



Introduction
During the Early and Middle Devonian, a particular marine fauna mostly recognized based on brachiopods and trilobites developed in southwestern Gondwana, which justified the recognition of the Malvinokaffric Realm. This major paleobiogeographical region had an almost circumpolar distribution, being recorded in southern South America, Antarctica, and South Africa (Boucot and Racheboeuf, Reference Boucot and Racheboeuf1993). Trilobites are fundamental to understand biogeographic patterns and paradigmatic evolutionary events in this realm (Eldredge and Ormiston, Reference Eldredge and Ormiston1979; Abe and Lieberman, Reference Abe and Lieberman2012). Most of the studies on trilobites, however, have been conducted on calmoniids, an entirely endemic family emblematic of the Malvinokaffric Realm that radiated spectacularly during the Early Devonian (Abe and Lieberman, Reference Abe and Lieberman2012), whereas phacopid trilobites, common and well-known elements of Devonian marine communities of Laurussian basins, have received less attention.
Devonian marine strata in Argentina, particularly those from the Precordillera Basin, are well developed and yield a diverse Malvinokaffric trilobite fauna, particularly in the Precordillera Basin (Baldis, Reference Baldis1975). In this basin, only few phacopids have been reported, namely“Phacops” chavelai Baldis and Longobucco, Reference Baldis and Longobucco1977, from the Middle Devonian Chigua Formation, and Reedops cf. bronni (Barrande, Reference Barrande1846) by Holloway and Rustán (Reference Holloway and Rustán2012) together with a couple of species of Paciphacops Maksimova, Reference Maksimova1972 (Rustán et al., Reference Rustán, Vaccari and Astini2011a, Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b), from the Lower Devonian Talacasto Formation.
Paciphacops species received renewed interest as they were interpreted as representing new Devonian species forming part of a Malvinokaffric Silurian-Devonian lineage that showed a particular infaunal moulting habit (Rustán et al., Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b). This behavior, together with a trend toward increasing size and spinosity, were interpreted as anti-predator adaptations, that evolved during an increase in global marine durophagous predation recorded in the Devonian (Rustán et al., Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b) known as the Paleozoic Marine revolution (Signor and Brett, Reference Signor and Brett1984). Moreover, those features evinced compellingly that trilobites were actively involved as preys in this major global bioevent (Rustán et al., Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b). However, the fact that infaunal moulting was limited to a few of phylogenetically related species prevented further inferences about the evolution of such behavior and the actual scope of the Marine Paleozoic Revolution.
Here we report on the taxonomic reappraisal of some of these Argentinian specimens, previously reported as Paciphacops n. sp. (Rustán et al., Reference Rustán, Vaccari and Astini2011a) and Paciphacops n. sp. B (Rustán et al., Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b), based on new and better materials from the Lochkovian of the Talacasto Formation. The fossils support a reassignment to a new species of the genus Echidnops Sandford, Reference Sandford2002 previously restricted to eastern Gondwana (Australia), refusing the previously supposed Paciphacops lineage (Rustán et al., Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b). In addition, new morphological information on the particular lens arrangement of this new Echidnops species, contributes to a discussion on the visual system of phacopids. In turn, paleobiogeographic implications of this first non-Australian Echidnops record in the Malvinokaffric context are discussed. The recognition of infaunal moulting in other taxa than Paciphacops, provides new insights into the extent of ethological predator-deterrent strategies among Devonian trilobites and some linked evolutionary aspects.
Geological setting
The Lower Devonian Talacasto Formation (Padula et al., Reference Padula, Rolleri, Mingramm, Criado Roqué, Flores and Baldis1967) is mainly exposed in the Central Argentine Precordillera of San Juan Province (Fig. 1), although isolated outcrops are also known in the northernmost part of the Precordillera, in La Rioja Province (Rustán et al., Reference Rustán, Vaccari and Astini2011a). This formation is composed of a marine succession of intensely bioturbated greenish-gray mudstones with intercalated sandstones. It begins with dark, fine-grained, muddy levels, passing upwards into sandy levels (Fig. 1). In its type locality at Quebrada de Talacasto, Sierra de Talacasto, in the San Juan Province (Fig. 1), the formation reaches around 300 m in thickness, but a maximum of more than 1000 m is recorded to the north at the Loma de los Piojos section, near Jáchal city.
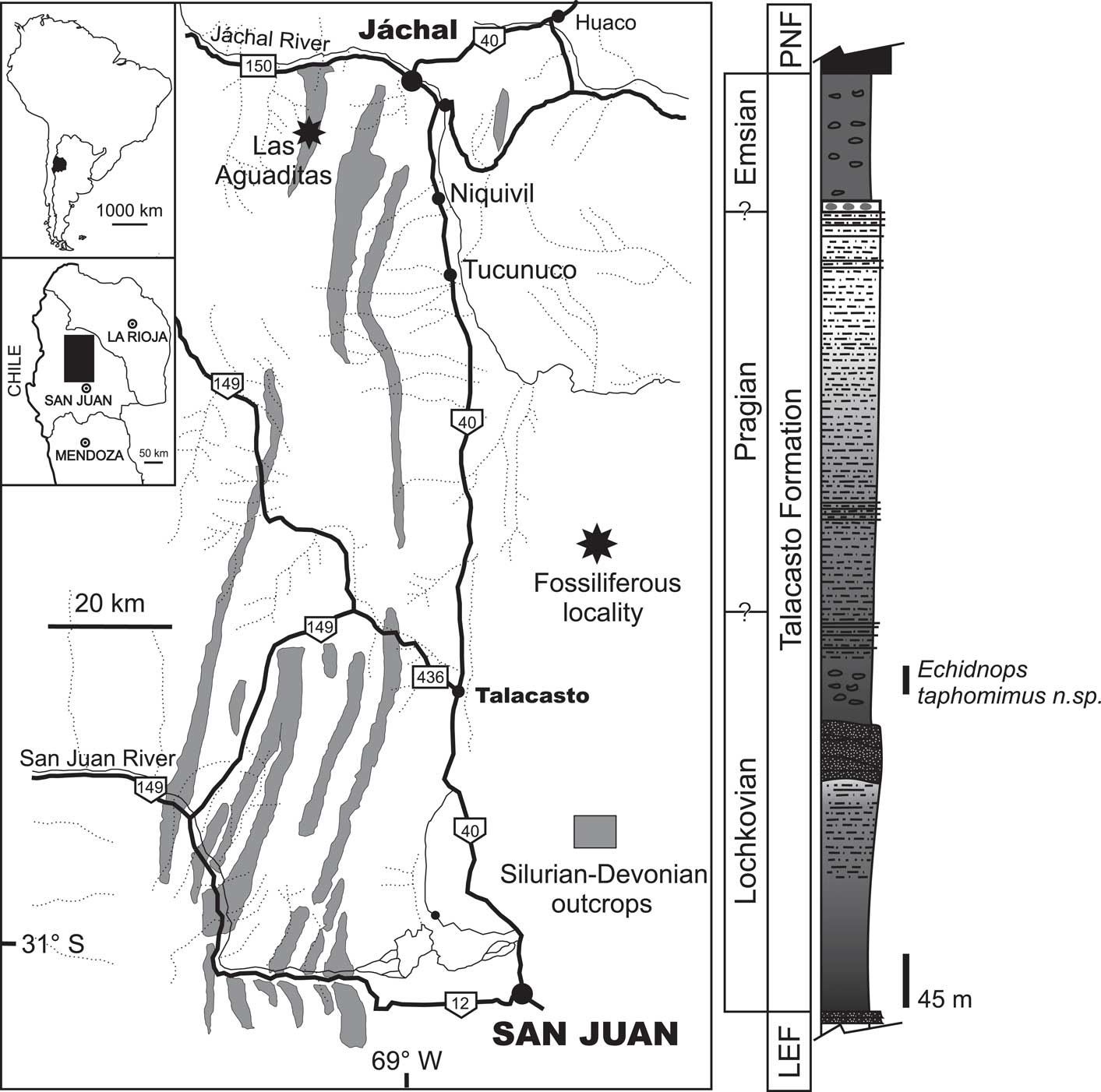
Figure 1 Location map and schematic stratigraphic column of the fossil locality recording Echidnops taphomimus n. sp. in the Lochkovian of the Talacasto Formation, Quebrada de Las Aguaditas, Sierra de Los Blanquitos, San Juan Province, Argentina. LEF=Los Espejos Formation (Upper Silurian); PNF=Punta Negra Formation (Middle Devonian).
The Talacasto Formation represents a muddy shelf depositional system developed during a highstand. This formation overlies the mainly Silurian shelf succession of the Los Espejos Formation (Astini, Reference Astini1991) and is overlied by the turbiditic system of the upper Lower-lower Upper? Devonian Punta Negra Formation (Bracaccini, Reference Bracaccini1949; Bustos, Reference Bustos1996; Bustos and Astini, Reference Bustos and Astini1997). The Talacasto Formation is the source of most of the Devonian macrofossils described from Argentina, sometimes exceptionally preserved. The main fossil groups include brachiopods, trilobites, bivalves, gastropods, cephalopods, echinoderms, corals, tentaculitids, ostracods, hyoliths, conulariids, abundant ichnofossils, and scarce plant remains.
According to brachiopod and palynological information (Benedetto et al., Reference Benedetto, Racheboeuf, Herrera, Brussa and Toro1992; Herrera, Reference Herrera1993; Racheboeuf and Herrera, Reference Racheboeuf and Herrera1994; Herrera and Bustos, Reference Herrera and Bustos2001; García-Muro and Rubinstein, Reference García-Muro and Rubinstein2015), the Talacasto Formation spans from the Lochkovian to the Emsian. The base of the Devonian is missing at most places due to the presence of a discontinuity between the Los Espejos and Talacasto formations. However, the Silurian-Devonian boundary can be recognized in the uppermost interval of the Los Espejos Formation in a few localities toward the north of the Talacasto area, at Cerro del Fuerte, La Chilca (Benedetto et al., Reference Benedetto, Racheboeuf, Herrera, Brussa and Toro1992; Carrera et al., Reference Carrera, Montoya, Rustán and Halpern2013), and Jáchal river sections (García-Muro and Rubinstein, Reference García-Muro and Rubinstein2015).
Localities
Most of the fossils studied herein come from the Quebrada de Las Aguaditas, Sierra de Los Blanquitos, approximately 12 km southwest of Jáchal, San Juan Province (Fig. 1). Fossiliferous horizons correspond to the same stratigraphic interval of the muddy lower part of the Talacasto Formation from which Edgecombe et al. (Reference Edgecombe, Vaccari and Waisfeld1994), described calmoniid trilobites such as Talacastops zarelae. Additional specimens come from equivalent layers of an isolated outcrop in the Sierra de Las Minitas, near Jagüé town, northernmost Precordillera, La Rioja Province (Rustán et al., Reference Rustán, Vaccari and Astini2011a). All specimens are preserved in dark grey to black nodules, included in a greenish-dark grey to black pelitic bioturbated interval, reported as Facies Association A by Astini (Reference Astini1991). Co-occurring trilobites include dalmanitids, odontopleurids, proetids, and calmoniids (Edgecombe et al., Reference Edgecombe, Vaccari and Waisfeld1994; Rustán et al., Reference Rustán, Vaccari and Astini2011a). This characteristic trilobite association is dominated by Echidnops taphomimus n. sp., which is mainly represented as articulated exuviae, whereas all other trilobites are preserved as disarticulated exoskeletal parts. The accompanying fauna, either in nodules or in the surrounding matrix, includes corals, crinoids, hyoliths, brachiopods, bivalves, cephalopods, conulariids, gastropods, and vertebrates (a small fragment of bone).
Age
Brachiopod data indicated an early Lochkovian age for the fossiliferous interval at the Las Aguaditas locality (Edgecombe et al. Reference Edgecombe, Vaccari and Waisfeld1994; Herrera, Reference Herrera1995 and references therein). Equivalent strata from the Sierra de Las Minitas (Rustán et al., Reference Rustán, Vaccari and Astini2011a), have been dated as late Lochkovian based on palynological content (Rubinstein et al., Reference Rubinstein, Monge, Rustán and Astini2010). The presence of the phacopid Reedops cf. bronni in proximity to the fossiliferous locality bearing E. taphomimus at Sierra de Las Minitas (Holloway and Rustán, Reference Holloway and Rustán2012), suggest that the succession would range into the Pragian.
Systematic paleontology
Illustrated specimens are housed in Argentina, in the paleontological collections of the CIPAL (Centro de Investigaciones Paleobiológicas), at the CICTERRA (Centro de Investigaciones en Ciencias de la Tierra: CONICET-Universidad Nacional de Córdoba), Córdoba (CEGH-UNC) Additional material is housed in the paleontological repository of the Museo de Ciencias Naturales at the Universidad Nacional de La Rioja, La Rioja (PULR).
Order Phacopida Salter, Reference Salter1864
Suborder Phacopina Struve in Richter et al., Reference Richter, Richter and Struve1959
Superfamily Phacopoidea Hawle and Corda, Reference Hawle and Corda1847
Family Phacopidae Hawle and Corda, Reference Hawle and Corda1847
Subfamily Phacopinae Hawle and Corda, Reference Hawle and Corda1847
Genus Echidnops Sandford, Reference Sandford2002
Type species
Echidnops wrighti Sanford, 2002 from the Lochkovian of the Humevale Formation, central Victoria, Australia, by original designation.
Echidnops taphomimus new species
Figures 2.1–2.13, 3.1–3.10, 4.1–4.13, 5.1–5.9
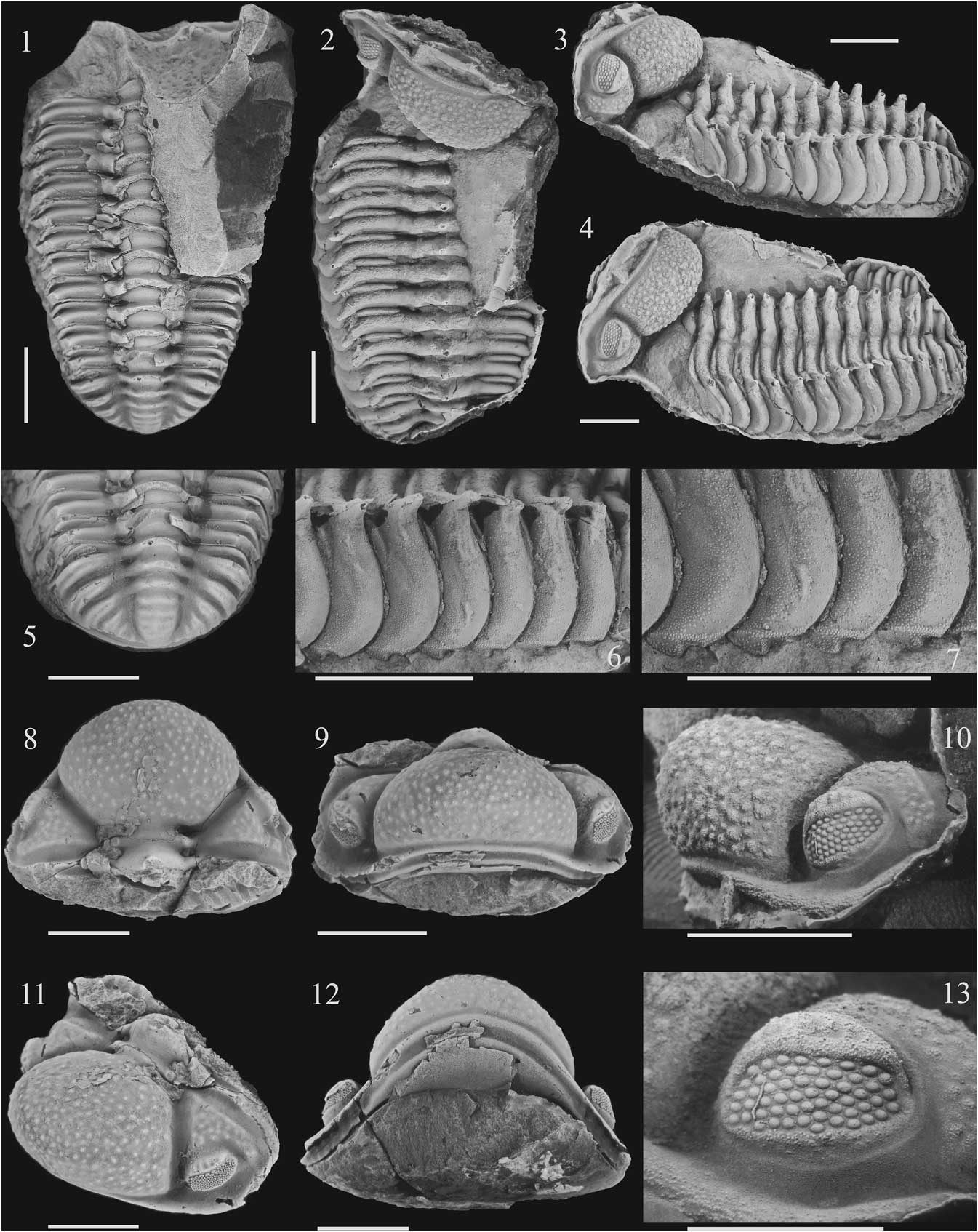
Figure 2 Echidnops taphomimus n. sp., holotype CEGH-UNC 24082, Lochkovian, Talacasto Formation, Quebrada de Las Aguaditas, Sierra de Los Blanquitos, San Juan Province, Argentina: (1) internal mold, dorsal view; note the infaunal molt pattern; (2) latex cast of external mold, (3) lateral view; (4) latero-dorsal view; (5) pygidium, dorsal view; (6) detail of thoracic fulcral spines and pleural tips in lateral view; (7) detail of ornamentation and uncinate ventral processes of pleural tips; (8) internal mold of cephalon, dorsal view; (9) internal mold of cephalon, frontal view; (10) latex cast of cephalon external mold, lateral view; (11) internal mold of cephalon, latero-dorsal view; (12) internal mold of cephalon, ventral view; (13) external mold of cephalon, detail of the eye, nearly antero-lateral view. Scale bars represent 1 cm in all photographs except (13), where represents 0.5 cm.

Figure 3 Cephalic morphology of Echidnops taphomimus n. sp., Lochkovian, Talacasto Formation, Quebrada de Las Aguaditas, the Sierra de Los Blanquitos, San Juan Province, Argentina: (1–3) cephalon, internal mold (paratype CEGH-UNC 24088), dorsal, anterodorsal and ventral views; (4, 5) cephalon, internal mold (paratype CEGH-UNC 12744), dorsal and postero-dorsal views; note the three dimensional arrangement, diagnostic of the infaunal molt pattern, involving the hypostome obliquely detached and the cephalon rotated up and backward; (6) the same specimen, detail of the (incomplete) hypostome; (7) cephalon, internal mold, dorsal view (paratype CEGH-UNC 26130); (8) latex cast of cephalon external mold, dorsal view, note S2 and S3 (paratype CEGH-UNC 26152); (9) latex cast of cephalon external mold, antero-dorsal view (paratype CEGH-UNC 12742); (10) cephalon, internal mold, ventral view (paratype CEGH-UNC 12740); (11) the same as (8), detail of tuberculation, note thick granulation of tubercles; (12) the same as (9), detail of tuberculation, note granulation and central pits in larger tubercles; (13) latex cast of cephalon external mold, dorso-lateral view, note the genal spine, (paratype CEGH-UNC 12741). Scale bars represent 5 mm in all photographs except in (12) and (13), where represents 1 mm.

Figure 4 Examples of typical infaunal molt patterns, and thoracic morphology of Echidnops taphomimus n. sp., Lochkovian, Talacasto Formation, Quebrada de Las Aguaditas, Sierra de Los Blanquitos, San Juan Province, Argentina: (1) latex cast of a nearly complete specimen, dorsal view, note the nearly vertical position of the cephalon over the thorax (paratype CEGH-UNC 24081); (2) the same in postero-dorsal view; (3) the same, detail of the nearly 90 degrees angle in the contact between the occipital ring lying over the first thoracic segment, (4) a nearly complete specimen, internal mold (paratype CEGH-UNC 12740); (5) latex cast of a nearly complete thoracopigydium, dorsal view, note the nearly vertical position of the cephalon over the thorax (paratype CEGH-UNC 26151); (6) a nearly complete specimen, internal mold, postero-dorsal view (paratype CEGH-UNC 24084); (7) a nearly complete specimen, internal mold, postero-lateral dorsal view (paratype CEGH-UNC 24083); (8) pygidium, internal mold, dorsal view (CEGH-UNC 26132); (9) latex cast of pygidium external mold, dorsal view (CEGH-UNC 12745); (10) the same as (4), detail of pygidium, dorsal view. Scale bars represent 1cm.
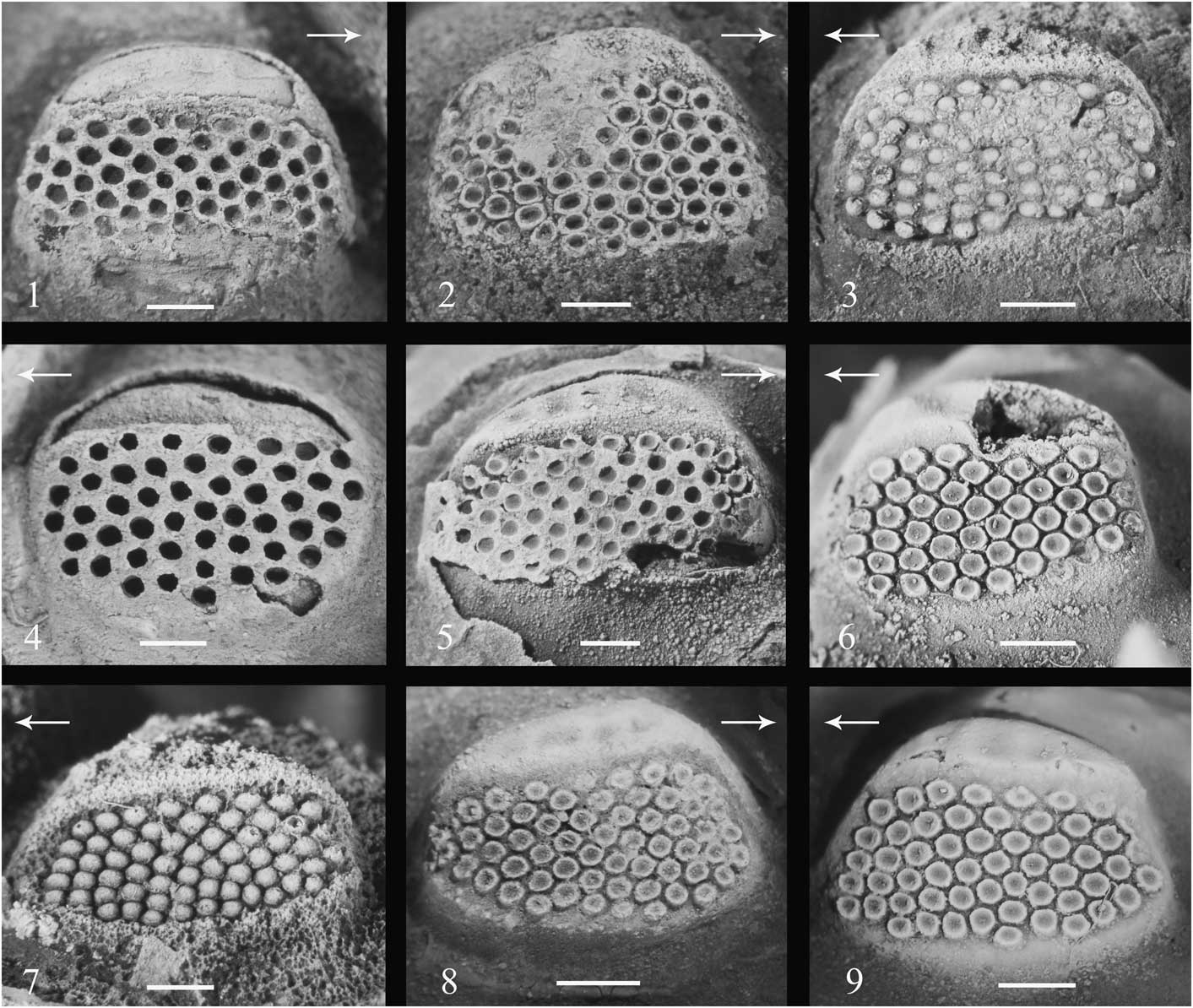
Figure 5 Eye morphology of Echidnops taphomimus n. sp., Lochkovian, Talacasto Formation, Quebrada de Las Aguaditas, Sierra de Los Blanquitos, San Juan Province, Argentina. Note the irregular arrangement (files are obscurely defined) and similar size of lenses: (1) right eye, internal mold, lateral view (CEGH-UNC 26131); (2) right eye, internal mold, lateral view (CEGH-UNC 12741); (3) left eye, with lenses preserved, lateral view (CEGH-UNC 12741); (4) left eye, internal mold, lateral view (CEGH-UNC 26130); (5) right eye, internal mold, lateral view (CEGH-UNC 26129); (6) left eye, internal mold, lateral view (CEGH-UNC 26133); (7) left eye, latex cast mold, lateral view (CEGH-UNC 12742); (8) right eye, internal mold, lateral view (CEGH-UNC 12740); (9) left eye, internal mold, lateral view (holotype CEGH-UNC 24082). Arrows point anteriorly. Scale bars represent 1 mm.
2011a Paciphacops (Paciphacops) n. sp. Reference Rustán, Vaccari and AstiniRustán et al., p. 229, pl. 2, figs. 1–9.
2011b Paciphacops (Paciphacops) n. sp. B Reference Rustán, Balseiro, Waisfeld, Foglia and VaccariRustán et al., p. 496, pl. 2, figs. A–D, DR 4.
Holotype
CEGH-UNC 24082, a nearly complete infaunal exuvia from the lower interval of the Talacasto Formation (Lochkovian) at Quebrada de Las Aguaditas, Sierra de Los Blaquitos , San Juan Province, Argentina (Fig. 2).
Paratypes
CEGH-UNC 12740-42, 12744, 24081, 24083-84, 24088, 26130, 26151-52.
Diagnosis
Large Echidnops (more than 9 cm of maximum total length) with upturned cephalic anterior region; small eyes with 50 up to 60 lenses, distributed in an irregular pattern of 10–14 poorly defined vertical files, with a maximum of six to seven lenses; arcuate and crenulated palpebral lobe; very stout and wide-based genal, intergenal, and occipital cephalic spines, and similar axial and fulcral thoracic ones; lateral lobe of the thoracic axial ring just weakly separated by a couple of shallow fossulae (not by a clear furrow); anterior pleural band very narrow (ridge-like), nearly straight and transversely oriented; posterior pleural band inflated and very wide (exsag.).
Ocurrence
Lower part of the Talacasto Formation (probably late Lochkovian) at Quebrada de Las Aguaditas, Sierra de Los Blaquitos (San Juan Province) and Sierra de Las Minitas, La Rioja Province, Argentine Precordillera.
Description
Rustán et al. (Reference Rustán, Vaccari and Astini2011a) gave an description of E. taphomimus n. sp. based on fossils from Las Minitas, here we focus on new information provided by materials from the Quebrada de Las Aguaditas.
Known maximum length of toracopygidium (CEGH-UNC 7057) ∽62 mm, and cephalon (CEGH-UNC 26135) ∽25 mm. Cephalon usually representing ∽30% of total length. Glabella with frontal lobe markedly inflated and directed upward, outstanding forward and projecting beyond the preglabellar furrow in lateral view (Figs. 2.8–2.12, 3.1–3.5, 3.7, 3.10). Cephalic tubercles bearing granules and usually a wide funnel-like depressed central area (Fig. 3.11, 3.12) that corresponds to a central pit in internal molds (Fig. 3.1). Anterior cephalic region upturned. Axial furrows highly divergent forward (70°–80°). Palpebral lobe narrow (tr.), sloping anterolaterally, gently arcuate to semilunar in outline, with large fossulae disposed nearly in a row laterally, giving it a crenulated appearance. Medium to small eye with length (exsag.) (∽30%–40% of sagittal cephalic length); visual surface with an irregular lenses arrangement, 10–14 poorly defined dorso-ventral files of lenses and a maximum of six to seven lenses (total lens number usually between 50 and 60, see Table 1). Cephalic posterior border as a strong ridge, deflected postero-laterally abaxial to fulcrum, which bears a stout, dorsolaterally-directed intergenal spine transversely elliptical in cross section at its base. Genal spine small, stout, slightly flattened in cross section at its base and posterolaterally directed. Vincular furrow well impressed, with anterior and posterior edges clearly defined, evenly arched upward in frontal view (Fig. 2.9), narrowest and shallowest immediately below the anterior ends of the axial furrows (Figs. 2.12, 3.3, 3.10), becoming distinctly crenulated posteriorly.
Table 1 Some measurements of the head and eyes of Echidnops taphomimus n. sp.

Hypostome large, widest (tr.) across anterior wings (Fig. 3.6). Hypostomal suture smooth, even, and gently bowed forward. Anterior wings very large, laterally extended and disposed in a very high angle that is almost transversely oriented respect of the hypostomal median body; with a strong distal inflation and separated by large and very deep lateral notches, more or less exsagittally oriented. Median body, slightly longer than wide (although its posterior part is broken), considerably inflated adventrally. Anterior lobe barely inflated, defined by a very faint furrow. Maculae not evident. Lateral border furrow moderately wide and deep. Small and shallow fossulae anterolaterally on the lateral border furrow, at junction between the anterior wings and the anterior lobe. Lateral border narrow and moderately high.
Distal portion of thoracic pleurae with a nearly blunt ventral uncinate-like process, posteriorly directed and separated from the lateral side of the pleural tip by a, transverse, nearly straight granulose ridge (Fig. 2.6, 2.7). Lateral side of the pleural tip with rounded posterior edge, exhibiting in the anterior side a nearly concave and sparsely granulose articulating facet, which shows a shallow and wide (dorsoventral) furrow posterolaterally directed, intersected by the curved (anteriorly concave) incision of the pleural furrow ending (Fig. 2.6).
Lateral lobe of the thoracic axial ring rather weakly separated from it by a pair of shallow notches (Figs. 2.2, 4.1–4.2).
Etymology
After the greek taphos: tomb, and mimos: imitator/mimic. In regarding the infaunal molt behavior, whereby articulated exoskeletal remains are buried by the trilobite during molting, thus generating a “false tomb.”
Materials
CEGH-UNC 7055-67, 7071-77, 12739, 12743, 12745-50, 24085-87, 24089, 26129, 26131-26150, 26153 from Quebrada de Las Aguaditas and PULR 118-126 from the Sierra de Las Minitas (Rustán et al., Reference Rustán, Vaccari and Astini2011a).
Remarks
In accordance with the diagnosis by Sandford (Reference Sandford2002), Argentinian studied specimens show the main morphological features of Echidnops, namely: median spines on the occipital ring, thoracic axial rings and the first pygidial axial ring; eyes short, ∽30%–40% cephalic length, placed with anterior margin opposite ∽60% cephalic length (measured from posterior to anterior), dorso-ventral files of lenses with up to six to seven lenses, and a moderately thickened sclera; wide (tr.) glabella ∽55% cephalic width; axial furrows diverging forward ∽75°–80°; vincular furrow deep medially with lateral notches very well impressed, and first pygidial axial rings bearing spine-like ridges transversally. Genal, intergenal, and fulcral spines, frequently present in Echidnops, are also clearly exhibited by E. taphomimus n. sp.
Rustán et al. (Reference Rustán, Vaccari and Astini2011a), already noted the continuing problem of differentiating Paciphacops, Kainops Ramsköld and Werdelin, Reference Ramsköld and Werdelin1991, Viaphacops Maksimova, Reference Maksimova1972 and Ananaspis Campbell, Reference Campbell1977 (overlooking references to Echidnops). Shared cephalic characters used for the assignment to Paciphacops included those originally proposed as diagnostic of Paciphacops (Paciphacops) by Maksimova (Reference Maksimova1972): preoccipital furrow bent forward and shallowest in its middle portion; more or less reduced preoccipital lobe appressed to the base of the glabella, and cut off behind by a straight occipital furrow; anterolateral angles of glabella drawn out; occipital ring broad (sag., exsag.); and pygidium moderately well segmented with rounded-triangular shape. Additional characters included a thickened sclera (character 23 as was suggested by Ramsköld and Werdelin, Reference Ramsköld and Werdelin1991). From the new available evidence, it is clear that these characters in particular are not sufficient for recognizing Paciphacops because most of them are shared with Echidnops, as was already noted by Sandford (Reference Sandford2002). In fact, taking into account cephalic characters except occipital, fulcral, and genal spines, most of the species currently assigned to Echidnops were previously included in Paciphacops (i.e., E. taphomimus n. sp., E. crawfordae, and E. serratus (see Wright and Haas, Reference Wright and Haas1990; Ramsköld and Werdelin, Reference Ramsköld and Werdelin1991; Ebach, Reference Ebach2002; Rustán et al., Reference Rustán, Vaccari and Astini2011a, Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b). The actual taxonomic concept of Echidnops highlights the relevance of postcephalic characters in phacopid systematic, as was already stated by Sandford (Reference Sandford2002). In this regard, we consider that the knowledge of some additional postcephalic, characters in Echidnops would help to better separate it from its close relatives such as Paciphacops, Kainops, Viaphacops, and Ananaspis. Relevant postcephalic characters include a thick exoskeleton, the strong downward deflection of the thoracic pleurae through almost 90° at the fulcrum, and a trend to develop fulcral shoulders and spines.
Our taxonomic reappraisal besides suggests that some cephalic characters of alleged taxonomic relevance are only of relative importance. For example, perforated tubercles, traditionally regarded as associated with large-eyed morphs, have been regarded as diagnostic of Paciphacops (Campbell, Reference Campbell1977; Wright and Haas, Reference Wright and Haas1990). However, E. taphomimus n. sp., which has relatively small eyes, clearly exhibits perforated glabellar tubercles (Fig. 3.11). We have not included this character in the diagnosis of E. taphomimus n. sp. because it seems to be present in other congeneric species (see Wright and Haas, Reference Wright and Haas1990, fig. 3A and Sandford, Reference Sandford2002, figs. 6B, 6C, 7R, 8F–8Q, and 10A).
In addition, we recognized a wide continuum of morphological cephalic variation in holaspides of Echidnops taphomimus belonging to a single population from the same stratigraphic level (nodules from a single narrow interval in the Quebrada de Las Aguaditas and Sierra de Las Minitas), ranging from slender and narrow (tr.) cephala to wider (tr.) and more robust ones. Such variation, classically taken into account for recognition of morphs, would indicate intraspecific variability, rather than sexual, ontogenetic, or taxonomic differences as previously proposed (Campbell, Reference Campbell1977, p. 10). Given that all specimens are preserved in nodules, undistorted, a possible taphonomic or collecting bias is also definitely disregarded.
Rustán et al. (Reference Rustán, Vaccari and Astini2011a) discussed E. crawfordae (Wright and Haas, Reference Wright and Haas1990) from the late Pragian Rosedale Shale, New South Wales, Australia, as the most closely related species to E. taphomimus n. sp.. However, the strong similarities with E. wrighti Sandford, Reference Sandford2002 from the late Lochkovian of the Humevale Formation, New South Wales, were overlooked by these authors. These three species are clearly the most closely related, mainly on the basis of the strong spine development and spine distribution pattern, while all remaining members of the group lack either the genal (E. serratus Sandford, Reference Sandford2002) or the fulcral spines (E. sherwini Sandford, Reference Sandford2002 and E. hollowayi Sandford, Reference Sandford2002).
Apart from its larger size, E. taphomimus n. sp. can be differentiated from both E. crawfordae and E. wrighti by a series of fossulae toward the external edge of the palpebral lobe (Fig. 2.8–2.10, 2.13), just a pair of shallow fossulae (not a true furrow) separating weakly the lateral lobes on the thoracic axial rings; and the nearly straight and ridge-like anterior pleural band (which is orientated nearly 90° with respect to the axial axis). Moreover, E. taphomimus n. sp. differs from E. wrighti in having smaller eyes, fewer dorso-vental files of lenses (approximately 10–14 versus nearly 16, respectively), less tuberculated glabella, fulcral pleural region ∽90° angle in transverse section, and pygidium with spine-like ridges on the first axial rings instead of nodes.
Discussion
Lens arrangement
A striking character of Echidnops taphomimus n. sp. is the irregular arrangement of lenses on the visual surface of the eye. Difficulties in recognizing files and counting lenses (at least in holaspides) are caused by a progressive curvature of the upper (dorsal) part of each lens file, as well as to the intercalation of lenses between files, particularly near the central-upper region of the visual surface (Fig. 5). Curvature of lens files is particularly noticeable when counting them from anterior to posterior and vice versa. As a result, the usual phacopid regular hexagonal close packing of lenses is not recognizable in E. taphomimus, so that reporting a complete lens formula cannot be assured. Nevertheless, we present approximate lens formulae for several specimens (Table 1) because this provides an idea of the overall arrangement, and allows comparisons with previous reports of closely related species.
Irregular packing of lenses is also present in most of Echidnops species, including E. crawfordae (see Wright and Haas, Reference Wright and Haas1990, figs. 3B, 3J, 4D), E. hollowayi (Sandford, Reference Sandford2002, fig. 8I, 8N), and E. serratus (Sandford, Reference Sandford2002, fig. 10G, 10H). It is not present in E. wrighti or E. sherwini (Sandford, Reference Sandford2002, figs. 6N, 9H).
The phylogenetic significance of this character is not clear, pending a cladistic analysis to explore its taxonomic relevance in distinguishing less spinose species of Echidnops from those belonging to closely allied genera such as Paciphacops, Kainops, or Viaphacops.
Although irregularities of lens arrangement have been reported in other members of Phacopina, such contributions have focused on considering them particular isolated exceptions to the regularity or bilateral symmetry, and thus, their taxonomic value has been not explored (see cases cited by Crônier and Clarkson, Reference Crônier and Clarkson2001, p. 110). Apart from, mostly unsupported, early hypotheses on sexual dimorphism, they have been attributed either to variation in timing of lens emplacement led by a not absolutely deterministic program of development, or abnormalities, stress or injuries during development (Thomas Reference Thomas1998; Crônier and Clarkson, Reference Crônier and Clarkson2001; Crônier et al., Reference Crônier, Feist and Auffray2004; Feist et al., Reference Feist, McNamara, Crônier and Lerosey-Aubril2009, and references therein). In contrast, the clear pattern of general irregularity of lens arrangement in Echidnops is stable at a specific level (observable in all individuals belonging to each species that exhibits the character), suggesting that it might be of taxonomic relevance.
Significantly, an irregular lens pattern seems to be present in other trilobites also at a specific level. For example Reedops bronni (Barrande, Reference Barrande1846) exhibits this feature as illustrated by Alberti (Reference Alberti1970, pl.14, fig. 2b), Chlupáč (Reference Chlupáč1977, pl. XXII, figs. 4, 9–11, 17), McKellar and Chatterton (Reference McKellar and Chatterton2009, pl. 1.1, figs. 1, 3, 7, 11), and Holloway and Rustán (Reference Holloway and Rustán2012, pl. 2, fig. 6); whereas other species, such as R. cephalotes (Alberti, Reference Alberti1983) and R. pembertoni McKellar and Chatterton, Reference McKellar and Chatterton2009, clearly exhibit a regular lens pattern.
Another analogous case seems to be Struveaspis Alberti, 1966, according to illustrations from Chlupáč (Reference Chlupáč1977, pls. 30 and 31), Alberti (Reference Alberti1970, pl. 13, fig. 10), Alberti (Reference Alberti2000, pl. 2, fig. 1b), and Corbacho et al. (Reference Corbacho, Badía and Quintilla2014, pl. 1, fig. 10).
The irregular lens pattern observed in Echidnops species resembles particularly that of the Ordovician basal phacopine Ormathops Delo, Reference Delo1935. Clarkson (Reference Clarkson1971) recognized two different kinds of eye disruptions in Ormathops, namely: (Reference Abe and Lieberman1) blocks of lenses with different orientation separated by vertical lines called caesurae, and (Reference Abe and Lieberman2) changes in the way lenses are packed in the upper part of the visual surface, allowing lens packing changes and curving of the files over toward the top. Although ceasure are not present in Echidnops, the lens distribution in Echidnops is identical to the second type of disruption (i.e., packing changes) observed in Ormathops by Clarkson (Reference Clarkson1971). Echidnops taphomimus n. sp. further shares with Ormathops a roughly constant lens size from the top to the bottom of the eye, which is a unique feature among schizochroal eyes (Crônier and Clarkson, Reference Crônier and Clarkson2001). As noted by Clarkson (Reference Clarkson1971), a constant lens size would cause a geometrical constraint to a regular lens packing because the visual surface approximates a cone segment. Therefore, the lens packing irregularities observed in Echidnops and Ormathops are a simple consequence of a constant lens size.
Clarkson (Reference Clarkson1971) argued that the lens arrangement present in Ormathops was an intermediate step in the early rapid evolution of schizochroal eyes, which never reappeared in the entire history of the clade, most probably because it was not as efficient as a fully regular eye. This hypothesis was later supported by Henry et al. (Reference Henry, VizcaÏno and Destombes1992) who recognized parallel evolutionary trends in Ormathops and Toletanaspis Rábano Reference Rábano1989 characterized by older Ordovician species with irregular eye patterns and younger Ordovician ones with “true” schizochroal regular eyes. Moreover, the alleged parallelism reported by these authors would suggest a markedly polyphyletic origin for the schizochroal eyes.
The irregular lens distribution in derived Devonian phacopids such as Echidnops, challenges the idea that a regular pattern would be the best spatial option for forming a proper image, and that an irregular pattern would be simply an evolutionary step towards regular eyes. It seems plausible that such irregular eyes were an alternative viable solution to functional efficiency of the schizochroal eye, which is in turn a more parsimonious interpretation since it does not involve a polyphyletic origin for a complex structure as schizochroal eyes. These new insights deserve further investigation.
Paleobiogeography
The Devonian, particularly the Lower Devonian, southern South American basins, have highly endemic trilobite fauna shared with South Africa, Ghana, Malvinas Islands and Antarctica. The faunal similarities led to the proposal of a nearly circumpolar, southern paleobiogeographic major area, known as the Malvinokaffric Realm (Richter and Richter, Reference Richter and Richter1942). The Malvinokaffric Realm was later also recognized based on the distribution of other marine groups (Boucot and Racheboeuf, Reference Boucot and Racheboeuf1993), such as brachiopods (Boucot et al., Reference Boucot, Johnson and Talent1969), ostracods (Lethiers et al., Reference Lethiers, Racheboeuf, Baudin and Vaccari2001; Salas et al., Reference Salas, Rustán. and Sterren2013), and hyoliths (Malinky and Racheboeuf, Reference Malinky and Racheboeuf2011). The Gondwanan records of New Zealand and Australia were excluded from the Malvinokaffric Realm because of their unrelated Early Devonian marine faunas (Boucot and Racheboeuf, Reference Boucot and Racheboeuf1993).
The endemic distribution of Early Devonian Malvinokaffric trilobites is notable and observed even at a familial level in calmoniids (Eldredge and Ormiston, Reference Eldredge and Ormiston1979; Abe and Lieberman, Reference Abe and Lieberman2009), dalmanitids (Braniša and Vanék, Reference Braniša and Vanék1973; Holloway and Carvalho, Reference Holloway and Carvalho2009; Rustán and Vaccari, Reference Rustán and Vaccari2012a), and homalonotids (Simões et al., Reference Simões, Leme and Soares2009). Endemicity is even recorded within cosmopolitan genera already present during the Silurian, which record radiations within the Malvinokaffric Realm in the Lower Devonian, as is the case of the endemic aulacopleurid Maurotarion (Malvinotarion) Rustán and Vaccari, Reference Rustán and Vaccari2010.
In this paleobiogeographical context, Echidnops exhibits an unusual distribution. All Echidnops species except the Argentinian E. taphomimus n. sp., are known from Australia (Sandford, Reference Sandford2002). Thus, Echidnops represents the first trilobite connecting the Early Devonian Australian basins with the South American Malvinokaffric coeval ones, and clearly suggest a particular link between the Precordillera Basin and Southeastern Australia.
Although unique, this striking Australian-Andean distribution pattern is consistent with a few other cases such as the brachiopod Boucotia Gill, Reference Gill1969 (according Herrera, Reference Herrera1995). Among trilobites from the Argentine Precordillera indicating extra-Malvinokaffric affinities, the probably late Pragian aulacopleurid Maurotarion megacephalum Rustán and Vaccari Reference Rustán and Vaccari2012b, suggests a biogeographical connection with the Eastern Americas Realm because it was considered most closely allied to Maurotarion periergum (Haas, Reference Haas1969), from the Pragian of the Great Basin, United States (Adrain, Reference Adrain2009; Rustán and Vaccari, Reference Rustán and Vaccari2012b). Independent evidence is also provided by the distribution of other Pragian trilobites such as Reedops, a cosmopolitan biostratigraphical indicator recorded in slightly higher stratigraphical intervals than Echidnops taphomimus n. sp. (Holloway and Rustán, Reference Holloway and Rustán2012).
The remarkable distribution pattern of Echidnops challenge the classical paleobiogeographical interpretations about these basins as isolated by means of dispersal barriers, particularly during periods of low sea levels, such as the Pragian (Rustán and Vaccari, Reference Rustán and Vaccari2010). In accordance, recent data on Lower-Middle Devonian trilobites from northern South American basins (including records from Colombia and Venezuela) indicate the presence of Malvinokaffric elements in faunal associations otherwise paleobiogeographically linked to the Devonian Eastern Americas Realm (Morzadec et al., Reference Morzadec, Mergl, VillarRoel, Janvier and Racheboeuf2015).
Infaunal molting and its role in the mid Paleozoic marine revolution
Infaunal molting behavior in trilobites has been only recently recognized mainly based on specimens of E. taphomimus n. sp. studied herein (Rustán et al., Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b). The infaunal molting pattern is typically characterized by an articulated thoracopygidium, frequently exhibiting a gentle dorsal concave-up bending; the cephalon separated and placed in front of and slightly above the first thoracic segment, being nearly perpendicularly oriented up to clearly rotated backward; and the hypostome being displaced. Because no other taphonomic agents can be invoked for explaining such a distinct three-dimensional arrangement as generated onto the sea bottom, this preservational pattern has been interpreted as biologically produced because of the trilobite being entirely buried in soft sediment during the molting process (Rustán et al., Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b, fig. 2.6; Fig. 4.1–4.7). The three-dimensional pattern described, provide unequivocal evidence of this ethological feature, in contrast with, earlier, functional inferences based on plausible life style of blind trilobites (Becker and Schreiber, Reference Becker and Schreiber1994).
Because the exuviation process is the most vulnerable period of the trilobite adult life, this behavior was understood as an evolutionary acquisition related to a predator-deterrent active (ethological) strategy (Rustán et al., Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b).
The infaunal molting pattern was originally recognized in a hypothetical late Silurian-Early Devonian phacopid lineage, involving three species of Paciphacops from western Argentina, namely P. argentinus, P. sp A and P. sp B (Rustán et al., Reference Rustán, Vaccari and Astini2011a, Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b). In addition, an evolutionary defensive trend toward a thickening of the carapace, a development of spinosity and an increasing size was documented for this putative lineage. However, the present reassignment to Echidnops rejects the idea of a Siluro-Devonian evolutionary trend of Paciphacops, as suggested by Rustán et al. (Reference Rustán, Vaccari and Astini2011a, Reference Rustán, Balseiro, Waisfeld, Foglia and Vaccari2011b). Hence, on the one hand, morphological characters in E. taphomimus n. sp. might be interpreted as plesiomorphies and not necessarily as adaptive responses against predation coupled to infaunalism. On the other hand, this implies that infaunal molting should have evolved independently several times. In accordance, preliminary reports (Rustán et al., Reference Rustán, Balseiro, Iwasaki and Foglia2012a, Reference Rustán, Foglia, Balseiro and Iwasaki2012b) indicated that infaunal molting is also present in the phacopid Viaphacops orurensis (Bonarelli, Reference Bonarelli1921) and in the calmoniid Pennaia, from the Middle Devonian (Givetian?) of the Sica Sica Formation in Bolivia. Additional support comes from the phacopid “Phacops” chavelai and the calmoniid Punillaspis argentina, from the Middle Devonian Chigua Formation of Argentina (Rustán et al., Reference Rustán, Vaccari and Balseiro2015). In addition, Błażejowski et al. (Reference Błażejowski, Gieszcz, Brett and Binkowski2015) recently reported an exceptional specimen of the blind phacopid Trimerocephalus, from the Late Devonian of Poland, caught in the act of molting infaunally. Finally, a number of molt ensambles coming (mainly) from the Lower Devonian of Germany-Luxemburg (Alberti, Reference Alberti2015a, Reference Albertib), although in many cases differ from the typical infaunal molt pattern reported herein, suggest it could also be present in acastid, homalonotid, and proetid trilobites. Thus, evidences support that infaunal molting evolved independently in many different groups in mid Paleozoic seas as an analogue of the enrollment and coaptive structures earliest in the history of the trilobites (Esteve et al., Reference Esteve, Hughes and Zamora2011). This clear defensive conduct can be interpreted as evidence of escalation, in concert with the evolutionary-ecological context of the so-called mid Paleozoic marine revolution (Signor and Brett, Reference Signor and Brett1984), a major bio-event related to the sudden appearance of several groups of predators that greatly intensified predation pressure in Silurian-Devonian marine ecosystems, simultaneously triggering defensive evolutionary trends in a number of prey groups (Brett and Walker, Reference Brett and Walker2002; Brett, Reference Brett2003; Klug et al., Reference Klug, Kroeger, Kiessling, Mullins, Servais, Frýda and Turner2010). According the phylogenetic patterns preliminary observed in this major paleoecological scenario, trilobites probably played an active role in a more similar way to the evolution of infaunalism in bivalves than in irregular echinoids during the Mesozoic (Harper, Reference Harper2003).
Acknowledgments
The authors would like to thank D. Holloway for suggesting the reassigment of the studied specimens to Echidnops and to T. Becker for pointing out his contribution on infaunal molting in Trilobita; and B.G. Waisfeld, N.E. Vaccari, and Z.A. Herrera for collecting most of the studied specimens. Reviewers C. Crônier and D. Holloway greatly improved this paper. The Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) and the Universidad Nacional de Córdoba provided support and facilities used in this investigation. Financial support to J.J.R. was provided by the ANPyT (Agencia Nacional de Promoción Científica y Tecnológica)-FONCYT (grants: 2012 -PICT-1993).






