


Introduction
Chancelloriids are a group of organisms of problematic affinity, which are characterized by a sac-like body covered with spiny sclerites. Complete specimens (scleritomes) show them to be sessile, cylindrically symmetrical metazoans, having a general morphology reminiscent of that of a barrel cactus (Bengtson and Hou, Reference Bengtson and Hou2001; Janussen et al., Reference Janussen, Steiner and Zhu2002; Randell et al., Reference Randell, Lieberman and Hasiotis2005; Moore et al., Reference Moore, Li and Porter2014). They are found stratigraphically from the Terreneuvian to the Furongian in the Cambrian (Bengtson and Hou, Reference Bengtson and Hou2001; Janussen et al., Reference Janussen, Steiner and Zhu2002; Hong et al., Reference Hong, Choh and Lee2016). Well-preserved chancelloriid scleritomes from the early and middle Cambrian occur mainly in China, Canada, and the USA (Bengtson and Collins, Reference Bengtson and Collins2015). The late Cambrian witnessed a rapid decline of chancelloriids and only a few occurrences have been described (e.g., Mostler and Mosleh-Yazdi, Reference Mostler and Mosleh-Yazdi1976). By the end of the Cambrian, this problematic group had probably become extinct (Janussen et al., Reference Janussen, Steiner and Zhu2002).
The phylogenetic position of chancelloriids remains unsolved. They were first considered to be problematic fossils (Bornemann, Reference Bornemann1886; Rosén, Reference Rosén1919). Walcott (Reference Walcott1920) described chancelloriids as heteractinid sponges based on their sponge-like appearance and sclerite morphology. This taxonomy was accepted by a number of workers for more than half a century (e.g., Laubenfels, Reference Laubenfels1955; Sdzuy, Reference Sdzuy1969; Finks, Reference Finks1970; Mostler, Reference Mostler1980, Reference Mostler1985), but rejected by Goryansky (Reference Goryansky1973), who argued that chancelloriids have an external skeleton, rather than the internal skeleton typical of sponges. Chancelloriids, along with halkieriids, siphogonuchitids, and sachitids, were placed by Bengtson and Missarzhevsky (Reference Bengtson and Missarzhevsky1981) in the Coeloscleritophora. This proposal was followed by some researchers (e.g., Qian and Bengtson, Reference Qian and Bengtson1989; Bengtson et al., Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990; Bengtson, Reference Bengtson2005; Porter, Reference Porter2008; Moore et al., Reference Moore, Porter, Steiner and Li2010), but again rejected by others because of the differences in anatomy between chancelloriids and the halkieriids: the former are cylindrically symmetrical whereas the latter are bilaterally symmetrical (Conway Morris and Peel, Reference Conway Morris and Peel1995; Vinther and Nielsen, Reference Vinther and Nielsen2005). Subsequently, chancelloriids have been considered to be related to tunicates (Mehl, Reference Mehl1996), cnidarians (Randell et al., Reference Randell, Lieberman and Hasiotis2005), mollusks (Conway Morris and Peel, Reference Conway Morris and Peel1995; Vinther and Nielsen, Reference Vinther and Nielsen2005; Conway Morris and Caron, Reference Conway Morris and Caron2007; Vinther, Reference Vinther2009), or to lie elsewhere in the early branches of the Metazoa (Janussen et al., Reference Janussen, Steiner and Zhu2002; Sperling et al., Reference Sperling, Pisani and Peterson2007). After detailed studies, chancelloriids, with potential eumetazoan plesiomorphy (coelosclerites), were suggested likely to belong to a parayphyletic Coeloscleritophora (Bengtson and Collins, Reference Bengtson and Collins2015).
In recent years, more and more articulated scleritomes of chancelloriids have been described in early and middle Cambrian faunas, which have definitely helped our understanding of the original appearance of these problematic animals. Here, we describe a new species of chancelloriid and a new chancelloriid-like metazoan. Both species occur in great numbers and are well preserved.
The feeding mode of chancelloriids remains unclear; they have been considered variously as predators, filter feeders, feeders of dissolved organic matter, and symbiotrophs (Chen et al., Reference Chen, Zhou, Zhu and Yeh.1996; Bengtson and Hou, Reference Bengtson and Hou2001; Janussen et al., Reference Janussen, Steiner and Zhu2002; Randell et al., Reference Randell, Lieberman and Hasiotis2005; Sperling et al., Reference Sperling, Pisani and Peterson2007; Kloss et al., Reference Kloss, Dornbos and Chen2009). The feeding mode of chancelloriids is discussed here, based on these well-preserved specimens.
Geological setting
The Guanshan biota occurs in the Wulongqing Formation (Cambrian Series 2, Stage 4), which is widely distributed in eastern Yunnan, including the Kunming-Wuding and Malong-Yiliang areas. The best-studied sections are the Gangtoucun (25.1021°E, 102.7213°N) and Gaoloufang (24.95916°E, 102.80539°N) sections of Kunming, the Sapushan (25.58819°E, 102.41071°N) section of Wuding, the Lihuazhuang (24.93173°E, 103.05454°N) section of Yiliang and the Wulongqing (25.3975°E, 103.49648°N) section of Malong (Luo et al., Reference Luo, Fu, Hu, Li, Hou, Liu, Chen, Li, Pang and Liu2008; Hu et al., Reference Hu, Zhu, Luo, Steiner, Zhao, Li, Liu and Zhang2013) (Fig. 1).

Figure 1 Location map of the best studied fossil sections of Guanshan Biota: 1, Gaoloufang; 2,Gangtoucun; 3, Sapushan; 4, Liuazhuang; 5, Wulongqing.
The specimens described in this paper were excavated from the Wulongqing Formation, Palaeolenus biozone, Gaoloufang section, which is ~15 km southeast of Kunming (Fig. 1). The basal Cambrian succession in this section mainly consists of the Hongjingshao and Wulongqing formations. The latter comprises several layers of dark brown sandstones with interbedded yellow-greenish, thin-bedded argillaceous shale (Hu et al., Reference Hu, Zhu, Steiner, Luo, Zhao and Liu2010, Reference Hu, Zhu, Luo, Steiner, Zhao, Li, Liu and Zhang2013). Three kinds of chancelloriids were reported from the Guanshan biota from the Yunnan province. They are: Archiasterella sp., Allonnia sp., and a new form of chancelloriid (Hu et al., Reference Hu, Zhu, Luo, Steiner, Zhao, Li, Liu and Zhang2013). The last two are confirmed herein and discussed below.
Materials and methods
All the fossils described here were collected in the first two months of 2017, and comprise more than100 specimens in all. Some were prepared using a needle under a binocular microscope, revealing parts covered by the matrix. Digital photographs were taken by using a Canon EOS 5D MkIII camera and 50 mm macro lens, dry under cross-polarized light and processed in Adobe Photoshop CS 5. Terminology follows Sdzuy (Reference Sdzuy1969), Randell et al. (Reference Randell, Lieberman and Hasiotis2005), and Bengtson and Collins (Reference Bengtson and Collins2015).
Repository Information and Institutional Abbreviation
All the specimens are housed at the School of Earth Sciences and Resources, China University of Geosciences (Beijing) (CUGB). The abbreviations in the specimen number refer to the location of the fossil section (YN, the province of Yunnan, GLF, the village where the quarry lies) and biozone (PAL, Palaeolenus zone).
Systematic paleontology
Phylum and Class uncertain
Order Chancelloriida Walcott, Reference Walcott1920
Family Chancelloriidae Walcott, Reference Walcott1920
Genus Allonnia Doré and Reid, Reference Doré and Reid1965
Type species
Allonnia tripodophora Doré and Reid, Reference Doré and Reid1965 (based on isolated sclerites).
Diagnosis
As in Bengtson and Collins (Reference Bengtson and Collins2015).
Allonnia tenuis new species
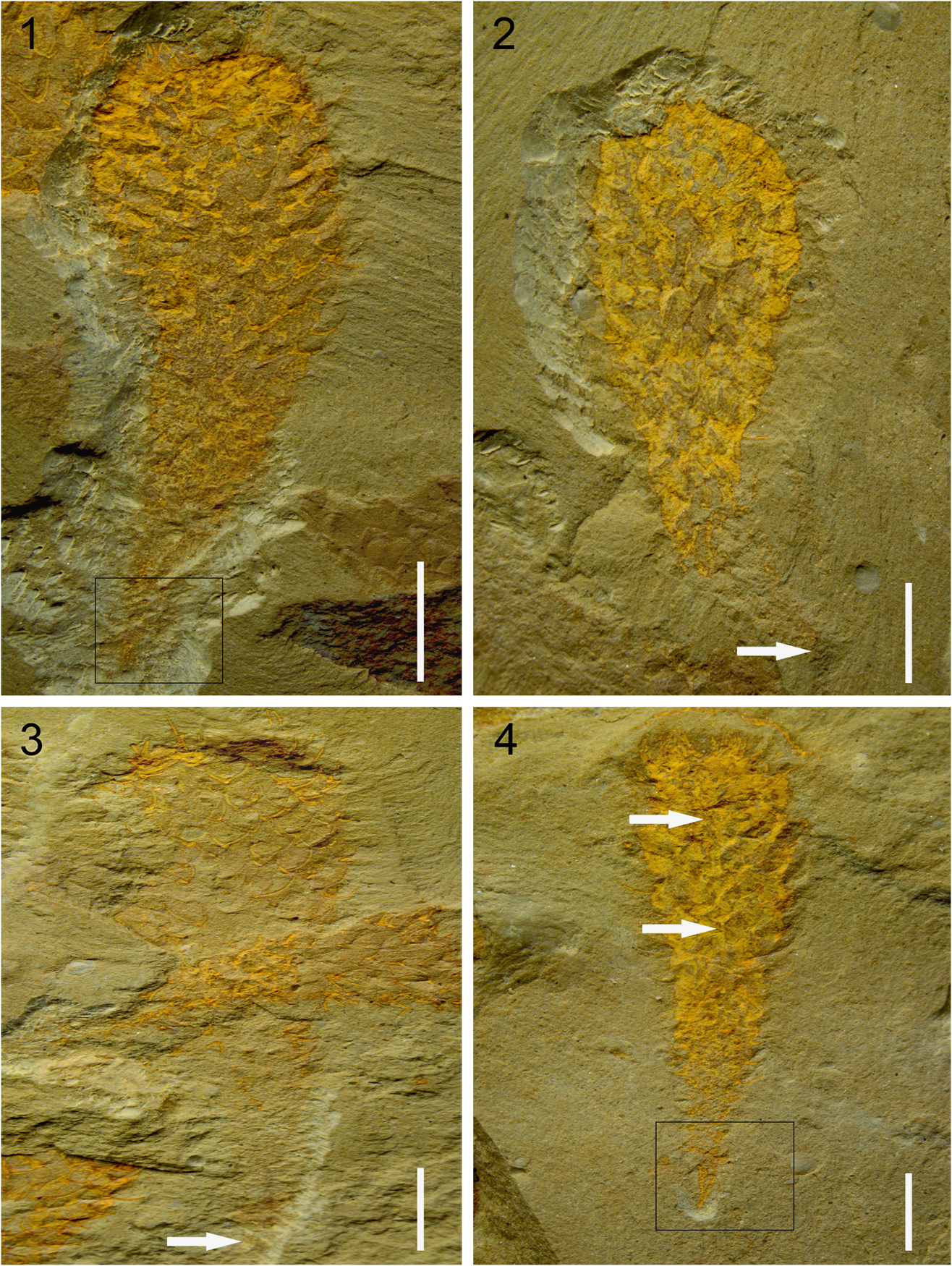
Figure 2 Allonnia tenuis n. sp. (1) holotype, YN-GLF-PAL-43; box indicates magnified view shown in Figure 3.3; (2) YN-GLF-PAL-45, white arrow points to the basal point; (3) YN-GLF-PAL-47, white arrow points to the basal point; (4) YN-GLF-PAL-06, white arrows point to the longitudinal fold; box indicates magnified view shown in Figure 3.2. Scale bars=5 mm.
Holotype
The specimen YN-GLF-PAL-3 (Fig. 2.1) is hereby designated as the holotype for this species. The holotype specimen is found in Cambrian Series 2, Stage 4, Palaeolenus Zone, Wulongqing Formation, Gaoloufang section, Kunming City, Yunnan Province, China.
Diagnosis
Small, baseball bat-like Allonnia with slender 3+0 sclerites (Bengtson and Collins, Reference Bengtson and Collins2015) and inconspicuous apical tuft; sclerites densely arranged around the apical end, but loosely elsewhere on the body; stalk slender, prolonged and well-developed, with the lower part devoid of sclerites; the stalk tapers to a point.
Description
The overall profile of Allonnia tenuis n. sp. is usually baseball bat-shaped, with the upper part obviously wider and more inflated than the lower part (Fig. 2). The lower part constricts abapically into a slender and prolonged stalk, which evenly narrows to a basal point (Figs. 2, 3). This species is the smallest among the whole-body preserved Allonnia in the literature: Allonnia phrixothrix Bengtson and Hou, Reference Bengtson and Hou2001; Allonnia tintinopsis Bengtson and Collins, Reference Bengtson and Collins2015; and Allonnia erjiensis Yun, Zhang, and Li, Reference Yun, Zhang and Li2017. The height is 23–29 mm and the maximum width (on upper part of body) is 5–10 mm; the minimum width of 0.1–0.2 mm lies at the basal point. Within this general morphology, some variation in body profiles exists: (1) YN-GLF-PAL-06 (Fig. 2.4) shows a more slender body profile: the upper part is less inflated while the lower part is similar to others; (2) YN-GLF-PAL-01 (Fig. 4.1) is spindle-shaped and the maximum width lies at the center while both ends appear narrower; and (3) three specimens show that the main body is deflected from the basal point (Figs. 2.1, 2.2, 3.1).

Figure 3 Allonnia tenuis n. sp. (1) YN-GLF-PAL-48; (2) detail of Figure 2.4 (position marked by frame); (3) detail of Figure 2.1 (position marked by frame). White arrows point to the basal point and yellow arrows point to the stalk’s basal part devoid of sclerites. Scale bars=5 mm for (1), 1 mm for (2, 3).

Figure 4 Allonnia tenuis n. sp. (1) YN-GLF-PAL-01; (2) YN-GLF-PAL-07. Scale bars=5 mm.
Sclerites of this new species, comprised of two lateral rays and one ascending ray, belong to the sclerite mode of Allonnia: 3+0 (Bengtson and Collins, Reference Bengtson and Collins2015). Rays are slender with their tips toward the apical end of the body. Usually only the lateral rays are preserved on the body surface, which thus have the shape of a lyre, similar to Al. tintinopsis (Bengtson and Collins, Reference Bengtson and Collins2015, fig. 2). The fully developed ascending rays can only be observed on the folded body margin where they protrude laterally, with the tips curved adapically (Fig. 5.2, 5.3). Some specimens show that the ascending ray on the body surface leaves a tiny three-dimensionally preserved basal part or a pit beside the basal disk of the lateral rays (Fig. 5.4). A few isolated sclerites (Fig. 6) are also preserved only with lateral rays, and longitudinal cracks along the lateral ray, thus exposing the internal cavity, which is one of the defining features of the chancelloriids.

Figure 5 Allonnia tenuis n. sp. (1) YN-GLF-PAL-34; (2) YN-GLF-PAL-66, white arrow points to an ascending ray on the body margin; (3) YN-GLF-PAL-44, white arrow points to an ascending ray on the body margin; (4) detail of (1) (position marked by frame), white arrow points to the three-dimensionally preserved basal part of an ascending ray and yellow arrow points to the pit left by an ascending ray. Scale bars=5 mm for (1), 1 mm for (2–4).

Figure 6 Isolated sclerites of Allonnia tenuis n. sp. (1) YN-GLF-PAL-12; (2) YN-GLF-PAL-09. White arrows point to the internal cavity of a lateral ray. Scale bars=1 mm.
Due to the missing ascending ray, the size of the sclerites on the body surface is defined herein only by the two lateral rays. Measurements show that on the stalk, the average ray length is 0.8 mm increasing to 1.5 mm on the upper part of the body. The average length of the ascending ray preserved on the folded margin increases from 1 mm on the stalk to 2 mm on the upper part. The mean angle between lateral rays increases from 109° to 125°. Based on these features, the sclerite size of this new species might increase adapically, but a size plateau, as observed in Allonnia tintinopsis (Bengtson and Collins, Reference Bengtson and Collins2015, fig. 18), cannot be obtained.
Most of the specimens show that sclerites are distributed over the body surface with a spacing of 1–1.5 mm, and only a few difer: smaller in one specimen (0.5–0.6 mm in YN-GLF-PAL-06, Fig. 2.4) and wider in two specimens (1.5–1.8 mm in YN-GLF-PAL-01, Fig. 4.1; 2 mm in YN-GLF-PAL-07, Fig. 4.2). A remarkable feature of Allonnia tenuis n. sp. can be observed from the specimens preserved with the whole stalk and the basal point: the lowermost part of stalk is devoid of sclerites (Fig. 3).
Compared to other specimens, the sclerites of YN-GLF-PAL-07 (Fig. 4.2) are clearly bigger both in size and angle: adapically the length of lateral ray increases from 1.5 to 2 mm and the angle in between from 117° to 130°.
The integument of this new species is smooth, and no granules, such as those observed in Al. phrixothrix (Bengtson and Hou, Reference Bengtson and Hou2001, figs. 7A, 8C), Al. tintinopsis (Bengtson and Collins, Reference Bengtson and Collins2015, fig. 31.1–31.3), and Al. erjiensis (Yun et al., Reference Yun, Zhang and Li2017, fig. 3A, 3B), can be seen over the whole body surface, but longitudinal folds are rather common, e.g. YN-GLF-PAL-06 (Fig. 2.4). As with other species of chancelloriids, openings, except for the apical orifice, cannot be observed in the integument of Allonnia tenuis n. sp.
The apical tuft and orifice are tiny and inconspicuous in Allonnia tenuis n. sp. They are difficult to discern in specimens compressed parallel to the matrix because the long, slender rays of the 3+0 sclerites are densely distributed at the apical end and obscure the structures behind (Fig. 7.1–7.3). YN-GLF-PAL-03 (Fig. 7.4) is a specimen compressed top-down and clearly shows the apical structures. A discernible subcircular orifice is located at the center and is surrounded concentrically by lyre-shaped sclerites. Closer to the orifice, the sclerites become denser and smaller. At the very edge of the orifice, the sclerites are modified to be a single element.

Figure 7 Allonnia tenuis n. sp. (1) YN-GLF-PAL-05, white arrow points to the densely distributed 3+0 sclerites at the apical end; (2) YN-GLF-PAL-21, white arrow points to the inconspicuous tuft; (3) YN-GLF-PAL-31, white arrow points to the inconspicuous tuft; (4) YN-GLF-PAL-03, white arrow points to the apical orifice and yellow arrow points to the single-elemented sclerites around the apical orifice. Scale bars=5 mm for (1–3), 1 mm for (4).
Etymology
From the Latin word tenuis, slender, referring to the slender stalk of this species.
Materials
89 specimens including the holotype.
Remarks
More than 80 nominal species and 13 genera of chancelloriids have been described in the literature, but many taxa are based on isolated sclerites (Moore et al., Reference Moore, Li and Porter2014; Bengtson and Collins, Reference Bengtson and Collins2015). Although having distinct features, these isolated sclerites cannot provide much information about the composition of the scleritomes. Three genera of Chancelloriidae (Chancelloria Walcott, Reference Walcott1920, Allonnia Doré and Reid, Reference Doré and Reid1965, and Archiasterella Sdzuy, Reference Sdzuy1969) have hitherto been found distributed worldwide and established or restudied based on whole-body preservation (Bengtson and Collins, Reference Bengtson and Collins2015). As the most distinctive feature, the morphology of sclerites, including the number and arrangement of rays, is commonly used to distinguish the genera and species of chancelloriids. Allonnia is characterized by a 3+0 arrangement of sclerites, long and apically directed rays, an apical orifice, and a tuft (Bengtson and Collins, Reference Bengtson and Collins2015), all of which can be observed in the new species, and therefore its genus placement is confirmed.
Apart from the type species, Allonnia tripodophora Doré and Reid, Reference Doré and Reid1965, ten other species and an indeterminate species have been recognized as Allonnia. Among these, only three species have been studied based on whole-body preservation: Al. phrixothrix Bengtson and Hou, Reference Bengtson and Hou2001; Al. tintinopsis Bengtson and Collins, Reference Bengtson and Collins2015; and Al. erjiensis Yun et al., Reference Yun, Zhang and Li2017. The species described herein is different from others in that it possesses the following features: the body is baseball bat-shaped; the tuft inconspicuous; the stalk slender, and is prolonged and devoid of sclerites near the basal part. These features help distinguish Al. tenuis n. sp. from other whole-body preserved species and establish it as a new one, and are discussed in more detail below.
Perfectly preserved specimens always have a slender stalk and a narrow point located at its basal end (Figs. 2, 3), which indicates that the stalk is the fixed structure of Allonnia tenuis n. sp., instead of being a possible temporary contraction of the body, as is seen in Al. tintinopsis (Bengtson and Collins, Reference Bengtson and Collins2015).
Besides the slender profile, another distinct feature of the stalk is that the lowermost part (~3 mm above the basal point in the holotype) is devoid of sclerites (Fig. 3). Compared to the natural exposure of the upper part of the body, the lower part can only be exposed by removing the overlying sediment (i.e., when slabs bearing the specimens are split during field excavation, the upper part of this new species is always exposed so it needs no further preparation in the laboratory; this is called the “natural exposure”). These features suggest that Al. tenuis n. sp. might be a kind of sessile benthos with its lowermost part being implanted in the sediment. Although no specimens of this new species have been found anchored to shell debris nor other metazoans (e.g. sponges), as observed in Al. tintinopsis (Bengtson and Collins, Reference Bengtson and Collins2015, fig. 28.1), an epizoan life mode cannot be excluded.
Allonnia tenuis n. sp. is numerous in the study material and usually found distributed in groups (Fig. 8); neither larger bodies nor larger isolated sclerites have been found; autochthonous burial, which could help preserve the original appearances of organisms, is the main preservation of Guanshan biota (Hu et al., Reference Hu, Zhu, Luo, Steiner, Zhao, Li, Liu and Zhang2013), all of which indicate that this new species is constant in small size, instead of being the juvenile of larger ones.

Figure 8 Allonnia tenuis n. sp. specimens on the same slab. White arrow points to specimen YN-GLF-PAL-47. Scale bar=10 mm.
Although YN-GLF-PAL-07 (Fig. 4.2) shows a different size and spacing of the sclerites, the overall morphology and size of the body are quite similar to other specimens, even the variation of the sclerite size (adapically the sclerite size increases) over the body surface remains the same. These indicate that the differences belong to intraspecific variation.
Chancelloriids have long been interpreted to have an apical orifice connecting the internal body cavity and the outside environment, but little evidence of this has been forthcoming because most of the specimens are disarticulated sclerites. Among articulated scleritomes, most are laterally compacted, so the orifice cannot be discerned clearly. Only Al. tintinopsis, from the middle Cambrian Burgess Shale, clearly shows the morphology and features of the apical orifice (Bengtson and Collins, Reference Bengtson and Collins2015, fig. 21). Likewise, most of the articulated scleritomes of chancelloriids from the early Cambrian, such as those found in the Chengjiang Lagerstätte, are preserved laterally, and no clear features of the apical end can be seen. Only one specimen of Allonnia junyuani Janussen et al., Reference Janussen, Steiner and Zhu2002 (synonymous with Al. phrixothrix Bengtson and Hou, Reference Bengtson and Hou2001) is compacted top-down and shows some sclerites radially arranged at the apical end (Kloss et al., Reference Kloss, Dornbos and Chen2009). Hitherto, all known specimens of chancelloriids from the Guanshan Biota (e.g., Luo et al., Reference Luo, Fu, Hu, Li, Hou, Liu, Chen, Li, Pang and Liu2008; Hu et al., Reference Hu, Zhu, Luo, Steiner, Zhao, Li, Liu and Zhang2013) are compacted laterally and the apex is densely ornamented with sclerites. YN-GLF-PAL-03 (Fig. 7.4) is the first specimen clearly showing the structures of the apical orifice in chancelloriids from the Guanshan Biota which is morphologically similar to that of Al. tintinopsis (e.g,. Bengtson and Collins, Reference Bengtson and Collins2015, figs. 21, 26). These observations confirm that the apical orifice, connecting the internal body cavity with the outside environment, is an integral part in chancelloriids (especially Allonnia from both early and middle Cambrian).
Phylum uncertain
? Order Chancelloriida Walcott, Reference Walcott1920
Genus Nidelric Hou et al., Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014
Type species
Nidelric pugio Hou et al., Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014.
Diagnosis
Chancelloriid-like metazoan with a subovoid body profile; spines are single-element, triangular, and outward projecting. Spine surface devoid of any ornament or bearing triangular scales that are absent from the spine base.
Nidelric gaoloufangensis new species

Figure 9 Nidelric gaoloufangensis n. sp. (1) YN-GLF-PAL-17, white arrow points to the area where marginal spines are normal or skewed to the body margin, frame 1 shows the spines at the abapical end; (2) detail of (1) (position marked by frame 2); (3) detail of (1) (position marked by frame 3), white arrow points to the broken mark on the top of a node-like structure. (4) detail of (1) (position marked by frame 4), white arrow points to the longitudinal groove in the spine. Scale bars=5 mm for (1), 1 mm for (2, 3), 0.5 mm for (4).

Figure 10 Nidelric gaoloufangensis n. sp. (1) YN-GLF-PAL-18; (2) YN-GLF-PAL-99, white arrow points to the depression on the margin; (3) YN-GLF-PAL-100, white arrow points to the depression on the margin and yellow arrow points to the transverse folds on the body surface, frame 1 shows the spines at the abapical end; (4) detail of (1) (position marked by frame 4); (5) detail of (1) (position marked by frame 5); (6) detail of (2) (position marked by frame 6), white arrow points to the tuft-like structure at the apical end; (7) detail of (2) (position marked by frame 7), white arrows point to the longitudinal folds on the body surface; (8) detail of (3) (position marked by frame 8), white arrows point to the spines preserved on the body surface. Scale bars=5 mm for (1–3), 1 mm for (4–8).

Figure 11 Nidelric gaoloufangensis n. sp. (1) YN-GLF-PAL-16, white arrows point to the spines preserved on the body surface, frame 1 shows the spines at the abapical end; (2) YN-GLF-PAL-98, white arrow points to the tuft-like structure at the apical end; (3) YN-GLF-PAL-14, white arrow points to the tuft-like structure at the apical end. Scale bars=5 mm.

Figure 12 Nidelric gaoloufangensis n. sp. (1) YN-GLF-PAL-101, white arrow points to the relief and yellow arrows point to the depressions on the body surface; (2) detail of (1) (position marked by frame 2), white arrow points to the longitudinal groove in the spine; (3) YN-GLF-PAL-19, white arrow points to the relief and yellow arrow points to the depression on the body surface. Scale bars=5 mm for (1, 3), 1 mm for (2).

Figure 13 Nidelric gaoloufangensis n. sp. (1) YN-GLF-PAL-102, white arrow points to one of the node-like structures and yellow arrow points to one of the pits on the body surface; (2) YN-GLF-PAL-106, white arrow points to one of the node-like structures and yellow arrow points to one of the pits on the body surface; (3) YN-GLF-PAL-108, white arrow points to the relief and yellow arrow points to the depression on the body surface; (4) YN-GLF-PAL-20. Scale bars=5 mm.

Figure 14 Nidelric gaoloufangensis n. sp. specimens on the same slab. White arrow points to the holotype, YN-GLF-PAL-17. Scale bar=10 mm.
Holotype
The specimen of YN-GLF-PAL-17 (Fig. 9.1) is hereby designated as the holotype for this species. The holotype specimen is found in Cambrian Series 2, Stage 4, Palaeolenus Zone, Wulongqing Formation, Gaoloufang section, Kunming City, Yunnan Province, China.
Diagnosis
Small species of Nidelric bearing single-element spines; body profile subovoid with the apical end narrowest; a tuft-like structure, formed by at least four oriented spines with their tips projecting towards the axis of the body, is located at the apical end; both ends gently curved; spine surface devoid of any ornament.
Description
Nidelric gaoloufangensis n. sp. is subovoid in general body profile, with the upper end narrower than the lower end (Figs. 9.1, 10.1). Height ranges from 11 mm (YN-GLF-PAL-98, Fig. 11.2) to 24 mm (YN-GLF-PAL-18, Fig. 10.1) and the greatest width (at the center of the body) ranges from 4 mm to11 mm. Different body profiles also exist (e.g., YN-GLF-PAL-99 is slenderer than others, with the margin in the center area being depressed to some extent, Fig. 10.2; YN-GLF-PAL-100 is also depressed in the center area, which gives the whole body a shoe shape, Fig.10.3).
Spines are of the single-element type are best preserved along the folded body margin (Figs. 9.1, 10.1–10.3, 11.1–11.3, 12.1, 13). Only a few specimens show some spines preserved on the body surface (e.g., YN-GLF-PAL-100, Fig. 10.8; YN-GLF-PAL-16, Fig. 11.1). Spines are broadly based and triangular in profile (Figs. 9.1, 9.4, 10.4, 10.6, 12.2). Their length ranges from 1 to 2 mm and the average width of the base is ~0.5 mm.
Most of the spines on both sides of the body margin have their tips projected towards the apical end (Figs. 9.1, 10.1, 10.2), whereas others are either normal or skewed to the body margin with their tips projecting outwards (Fig. 9.1). At least four spines are located at the apical end, with their tips towards the axis of the body, and form an inconspicuous structure (Figs. 9.2, 10.5, 11.2, 11.3), which is quite similar to the tuft of chancelloriids (e.g., Allonnia tenuis n. sp.; Fig.7.2, 7.3). Spines also exist at the abapical end and, bounded by the axis of the body, these spines project at an acute angle from the body surface (Figs. 9.1, 10.3, 11.1). Some highly three-dimensionally preserved spines (Figs. 9.4, 12.2) are cracked along the axis and show longitudinal grooves that are quite similar with those internal cavities found in the lateral rays of Allonnia tenuis n. sp. (Fig.6).
Folds, relief, and depressions can be discerned on the body surface of Nidelric gaoloufangensis n. sp. (e.g., longitudinal folds [Fig. 10.7], transverse folds [Fig. 10.3], relief, and depressions [Figs.12.1, 12.3, 13.3]). One specimen (Fig. 13.4) shows a greatly twisted body, but no crack can be observed on the surface. Due to the flexible and ductile body, this new species can be preserved in different body profiles.
Node-like structures and pits can be observed on the body surface (Figs. 9.1, 13.1, 13.2) and they are subcircular in profile with diameters ranging from 0.4 to 0.7 mm, and average spacing between of 0.5 mm. These “nodes” are not complete structures because broken marks are discernible on their tops (Fig. 9.3).
Etymology
After the Gaoloufang section, where the specimens were collected.
Materials
18 specimens including the holotype.
Remarks
Nidelric gaoloufangensis n. sp. was mentioned by Hu et al. (Reference Hu, Zhu, Luo, Steiner, Zhao, Li, Liu and Zhang2013) and considered to be a new kind of chancelloriid, but no detailed studies were provided. Hou et al. (Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014) described a chancelloriid-like metazoan from the Chengjiang Lagerstätte and established the genus Nidelric, which has many similarities to the metazoan described here. Nidelric pugio Hou et al., Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014 is the type and only other known species. The diagnosis of Nidelric is emended from Hou et al. (Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014), based on the complexity of spines, which is discussed below.
Both Nidelric pugio and N. gaoloufangensis n. sp. are more or less ovoid in profile. The spines, which consist of a single element, are triangular and broadly based, with most being preserved along the body margin. Only a few spines can be discerned on the body surface. Some spines are normal or skew to the margin, and the tips of most spines are deflected towards the narrower end of the body. Based on these similarities, the metazoan in this paper is placed in the genus Nidelric. This indicates that Nidelric is not restricted to the Chengjiang Lagerstätte and, instead, this taxon is rather long-lived and was widespread in early Cambrian Chinese faunas (Hou et al., Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014).
The most obvious difference between Nidelric pugio and N. gaoloufangensis n. sp. lies in the complexity of the spines. The spines of N. pugio are more complex, and are covered with small triangular scales on the upper or lower surface; the spines of N. gaoloufangensis n. sp. are relatively simple and devoid of ornamentation. The size of the spines in N. pugio increases from the narrower end to the broader end of the body, whereas the spines of N. gaoloufangensis n. sp. do not show this pattern. Both ends of the body of N. pugio are more or less triangular in profile, whereas the body of N. gaoloufangensis n. sp. has gently curved ends. Based on these differences, the metazoan in this paper should be established as a new species of Nidelric.
Although all the specimens show that N. gaoloufangensis n. sp. is small in body size (the maximum height is 24 mm), the size variation between individuals can be as much as twice that, (e.g., YN-GLF-PAL-18, Fig. 10.1; YN-GLF-PAL-98, Fig. 11.2). Without major morphological differences, this kind of variation should be considered either intraspecific or possibly ontogenetic.
Archiasterella fletchergryllus Randell, Lieberman, and Hasiotis, Reference Randell, Lieberman and Hasiotis2005 shows short stalks on the body surface, which were considered to be the joints between the sclerites and the integument (Randell et al., Reference Randell, Lieberman and Hasiotis2005). These structures were first discovered and interpreted by Chen and Zhou (Reference Chen and Zhou1997). Similar structures can also be observed on the body surface of N. gaoloufangensis n. sp. Both the node-like structures and pits of N. gaoloufangensis n. sp. have the same diameter range (0.4–0.7 mm), which is very similar with the base width (0.5 mm) of the spines preserved on the body margin and surface. More importantly, broken marks can be observed on almost all the tops of the node-like structures. This strongly suggests that the node-like structure and pit are the remains of broken spines: the node-like structure is the basal part and the pit is formed when the whole spine is removed. These remains are quite similar to those found in Allonnia tenuis n. sp. (Fig. 5.4). The circular structure seen at the proximal end of a marginal spine of Nidelric pugio (Hou et al., Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014, figs. 1d, 2) further confirms this relationship between the spines and these structrures.
A concentration of spines was found at the broader end of Nidelric pugio and thought to be possibly analogous to the tuft seen in chancelloriids (Hou et al., Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014). Conversely, the whole-body preserved specimens of N. gaoloufangensis n. sp. show a clear tuft-like structure at the narrower end and spines at the broader end, being bounded by the axis of the body, almost project towards left and right, which are apparently not the classic feature of a tuft. Besides, most of the marginal spines project towards the tuft-like structure, and this is consistent with the general sclerite orientation of chancelloriids. All these features strongly suggest that the narrower end with the tuft-like structure should be the apical end. But still another problem remains: no stalk or basal attachment, which could indicate a definite abapical end, has been found in either species of Nidelric.
Discussion
Preservation of Nidelric gaoloufangensis n. sp
N. gaoloufangensis is preserved in yellowish green mudstones and, due to iron oxide, the fossils show reddish-brown staining on the body surface, which is one of the characteristic preservational features of Guanshan fossils (Hu et al., Reference Hu, Zhu, Steiner, Luo, Zhao and Liu2010). The body is laterally compressed with folds, reliefs, and pits being clearly discernible on the surface.Nidelric gaoloufangensis n. sp. is generally preserved in groups on the mudstone slab with individuals of different sizes mixed together (Fig. 14). Individuals are preserved articulated, and some incomplete individuals (e.g., Fig. 13.1, 13.2) were formed due to the cracking of the slab during the excavation. All these indicate that N. gaoloufangensis is buried in situ, and this is also the main preservation of Chengjiang Lagerstätte, which yields N. pugio. Under such preservation, no internal organs have been found in either species of Nidelric, which indicates that that Nidelric was a metazoan without internal organs, as proposed by Hou et al. (Reference Hou, Williams, Siveter, Gabbott, Holwell and Harvey2014), sounds reasonable.
Spines on the body surface of N. gaoloufangensis n. sp. are generally broken and missing, leaving node-like structures and pits (e.g., Fig. 13.2). Most of the spines along the folded body margin are generally preserved in flat and triangular profile. Some spines are preserved three-dimensionally and show longitudinal grooves (Figs. 9.4, 12.2), suggesting that the morphology of the spines was not flat, but that they were deformed during preservation. Considering the subcircular node-like structures and pits preserved on the body surface of N. gaoloufangensis n. sp. and the circular structures at the base of some spines of N. pugio, the original appearance of Nidelric spines would have been a cone shape with the tips being deflected somewhat, which is quite similar to the sclerite rays of chancelloriids.
Affinities of Nidelric
Interpreting Nidelric as an unusual form of chancelloriid is reasonable based on their similarities and differences. Similarities include: (1) the cylindrically symmetrical body and flexible body surface; (2) internal cavity inside chancellorid sclerite rays and possible internal cavity inside the spines of N. gaoloufangensis n. sp.; (3) single-element spines in both Nidelric and some whole-body preserved chancelloriids (e.g., the modified ones around the apical orifice in Allonnia tintinopsis; Bengtson and Collins, Reference Bengtson and Collins2015, fig. 22.3); and (4) no ostium-like openings on body surfaces. Differences include: (1) body profiles; (2) the complexities and orientations of sclerites (e.g., the spines of Nidelric are less complicated and not all the spine tips project towards the apical end of the body, but whole-body preserved chancelloriids show that at least one ray of the sclerite projects apically); and (3) presence or absence of a stalk or basal attachment (e.g., the whole-body preserved chancelloriids always show a stalk or basal attachment, which cannot be obviously observed in the specimens of Nidelric).
Feeding mode of chancelloriids and Nidelric
The feeding mode of chancelloriids has long been in discussion. Although many proposals have been put forward, none has been accepted universally. Based on the fact that the apical orifice is the only opening in the body, some researchers (e.g., Chen et al., Reference Chen, Zhou, Zhu and Yeh.1996) have suggested that chancelloriids captured prey or organic grains by expanding the body to suck seawater into the internal body cavity, and then compressed the body to expel the waste water. A similar proposal that chancelloriids might periodically have contracted the lower part of the body to extrude waste material was put forward by Bengtson and Collins (Reference Bengtson and Collins2015) and supported to some extent by Yun et al. (Reference Yun, Zhang and Li2017). This kind of feeding requires at least the integument to be flexible, and chancelloriids, indeed, possess a flexible integument as evidenced by folds in Al. phrixothrix (Bengtson and Hou, Reference Bengtson and Hou2001, fig. 4), longitudinal stripes in Al. tintinopsis (Bengtson and Collins, Reference Bengtson and Collins2015, fig. 32), transverse stripes in Al. erjiensis (Yun et al., Reference Yun, Zhang and Li2017, fig.2), and the greatly bent body without apparent cracks seen in Chancelloria eros Walcott, Reference Walcott1920 (Bengtson and Collins, Reference Bengtson and Collins2015, fig. 9), Al. tintinopsis (Bengtson and Collins, Reference Bengtson and Collins2015, fig. 27), and Al. erjiensis (Yun et al., Reference Yun, Zhang and Li2017, fig.1). The specimens of Al. tenuis n. sp. seem to support this proposal. This species also has flexible integument, which can be observed by the deflection of the main body from the basal point without cracks (Figs. 2.1, 2.2, 3.1) and the longitudinal folds over the body surface in some specimens. YN-GLF-PAL-06 (Fig. 2.4) has a similar height as the others, but the upper part is apparently less inflated in profile. More importantly, a long longitudinal fold can be discerned on the upper part of the body surface, and the spacing between sclerites around the fold is smaller, which produces a higher density of sclerites. YN-GLF-PAL-01 (Fig. 4.1) is a specimen with the lower part missing, but its main body is apparently more inflated and the spacing between sclerites is wider (lower density). All these features suggest that, with the assistance of a flexible integument, chancelloriids, especially Allonnia from early and middle Cambrian, may have had the ability to partially or completely expand and compress the body, which could help suck seawater in and expel it out. Chancelloriids may have fed during this process, but this needs to be confirmed by more solid evidence.All specimens of Nidelric are laterally compacted and no apical orifice can be discerned, but some spines form a tuft-like structure at the apical end, which is quite similar to chancelloriids. Therefore, the presence of an apical orifice cannot be excluded from this metazoan, and given the other morphological similarities between Nidelric and other chancelloriids (e.g., Allonnia), their feeding modes might have also been very similar.
Conclusions
New well-preserved specimens not only provide new members of the chancelloriids, but also help disclose the original appearance of this enigmatic group. Chancelloriids, especially Allonnia from early and middle Cambrian, may have had the ability to partially or completely expand and compact the body, which might have played an important role in feeding. Based on new specimens, Nidelric shows many similarities with chancelloriids in the morphology of body and spines, which suggests that it is an unusual form of chancelloriid, but still this needs to be confirmed by more solid evidence.
Acknowledgments
This research is supported by the Strategic Project of Science and Technology of Chinese Academy of Sciences (XDB03010403) and the State Scholarship Fund of China (41272030, 40972026). We appreciate the constructive comments from reviewers and the editor, which helped improve the manuscript significantly. We thank L. Fei and J. Lei for their assistance during field work.














