedrow Member (New York State, USA), but this seems doubtful because the specimen illustrated by Bultynck (Reference Bultynck2003, pl. 2, fig. 33) does not possess a depression in the posterior part of the spindle (Fig. 4.21, showing lateral view), and thus cannot be included in I. retrodepressus. The first representatives of this species in North America were reported from an upper part of the costatus Zone at the base of the Dundee Formation in the Michigan Basin. The uppermost range of I. retrodepressus has been recognized in the Spillville section, in sample 2 from the lower part of the Spillville Formation (Klapper and Barrick, Reference Klapper and Barrick1983). The age of the sediments is attributed to the upper (but not uppermost) part of the kockelianus Zone (see Regional biostratigraphic comparisons; Table 1). So far, I. retrodepressus has not been recognized in the lower part of the costatus Zone, in the australis Zone, or in the lower part of the kockelianus Zone.
The earliest occurrence of Icriodus michiganus n. sp. has been noted in the sandy facies of the Anderdon Member, Lucas Formation (Uyeno et al., Reference Uyeno, Telford and Sanford1982). This unit was attributed to the interval comprising the partitus Zone to a lower part of the costatus Zone (see above; Table 1). The lower boundaries of both zones have not been recognized in the investigated strata. The present authors suggest that I. michiganus n. sp. first appears in the lower part of the costatus Zone, and possibly even earlier, in the partitus Zone. The latest occurrence of the species has been noted in the lower Spillville Formation dated as the upper (but not uppermost) part of the kockelianus Zone (Racine section, samples 2A, 3, and Albion locality, sample 1; Klapper and Barrick, Reference Klapper and Barrick1983, figs. 4, 5). So far, I. michiganus n. sp. has not been found in the australis and lower part of the kockelianus zones.
Icriodus orri first appears in the lower Spillville Formation above the last occurrence of P. curtigladius (Racine section, sample 3, Klapper and Barrick, Reference Klapper and Barrick1983, fig. 4). These strata were dated as the uppermost part of the kockelianus Zone. The last occurrence is in the ensensis Zone based on data from the Pripyat Graben of Belarus (Narkiewicz and Kruchek, Reference Narkiewicz and Kruchek2008).
Paleogeographic controls on species distribution
When projected on the paleogeographic map of Scotese (Reference Scotese2002), the distributions of Icriodus retrodepressus, I. michiganus n. sp., and I. orri appear concentrated mainly in shallow epicontinental marine areas along the southern margin of the Euramerican continent (Fig. 1). None of the three species has been found in Gondwana, e.g., in the well-studied Devonian sections of the Moroccan Anti-Atlas.
It can be noticed that particular species have a characteristic distribution in time and space. The earliest occurrences of Icriodus retrodepressus, in the lowermost Eifelian, have been noted only in the European part of Euramerica, in the Ardennes (Bultynck, Reference Bultynck1970) and in the Eifel Mountains (Weddige, Reference Weddige1977; Werner and Ziegler, Reference Werner and Ziegler1982). The relatively high number of the species in the Eifel Mountains area (Weddige, Reference Weddige1977, p. 291) suggests that this was the region where the species evolved and from which it expanded into other basins. It can be assumed that the most favorable habitats for the species were the neritic facies with some pelagic influences, as represented by the crinoidal limestones with brachiopods, trilobites, and rugose corals, occurring in the lower part of the Lauch Formation (Werner and Ziegler, Reference Werner and Ziegler1982, p. 27; Weddige, Reference Weddige1988, p. 114). The disappearance of the species corresponds to a deepening of the basin and introduction of numerous representatives of I. corniger corniger Wittekindt, Reference Wittekindt1966 known from the goniatite-bearing pelagic facies (Wittekindt, Reference Wittekindt1966).
In the Ardennes, rare specimens of Icriodus retrodepressus (see Bultynck, Reference Bultynck1970, pl. 30) have been found in the alternating calcareous and shaly beds with brachiopods and tabulate corals. The shaly interbeds disappear upward, replaced by a biostromal facies with domal and platy stromatoporoids and corals (Bultynck, Reference Bultynck1970, pl. 35; Bultynck and Godefroid, Reference Bultynck and Godefroid1974, p. 6). The last representative of I. retrodepressus has been found in the basal part of these sediments. The gradual shallowing and the onset of biostromal facies are considered unfavorable for the species in question, leading to its disappearance in that area. The frequency of the species increased within the costatus Zone, characterized by shallow-marine, quiet-water sedimentation in the intracratonic Michigan Basin (Bultynck, Reference Bultynck1976). Numerous representatives of the species have also been found in the shaly calcareous sediments of the Speeds Member in the upper part of the kockelianus Zone (Klug, Reference Klug1983).
The geographic range of Icriodus michiganus n. sp. includes shallow-marine epeiric seas of the inner part of the North American craton (Fig. 5, Table 1) and on the eastern margin of Euramerica (Transcaucasus) (Fig. 1, Table 1). The species probably first appeared in the American part of Euramerica and thereafter migrated along the northern continental margin not reaching the European part. Icriodus orri has a similar distribution except that it was also found in the European part of Euramerica (Belarus) in addition to the North American area (Iowa, British Columbia).
The first appearance of Icriodus retrodepressus might have been related to the transgressive Choteč Event, connected to considerable global biotic changes (Walliser, Reference Walliser1996). The appearance of I. michiganus n. sp. seems to be unrelated to any known eustatic and/or global biotic event. The introduction of I. orri correlates approximately with the transgression of the eustatic cycle Ie (Johnson et al., Reference Johnson, Klapper and Sandberg1985, Reference Johnson, Klapper and Elrick1996). The occurrence of the last species in the ensensis Zone indicates that its development was unaffected by the initial Kačak Event (House, Reference House1985) corresponding to the If transgression at the kockelianus-ensensis zonal boundary. On the other hand, the If transgression could have facilitated and fostered the migration of I. orri through the ‘northern route’ around Euramerica to the European part of the continent.
Conclusions
The comparative study of abundant Eifelian icriodid assemblages from eastern Europe (Belorussian Basin) and North America (Michigan Basin) has permitted revision of the earlier concept of Icriodus orri (see Klapper and Barrick, Reference Klapper and Barrick1983) and related species. Morphological features in conjunction with stratigraphic and geographic considerations led to a more restricted definition of I. orri and its more rigorous separation from I. retrodepressus. The original concept of I. retrodepressus has been revised and its diagnosis emended by stressing the presence of a triangular and elongated spindle with a pronounced posterior depression and the basal cavity displaying a well-developed oblique spur on its inner side and a rounded posterior expansion on its outer side. Some forms originally included in Morphotype I of I. orri (of Klapper and Barrick, Reference Klapper and Barrick1983) are described herein as I. michiganus n. sp. The diagnostic features of the latter are the lachrymiform spindle with a wide, biconvex posterior part showing a short, shallow depression and a narrow, elongated, pointed anterior part, more or less curved inward. The basal cavity is nearly symmetrical with a rounded spur on its inner side.
Analysis of our collections and re-evaluation of published materials has made verification of the stratigraphic ranges of the studied icriodid species possible, and, for the first time, documentation of their significance for biostratigraphic correlation of shallow-marine strata. Icriodus retrodepressus ranges from the lowermost Eifelian (slightly above the base of the partitus Zone) to the upper (but not uppermost) part of the kockelianus Zone. Its appearance and dispersal might have been related to the transgressive/biotic Choteč Event (Walliser, Reference Walliser1996). Icriodus michiganus n. sp. appeared in the lowermost costatus Zone (and perhaps earlier in the partitus Zone) and disappeared close to the last occurrence of I. retrodepressus. Icriodus orri has the narrowest stratigraphic range, from the uppermost part of the kockelianus Zone to the top of the ensensis Zone (=top Eifelian). Its first appearance in the Michigan Basin coincides with the eustatic Ie transgression, whereas its migration to eastern Euramerica could have been related to the If transgressive pulse (=initial Kačak Event).
This study reaffirms the need for a comprehensive approach to a conodont species definitions, including not only unequivocal diagnostic morphology but also well-established stratigraphic and geographic constraints.
Acknowledgments
We thank M. Narkiewicz for discussion and help with preparation of the English manuscript, to J. Turczynowicz for performing computer graphics, and to L. Giro for taking the SEM photos. We are grateful to G. Klapper and T. Uyeno for their comments and suggestions that led to improvement of the manuscript. This research was supported by the Polish National Research Centre (grant no. UMO-2011/03/B/ST10/05468). It is a contribution to the IGCP Project 596 ‘Climate change and biodiversity patterns in the Mid-Palaeozoic.’






