



Introduction
The fossil record of annelid worms is generally sparse because many annelids do not possess easily preservable tissues. The fossil record of annelids dates back to the early Cambrian, with complete worms being known from several Cambrian Konservat-Lagerstätten (Parry et al., Reference Parry, Tanner and Vinther2014). Molecular clock evidence suggests that crown-group annelids arose during the late Cambrian–Ordovician interval (Parry et al., Reference Parry, Tanner and Vinther2014); thus, the Ordovician annelid record is crucial to understanding the evolution of the group. The scolecodont record suggests substantial diversification of jawed polychaetes during the Ordovician Period (Hints and Eriksson, Reference Hints and Eriksson2007). Machaeridians (now known to be annelids; Vinther et al., Reference Vinther, Van Roy and Briggs2008) also appear to have diversified during the same interval (Hints et al., Reference Hints, Eriksson, Högstrom, Kraft, Lehnert, Webby, Paris, Droser and Percival2004). However, there are few annelid body fossils known from Ordovician rocks, making it difficult to assess the evolution of the group as a whole during this time.
A number of fossils identified as annelids or other types of worm were described from Ordovician strata in the nineteenth and earlier part of the twentieth centuries (e.g., Ulrich, Reference Ulrich1878; Ruedemann, Reference Ruedemann1901, Reference Ruedemann1934; Moberg and Segerberg, Reference Moberg and Segerberg1906; Hadding, Reference Hadding1913). The majority of these fossils have never been restudied or adequately illustrated, and it is unclear whether they are indeed worms and, if they are worms, to which phylum they belong. Some of these fossils, such as those that are the subject of this paper, have been included in paleontological databases such as the Paleobiology Database (PBDB, source: fossilworks.org; search conducted 28 April 2019) and compilations (Sepkoski, Reference Sepkoski2002), at which point rare taxa in particular can influence perceived global patterns, even when not explicitly cited or discussed. For members of rare fossil groups, erroneous data points can become significant.
Rudolf Ruedemann (1864–1956) was an extremely prolific paleontologist, with more than 140 publications during his lifetime (Rodgers, Reference Rodgers1974). Much of his work has stood the test of time; however, in some cases he described structures in fossils that, upon subsequent examination, are not present (e.g., Simonetta, Reference Simonetta1961; Maletz, Reference Maletz2015), and it appears that many of his specimens are of inorganic origin (Tollerton, Reference Tollerton2006). One of Ruedemann's putative worms is Haileyia adhaerens Ruedemann, Reference Ruedemann1934, from the Ordovician Normanskill Shale of Idaho, USA. Herein we re-describe and illustrate photographically the type (and only known) material of Haileyia adhaerens and assess its interpretation.
Materials and methods
The type material of Haileyia adhaerens Ruedemann, Reference Ruedemann1934 consists of two specimens: the holotype (USNM PAL 90853; Fig. 1) and the paratype (USNM PAL 706443; Fig. 2), both of which lack counterparts. Both specimens are from the Normanskill Shale (upper Middle Ordovician), near the head of Fall Creek, Idaho, USA. We could find no references to additional material of Haileyia adhaerens in the literature, so the re-description is based solely on the type series. The specimens were examined both dry and wet under both low- and high-angle light. Photographs were obtained with a Nikon D80 camera and a Maozua 5 MP USB microscope.
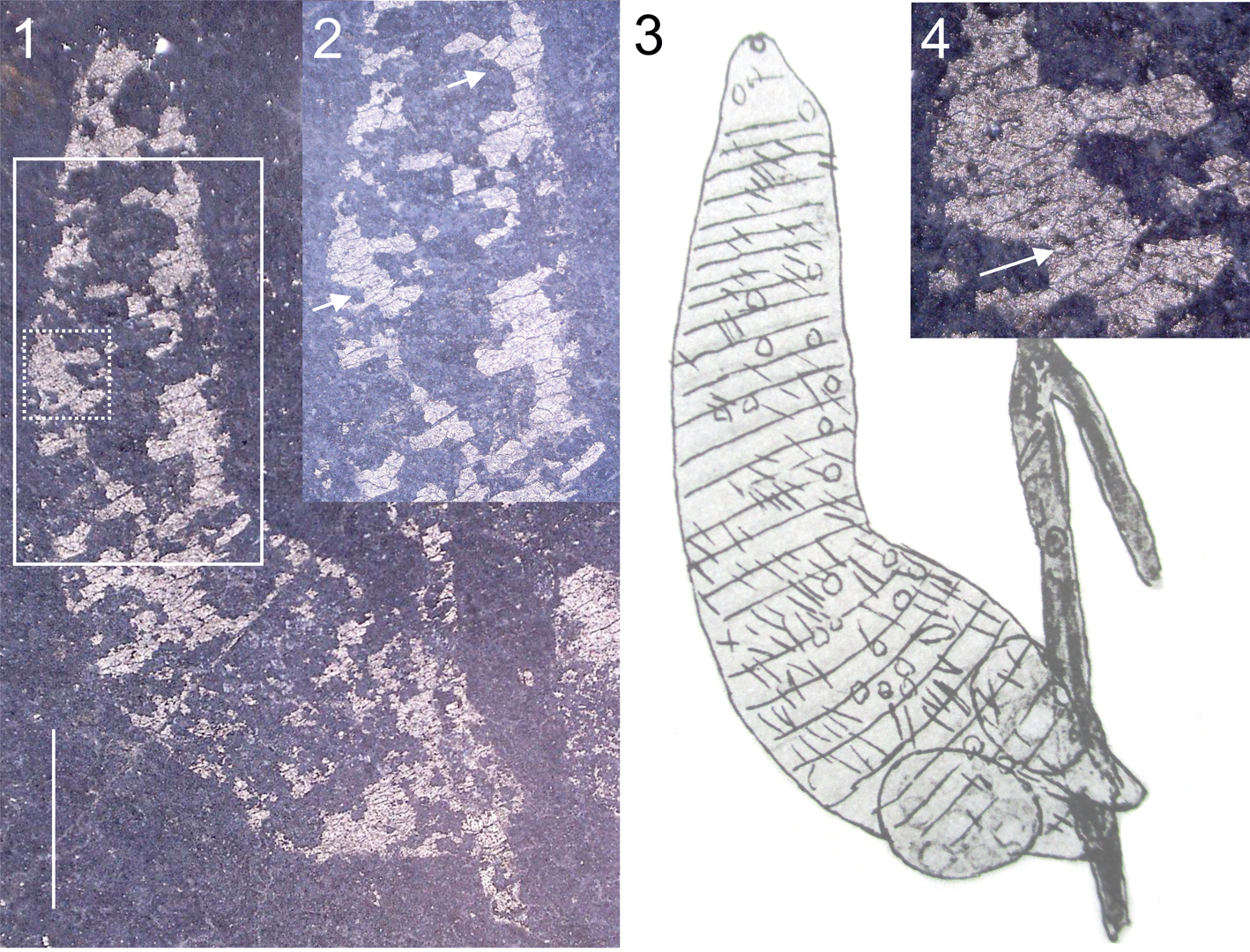
Figure 1. Haileyia adhaerens Ruedemann, Reference Ruedemann1934. (1, 2, 4) Holotype, USNM PAL 90853, entire specimen, photographed under water; (2) same, enlargement of the area indicated by the dashed white rectangle in (1), photographed under water, scattered bumps on film indicated by white arrow; (4) same, enlargement of the area indicated by the solid white rectangle in (1), photographed under water, showing the transverse structures on the specimen; the white arrows in (2, 4) indicate irregular cracking; (3) reproduction of the original illustration of the holotype (Ruedemann, Reference Ruedemann1934, pl. 21, fig. 11). The original illustration has been rotated to match the orientation of the photograph. Original image is copyright Geological Society of America and used with permission. Scale bar represents 2 mm.

Figure 2. Haileyia adhaerens Ruedemann, Reference Ruedemann1934, paratype, USNM PAL 706443. (1) Photographed dry; note that the contrast has not been digitally enhanced, so that the image accurately reflects the actual appearance of the fossil; (2) reproduction of the original illustration of the paratype (Ruedemann, Reference Ruedemann1934, pl. 21, fig. 10). The original illustration has been rotated to match the orientation of the photograph. Original image is copyright Geological Society of America and used with permission. Scale bar represents 2 mm.
Repository and institutional abbreviation
The specimens are housed in the National Museum of Natural History, Washington DC, USA (NMNH).
Description of material
The Haileyia material is preserved as reflective films (presumed to be carbonaceous, perhaps with minor iron mineralization) on shale bedding planes, broadly similar in appearance to graptolites on the same slabs.
The holotype specimen tapers in both directions (abruptly in one direction) and is slightly curved (Fig. 1.1). It is ~10 mm long and ~3 mm wide at the widest point, tapering to a blunt point at the thin end; the wider end has a blunt termination (Fig. 1.1). Transverse structures extend across part of the width of the specimen, but are not entirely parallel and show irregularity typical of taphonomic cracking of a brittle sheet (Fig. 1.2). The surface of the specimen is irregular, and bears minute scattered bumps (Fig. 1.4). No other structures are visible within or around the margin of the specimen. The wider end of the specimen underlies a graptolite stipe.
The paratype (Fig. 2) is ~13 mm long and irregularly cylindrical, with a width of ~2 mm over most of its length. No structures are visible within or around the specimen, and the margins are diffuse. One end of the specimen is overlain by a graptolite stipe and the other is in contact with a dicellograptid graptolite.
Discussion
In the original description, Haileyia is stated to possess segmentation, parapodia, setae, and papillae (Ruedemann, Reference Ruedemann1934). Transverse structures visible on the holotype were interpreted by Ruedemann (Reference Ruedemann1934) as sutures separating segments; however, the transverse structures are not parallel (Fig. 1.2), as would be expected for the boundaries of annuli/segments, the dark lines are visible rock between broken plates of the organic film, and there are numerous oblique and perpendicular structures that indicate abiological cracking rather than original structure. Accordingly, these features are reinterpreted as cracks in the thin film that constitutes the specimen, and not as biological features. The putative parapodia illustrated by Ruedemann (Reference Ruedemann1934) on the wider end of the holotype (Fig. 1.3) are not visible. Ruedemann (Reference Ruedemann1934, p. 86) stated that “minute papillae and delicate setae are irregularly distributed over the body.” The ‘setae’ could not be detected under the microscope, but the ‘papillae’ are probably a reference to minute, scattered bumps on the film that appear to be impressions of pyrite framboids (Fig. 1.4). The surface of the film that forms the specimen is markedly irregular, but there is no suggestion that this indicates the presence of original structures.
Ruedemann (Reference Ruedemann1934) stated that the putative worms were attached to graptolites and hypothesized that this was their mode of life. Both specimens are overlain by planktonic graptolites (Figs. 1.1, 2.1), and there is no sign of any parts of the putative worms overlying the graptolites, as would be expected if the worm had been clasping the graptolite. We interpret these associations as having arisen by chance because graptolites are very abundant on the bedding surfaces. For example, on the paratype slab, which measures 55 mm by 50 mm, there are at least 20 graptolites and at least 10 pieces of reflective film similar to those forming the type material of Haileyia. It is thus not surprising that some of these films are in contact with graptolites.
Haileyia adhaerens was identified by Ruedemann (Reference Ruedemann1934) as a worm, but not firmly assigned to any group; nor was it assigned to a phylum, class, order, or family by Howell (Reference Howell and Moore1962, p. W170), although it is listed as a polychaete annelid by the PBDB (source: fossilworks.org; search conducted 28 April 2019). The PBDB lists 22 Ordovician polychaete genera (source: fossilworks.org; search conducted 28 April 2019, search terms Polychaeta and Ordovician), including Haileyia. Thus, this occurrence forms part of (and biases) our knowledge of large-scale patterns in the fossil record. Although this is only one record, it is not known how many other records are also erroneous, and so the re-interpretation of Haileyia serves as a reminder that data should not be used uncritically.
Conclusions
The type material of Haileyia adhaerens does not show any features that allow identification as a worm. Although the specimens probably represent organic remains, it is not possible to tell what type of organism may have produced them; such organic debris is extremely common in Ordovician black mudstone sequences, and is typically rejected as being undeterminable. As a result, both the generic and specific names should be abandoned, and the record should be rejected as evidence for any taxonomic group.
Acknowledgments
We are grateful to M. Florence, National Museum of Natural History, Washington DC, USA, for organizing the loan of the type material of Haileyia, and C. Howells, Amgueddfa Cymru, Cardiff, UK, for facilitating the loan. O. Vinn and an anonymous reviewer are thanked for their supportive reviews; we are also grateful to the handling editor, A. Liu. No funding was received for this research.


