



Introduction
Lower Devonian (Lochkovian) stromatoporoids have been described from several areas of North America, where in many cases they are highly abundant. These include those from New York (Stock, Reference Stock1991, Reference Stock1997), Virginia (Stock and Holmes, Reference Stock and Holmes1986), and Bathurst Island, Arctic Canada (Stearn, Reference Stearn1990). Additional previously undescribed stromatoporoid faunas are also present in Maine and Ellesmere Island, Arctic Canada. Habrostroma is the dominant stromatoporoid genus in all of these strata.
Much of the taxonomic history of the species involved was summarized by Stock and Burry-Stock (Reference Stock and Burry-Stock2001), who also summarized the history of the application of statistics to stromatoporoid systematics. They used multivariate statistics to analyze specimens of Habrostroma in the Lochkovian of New York, and determined that there were two closely related species, H. centrotum (Girty, Reference Girty1895) and H. consimile (Girty, Reference Girty1895). In the present study, their work is extended to include additional specimens from Virginia, Maine, Bathurst Island, and Ellesmere Island, and three species are described: H. centrotum, H. consimile, and H. microporum (Girty, Reference Girty1895). Only Habrostroma centrotum is found in all five areas, therefore both realms, which is highly unusual. For a single species to occur in two realms, which are defined by genera unique to just one, there must have been some sort of corridor across the inter-realm barrier.
Geography and stratigraphy
New York
Stromatoporoids were collected from 29 localities in the Manlius and Coeymans formations of New York, which according to Rickard (Reference Rickard1962, Reference Rickard1975) are Lochkovian. In eastern New York the specimens came from the Thacher Member of the Manlius Formation, and in central New York from the Thacher, Olney, Elmwood, Clark Reservation, and Jamesville members of the Manlius Formation, and the Dayville and Deansboro members of the Coeymans Formation (Fig. 1). In addition to the six Girty (Reference Girty1895) type specimens, the stratigraphic occurrence of which is not certain, 97 specimens, 77 from the Manlius and 20 from the Coeymans, are included in this study. Rickard (Reference Rickard1962, Reference Rickard1975) placed the Silurian-Devonian boundary within the Rondout Formation (Fig. 1.1); however, Ebert and Matteson (Reference Ebert and Matteson2003) indicated the Silurian-Devonian boundary lies above the Manlius Formation and the Dayville Member of the Coeymans Formation, and below the Deansboro Member of the Coeymans Formation (Fig. 1.2), rather than beneath the Manlius Formation, within the Chrysler Member of the Rondout Formation. We prefer the Rickard-based stratigraphy, but also recognize that Ebert and Matteson could be right. Neither scenario invalidates the conclusions of this paper.
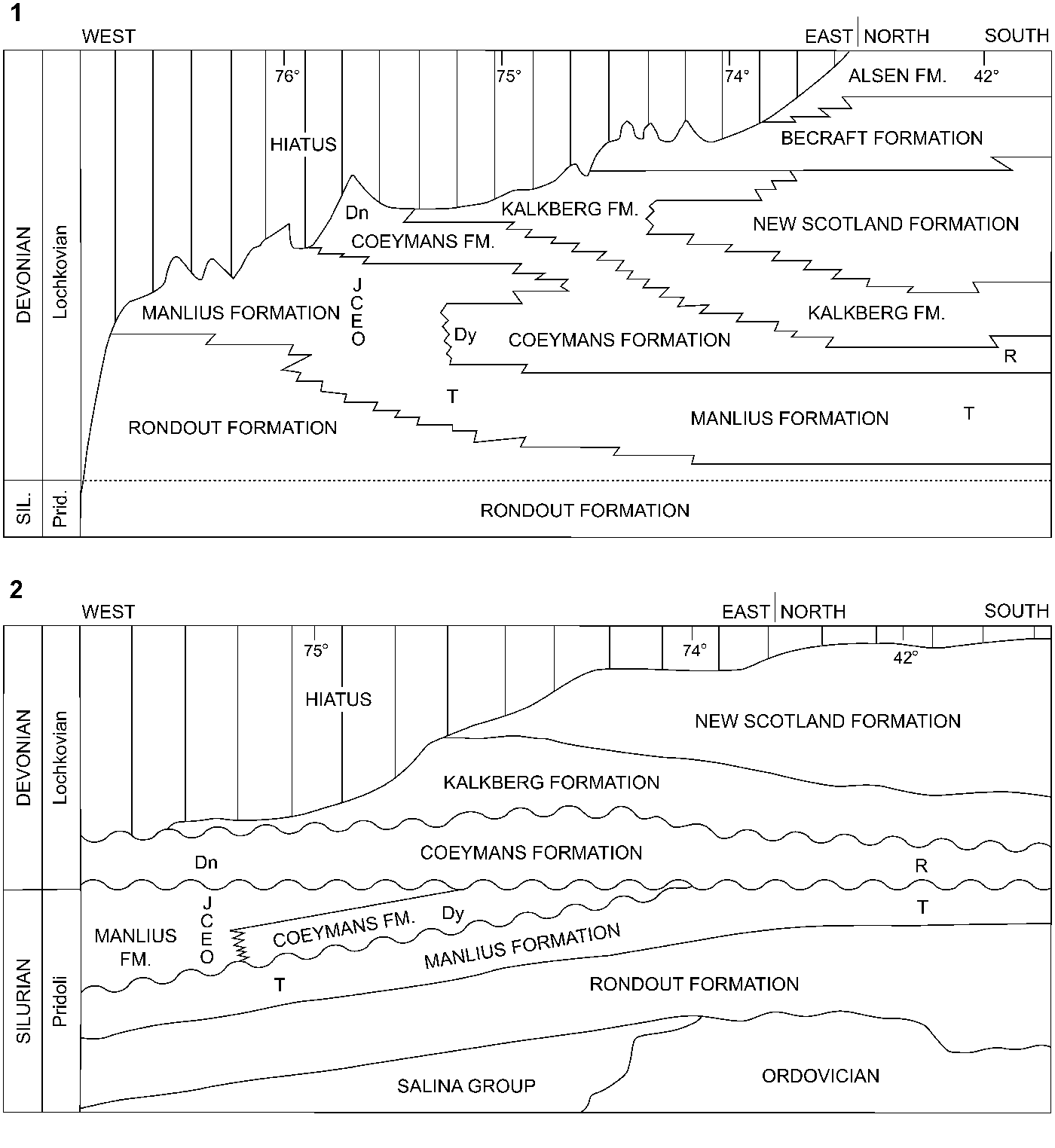
Figure 1. Stratigraphy of the Manlius and Coeymans formations and adjacent strata in central and eastern New York. (1) After Rickard (Reference Rickard1975); (2) after Ebert and Matteson (Reference Ebert and Matteson2003). Dn = Deansboro Mbr.; Dy = Dayville Mbr.; C = Clark Reservation Mbr.; E = Elmwood Mbr.; J = Jamesville Mbr.; O = Olney Mbr.; R = Ravena Mbr.; T = Thacher Mbr.
Virginia
At Mustoe, western Virginia, stomatoporoids are abundant in the Jersey Shore Member, and are common in the LaVale Member of the Keyser Formation (Fig. 2). The sedimentological setting of this locality was described by Smosna and Warshauer (Reference Smosna and Warshauer1979). At the time of their publication, the portion of the Keyser at Mustoe was thought to be Lower Devonian; however, Denkler and Harris (Reference Denkler and Harris1988) demonstrated that the Silurian-Devonian boundary is very high in the Keyser, close to the top of the Formation. Three specimens from probably uppermost Silurian and two specimens from probably lowermost Devonian are included in the present study—they had been called Parallelostroma barretti (Girty, Reference Girty1895) or P. cf. P. barretti (Girty, Reference Girty1895) by Stock and Holmes (Reference Stock and Holmes1986).
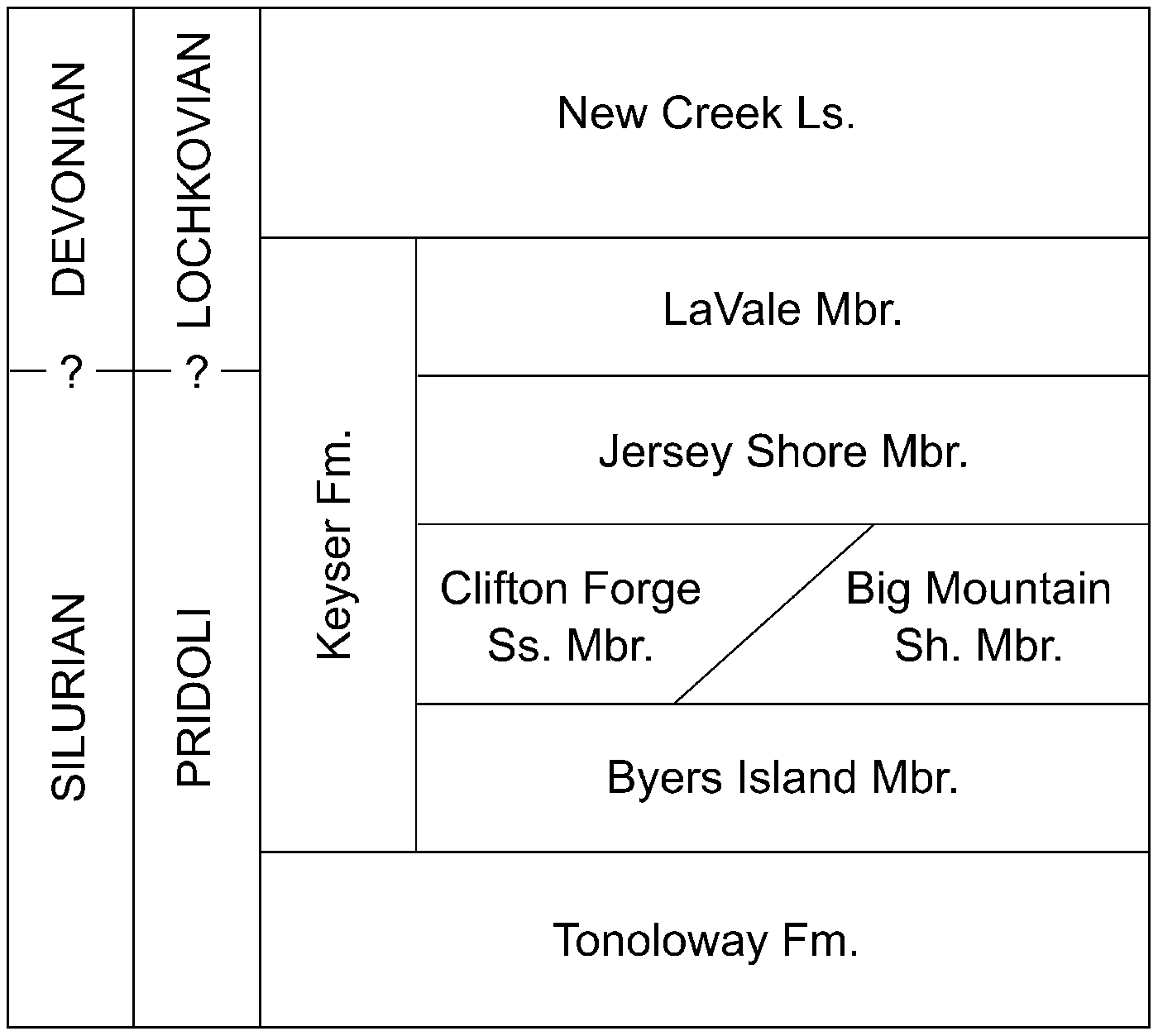
Figure 2. Stratigraphy of the Keyser Formation and adjacent strata at Mustoe, Virginia.
Maine
The Beck Pond Limestone occurs in a small area (~46 ha) of western Maine (Fig. 3.1), and lies unconformably on granitic basement (Boucot, Reference Boucot1961). The Beck Pond Limestone is located in an area in which, according to Williams and Hatcher (Reference Williams, Hatcher, Hatcher, Williams and Zietz1983), any of three suspect terranes are present: Dunnage; Gander; and Chain Lakes. They indicated that these terranes reached their current position relative to the craton of North America during the Early to Middle Ordovician. Zen (Reference Zen, Hatcher, Williams and Zietz1983, fig. 1) showed that the location of the Beck Pond Limestone is within the Chain Lakes massif, which he viewed as a possible suspect terrane that accreted during the Late Ordovician Taconic orogeny. Lavoie and Asselin (Reference Lavoie and Asselin2004) placed the Beck Pond area in either the Bronson Hill anticlinorium or Keasarge-Central Maine synclinorium tectonostratigraphic domains, whereas Malo (Reference Malo2004) included the Beck Pond area in the Boundary Mountain anticlinorium tectonostratigraphic zone. Beck Pond Limestone stratigraphy and environments of deposition were described by Boucot et al. (Reference Boucot, Harper and Rhea1959, Reference Boucot, Harper and Rhea1966), who divided the formation into five numbered members (in ascending order, 1–5). Stromatoporoids are present in all five members—the six specimens included in this study are from members 2, 4, and 5. The age of the Beck Pond Limestone is believed to be equivalent to that of the New Scotland Formation of eastern New York (Boucot et al., Reference Boucot, Harper and Rhea1959), which correlates with the upper parts of the Manlius and Coeymans formations in central New York (Rickard, Reference Rickard1975), or is found above the formations in eastern and central New York (Ebert and Matteson, Reference Ebert and Matteson2003).
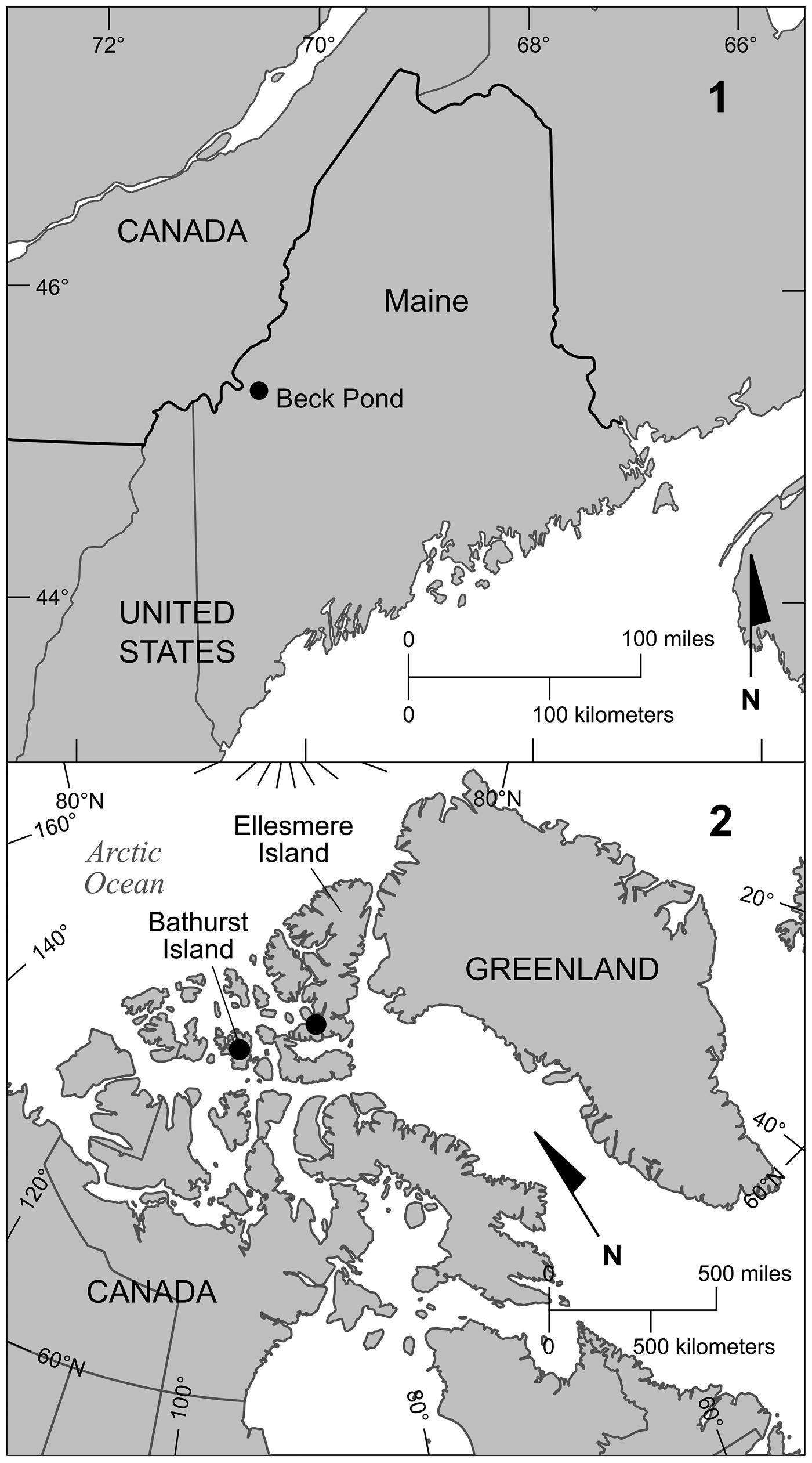
Figure 3. (1) Location of Beck Pond in western Maine. (2) Location of collecting areas on Bathurst Island and Ellesmere Island.
Bathurst Island
The Stuart Bay Formation on Bathurst Island, Arctic Canada (Fig. 3.2), is a Pragian–Emsian-age siltstone (Fig. 4.1) that contains limestone olistostromes that are interpreted to have been transported to their current location from a Lochkovian-age reef that developed on the Boothia Uplift (Polan and Stearn, Reference Polan and Stearn1984)—the source of these rocks was removed by subsequent erosion. Stromatoporoids and corals are abundant in the reef masses. Five of the 38 specimens of Habrostroma (as Parallelostroma microporum), included by Stearn (Reference Stearn1990) in his description of the Stuart Bay stromatoporoid fauna, are included in this study.

Figure 4. (1) Lower Devonian stratigraphy of Bathurst Island. (2) Upper Silurian and Lower Devonian stratigraphy of southwestern Ellesmere Island (after de Freitas and Mayr, Reference de Freitas, Mayr, Mayr, de Freitas, Beauchamp and Eisenbacher1998, fig. 4.2). G.F. = Goose Fiord Fm.; P.A. = Prince Alfred Fm.
Ellesmere Island
In 1989, T.A. de Freitas, then of the Geological Survey of Canada, collected 103 stromatoporoids from four measured sections of upper Silurian–Lower Devonian strata of the Goose Fiord Formation (Fig. 4.2) on southern Ellesmere Island, Arctic Canada (Fig. 3.2) (Norris, Reference Norris1996). The exact location of the Silurian-Devonian boundary within the stratigraphic sequence has not been determined. Eight specimens of Habrostroma, six from section 67 and two from section 63 (Appendix), are included in this study.
Other possibilities
Stromatoporoids in close proximity to the Silurian-Devonian boundary within the Roberts Mountains Formation of central Idaho were reported by Skipp and Sandberg (Reference Skipp and Sandberg1975). Although Morrow (Reference Morrow1984) and Morrow and Cook (Reference Morrow and Cook1987) reported stromatoporoids in the Lochkovian-age strata in the Root Basin and Prairie Creek Embayment of Northwest Territories, Canada, they apparently did not collect any specimens.
Stromatoporoid-bearing strata near the top of the West Point Formation of the Gaspé Peninsula of Quebec, Canada are believed to be lower Lochkovian (e.g., Bourque et al., Reference Bourque, Malo and Kirkwood2001). Pope (Reference Pope1986) described the stromatoporoids of the West Point Formation. Some of the specimens that she placed in Parallelostroma and Anostylostroma may belong to Habrostroma. Unfortunately, her collection is not available for study.
Materials, methods, and data analysis
Methods
Internal measurements and counts are made by means of a light microscope equipped with a calibrated ocular micrometer. Nine skeletal dimensions were obtained from each of the 127 specimens (Table 1).
Table 1. Skeletal dimensions of morphologies obtained from specimens of Habrostroma.

Counts of laminae per mm are begun in the middle of a lamina. If four additional laminae are included within the interval of 1 mm, the count is recorded as 5 laminae per mm. If four and a portion of a fifth additional lamina are included, the count is 5.5 laminae per mm. The same method is employed for counting microlaminae and pillars per mm. Determination of the percent of cyst-like microlaminae (Fig. 5) present in longitudinal sections is made from a single traverse from the top to the bottom of an individual thin section. Distance between pillars is taken from the centers of adjacent pillars, to avoid the effect of pillar diameter. Laminar thickness and gallery height are measured at the same time, to insure proper proportions between the two when compared.

Figure 5. Photomicrograph of longitudinal section of a portion of Habrostroma consimile (Girty) USNM 408733 from the Thacher Member of the Manlius Formation of New York. Note presence of convex-upward localized cyst-like microlaminae. Scale bar = 0.5 mm.
At least 20 measurements or counts are taken for each character in most specimens. In a few specimens the small size or poor preservation of the skeleton precludes a minimum of 20. Where mean value for a particular character in a species is given, the mean of the means (the grand mean) obtained for each specimen is treated equally, thus insuring equal weighting of each specimen, regardless of this size.
Data analysis
The statistical analyses employed here (cluster analysis, discriminant analysis, canonical correlation) are described in detail by Stock and Burry-Stock (Reference Stock and Burry-Stock2001), so only a brief summary will be given here. Cluster analysis answers the question, “How many species are there in these 127 specimens?” Discriminant analysis answers the question, “Which among the nine morphologies obtained contribute most to species separation?” Canonical correlation tells us to what degree the nine morphologies and species membership are related. These analyses were run on the microcomputer version of SPSS 11.0.
We used the average linkage cluster analysis procedure to determine how many groups are in this data set. We found that with 127 specimens, measured on nine morphologies, there are three groups (Fig. 6). Figure 7 is the resulting dendrogram that illustrates the average linkage of how the specimens separate into three groups. Each specimen is located on the vertical axis, and the graphic representation of the rescaled Euclidean distance is located on the horizontal axis on a scale of 0 to 25. Thus, the first stage of the data analysis suggests that there are three groups/species.
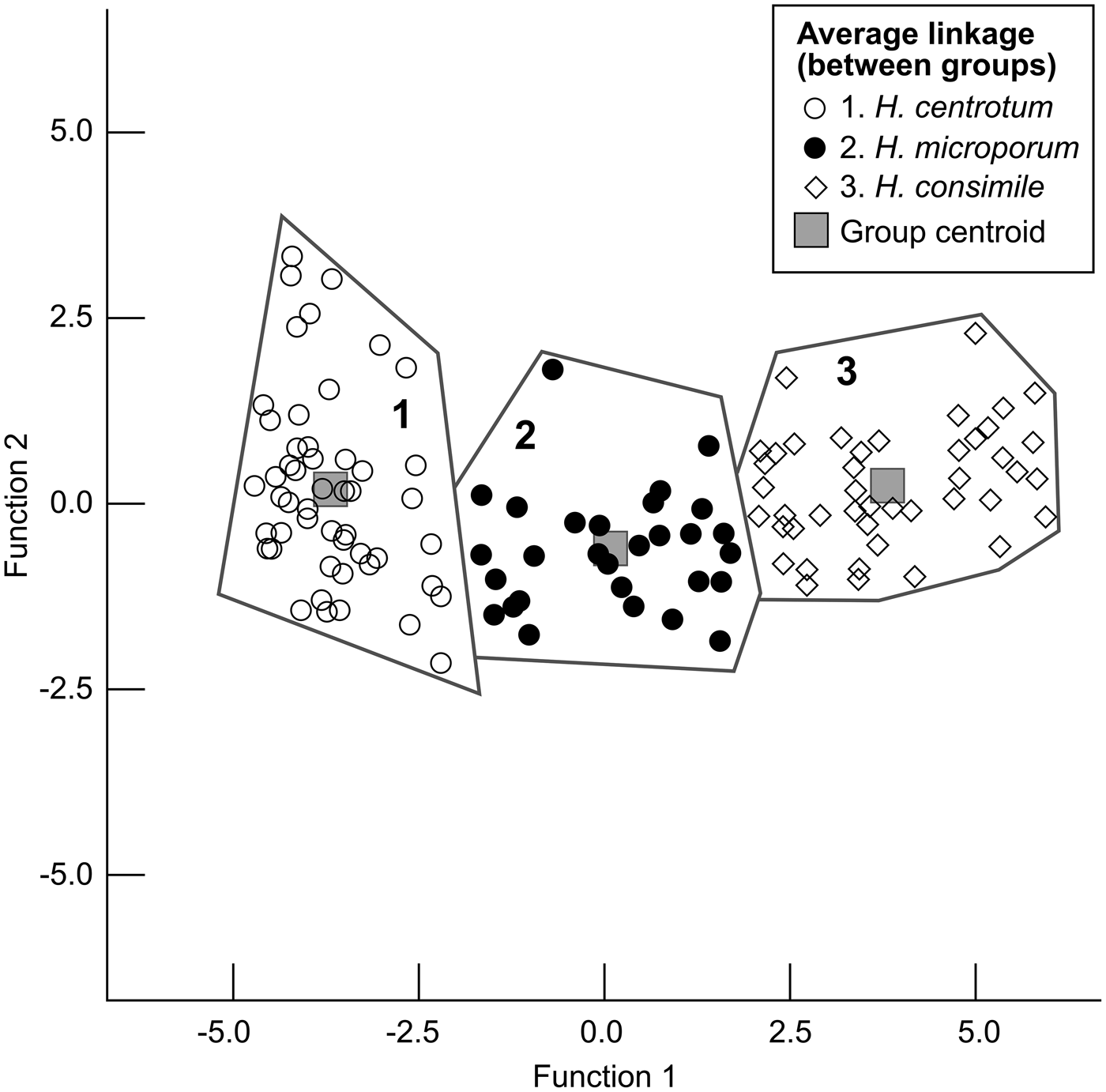
Figure 6. Discriminant analysis on the 127 studied specimens of Habrostroma. Function 1 represents percent cyst-like microlaminae, and Function 2 represents microlaminae/mm.
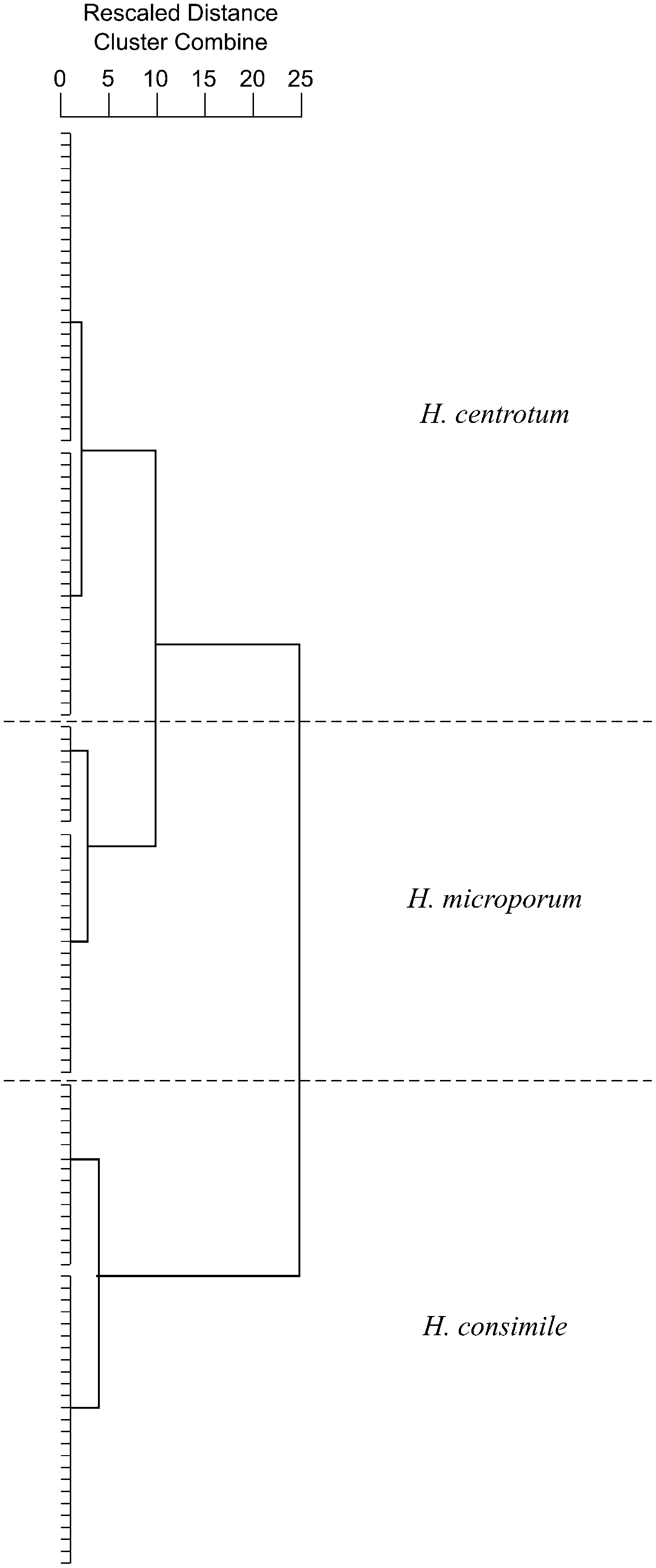
Figure 7. Dendrogram resulting from average linkage cluster analysis.
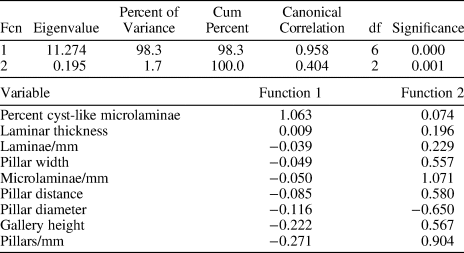
Group membership from the three groups was retained from the cluster analysis and entered with the skeletal dimensions of nine morphologies into a three-group discriminant analysis. The discriminant functions are listed in Table 2 and are standardized, which means that all of the morphologies have a mean of 0 and a standard deviation of 1. These weights are used to predict membership in one of the three groups.
Table 2. Canonical discriminant function and standardized canonical discriminant function coefficients.
Fcn = function; df = degrees of freedom.
The overall statistic for this analysis is Wilks’ lambda, also called the U statistic, which is 0.219 where p < 0.0000. Wilk's lambda and the F test for each morphology are detailed in Table 3. The F tests answer a related research question: “Which morphologies contribute to group separation?” Five variables are statistically significant at alpha = 0.01: percent cyst-like microlaminae; microlaminae/mm; gallery height; laminae/mm; and laminar thickness. These tests imply that these five morphologies are doing most of the prediction of group membership of the three species. Using the level of significance, it is easy to tell the magnitude of the contribution, which from greatest to least are: (1) percent cyst-like microlaminae, (2) microlaminae/mm, (3) laminae/mm, (4) laminar thickness, (5) gallery height, (6) pillar width, (7) pillars/mm, (8) pillar diameter, and (9) pillar distance.
Table 3. Discriminant analysis Wilks’ lambda = 0.068 where p < 0.000, .837 where p < 0.006, and univariate F-ratio for morphologies.

* p < 0.01
Canonical correlation analysis is used as a procedure that examines the interdependence between two sets of variables, the predictor variables (independent variables), and the criterion variables (dependent variables) (Pedhazur, Reference Pedhazur1997). In our case, the predictor variables are the morphologies and the criterion variables are the group memberships in group 1, group 2, or group 3. The canonical correlations for our three groups and nine morphologies are 0.958 and 0.404, where p < 0.000 and 0.001, respectively. The canonical correlation is interpreted as a simple correlation coefficient, where a value of 0 implies that there is no relationship between variables, and 1 implies there is a perfect relationship. There is only one set of predictor variables in this analysis, which is labeled Function 1 in Table 2. These functions are standardized, which means that they have a mean of 0 and a standard deviation of 1. For Function 1, the order from greatest to least is in determining the interdependence between group membership and the morphologies are: percent cyst-like microlaminae; laminar thickness; laminae/mm; pillar width; microlaminae/mm; pillar distance; pillar diameter; gallery height; and pillars/mm. For Function 2, the order from greatest to least is: microlaminae/mm; pillars/mm; pillar distance; gallery height; pillar width; laminae/mm; laminar thickness; percent cyst-like microlaminae; and pillar diameter. Note that the order of magnitude is different here because this listing describes the interdependence of the predictor variables’ values associated with the prediction.
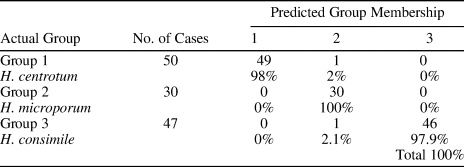
Table 4 is the result of discriminant analysis for predicting group membership from the nine morphologies. There was a total of 127 specimens included in the analysis. No cases were excluded for missing or out-of-group codes. Of the specimens that were grouped in group 1, 98% were grouped accurately, of the specimens that were grouped in group 2, 100% were classified correctly, and of the specimens that were grouped in group 3, 97.9% were classified correctly. The total classification was 100%. This value is sometimes referred to as the “hit rate” for the analysis, and it provides yet another check on the accuracy of predicting three groups from the nine morphologies.
Table 4. Classification of Habrostroma centrotum, H. microporum, and H. consimile.
Wolniewicz (Reference Wolniewicz2010, Reference Wolniewicz2013) took issue with some of the standard skeletal measurement and data interpretation employed by previous stromatoporoid workers. He was able to detect two species of Gerronostromaria from the Upper Devonian (Famennian) of Poland, using image analysis software. Integral to the latter was the inclusion of stereological data (Wolniewicz, Reference Wolniewicz2013). His techniques are fascinating, and show some potential for stromatoporoid identification. Unfortunately, he is the only worker who has employed this technique, and did so on only one genus, the rectilinear Gerronostromaria. Wolniewicz acknowledged some shortcomings in his method: The skeleton must have a network of horizontal laminae and vertical pillars (Wolniewicz, Reference Wolniewicz2010, p. 555), and the results he obtained are, “… valid for only the populations under study” (Wolniewicz, Reference Wolniewicz2013, p. 955). We have taken the position that the statistics we employed are useful and valid. Wolniewicz (Reference Wolniewicz2013, p. 954) agreed with us, stating, “The value of traditional features also increases when multivariate techniques are used, e.g. a canonical discriminant analysis.”
Repositories and institutional abbreviations
GSC, Geological Survey of Canada, Ottawa, Ontario, Canada; RN, Redpath Museum, McGill University, Montreal, Quebec, Canada; USNM, U.S. National Museum of Natural History, Smithsonian Institution, Washington, D.C., U.S.A.; YPM, Yale Peabody Museum, New Haven, Connecticut, U.S.A.
Systematic paleontology
Phylum Porifera Grant, Reference Grant and Todd1836
Class Stromatoporoidea Nicholson and Murie, Reference Nicholson and Murie1878
Order Syringostromatida Bogoyavlenskaya, Reference Bogoyavlenskaya1969
Family Coenostromatidae Waagen and Wentzel, Reference Waagen, Wentzel, Waagen and Salt-Range Fossils1887
Genus Habrostroma Fagerstrom, Reference Fagerstrom1982
Type species
Stromatopora proxilaminata Fagerstrom, Reference Fagerstrom1961, p. 8, pl. 1, figs. 4–6; Middle Devonian (Eifelian), Formosa Reef Limestone, Formosa, Ontario, Canada.
Diagnosis
“Laminae continuous, composed of one or more microlaminae, connected by regular to irregular pillars; pillars may be superposed. Phases may be developed in conjunction with latilaminae; where laminae far apart coenostromes and coenosteles present. Typically laminae and coenostromes thinner than galleries high. Microstructure acosmoreticular” (Stock, Reference Stock1997, p. 545).
Remarks
The recommended terms pachystrome and pachystele have replaced coenostrome and coenostele, respectively (Webby, Reference Webby and Selden2015, p. E409, E410).
Disagreement persists as to which genus the species described in this paper belong. Stearn (Reference Stearn1990) placed his specimens from Bathurst Island in Parallelostroma microporum (Girty); Stock (Reference Stock1988) had questionably assigned this species to Parallelostroma, but later reassigned it to Habrostroma (Stock, Reference Stock1991). Stearn (Reference Stearn2001) changed his species assignment of the Bathurst Island specimens to Parallelostroma centrotum (Girty). The five Bathurst Island specimens included in this paper (GSC 95774–95776; RM 20.6801, 20.6802), and other specimens examined by Stock that were not included, all contain skeletal macrostructures consisting of acosmoreticular microstructure, and galleries that are higher than laminae are thick, both characteristics of Habrostroma. Parallelostroma is characterized by orthoreticular microstructure (Stearn et al., Reference Stearn, Webby, Nestor and Stock1999, p. 54; Stearn, Reference Stearn and Selden2015, p. E819), and laminae that are thicker than galleries are high.
Careful reading of Stearn (Reference Stearn and Selden2015, p. E800–E820) reveals characteristics of genera that are similar to Habrostroma, but which differ in some traits. Briefly, most other genera of the same age can be differentiated from Habrostroma on the basis of skeletal microstructure. Stromatopora and Syringostromella have cellular, not microreticulate, microstructure. The dominant microstructure in Parallelostroma is orthoreticular, and Atopostroma contains a mixture of orthoreticular and clinoreticular microstructure. The microstructure of the pillars in Columnostroma is clinoreticular, as is that in the long subcolumns of Syringostroma. Only Habrostroma and what Stock (Reference Stock1997) incorrectly called Coenostroma of the Pridoli-Lochkovian stromatoporoids have acosmoreticular microstructure. “Coenostroma” has more robust pachystromes and pachysteles relative to Habrostroma; both long and short pachysteles are distributed throughout the skeleton of “Coenostroma,” whereas the thinner pachysteles in Habrostroma are distributed in laminar zones.
Habrostroma centrotum (Girty, Reference Girty1895)
Figure 8
- Reference Girty1895
Syringostroma centrotum Girty, p. 293, figs. 1, 2.
- Reference Fagerstrom1982
Habrostroma centrotum; Fagerstrom, p. 13.
- Reference Stock and Holmes1986
Parallelostroma barretti; Stock and Holmes, p. 564, figs. 4.7, 4.8, 5.1–5.3.
- Reference Stearn1990
Parallelostroma microporum; Stearn, p. 506, figs. 4.8, 7.1–7.4, 8.4.
- Reference Stock1991
Habrostroma centrotum; Stock, p. 905, fig. 5.1–5.7.
- Reference Stock1997
Habrostroma centrotum; Stock, p. 545, fig. 4.1, 4.2.
- Reference Stearn2001
Parallelostroma centrotum; Stearn, p. 22.

Figure 8. Photomicrographs of specimens of Habrostroma centrotum (Girty) from Maine, Bathurst Island, and Ellesmere Island, respectively, none of which has been illustrated previously. Difference in appearance may exist, due to differences in preservation; skeletal dimensions are not affected by this. (1, 2) USNM 526377 from Maine; (1) longitudinal section; (2) tangential section. (3, 4) USNM 526380 from Maine; (3) longitudinal section; (4) tangential section; (5, 6) RM 20.6801 from Bathurst Island; (5) longitudinal section; (6) tangential section. (7, 8) GSC 10767 from Ellesmere Island; (7) longitudinal section; (8) tangential section. Bar scales = 1.0 mm.
Holotype
YPM 22449 collected from the “Lower Pentamerus by Girty, 1985 limestone” at “Cedarville, N.Y.” (Girty, Reference Girty1895, p. 295). The exact collecting locality and stratum could not be determined, but could be close to Locality E of Stock (Reference Stock1988), at which the stromatoporoid-bearing lithostratigraphic unit is the Clark Reservation Member of the of the Manlius Formation.
Occurrence
Uppermost Silurian (upper Pridoli) or lowermost Devonian Jersey Shore Member of the Keyser Formation of Virginia; and lower Lower Devonian (Lochkovian) LaVale Member of the Keyser Formation of Virginia; Thacher, Olney, Elmwood, Clark Reservation, and Jamesville members of the Manlius Formation, and Deansboro Member of the Coeymans Formation of New York; Beck Pond Limestone of Maine; Stuart Bay Formation of Bathurst Island; and Goose Fiord Formation of Ellesmere Island (Fig. 9).
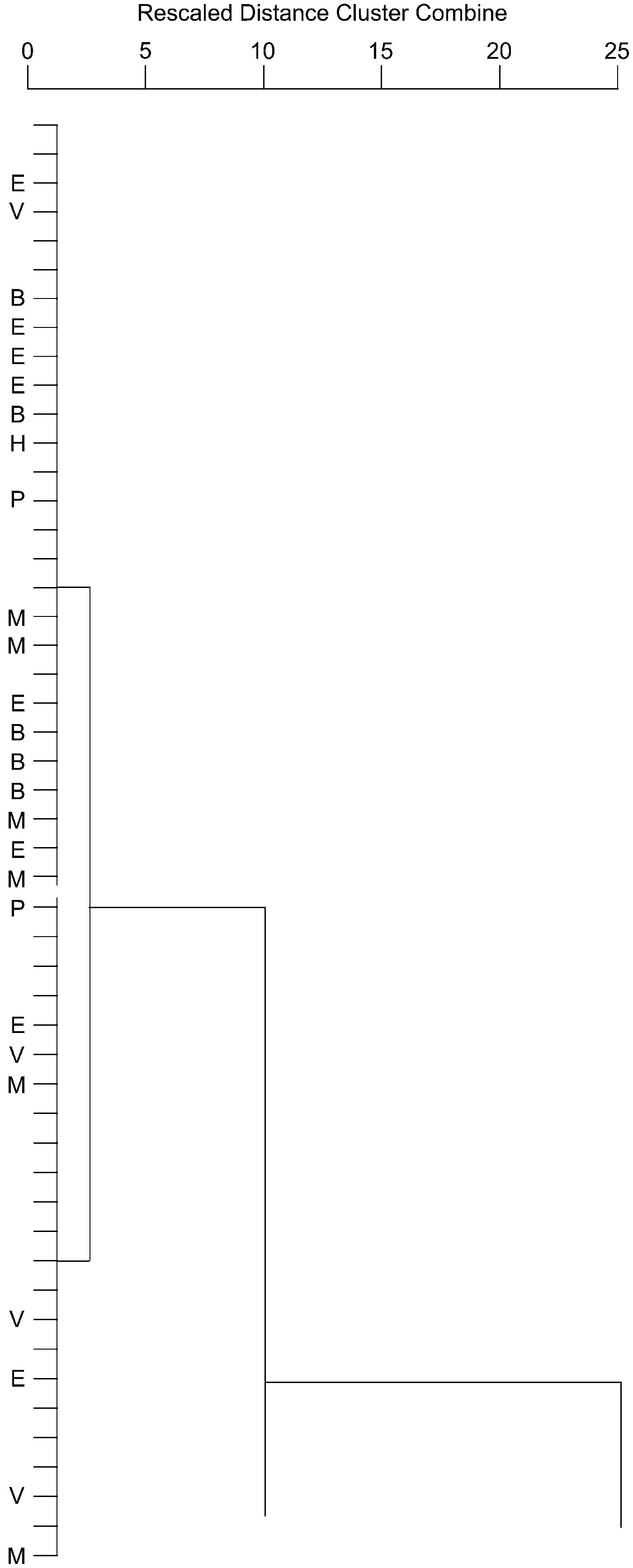
Figure 9. Portion of Figure 7 representing Habrostroma centrotum. H = holotype; P = paratype; B = Bathurst Island; E = Ellesmere Island; M = Maine; V = Virginia; remaining unlabeled specimens from New York.
Description
Combined skeletal dimensions for the 50 specimens included in this study are given in Table 5.
Table 5. Combined skeletal dimensions (in mm) of 50 specimens of Habrostroma centrotum (Girty, Reference Girty1895) included in this study.

R = total range; r = range of means; $\bar{\rm x}$ = grand mean.
= grand mean.
Materials
Holotype, YPM 22449 (New York); paratypes, YPM 22419, 22420 (New York); hypotypes, YPM 22454 (New York); USNM 364708-364711 (Virginia); USNM 408724–408728, 432830, 446774, 446786, 446787 (New York); USNM 526376–526381 (Maine); GSC 95774–95776 (Bathurst Island); GSC 10762–10769 (Ellesmere Island); RM 20.6801, 20.6802 (Bathurst Island); and 14 uncatalogued specimens (New York).
Remarks
For additional photomicrographs of Habrostroma centrotum, the reader is referred to the work of Stock and Holmes (Reference Stock and Holmes1986), Stock (Reference Stock1988, Reference Stock1991, Reference Stock1997), and Stearn (Reference Stearn1990).
Of the three species, Habrostroma centrotum is characterized by skeletons containing the lowest development of cyst-like microlaminae (0–22%; mean 7.3%). It also has the most laminae per mm (6.5) and the most microlaminae per mm (7.3).
Habrostroma microporum (Girty, Reference Girty1895)
Figure 10.1, 10.2
- Reference Girty1895
Syringostroma microporum Girty, p. 296, pl. 6, fig. 7.
- Reference Fagerstrom1982
Habrostroma microporum; Fagerstrom, p. 13.
- Reference Fagerstrom1991
Habrostroma microporum; Stock, part, p. 906, fig. 7.7, 7.8, not fig. 7.3–7.6.
- Reference Stock1997
Habrostroma microporum; Stock, p. 546, not fig. 4.7, 4.8.

Figure 10. (1, 2) Photomicrographs of a specimen of Habrostroma microporum (Girty) from New York; USNM 446789. (1) Longitudinal section; (2) tangential section. (3, 4) Photomicrographs of a specimen of Habrostroma consimile (Girty) from New York; USNM 446790. (3) Longitudinal section; (4) tangential section. Scale bars = 0.5 mm.
Holotype
YPM 9195 collected from the “Lower Pentamerus by Girty, 1895 limestone” at “Cedarville, N.Y.” (Girty, Reference Girty1895, p. 296). The exact locality and stratum could not be determined, but could be close to Locality E of Stock (Reference Stock1988), at which the stromatoporoid-bearing lithostratigraphic unit is the Clark Reservation Member of the Manlius Formation.
Occurrence
Lower Lower Devonian (Lochkovian) Thacher, Olney, Elmwood, Clark Reservation, and Jamesville members of the Manlius Formation, and Dayville and Deansboro members of the Coeymans Formation of New York.
Description
Combined skeletal dimensions for the 30 specimens included in this study are given in Table 6.
Table 6. Combined skeletal dimensions (in mm) of 30 specimens of Habrostroma microporum (Girty, Reference Girty1895) included in this study.

R = total range; r = range of means; $\bar{\rm x}$ = grand mean.
= grand mean.
Materials
Holotype YPM 9195 (New York); hypotypes USNM 408730–408732, 408734, 408735, 446789 (New York); and 23 uncatalogued specimens (New York).
Remarks
For photomicrographs of Habrostroma microporum, the reader is referred to the work of Stock (Reference Stock1988, Reference Stock1991).
Of the three species, Habrostroma microporum is characterized by skeletons containing moderate development of cyst-like microlaminae (26–58%; mean 46.2%). It also has the fewest laminae per mm (5.8) and the fewest microlaminae per mm (4.7).
Habrostroma consimile (Girty, Reference Girty1895)
Figures 5, 10.3, 10.4
- Reference Girty1895
Syringostroma consimile Girty, p. 297, pl. 7, figs. 3, 4.
- Reference Stock1991
Habrostroma microporum; Stock, part, fig. 7.3–7.6, not fig. 7.7, 7.8.
- Reference Stock1997
Habrostroma microporum; Stock part, fig. 4.7, 4.8.
Holotype
YPM 5527 collected from the “Lower Pentamerus by Girty, 1985 limestone” at the “outlet of Skaneateles Lake, N.Y.” (Girty, Reference Girty1895, p. 298). Locality I of Stock (Reference Stock1988) in Skaneateles Falls is the stromatoporoid-bearing outcrop closest to the outlet of Skaneateles Lake; stromatoporoids are found in both the Olney and Jamesville members of the Manlius Formation.
Occurrence
Lower Lower Devonian (lower Lochkovian) LaVale Member of the Keyser Formation of Virginia; and lower Lower Devonian (Lochkovian) Thacher, Olney, Elmwood, Clark Reservation, and Jamesville members of the Manlius Formation, and Dayville and Deansboro members of the Coeymans Formation of New York.
Description
Combined skeletal dimensions for the 48 specimens included in this study are given in Table 7.
Table 7. Combined skeletal dimensions (in mm) of 48 specimens of Habrostroma consimile (Girty, Reference Girty1895) included in this study.
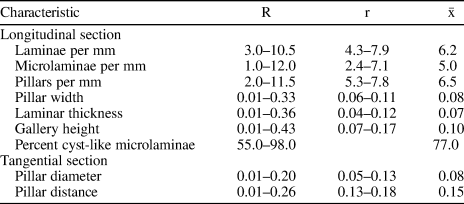
R = total range; r = range of means; $\bar{\rm x}$ = grand mean.
= grand mean.
Materials
Holotype YPM 5527 (New York); hypotypes USNM 364712 (Virginia); 408729, 408733, 446790, 446791, 482831 (New York); and 41 uncatalogued specimens (New York).
Remarks
For photomicrographs of Habrostroma consimile, the reader is referred the work of Stock and Holmes (Reference Stock and Holmes1986) and Stock (Reference Stock1988, Reference Stock1991, Reference Stock1997).
Of the three species, Habrostroma consimile is characterized by skeletons containing the highest development of cyst-like microlaminae (55–98%; mean 77%).
Systematic results
Composition of the three groups defined by the average linkage cluster analysis does not coincide exactly with the two species that were determined by Stock (Reference Stock1991, Reference Stock1997) for New York. In those earlier studies, the holotype of Habrostroma centrotum was included in one species-group, and the holotypes of H. microporum and H. consimile were part of a second species-group. Because H. microporum is the senior synonym of the latter two species, the second group bore its name. Stock and Burry-Stock (Reference Stock and Burry-Stock2001) also determined that there were two species from New York, H. centrotum and H. consimile, because the holotype of H. microporum was classified with H. centrotum.
The additional 24 specimens added here from Virginia, Maine, Bathurst Island, and Ellesmere Island, to the 103 examined by Stock and Burry-Stock (Reference Stock and Burry-Stock2001), accentuated the differences among specimens in this enlarged collection. This is especially true for the discrimination of Habrostroma microporum from H. consimile. The three species groups (Fig. 6) do not overlap, but are close to each other, which may represent that H. centrotum gave rise to H. microporum and H. consimile in the Appalachian Basin.
Paleobiogeography
Realms
The tropical seas of the Early Devonian are divided into two faunal realms: (1) the Eastern Americas Realm (EAR), consisting of maritime Canada, the eastern United States, and northeastern South America; and (2) the Old World Realm (OWR), consisting of western and northern North America, Europe, Asia, and eastern Australia. These realms were separated by a North American land barrier, commonly known as the Transcontinental Arch (Stock et al., Reference Stock, Nestor, Webby and Selden2015), and by the Canadian Shield. Realms are defined by unique assemblages of genera (Servais et al., Reference Servais, Cecca, Harper, Isozaki, Mac Niocaill, Harper and Servais2013). According to Stock (in Stock et al., Reference Stock, Nestor, Webby and Selden2015), during the Lochkovian, the EAR contained four unique stromatoporoid genera: Stelodictyon, Intexodictyides (as Intexodictyon), Columnostroma, and Coenostroma. The OWR contained 15 unique genera. Five genera occurred in both realms, including Habrostroma.
Factors controlling stromatoporoid paleobiogeography
Stromatoporoids were sessile benthic sponges with a calcareous exoskeleton that lived from the Middle Ordovician to the Late Devonian. Generally, the environment they inhabited displayed several characteristics: (1) warm water, tropical to subtropical latitudes; (2) normal salinity; (3) shallow subtidal depth, some of the shallowest known in the Silurian and Devonian; (4) medium to high turbulence, increased dissolved O2 in seawater; (5) firm substrate, colonization sites for larvae; and (6) low turbidity, which precluded clay-size particles from clogging the aquiferous canal systems, and allowed sunlight for photosymbionts, if such were the case. Stromatoporoids are known to be the principal reef-builders of their time, but the majority of them inhabited level-bottom environments.
The characteristics noted above do not appear to provide much potential for stromatoporoids as index fossils. They were sessile benthic, could not tolerate a soft seafloor, nor most typical siliciclastic depositional environments. Their depth range was relatively narrow. The use of biostratigraphy for chronostratigraphic purposes is typically limited to planktonic (e.g., certain foraminiferans, graptoloid graptolites) or nektonic (e.g., ammonoid cephalopods, conodonts) animals because these organisms are not dependent on depth nor particular substrate conditions. In cases where plankton and nekton are absent, benthic organisms with high population numbers and wide environmental tolerances, such as brachiopods, have proven useful.
One way that sessile benthic organisms can spread their geographic range is through the dispersal of planktonic larvae. According to Thorson (Reference Thorson1971, p. 79) ~74% of marine benthic animals today produce planktonic larvae. Bergquist (Reference Bergquist1978, p. 106) stated that many living sponges produce free-swimming larvae that remain in the plankton for 3–48 hours.
Stromatoporoids, as dwellers in shallow-water environments, benefit when eustatic sea level is high, and large land areas are flooded by shallow seas. When seas are especially high, land-area barriers to migration of marine organisms are compromised. Such a situation existed, beginning with the Taghanic onlap mid-way in the Givetian, when there was no longer a separation between the OWR and EAR. On the other hand, eustatic sea level was especially low during the Early Devonian and earliest Middle Devonian (Johnson et al., Reference Johnson, Klapper and Sandberg1985).
The close of the Lochkovian Age coincides with the end of the Tippecanoe sequence of Sloss (Reference Sloss1963), and the beginning of his Kaskaskia sequence (Johnson et al., Reference Johnson, Klapper and Elrick1996). At this time, eustatic sea level began to rise, but again fell at the end of the Pragian, almost as much as at the beginning of the Pragian. Elrick et al. (Reference Elrick, Berkyová, Klapper, Sharp, Joachimski and Fryda2009, p. 179) concluded that Early–Middle Devonian changes in sea level were the result of My-scale sequences, which were caused by “paleoclimatically driven glacio-eustasy and thermo-eustasy.”
Johnson (Reference Johnson1971) introduced a concept he called the “Antler effect.” Essentially, the idea is that when the margins of a tectonic plate are forced upward by orogeny, the central part of the plate sinks downward in order to restore isostatic equilibrium (epeirogeny). He noted three tectophases that affected the margins of North America in the Devonian, which were associated with the Antler Orogeny of the West, what is now called the Ellesmerian Orogeny of the North, and the Acadian Orogeny of the East. When there was no orogeny, there was epeirogenic uplift in the midcontinent, causing seas to retreat to the continental margins. A year later, Johnson (Reference Johnson1972) realized his idea was not new—he had independently rediscovered the “Haug effect,” which was published 70 years earlier.
North America was tectonically quiescent from the late Silurian (Ludlow Epoch) (Ettensohn and Brett, Reference Ettensohn, Brett, Landing and Johnson1998) through the Early Devonian, as noted above. No Silurian differentiation into the EAR and OWR was detected for brachiopods (Boucot and Johnson, Reference Boucot, Johnson and Hallam1973), rugose corals (Oliver, Reference Oliver1977), or stromatoporoids (Nestor, Reference Nestor, McKerrow and Scotese1990). The effect of Early Devonian tectonism no doubt led to a withdrawal of the sea from North America, a consequence that was enhanced by the exceptionally low sea level associated with the Tippecanoe-Kaskaskia transition.
Early Devonian paleobiogeography
Most workers (e.g., Johnson and Boucot, Reference Johnson, Boucot and Hallam1973; Oliver, Reference Oliver1977) invoked the presence of a land barrier in North America that separated the EAR from the OWR during the Early Devonian. This barrier was composed of the Canadian Shield and a connected ridge that extended from Minnesota southwest to Arizona (Fig. 11.1), known as the Transcontinental Arch. This situation persisted through the Eifelian, but to a progressively lesser degree.

Figure 11. Lochkovian paleogeography and paleobiogeography of North America. (1) Map with location of the Old World Realm, Eastern Americas Realm, Canadian Shield, Transcontinental Arch, based on work of Oliver (Reference Oliver1977). Circles represent location of the five areas from which specimens described in this paper were collected, and of three kimberlite pipes (circles with dots) containing Paleozoic xenoliths. (2) Map showing the proposed location of a shallow seaway across the Canadian Shield.
Oliver (Reference Oliver1977, p. 89–92) summarized the history of ideas regarding Devonian paleobiogeography, especially for North America. The “modern era” of Devonian paleobiogeography began over 50 years ago, with the work of Boucot et al. (Reference Boucot, Johnson, Talent and Oswald1968, Reference Boucot, Johnson and Talent1969). They were the first to elucidate the three Devonian faunal realms, based on the distribution of brachiopods; however, they called the realms provinces, and the EAR was called the Appalachian Province. Oliver (Reference Oliver1977) recognized the three biogeographic areas, and was the first to call them realms, coining the term Eastern Americas Realm. His findings were based on the distribution of Devonian rugose corals. His acceptance of the tripartite division of the Devonian marine faunas remained essentially unchanged in subsequent years (e.g., Oliver and Pedder, Reference Oliver, Pedder, Gray and Boucot1979, Reference Oliver and Pedder1984, Reference Oliver and Pedder1989).
Devonian conodont paleobiogeography was discussed by Klapper and Johnson (Reference Klapper and Johnson1980), who concluded that conodonts were absent from the Malvinokaffric Realm, but were widespread in the OWR and EAR. They did not see genus-level differences between the two tropical-subtropical realms, but noted endemism at the species level, especially among those in the near-shore conodont biofacies. Most stromatoporoids occur in near-shore facies.
Blodgett et al. (Reference Blodgett, Rohr, Boucot, McKerrow and Scotese1990, p. 277) found that the distribution of gastropods in the Early and Middle Devonian were “remarkably similar” to those of the brachiopods, rugose corals, and trilobites. According to Stock (Reference Stock, McKerrow and Scotese1990), stromatoporoid distributions supported separate OWR and EAR during the Early Devonian, although stromatoporoids vacated the EAR and North American portions of the OWR during the Pragian.
Breaching the barrier
Cox and Moore (Reference Cox and Moore2000, p. 37) described three kinds of pathways across barriers. The first is a corridor, which they defined as an easily traversed pathway for the majority of organisms at both ends—one that contains a wide variety of habitats. The second type, the filter, contains fewer habitats, restricting organisms that traverse it to those that can inhabit the limited number of habitats. The most restrictive pathway is the sweepstakes route, where two isolated, but similar, habitats are connected for a short period of time; normally the entire life history of an organism is not spent by this means (e.g., only planktonic larvae cross the barrier).
Some hints of a breaching of the inter-realm barrier have been published. Sandberg (Reference Sandberg1970) proposed a northwest-southeast oriented strait, which was a graben crossing the Transcontinental Arch, providing a marine connection between the Williston Basin (OWR) and the North Kansas Basin (EAR). He believed the strait existed from the Ordovician through the Mississippian. Sandberg (personal communication, 2018) saw the strait as an inter-realm breach because Devonian fossils in the Williston Basin also occurred in the Cedar Valley Formation of Iowa. Since then, the Cedar Valley has been raised to group status (e.g., Day and Witzke, Reference Day, Witzke and Montenari2017), including three formations in ascending order: Little Cedar Formation, Coralville Formation, and Lithograph City Formation. At the time of Sandberg's (Reference Sandberg1970) work, the term Cedar Valley Formation applied to the Givetian-age Little Cedar and Coralville formations (e.g., Koch, Reference Koch1970), which were deposited immediately following the Taghanic Onlap. The strait, now named the South Dakota-Nebraska Strait, appeared in some of Sandberg's subsequent work as the Paleozoic Strait without any explanation (e.g., Sandberg et al., Reference Sandberg, Poole, Johnson, McMillan, Embry and Glass1989; Johnson et al., Reference Johnson, Sandberg, Poole, Cooper and Stevens1991).
In a recent publication, McCormick et al. (Reference McCormick, Chamberlain and Paterson2018) described a series of faults in southeastern South Dakota and southwestern Minnesota oriented northwest-southeast. They believed the faults date back to the late Mesoproterozoic, temporally related to the Midcontinent Rift. It is possible that these faults are related to the similarly oriented South Dakota-Nebraska Strait. In another recent publication, Stein et al. (Reference Stein, Stein, Elling, Keller and Kley2018) illustrated a revised and extended map of the Midcontinent Rift.
Chatterton and Perry (Reference Chatterton and Perry1977) reported that Alberti et al. (Reference Alberti, Haas and Ormiston1972) suggested the Lochkovian trilobite Warburgella rugulosa eureka Alberti, Haas, and Ormiston, Reference Alberti, Haas and Ormiston1972 may have traveled between Europe and Nevada (both OWR) via the Appalachian region (EAR); however, they felt that other benthic organisms of that time do not support the Alberti et al. (Reference Alberti, Haas and Ormiston1972) route. In the same paper, Chatterton and Perry (Reference Chatterton and Perry1977) reported the presence of the trilobite Acanthopyge cf. consanguinea (Clarke, 1894) from the Lochkovian of Northwest Territories, Canada. The type specimens of A. consanguinea are from the Lochkovian-age New Scotland Formation of New York, which overlies the Coeymans Formation (Fig. 1).
In the same paper Chatterton and Perry (Reference Chatterton and Perry1977) described the conodont Ozarkodina remscheidensis remscheidensis (Ziegler, Reference Ziegler1960) from the early Lochkovian-age Road River Formation of Yukon Territory, and Delorme Formation of Northwest Territories. They noted what criteria they used to discriminate O. r. remscheidensis from its putative ancestor O. r. eosteinhornensis (Walliser, Reference Walliser1964). Denkler and Harris (Reference Denkler and Harris1988, p. B6) pointed out that attempts to separate the two subspecies of O. remscheidensis “…based on morphometric studies or subspecies distribution…have been unsuccessful.” They described O. remscheidensis from the Pridoli and lower Lochkovian of the northern Appalachians. Both of the above-mentioned papers reported the conodont Icriodus woschmidti Ziegler, Reference Ziegler1960, a biostratigraphic indicator for the base of the Lochkovian, in northwestern Canada and the Appalachians. Neither Chatterton and Perry (Reference Chatterton and Perry1977) nor Denkler and Harris (Reference Denkler and Harris1988) suggested an inter-realm route for the conodonts. The latter two authors did, to some extent, invoke the pelagic nature of I. woschmidti to explain its entry into what is now the southwestern end of the Appalachian Basin.
How large was the barrier?
Newer research that followed the establishment of the Canadian Shield-Transcontinental Arch barrier between the OWR and EAR in Laurussia, indicates that at least the Canadian Shield was not as prominent a physiographic feature as was once thought. Lower Devonian stromatoporoid-bearing deposits on Ellesmere Island (Norris, Reference Norris1996) and in southwestern Northwest Territories (Morrow and Cook, Reference Morrow and Cook1987) are located somewhat landward of Oliver's (Reference Oliver1977) shoreline (Fig. 11.1).
Even more impactful are the discoveries of Devonian-age xenoliths composed of normal-marine sedimentary rocks in kimberlites in the Canadian Shield of Northwest Territories (Cookenboo et al., Reference Cookenboo, Orchard and Daoud1998), the Ontario-Quebec boundary area (McCracken et al., Reference McCracken, Armstrong and Bolton2000), and southern Baffin Island, Nunavut (Zhang and Pell, Reference Zhang and Pell2014) (Fig. 11.2). All are located at least 100 km within the land area depicted by Oliver (Reference Oliver1977) for the Lochkovian (Fig. 11.1).
Using well logs, Thorpe (Reference Thorpe, McMillan, Embry and Glass1989) described the occurrence of the Pridoli–Lochkovian Kenogami Formation in the Hudson Bay Basin and western adjacent Moose River Basin; both are surrounded by Precambrian rocks of the Canadian Shield. He stated that the two basins were isolated from other basins to the west during the Devonian, and that they experienced “intermittent marine influxes and continuing dry conditions…” (Thorpe, Reference Thorpe, McMillan, Embry and Glass1989, p. 153).
Discussion
It is clear that during the Early Devonian, stromatoporoids were able to overcome the barrier presented by the Canadian Shield and the Transcontinental Arch. It was not likely that stromatoporoids were able to circumnavigate the barrier by going around either the now southwestern end of the Arch or the eastern side of the Shield. Surely planktonic and nektonic organisms could have achieved such a passage much more easily than the sessile benthic stromatoporoids. It is also unlikely that planktonic stromatoporoid larvae could float around Laurussia, or pass through/over the barrier, because a majority of marine organisms almost certainly had planktonic larvae, and there is little evidence on non-stromatoporoids existing concurrently on both sides of the barrier during the Early Devonian.
This leaves one plausible option—the stromatoporoids crossed the inter-realm barrier in several generations. Their larvae could travel by currents at least part of the way, as no doubt did the larvae of other organisms; however, only stromatoporoid adults could inhabit the very shallow conditions present in such a passage. This scenario coincides with the definition of a filter, which contained only a few habitats, restricting traverse to the few kinds of organisms capable of tolerating those conditions.
Conclusions
Multivariate statistical analysis, in this case average linkage cluster analysis followed by discriminant and canonical analyses, proved useful in differentiating among three closely related species of the stromatoporoid Habrostroma. Stock's (Reference Stock1988, Reference Stock1991, Reference Stock1997) use of the percentage of the skeleton occupied by localized, cyst-like microlaminae as the primary morphological characteristic in distinguishing among the three species of Habrostroma is strongly supported; however, it is not the only criterion that should be employed in differentiating among the three species. There are three species of Habrostroma in the Lochkovian of North America: H. centrotum (Girty), H. microporum (Girty), and H. consimile (Girty). All three species occur in the Manlius and Coeymans formations of New York.
Four specimens from the Keyser Formation of Virginia are H. centrotum, and a fifth is H. consimile. All six specimens from the Beck Pond Limestone of Maine are H. centrotum. All five specimens from the Stuart Bay Formation of Bathurst Island are H. centrotum. All eight specimens from the Goose Fiord Formation of Ellesmere Island are H. centrotum.
Habrostroma centrotum is the most common species present in New York, Maine, Bathurst Island, and Ellesmere Island, and is one of the two most common in Virginia. The coeval occurrence of Habrostroma centrotum, but neither H. microporum nor H. consimile, in both the Eastern Americas and Old World realms indicates a limited connection between the two biogeographic realms.
Evidence from xenoliths in kimberlite pipes in the Canadian Shield, and strata beneath Hudson Bay, indicates deposition of normal-marine sediments well onto the Shield. There was a shallow-water passage across the Canadian Shield part of what had been seen as a cross-continental barrier, which represented a filter, a seaway containing few habitats, restricting the kinds of organisms to those able to inhabit those depositional settings.
Acknowledgments
Field work and preparation of specimens from New York were supported by the U.S. National Science Foundation grant EAR-8404018, and the University of Alabama Research Grants Committee grant 1212, both to Stock; field work in Maine was supported by the University of Alabama Research Grants Committee grant 1399 to Stock; field work and preparation of specimens from Virginia were supported by a grant from Sigma Xi to A.E. Holmes; these contributions are gratefully acknowledged. R.D. White of the Yale Peabody Museum, E. Landing of the New York State Museum, J. Thompson, S. Twelker, and the late T.E. Bolton of the Geological Survey of Canada, and I. Birker of the Redpath Museum, McGill University facilitated loans of specimens. T.A. de Freitas is thanked for having the insight to collect stromatoporoids from Ellesmere Island, whereas others may not have. J.A. Adrain is thanked for telling the authors of the Chatterton and Perry (Reference Chatterton and Perry1977) article. Constructive comments by reviewers S. Kershaw and K. Liang helped to improve the manuscript.
Appendix. Collecting localities and specimen occurrences
New York
Locality numbers are those employed by Stock (Reference Stock1991) for the Manlius Formation and Stock (1997) for the Coeymans Formation.
Manlius Formation:
4. Cliff face 20–30 m north of western end of Indian Ladder Trail, John Boyd Thacher State Park; Albany County; 42°39'22''N, 74°1'0''W; Locality 59 of Rickard (Reference Rickard1962); Thacher Mbr.: USNM 408732.
5. Roadcut on south side of NY State Rt. 146 and ledges in adjacent woods, 0.7 km east of West Township, and 8.2 km east-northeast of Gallupville; Albany County; 42°42'0''N, 74°8'30''W; Locality 65 of Rickard (Reference Rickard1962); Thacher Mbr.: USNM 408733, 408734, 446789.
9. Abandoned quarries in pasture, 0.2 km west of road, 1.7 km south of Deck, and 4.2 km northeast of Crains Corners; Herkimer County; 42°56'6''N, 74°52'37''W; Locality 110 of Rickard (Reference Rickard1962); Thacher Mbr.: USNM 446790.
13. Roadcut and small abandoned quarry on east side, and outcrops in pasture on west side, of Columbia Center Road, 1.2 km west-southwest of Getman Corners, and 7.0 km east-northeast of Cedarville; Herkimer County; 42°56'50''N, 75°1'16''W; Locality 121A of Rickard (Reference Rickard1962); Clark Reservation Mbr.: USNM 408729; Jamesville Mbr.: USNM 408725–408728.
14. Roadcut and abandoned quarry on north side of road, 70 m east of T-intersection, and 1.1 km southwest of Cedarville; Herkimer County; 42°55'35''N, 75°7'31''W; Clark Reservation Mbr.: USNM 408724.
15. Roadcut on west side of road, 1.3 km northwest of Cedarville; Herkimer County; 42°56'18''N, 75°7'32''W; Elmwood(?) Mbr.: USNM 408731.
16. Abandoned quarry 0.2 km south of road, and 0.6 km west of Cranes Corners; Herkimer County; 42°59'13''N, 75°7'51''W; Locality 124 of Rickard (Reference Rickard1962); Clark Reservation Mbr.: USNM 408730.
17. Quarries north and south of County Rt 9, just west and south of junction with County Rt 16, 1.15 km southeast of McConnell Corners, and 3.8 km west-southwest of Paris; Oneida County; 42°59'25''N, 75°21'45''W; Locality 128 of Rickard (Reference Rickard1962); Olney Mbr.: USNM 446786.
18. Outcrops along bank of brook 0.1 km west of Cook Road, and 2.15 km east of Stockbridge; Oneida County; 42°59'32''N, 75°34'21''W; Locality 135 of Rickard (Reference Rickard1962); Elmwood Mbr.: USNM 446774.
23. Abandoned quarry 0.6 km northwest of junction of NY State Rt 173 with North Street (Duguid Road) in Manlius, and just northeast of abandoned railroad right-of-way; Onondaga County; 43°0'25''N, 75°58'18''W; Locality 148B of Rickard (Reference Rickard1962); Jamesville Mbr.: USNM 446791.
25. Roadcuts on median and east side of Interstate Rt 81, 0.3 km south of NY State Rt 173 underpass; Onondaga County; 42°59'58''N, 76°7'7''W; Olney Mbr.: USNM 446787.
26. Abandoned quarry, now parking lot, in Skaneateles Falls on east side of Jordan Road, 0.15 km north of intersection with Stump Road, and 6.0 km north-northwest of Skaneateles; Onondaga County; 42°59'45''N, 76°27'26''W; Locality 159 of Rickard (Reference Rickard1962); Jamesville Mbr.: USNM 408735.
Coeymans Formation:
1. Outcrops in pasture on west side of Columbia Center Road, 1.2 km west-southwest of Getman Corners, and 7.0 km east-northeast of Cedarville; Herkimer County; 42°56'52''N, 75°1'13''W; part of Locality 121A of Rickard (Reference Rickard1962); Dayville Mbr.: USNM 432831.
5. Upper end of roadcut 0.35 km east of Munns, and 50 m east of Mosquito Point; Oneida County; 42°58'48''N, 75°34'21''W; part of Locality 136 of Rickard (Reference Rickard1962); Deansboro Mbr.: USNM 432830.
Virginia
Keyser Formation: Roadcut along northeast side of VA State Rt 607, Mustoe; Highland County; 38°19'18''N, 79°38'18''W; Jersey Shore Mbr.: USNM 364710, 364711; LaVale Mbr.: USNM 364708, 364709, 364712.
Maine
Locality numbers are those employed by Boucot et al. (Reference Boucot, Harper and Rhea1959; Reference Boucot, Harper and Rhea1966); all are outcrops in the forest, in Somerset County.
Beck Pond Limestone:
2. 204 m south-southwest of the south end of Beck Pond; 45°22'5''N, 70°20'22”W; Mbr. 4: USNM 526378, 526379.
10. 122 m south of the south end of Beck Pond; 45°22'5''N, 70°20'17''W; Mbr. 5: USNM 526380, 526381.
22. 329 m west of the south end of Beck Pond; 45°22'8''N, 70°20'31''W; Mbr. 2: USNM 526376, 526377.
Bathurst Island
Locality designations are those employed by Stearn (Reference Stearn1990).
Stuart Bay Formation:
AS. “Arthur's Seat;” 75°51'N, 98°31'W; GSC 95775.
a. Polar Bear Pass; 75°45'N, 98°20'W; Block 23: RM 20.6802
MR. Moses Robinson River; 76°5'N, 97°58'W; Block 5: GSC 95776; Block 14: GSC 95774; Block 16: RM 20.6801.
Ellesmere Island
Locality Numbers are those employed by Norris (Reference Norris1996).
Goose Fiord Formation:
63. Mackinson Inlet area, southern Ellesmere Island; 77°43'N, 82°6'W; 194 m above base of section: GSC 10762, 10763.
67. Vendon River area, southern Ellesmere Island; 77°47'N, 82°24'30''W; 278–336 m above base of section: GSC 10764; 534–540 m above base of section: GSC 10765, 10766; 540 m above base of section: GSC 10767; 690 m above base of section: GSC 10768, 10769.


















