Introduction
Knowledge of lepidosaurian (lizards, snakes, amphisbaenians, and tuatara) origins in the Triassic is largely based on fossils from Europe and South America (e.g., Fraser and Benton, Reference Fraser and Benton1989; Jones et al., Reference Jones, Anderson, Hipsley, Müller, Evans and Schoch2013; Whiteside et al., Reference Whiteside, Duffin and Furrer2017; Simões et al., Reference Simões, Caldwell, Tałanda, Bernardi, Palci, Vernygora, Barnardini, Mancini and Nydam2018; Hsiou et al., Reference Hsiou, Nydam, Simões, Pretto, Onary, Martinelli, Liparini, Romo de Vivar Martinez, Soares, Schultz and Calwell2019; Romo de Vivar et al., Reference Romo de Vivar, Martinelli, Hsiou and Soares2020). The record of this clade in the Upper Triassic rocks of western North America is comparatively incomplete. This is surprising because of the extensive paleontological research on Upper Triassic strata in the Chinle and Dockum basins, and is likely a result of biases towards collection of large-bodied tetrapod taxa such as metoposaurids, aetosaurs, phytosaurs, and dinosaurs (Kligman et al., Reference Kligman, Parker and Marsh2017, Reference Kligman, Marsh and Parker2018; Stocker et al., Reference Stocker, Nesbitt, Kligman, Paluh, Marsh, Blackburn and Parker2019). Triassic-aged lepidosaurs are reported from at least 12 localities in western North America, suggesting they were common elements of these ecosystems, but occurrences are almost entirely comprised of highly fragmentary dentigerous material that severely limits understanding of their diversity, biogeography, and ecology (Murry, Reference Murry1987a, Reference Murryb; Murry and Long, Reference Murry, Long, Lucas and Hunt1989; Kirby, Reference Kirby1991; Kaye and Padian, Reference Kaye, Padian, Fraser and Sues1994; Polcyn et al., Reference Polcyn, Winkler and Jacobs2002; Heckert, Reference Heckert2004; Heckert et al., Reference Heckert, Lucas, Rinehart and Hunt2008; Martz et al., Reference Martz, Mueller, Nesbitt and Stocker2013; Britt et al., Reference Britt, Chure, Engelmann and Shumway2016; Lucas et al., Reference Lucas, Rinehart, Heckert and Hunt2016).
Here we describe Trullidens purgatorii n. gen. n. sp. (DMNH EPV.135990), a sphenodontian rhynchocephalian from Upper Triassic strata of the Purgatoire River valley, southeastern Colorado. Trullidens purgatorii n. gen. n. sp. is the first lepidosaur from the Triassic of western North America with sufficient morphological information to analyze phylogenetic relationships, revealing its inclusion within Opisthodontia (Lepidosauria: Sphenodontia). This is the earliest record of this clade from North America and one of the earliest records globally. The new taxon possesses a masticatory apparatus specialized for herbivory similar to that in other opisthodontians from the Triassic of Europe and South America, including Sphenotitan leyesi Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013 and Fraserosphenodon latidens (Fraser, Reference Fraser1993) (Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013; Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018). The lower jaw bears an elongate ‘incisor-like’ tooth, a morphology unique among lepidosaurs, indicative of a novel ecological function within this clade. The occurrence of this taxon in the Late Triassic of North America demonstrates that opisthodontians were a globally distributed clade of small-bodied herbivores with an unknown biogeographic and evolutionary history prior to the Late Triassic.
The stratigraphic context of Trullidens purgatorii n. gen. n. sp. is in a poorly known sequence of Upper Triassic red beds originally mentioned by Stanton (Reference Stanton1905), and recently re-discovered and re-interpreted (Schumacher, Reference Schumacher2002; Heckert et al., Reference Heckert, Sload, Lucas and Schumacher.2012). The sequence is poorly known due to limited exposure and difficult accessibility. The strata represent a fluvio-lacustrine depositional setting and preserve a fossil biota typical of the Late Triassic of western North America (Heckert et al., Reference Heckert, Sload, Lucas and Schumacher.2012). The geographic isolation of these rocks from other Upper Triassic sequences in western North America renders exact stratigraphic and geochronologic correlation difficult, but limited biochronologic evidence suggests a middle Norian age. The unique fossil biota of these rocks, including Trullidens purgatorii n. gen. n. sp. and early sauropodomorph trackways (McClure et al., Reference McClure, Lockley, Schumacher and Korbitz2021), suggests biogeographic heterogeneity and endemism between Upper Triassic basins in western North America (Small and Martz, Reference Small and Martz2013; Martz and Small, Reference Martz and Small2019).
Geological setting
The specimens discussed herein are from Upper Triassic rocks exposed in a limited area of canyonlands along the main canyon of the Purgatoire River and several tributary canyons in Las Animas County, Colorado (Fig. 1.1). These rocks are mostly red in color, and consist of fluvial sandstones and conglomerates, floodplain mudstones, and paleosols (Fig. 1.2). A distinctive bed of vertebrate bone-bearing, lenticular, limestone-pebble conglomerate is present as the basal unit of the Upper Triassic sequence directly overlying the top of the Jelm Formation (Heckert et al., Reference Heckert, Sload, Lucas and Schumacher.2012). These rocks are exposed as cliffs and benches along the Purgatoire River and its side canyons. The Upper Triassic rocks unconformably overlie aeolianites of the Middle Triassic Red Draw Member of the Jelm Formation and are unconformably overlain by gypsiferous beds of the Middle Jurassic Ralston Creek Formation (de Albuquerque, Reference de Alburqueque1988; Heckert et al., Reference Heckert, Sload, Lucas and Schumacher.2012).
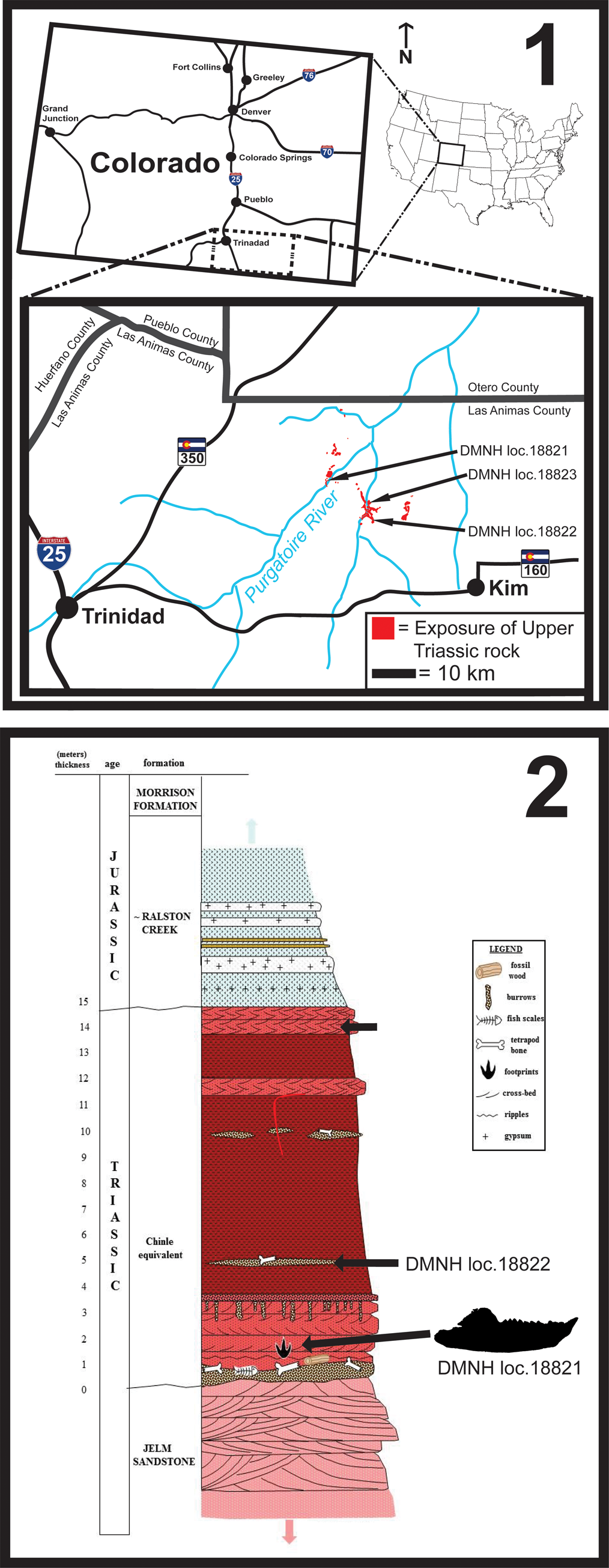
Figure 1. Geographic (1) and stratigraphic (2) locations of the holotype specimen of Trullidens purgatorii n. gen. n. sp. (DMNH EPV.135990; DMNH loc. 18821), and aetosaur osteoderms used for biostratigraphic correlation, including Paratypothorax sp. (DMNH EPV.135991; DMNH loc. 18822) and Aetosauria indet. (DMNH EPV.135992; DMNH loc. 18823).
The Upper Triassic sediments are likely derived from the Ancestral Front Range, which was located to the west in south-central Colorado in the Late Triassic (Dubiel, Reference Dubiel, Caputo, Peterson and Franczyk1994). The Ancestral Front Range may have separated this depositional sequence of Upper Triassic strata exposed in the Purgatoire River canyonlands from the Chinle Formation in Arizona and Utah, as well as other Triassic basins in Western Colorado and Wyoming (e.g., Eagle Basin and Popo Agie Formation; Dubiel, 1994). The presence of abundant conglomerate beds and lenticular sandstone bodies in these strata indicate deposition in a high-energy fluvial setting, possibly suggesting a closer proximity to the Ancestral Front Range source area than the nearest exposures of Upper Triassic rocks in Union County, New Mexico and neighboring Oklahoma, where strata are dominated by finer-grained sediments deposited in a lower-energy setting (Stovall and Savage, Reference Stovall and Savage1939). Heckert et al. (Reference Heckert, Sload, Lucas and Schumacher.2012) proposed that these rocks may be a geographic outlier of the Chinle Formation. Further investigation is needed to develop a clearer understanding of the depositional connection of the Purgatoire canyonlands deposits to the Chinle Formation.
The Upper Triassic rocks of the Purgatoire canyonlands include a diverse fossil biota of invertebrates, plants, and vertebrates. Invertebrate trace fossils attributed to Scoyenia are common on sandstone bedding planes, particularly in the lower half of the strata. Vertebrate ichnofossils include locally abundant vertical burrows, possibly constructed by dipnoans (or other vertebrates), and a vertebrate trackway from a large tetrapod with likely sauropodomorph affinities (McClure et al., Reference McClure, Lockley, Schumacher and Korbitz2021). During field work conducted in 2019, plant macrofossils, paleoniscoid fish, and abundant phytosaur and aetosaur material were collected, further supporting a Late Triassic age for these strata, as reported by Heckert et al. (Reference Heckert, Sload, Lucas and Schumacher.2012).
Two aetosaur osteoderms collected in 2019 allow for biostratigraphic correlation with the Chinle Formation. DMNH EPV.125991 is a right lateral dorsal osteoderm of a paratypothoracine aetosaur (Parker, Reference Parker2007) that we refer to Paratypothorax sp. (Fig. 2.1) based on the following synapomorphies: (1) dorsal and lateral flanges that connect to form a laterally projected horn, (2) presence of radial grooves radiating longitudinally from an acute horn, and (3) a reduced dorsal flange (Long and Ballew, Reference Long and Ballew1985; Hunt and Lucas, Reference Hunt and Lucas1992; Parker, Reference Parker2007). Occurrences of aetosaur fossils referable to Paratypothorax in the Chinle Formation are limited to within the Sonsela Member (‘upper subbiozone’ of the Adamanian holochronozone and the Revueltian holochronozone; Parker and Martz, Reference Parker and Martz2010; Martz and Parker, Reference Martz, Parker, Ziegler and Parker2017). The lowest documented stratigraphic occurrence of Paratypothorax is in the Lot's Wife Beds of the Sonsela Member at the Placerias Quarry (MNA 207-2) where the taxon is represented by lateral and paramedian osteoderms (MNA V3202; Parker, Reference Parker2005). The Placerias Quarry is isotopically dated to 219.317 ± 0.080 Ma (Ramezani et al., Reference Ramezani, Fastovsky and Bowring2014). The highest documented stratigraphic occurrence of Paratypothorax in the Chinle Formation is a partial carapace (PEFO 3004) from the Crystal Forest Buttes locality (PFV 173) in the Martha's Butte Beds of the Sonsela Member (Parker and Martz, Reference Parker and Martz2010), a locality bracketed by isotopic ages of 209.926 ± 0.26 and 212.81 ± 1.25 Ma (Ramezani et al., Reference Ramezani, Hoke, Fastovsky, Bowring, Therrien, Dworkin, Atchley and Nordt2011; Kent et al., Reference Kent, Olsen, Rasmussen, Lepre, Mundil, Irmis, Gehrels, Giesler, Geissman and Parker2018). DMNH EPV.125991 was collected from a lens of limestone pebble conglomerate ~5 m above the base of the Upper Triassic Sequence in the Picket Wire Canyonlands (DMNH loc. 18822; Fig. 1.2).

Figure 2. (1) DMNH EPV.135991, Paratypothorax sp. right lateral osteoderm in ventral view; (2) DMNH EPV.135992, Aetosauria indet., right dorsal paramedian osteoderm in dorsal view. Scale bars = 1 cm.
The second aetosaur specimen, DMNH EPV.135992 (Fig. 2.2), is a right dorsal paramedian osteoderm similar to those of Stenomyti huangae Small and Martz, Reference Small and Martz2013 (DMNH EPV.60708) of the Eagle Basin in Colorado and Aetosaurus ferratus Fraas, Reference Fraas1877 of Germany and Italy, based on the small size of the osteoderm, presence of weak radiating grooves on its dorsal surface, a weak dorsal eminence that does not reach the anterior bar, and a 5:2.7 length to width ratio (Schoch, Reference Schoch2007; Small and Martz, Reference Small and Martz2013). DMNH EPV.135992 was collected from a lens of limestone pebble conglomerate, one meter below the contact of Upper Triassic rocks with the overlying Ralston Creek Formation (DMNH loc. 18823; Fig. 1.2). The occurrence of Stenomyti huangae in the Eagle Basin is biostratigraphically correlated to the Revueltian holochronozone (sensu Martz and Parker, Reference Martz, Parker, Ziegler and Parker2017) of the Chinle Formation based on the presence of the procolophonid Libognathus sheddi Small, Reference Small1997, the aetosaur Rioarribosuchus chamaensis Lucas, Hunt, and Spielmann, Reference Lucas, Hunt and Spielmann2006, and the lagerpetid Dromomeron romeri Irmis et al., Reference Irmis, Nesbitt, Padian, Smith, Turner, Woody and Downs2007 (Small and Martz, Reference Small and Martz2013; Martz and Small, Reference Martz and Small2019). Therefore DMNH EPV.135992 adds a second, though tentative, biostratigraphic link to the Revueltian holochronozone.
Given these biostratigraphic correlations, we assign the vertebrate assemblage of the Upper Triassic strata from the Purgatoire River valley to the upper subzone of the Adamanian holochronozone and the Revueltian holochronozone with geochronologic bounds of ca. 219 to ca. 209 Ma (middle Norian). The basal conglomerates of these strata were tentatively correlated to the Cobert Canyon Bed from the base of the Chinle Formation in far western Oklahoma and northeast New Mexico or the Gartra Formation of Wyoming, which are of uncertain Late Triassic age (Heckert et al., Reference Heckert, Sload, Lucas and Schumacher.2012).
DMNH EPV.135990 (Trullidens purgatorii n. gen. n. sp.) was found embedded in the matrix of a red-brown colored limestone pebble conglomerate bed with sand-sized matrix that was exposed on the banks of the Purgatoire River (DMNH loc. 18821; Fig. 1.2; 37°27'53.67"N, 103°45'43.11W GCS). The sub-rounded imbricated limestone rip-up clasts of up to 7 cm are matrix-supported. Other vertebrate material is common in this horizon, mainly consisting of bone fragments up to 5 cm in length that are too worn or incomplete for identification. A stratigraphically equivalent sandstone bed at the same locality preserves a large sized trackway assigned to a sauropodomorph dinosaur (McClure et al., Reference McClure, Lockley, Schumacher and Korbitz2021).
Material and methods
Field and lab methods
DMNH EPV.135990 was collected by hand and trimmed of excess rock matrix using a battery powered rotary tool with a diamond blade. The specimen was subsequently scanned using x-ray microcomputed microtomography (μCT scanning) at the Duke Shared Materials Instrumentation Facility on a Nikon XTH 225 ST High Resolution X-ray Computed Tomography Scanner using the following parameters: source voltage (kV) = 173, source current (μA) = 81, image pixel size (μm) = 17.42. Digital three-dimensional models were generated in Materialize Mimics 20.0 and imaged in Meshlab 2016.
Phylogenetic methods
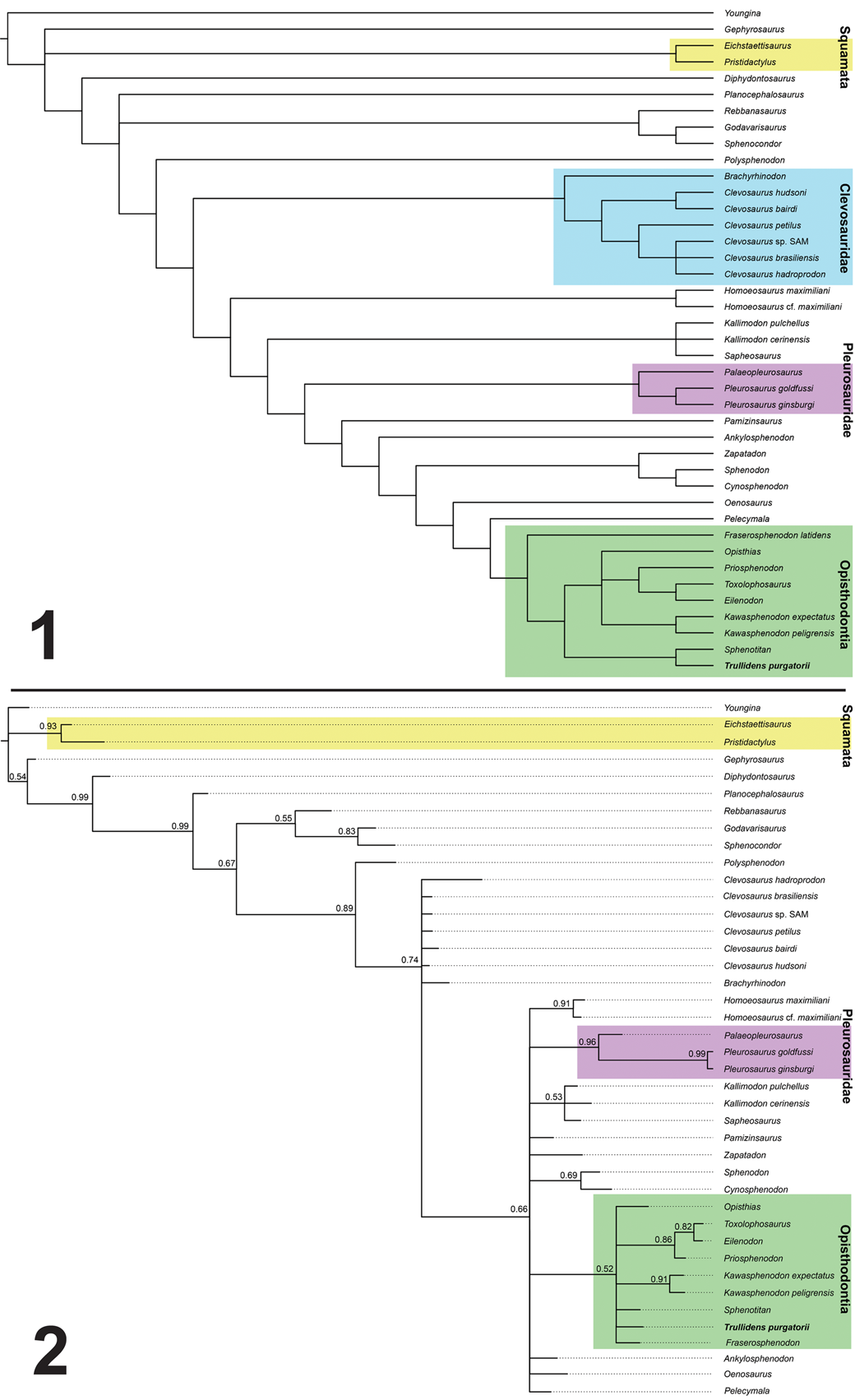
Evolutionary relationships of DMNH EPV.135990 were tested using a recent rhynchocephalian dataset (analysis 2 dataset of Hsiou et al., Reference Hsiou, Nydam, Simões, Pretto, Onary, Martinelli, Liparini, Romo de Vivar Martinez, Soares, Schultz and Calwell2019). We added DMNH EPV.135990 to this dataset and performed a parsimony analysis using TNT v. 1.5 (Goloboff et al., Reference Goloboff, Farris and Nixon2008) under the following parameters: new technology search option with four rounds of tree fusing, 10 rounds of drifting, and 200 ratcheting iterations. Seven resulting most parsimonious trees were found with 253 steps each (consistency index = 0.423; retention index = 0.695). The most parsimonious trees were summarized in a maximum parsimony 50% majority rule consensus tree using traditional tree bisection and reconnection branch swapping. A Bayesian inference analysis was performed in MrBayes 3.2.6 (Ronquist et al., Reference Ronquist, Teslenko, Mark, Ayres, Darling, Höhna, Larget, Liu, Suchard and Huelsenbeck2012) under the Mkv evolutionary model. The analyses used two independent runs with four Markov chains each, sampling every 1000 generations with a total of 10 million generations. The burn in fraction was set to 25% and rate variation across characters was sampled from a gamma distribution. A 50% majority rule consensus tree was produced to summarize this analysis. A time-scaled tree was produced in the package strap (Bell and Lloyd, Reference Bell and Lloyd2015) using the consensus tree topology from the parsimony analysis and the temporal ranges of rhynchocephalians used in this analysis. Temporal range data are derived from the supplementary information of Hsiou et al. (Reference Hsiou, Nydam, Simões, Pretto, Onary, Martinelli, Liparini, Romo de Vivar Martinez, Soares, Schultz and Calwell2019) and incorporates new temporal range data for Trullidens purgatorii n. gen. n. sp., and revisions of temporal range data for Brachyrhinodon taylori Huene, Reference Huene1910. Method summary and data sets used in phylogenetic analyses are available in the supplemental data file.
Anatomical terminology
Terms describing tooth generations in rhynchocephalians (i.e., additional, successional, hatchling) follow the terminology of Robinson (Reference Robinson, Bellairs and Cox1976) and Apesteguía et al. (Reference Apesteguía, Gómez and Rougier2012). Herein, teeth in each of the dental generations are numbered sequentially anterior to posterior (the anterior-most tooth in each generation is tooth one, and numbers increase posteriorly).
Repositories and institutional abbreviations
University of Aberdeen Paleontology Collection (AUP), Aberdeen, UK; Denver Museum of Natural History Vertebrate Paleontology Collection (DMNH EPV), Denver, USA; Field Museum of Natural History (FMNH), Chicago, USA; Museu Municipal Aristides Carlos Rodrigues (MMACR), Rio Grande do Sul Province, Brazil; Museum of Northern Arizona (MNA), Flagstaff, USA; Museo Carlos Ameghino, Cipolletti, (MPCA) Rio Negro Province, Argentina; Petrified Forest National Park (PEFO), Petrified Forest, USA; Division of Paleontology of the Museo de Ciencias Naturales, Universidad Nacional de San Juan (PVSJ), San Juan, Argentina; Virginia Museum of National History (VMNH), Virginia, USA.
Systematic paleontology
Lepidosauria Haeckel, Reference Haeckel1866
Rhynchocephalia Günther, Reference Günther1867
Sphenodontia Williston, Reference Williston1925
Opisthodontia Apesteguía and Novas, Reference Apesteguía and Novas2003
Trullidens new genus
Type species
Trullidens purgatorii n. gen. n. sp. (by monotypy).
Diagnosis
As for type species.
Occurrence
DMNH loc. 18821, a fossiliferous horizon in the Upper Triassic rocks of the Purgatoire River valley, J.E. Canyon Ranch, Las Animas County, Colorado, USA (Fig. 1). Estimated middle Norian age, estimated Revueltian holochronozone (sensu Martz and Parker, Reference Martz, Parker, Ziegler and Parker2017). Detailed locality information is on file from DMNH and available to qualified researchers.
Etymology
Named for the Latin word for trowel, ‘trulla’ for its trowel-like anterior-most mandibular tooth, and the Latin word ‘dens’ for tooth.
Remarks
Trullidens n. gen. is referred to Sphenodontia Williston, Reference Williston1925 based on acrodont implantation of the entire dental series in the hypodigm, where the teeth are attached to the dorsal crest of the dentary, lacking a bony wall on their labial and lingual sides. Unlike the dentition in some parareptiles, which has been described as ‘acrodont,’ the teeth in Trullidens n. gen. and other sphenodontians are not replaced, and instead new teeth are added at the posterior position in the jaw (Jenkins et al., Reference Jenkins, Jones, Zikmund, Boyde and Daza2017). Erection of this new genus is warranted given the distinct characteristics of the current hypodigm, which includes a series of features unique among sphenodontian taxa (detailed below in the species diagnosis section), particularly the elongate anterior-most tooth.
Holotype
DMNH EPV.135990, a complete right dentary.
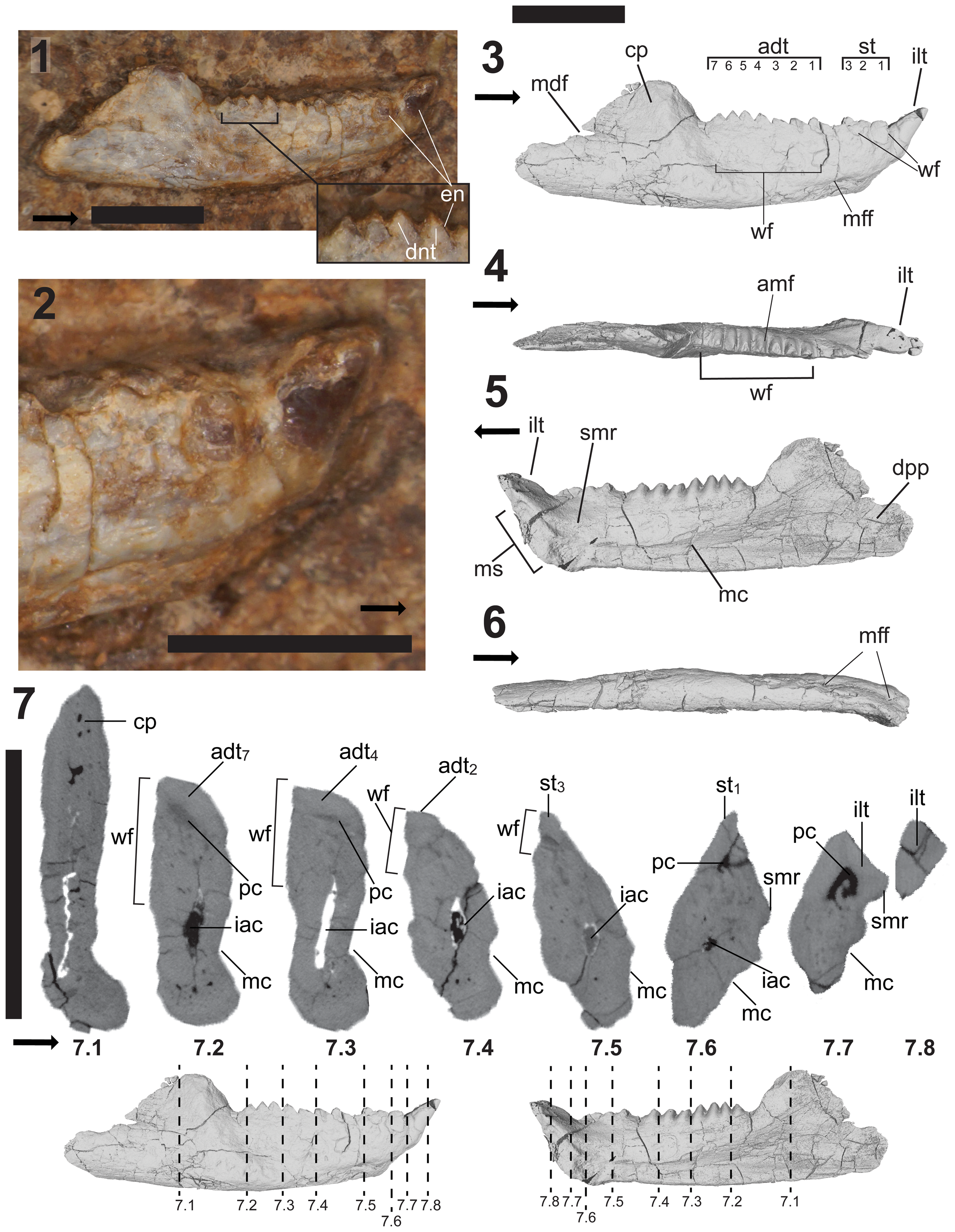
Figure 3. DMNH EPV.135990, holotypic right dentary of Trullidens purgatorii n. gen. n. sp. (1) Labeled photograph of lateral surface of jaw embedded in rock matrix; (2) magnified photograph of the anterior end of the jaw in lateral view; (3–6), labeled images of 3D reconstruction of jaw in; (3) lateral view; (4) dorsal view; (5) medial view; (6) ventral view. Coronal μCT cross sections of DMNH EPV.135990 (7), locations of cross section indicated by dashed lines labeled 7.1–7.8; dashed lines correspond to μCT cross sections with the corresponding label (7.1–7.8). Abbreviations: adt, additional teeth; amf, anteromedial flange; cp, coronoid process; dpp, dentary posterior process; dnt, dentine; en, enamel; iac, inferior alveolar canal; ilt, ‘incisor-like’ tooth; mdf, mandibular foramen; mc; Meckelian canal; mff, mental foramina; ms, mandibular symphysis; pc, pulp cavity; smr, supra-Meckelian ridge; st, successional teeth; wf, wear facet. Numbers indicate tooth position in each dental generation. Arrows indicate anterior direction (1–6) and medial direction (7). Scale bars = 5 mm.
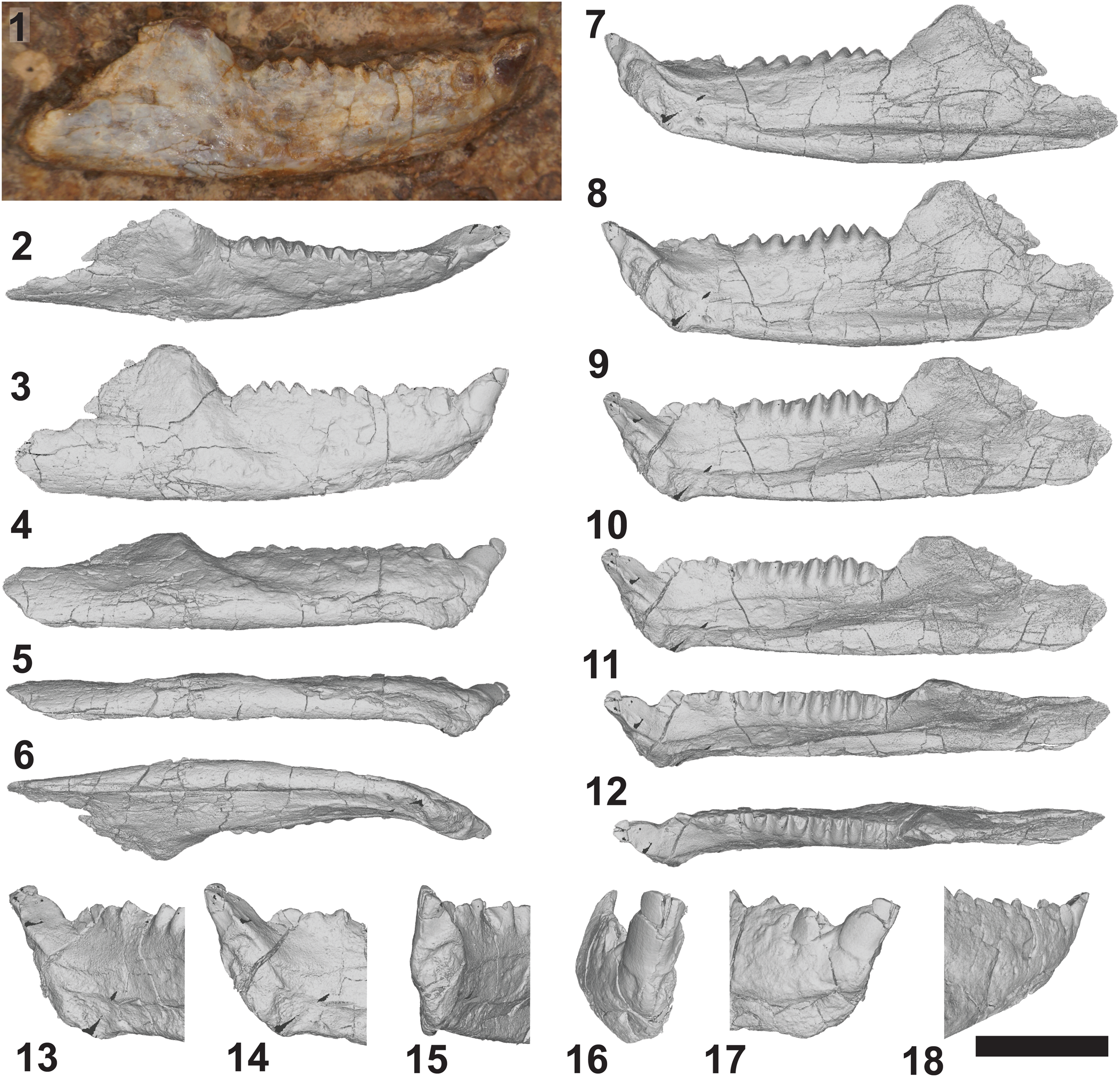
Figure 4. DMNH EPV.135990, holotypic right dentary of Trullidens purgatorii n. gen. n. sp. (1) Photograph of lateral surface of jaw. Images of 3D reconstruction of jaw (2–12), in: dorsolateral (2), lateral (3), ventrolateral (4), ventral (5), ventromedial (6, 7), medial (8), dorsomedial (9–11), and dorsal (12) views. 3D reconstructions of anterior end of the jaw showing ‘incisor-like’ tooth (13–18), in: posteromedial (13), medial (14), anteromedial (15), anterior (16), lateral (17), and posterolateral (18) views. Scale bar = 5 mm.
Diagnosis
Sphenodontian rhynchocephalian with a robust dentary differing from all known sphenodontians in possessing the following unique combination of autapomorphic features in the lower jaw: an antero-dorsally projecting, elongate ‘incisor-like’ tooth extending anteriorly and dorsally beyond the mandibular symphysis; in occlusal view, the long axis of the ‘incisor-like’ tooth is medial to and parallel to the rest of the dentition; a continuous wear surface on the labial side of the dentary from the apex of the additional teeth towards the ventrolateral surface of the dentary, occupying 70% of the dorsoventral depth of the jaw (formed by occlusion with the lingual surface of the maxillary dentition); a U-shaped wear facet on the lateral surface of the second and third successional teeth and a ventrally pointing triangular wear facet on the lateral side of the jaw between the first succession tooth and the ‘incisor-like’ tooth.
Description
Holotype dentary DMNH EPV.135990 is a complete right dentary (Figs. 3, 4) with an anteroposterior length of 1.81 cm. Although DMNH EPV.135990 was found and collected in situ from the outcrop at DMNH Loc. 18821 with its lateral side exposed, there is no apparent damage to the specimen from weathering. Shallow incised concavities on the lateral side of the dentary are similar to wear surfaces found in other rhynchocephalians, and are interpreted to be formed by occlusion of the lingual surface of the maxillary dentition on the labial surface of the dentary during mastication.
Dentary
In mediolateral view, the precoronoid dentary is an elongate anteriorly tapering rectangle with an anterior end inflected anterodorsally at a 45° angle. The jaw is mediolaterally thick with a maximum length-to-width ratio of 0.1. The anterior half of the lower jaw is medially curved towards the symphysis in dorsoventral view. The dorsoventral depth and mediolateral thickness of the dentary are similar to those of other opisthodontians, including Sphenotitan leyesi, Opisthias rarus Gilmore, Reference Gilmore1909, and Fraserosphenodon latidens (Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013; Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018). The anteroposterior length of the jaw is 1.81 cm, shorter than other opisthodontians with complete dentaries such as Opisthias rarus and Priosphenodon avelasi Apesteguía and Novas, Reference Apesteguía and Novas2003 that range from 2–4 cm in length, and much smaller than Sphenotitan leyesi that can reach over 6 cm in length.
In mediolateral view, the coronoid process rises posterodorsally from the dorsal crest of the precoronoid dentary at a 60° angle to an overall dorsoventral height of 140% greater than the height of the posterior-most tooth. The thin bone at the anterodorsal corner of the coronoid process is broken, but the posterodorsal corner is squared off. The dorsoventrally tall, trapezoidal, robust coronoid process differs from the dorsoventrally low, triangular coronoid process in other opisthodontians where this feature is preserved (e.g., Priosphenodon avelasi Apesteguía and Novas, Reference Apesteguía and Novas2003). An anteriorly directed notch in the posterior margin of the coronoid forms the anterior margin of the mandibular foramen (Fig. 3.3). In mediolateral view, the ventral margin of the dentary is almost straight with a slight anterior and posterior parabolic curve. The posterior process of the jaw is short and deep, occupying 15% of the total jaw length. The posterior margin is rounded and mostly complete, and there are no clear facets for the attachment of the postdentary bones.
The Meckelian groove extends from a point ventral to the coronoid process to the mandibular symphysis (Fig. 3.5). It is mediolaterally shallow and dorsoventrally tall, occupying 40% of the dorsoventral depth of the jaw ventral to the posterior-most additional tooth, and tapering anteriorly to a minimum height where it meets the mandibular symphysis where it occupies 12% of the dorsoventral depth of the jaw. As it approaches the symphysis the groove shallows in its mediolateral depth, a similar morphology to that of clevosaurs such as Clevosaurus hudsoni Swinton, Reference Swinton1939 (Fraser, Reference Fraser1988) as well as opisthodontians including Sphenotitan leyesi (PVSJ 887; Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013), Toxolophosaurus cloudi Olson, Reference Olson1960 (Throckmorton et al., Reference Throckmorton, Hopson and Parks1981), and Kaikaifilusaurus calvoi Simón and Kellner, Reference Simón and Kellner2003. The mandibular symphysis points anteromedially and rises slightly from the surrounding bone surface to form an oval shape with its long axis oriented tilted anterodorsally at a 45° angle. It occupies 83% of the dorsoventral depth of the anterior surface of the jaw (excluding the height of the ‘incisor-like’ tooth). The mandibular symphysis is dorsoventrally tall as in other opisthodontians and forms an extensive surface (e.g., Sphenotitan leyesi, Kaikaifilusaurus calvoi). On the medial surface of the dentary between the ventral margin of the teeth and the dorsal margin of the Meckelian canal is a ridge that is subparallel with the Meckelian canal, extending from the base of the 7th additional tooth to the mandibular symphysis (‘supra-Meckelian ridge’ in Fig. 3.5). A similar ridge is present in the dentary of Sphenotitan leyesi (PVSJ 887; Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013).
The lateral side of the dentary is marked by an extensive wear facet formed by occlusion from the lingual surface of the maxillary dentition (Fig. 3.3, 3.4, 3.7). It occupies 63% of the length of the dentigerous region of the jaw, from the posterior-most additional tooth to the anterior-most additional tooth. This wear surface extends down 70% of the dorsoventral depth of the jaw at its deepest ventral to the 5th additional tooth. The presence of a similar extensive wear facet on the labial side of the dentary teeth is shared with Pelecymala robustus Fraser, Reference Fraser1986, several opisthodontians, including Fraserosphenodon latidens (Fraser, Reference Fraser1993; Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018), and Sphenotitan leyesi (Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013). Two small wear facets occupy the lateral surface of the dentary below the successional teeth. A ventrally pointed, U-shaped wear facet occupies the labial side of the second and third successional teeth and the bone just below it (Fig. 3.3). A ventrally pointed triangular wear facet occupies the mesiolabial side of the first successional tooth and the dentary bone between that tooth and the ‘incisor-like’ tooth (Fig. 3.3). Two mental foramina are present on the ventrolateral side of the jaw, ventral to the first successional tooth and the gap between the successional and additional teeth (Figs. 3.3, 3.6).
Dentition
All teeth have acrodont implantation, and 11 teeth occupy the dentigerous region of the jaw. Three dental generations of teeth are present in anterior to posterior sequence (sensu Robinson, Reference Robinson, Bellairs and Cox1976): a single large ‘incisor-like’ tooth at the anterior end of the jaw, three successional teeth just posterior to the ‘incisor-like’ tooth, and seven labiolingually broadened additional teeth with extensive labial wear.
The ‘incisor-like’ tooth is wedge-shaped with a flat dorsal surface with sharp edges where it meets the rounded surface formed by the medial, ventral, and lateral sides of the tooth (Figs. 3.1–3.4, 3.7.8, 4.13–4.18). It is the only tooth in the jaw that lacks wear on its labial side, and therefore is the only tooth with extensive enamel on its labial side. Although mostly obscured by rock matrix, the apical surface of this tooth is enamel-less, exposing a flat dentine surface that meets the enamel-covered labial side at an acute angle (Figs. 3.2, 4.13–4.18). The lingual side of this tooth is a flat-planar surface forming a dorsal extension of the symphyseal surface, such that left and right ‘incisor-like’ teeth would form a single chisel-like spur in life. It is unclear whether enamel is present on the lingual side of this tooth because it is covered in rock matrix, and the μCT data lacks resolution to differentiate enamel from dentine (Fig. 3.7.7, 3.7.8), but the aforementioned symphyseal structure of the lingual surface may indicate the absence of enamel on this surface. When included in the dorsoventral depth of the anterior end of the jaw, this tooth is 45% of the total depth and projects anterodorsally beyond the end of the dentary bone. The ‘incisor-like’ morphology of this tooth is unique among lepidosaurs, with exception of the Norian-aged sphenodontian Clevosaurus hadroprodon Hsiou et al., Reference Hsiou, Nydam, Simões, Pretto, Onary, Martinelli, Liparini, Romo de Vivar Martinez, Soares, Schultz and Calwell2019 (MMACR PV-027-T), which bears an anterodorsally projecting tooth at the anterior-most position in the dentary. In Clevosaurus hadroprodon this tooth is less elongate than in Trullidens purgatorii n. gen. n. sp. Opisthodontian sphenodontians bear an anterodorsally projecting mandibular spur, exemplified in Sphenotitan leyesi (PVSJ 887), Priosphenodon avelasi, (MPCA 300; Apesteguía and Novas, Reference Apesteguía and Novas2003), and Toxolophosaurus cloudi (FMNH UR 619; Throckmorton et al., Reference Throckmorton, Hopson and Parks1981), although this spur is formed by edentulous bone of the dentary and not an enamel-bearing tooth. Within Rhynchocephalia, successional teeth in the anterior of the lower jaw that are substantially larger than adjacent teeth are referred to as ‘caniniform’ (Apesteguía et al., Reference Apesteguía, Gómez and Rougier2012) and are present in many taxa, including Theretairus antiquus Simpson, Reference Simpson1926, Sphenovipera jimmysjoyi Reynoso, Reference Reynoso2005, Sphenocondor gracilis Apesteguía, Gómez, and Rougier, Reference Apesteguía, Gómez and Rougier2012, and Sphenodon punctatus (Gray, Reference Gray1842) (Jones, Reference Jones2009). The anterior-most tooth in Trullidens purgatorii n. gen. n. sp. and Clevosaurus hadroprodon differs substantially from the aforementioned taxa with true caniniform teeth because of its anterior-most position, the robust chisel-like shape, and its anterodorsally projection beyond the anterior margin of the dentary.
Posterior to the ‘incisor-like’ tooth are three labiolingually compressed successional teeth with a triangular shape and rounded apices in lateral view. All three teeth are closely spaced with no gaps between them (Fig. 3.3). The apical margin of each tooth forms a continuous acute ridge extending from the anterior-most to posterior-most tooth. The first successional tooth has a small amount of enamel on its labial side (Fig. 3.2), but the enamel is not present where the triangular wear facet cuts the anterior surface of the tooth. This tooth resembles the second anterior-most tooth in the dentary of Fraserosphenodon latidens (AUP 1192; Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018). The second and third successional teeth are worn on their labial sides by the U-shaped wear facet (Fig. 3.3, 3.7.5).
No hatchling teeth are present between the successional and additional generations, suggesting a later stage in ontogeny for this individual. A gap separates the third successional tooth from the first additional tooth. The additional teeth all share a similar morphology, forming labiolingually widened rectangular outlines in occlusal view (Fig. 3.4). The labiolingual width of these teeth reduces anteriorly and posteriorly, with the third tooth being the widest. The teeth are closely spaced with no gaps between teeth. The teeth have anteromedially oriented flanges extending down the medial surface of the jaw (Fig. 3.4). The overall shape of each tooth is a loaf with a rounded apex that becomes more acute in the labial direction. Enamel covers the surface of each tooth except for the labial side. Wear from the lingual surface of the maxillary dentition has produced a vertical wear facet and exposes cross sections of the internal tissues of the teeth showing enamel and dentine layers (Fig. 3.1). The shape and size of these teeth resemble the posterior dentition of Fraserosphenodon latidens (Fraser, Reference Fraser1993; Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018), and Sphenotitan leyesi (Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013).
The resolution of the μCT scans prohibits differentiation of dental and mandibular tissue types (e.g., enamel, dentine, attachment tissue), but some features of the internal structure are apparent (Fig. 3.7). A sheet-like vascular bed (pulp cavity) floors the base of each additional tooth (‘pulp cavity’ in Fig. 3.7.2, 3.7.3). This resembles the structure of the sheet-like vascular beds in the eilenodontine opisthodontian Priosphenodon avelasi (LeBlanc et al., Reference LeBlanc, Apesteguía, Larsson and Caldwell2020), and in both Trullidens purgatorii n. gen. n. sp. and Priosphenodon avelasi these vascular beds progressively decrease in size anteriorly and are absent in the oldest anterior-most additional teeth (Fig. 3.7.4). In lateral view, the wear surface on the labial side of the additional tooth series exposes the internal dental structure (Fig. 3.1), revealing two features: (1) the enamel layer is ~0.2 mm thick; and (2) the distal side of each tooth overlaps the basal 1/2–1/3 of the posteriorly adjacent tooth (shown best in additional teeth four and five, Fig. 3.1). In Priosphendon avelasi, the enamel layer on the additional teeth reaches a thickness of 0.6 mm (LeBlanc et al., Reference LeBlanc, Apesteguía, Larsson and Caldwell2020), significantly thicker than the 0.2 mm thickness in Trullidens purgatorii n. gen. n. sp., however the dentary of Priosphendon avelasi is about twice the size in Trullidens purgatorii n. gen. n. sp. The additional teeth of Priosphendon avelasi and Trullidens purgatorii n. gen. n. sp. overlap and anchor to the enamel surface of the immediately adjacent posterior tooth, which is a feature also apparent in Fraserosphenodon latidens and Pelecymala robustus (Fraser, Reference Fraser1986; Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018).
Etymology
Named ‘purgatorii’ for the type locality on the bank of the Purgatoire River.
Remarks
The taphonomic setting of the bone-bearing unit (DMNH loc. 18821) that produced the holotype of Trullidens purgatorii n. gen. n. sp. (DMNH EPV.135990) makes discovery of additional material referable to this taxon unlikely. All vertebrate bone found in the limestone pebble conglomerate at DMNH loc. 18821 is highly worn, with the exception of DMNH EPV.135990. Although more material of this taxon is unlikely to be found at the holotypic locality, its presence indicates the future possibility of finding additional opisthodontian sphenodontian material from other Upper Triassic strata in western North America.
Results
Phylogenetic analyses
Both parsimony and Bayesian-inference phylogenetic analyses recover Trullidens purgatorii n. gen. n. sp. within the stem-based definition of Opisthodontia, lending strong support for its inclusion within this clade (Fig. 5.1, 5.2). All most-parsimonious trees (MPT's) recovered Trullidens purgatorii n. gen. n. sp. in a sister group relationship with Sphenotitan leyesi (Fig. 5.1), with the presence of a small mandibular foramen as a synapomorphy of this clade (Character 37.0). All MPTs recovered Oenosaurus muehlheimensis Rauhut et al., Reference Rauhut, Heyng, López-Arbarello and Hecker2012 as the earliest-diverging opisthodontian, with the clade Oenosaurus muehlheimensis + Opisthodontia sharing the following synapomorphies: an antorbital region length relative to skull length of one fourth or less (Character 1.2), a short central region between the two pterygoid rami (Character 25.0), and a well-developed and rounded Mentonian process of the mandibular symphysis (Character 33.3). The clade Pelecymala robustus + Opisthodontia (excluding Oenosaurus muehlheimensis) share marginal teeth with rectangular (wider than long) cross sections as a synapomorphy (Character 47.3). The clade Fraserosphenodon latidens + Opisthodontia (excluding Pelecymala robustus and Oenosaurus muehlheimensis) share the following synapomorphies: a maxilla excluded from the margin of the external naris by the posterodorsal process of the premaxilla (Character 7.1), and anteromedial flanges present in at least one mandibular tooth (Character 57.1). In all MPTs, Trullidens purgatorii n. gen. n. sp. + Sphenotitan leyesi are recovered in a sister relationship to the clade of derived opisthodontians including Opisthias rarus, Kawasphenodon peligrensis Apesteguía, Gómez, and Rougier, Reference Apesteguía, Gómez and Rougier2014, Kawasphenodon expectatus Apesteguía, Reference Apesteguía2005, Priosphenodon avelasi, Eilenodon robustus, and Toxolophosaurus cloudi, sharing the following synapomorphies: a well-developed, anterodorsally projected symphyseal spur (Character 36.1), and extensive posteromedial flanges on posterior maxillary teeth (Character 50.2). In the consensus tree the polytomy including Opisthias rarus, Kawasphenodon peligrensis, Kawasphenodon expectatus, Priosphenodon avelasi, Eilenodon robustus, and Toxolophosaurus cloudi shares enamel ornamentation of a few striae and wide grooves on additional mandibular teeth (Character 58.2).

Figure 5. Phylogenetic analyses supporting the position of Trullidens purgatorii n. gen. n. sp. within Opisthodontia using two optimality criteria. (1) 50% majority rule consensus tree, CI = 0.423, RI = 0.695; (2) 50% majority rule consensus tree from trees produced by Bayesian-inference analysis (clade credibility values in decimal proportions labeled on nodes).
In the Bayesian analysis, Opisthodontia was recovered as a polytomy including Opisthias rarus, Sphenotitan leyesi, Trullidens purgatorii n. gen. n. sp., Fraserosphenodon latidens, the sister taxa Kawasphenodon expectatus and Kawasphenodon peligrensis, and a clade including Priosphenodon avelasi, Eilenodon robustus, and Toxolophosaurus cloudi. Fraserosphenodon latidens was recovered within a monophyletic Opisthodontia, strengthening support for its inclusion within this clade (previous Bayesian analyses recovered Fraserosphenodon latidens outside Opisthodontia; Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018).
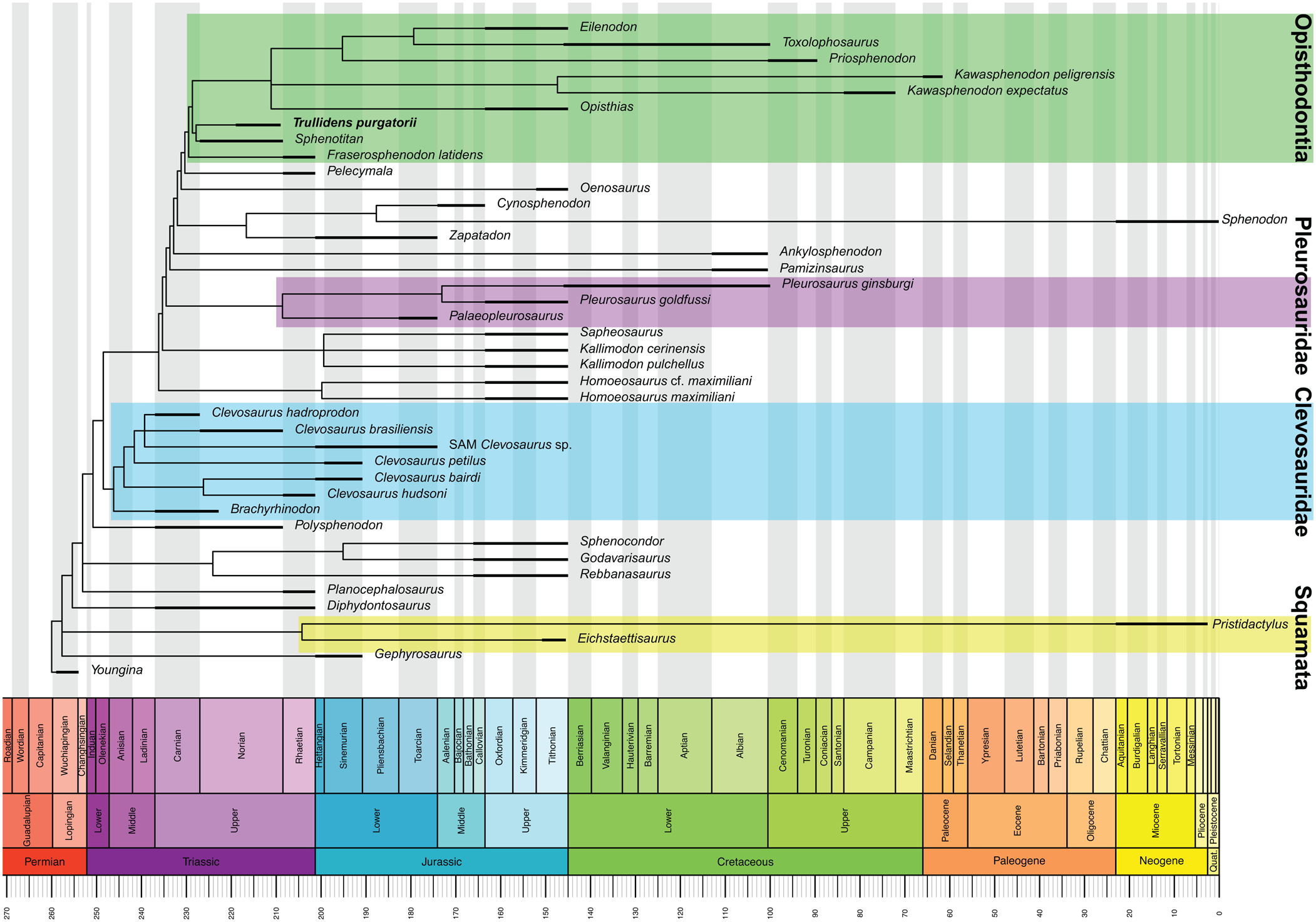
The timing of rhynchocephalian evolution was assessed using the parsimony analysis tree topology and taxon temporal range data to produce a time-scaled phylogeny of Rhynchocephalia (Fig. 6). This analysis demonstrates the presence of two opisthodontian taxa (Trullidens purgatorii n. gen. n. sp. and Sphenotitan leyesi) by the Norian and estimates the origin of Opisthodontian rhynchocephalians to have occurred sometime prior to the Norian.

Figure 6. Time-scaled 50% majority rule consensus tree of Rhynchocephalia (tree from parsimony analysis); thickened lines represent temporal range of each taxon.
Discussion
Evolutionary implications
Our findings suggest that in the Triassic opisthodontian sphenodontians were one of the first monophyletic lepidosaur sub-clades united by shared derived craniodental morphologies to achieve taxonomic diversity and a wide geographic distribution. Furthermore, opisthodontians appear to be exclusively herbivorous, and their Triassic representatives indicate the earliest radiation of any herbivorous lepidosaur clade. The only other clade of lepidosaurs showing a similar evolutionary pattern are clevosaurs, which are represented by six distinct species and several indeterminate occurrences in the Triassic (Table 1). The earliest clevosaurid is Clevosaurus hadroprodon from the Carnian of Gondwana (Hsiou et al., Reference Hsiou, Nydam, Simões, Pretto, Onary, Martinelli, Liparini, Romo de Vivar Martinez, Soares, Schultz and Calwell2019). Norian-aged clevosaurs include Clevosaurus brasiliensis Bonaparte and Sues, Reference Bonaparte and Sues2006 from Gondwana, and a possible occurrence in Equatorial Pangaea (Martz et al., Reference Martz, Mueller, Nesbitt and Stocker2013). Rhaetian-aged clevosaurs occur exclusively in high northern latitudes of Pangaea (Europe), and include Clevosaurus hudsoni, Clevosaurus sectumsemper Klein et al., Reference Klein, Whiteside, De Lucas, Viegas and Benton2015, Clevosaurus minor Fraser, Reference Fraser1988, and Clevosaurus cambrica Keeble, Whiteside, and Benton, Reference Keeble, Whiteside and Benton2018 (Fraser, Reference Fraser1988; Klein et al., Reference Klein, Whiteside, De Lucas, Viegas and Benton2015; Keeble et al., Reference Keeble, Whiteside and Benton2018). Broad evolutionary trends shared by Triassic opisthodontians and clevosaurids include: (1) Strong support for monophyly of each clade respectively, based on suites of derived craniodental features; (2) earliest records from the Carnian to middle Norian; and (3) wide distribution across Pangaea, north and south of the paleoequator by the Late Triassic. Clevosaurs and opisthodontians are characterized by relatively robust cranial elements and jaws compared to other Triassic rhynchocephalians with more delicate skeletons such as Diphydontosaurus avonis Whiteside, Reference Whiteside1986 or Lanceirosphenodon ferigoloi Romo de Vivar et al., Reference Romo de Vivar, Martinelli, Hsiou and Soares2020, and taphonomic or collection biases may account for the more complete Triassic record of opisthodontians and clevosaurids.
Table 1. Table of Triassic-aged rhynchocephalian occurrences organized by continent and from oldest to youngest, respectively, within continental groups. Occurrence nomens are displayed geographically in Figure 8.

*occurrence nomens (e.g., NA1, NA2, etc…) correspond to map in Figure 8
Comparison of the dental tissues in Trullidens purgatorii n. gen. n. sp. to that of Priosphenodon avelasi shows that features including the sheet-like vascular beds and the overlapping attachment of adjacent additional tooth bases have a deep evolutionary history dating to at least the middle Norian within opisthodontians. All opisthodontians except Fraserosphenodon latidens share a well-developed, anterodorsally projected symphyseal spur. In Trullidens purgatorii n. gen. n. sp., the symphysis bears an ‘incisor-like’ tooth that resembles the morphology of the symphyseal spur in other opisthodontians, but in Trullidens purgatorii n. gen. n. sp. it is hyperextended and enamel-covered on its labial and mesial surfaces. A tooth-bearing symphyseal spur may represent the plesiomorphic condition, where the tooth was lost in later-diverging opisthodontians, or it could also represent a derived state autapomorphic to Trullidens purgatorii n. gen. n. sp.
Functional morphology and paleoecology
Opisthodontians are the earliest lepidosaur clade to acquire specialized craniodental features for herbivory, a feeding ecology commonly seen in extant lizards. Therefore, the functional origins of herbivory in Lepidosauria dates back to the Late Triassic, when opisthodontians diversified and distributed. The broad distribution of opisthodontians by the Norian suggests that they may have played important roles in terrestrial ecosystems globally as small to medium body sized herbivores. Several features of the lower jaw and teeth shared in Triassic opisthodontians indicate herbivorous functions including: (1) additional teeth with rectangular tooth bases expanded along the labiolingual axis; (2) truncation of the tooth crowns on their labial margin by a wear facet (created by shearing occlusion with the lingual surface of the maxilla), exposing in cross section the enamel and dentin tissues; (3) continuation of the aforementioned dental wear facet onto the lateral side of the dentary, forming an extensive wear surface; (4) a robust dentary that is dorsoventrally deep and mediolaterally thickened; and (5) a thickened enamel layer on additional teeth (Throckmorton et al., Reference Throckmorton, Hopson and Parks1981; Jones, Reference Jones2008, Reference Jones2009; Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013). Teeth with expanded tooth bases, high surface area, and thickened enamel can withstand greater forces during mastication without breaking while concurrently providing a large surface area for food processing (Lucas and Luke, Reference Lucas, Luke, Chivers, Wood and Bilsborough1984; Jones, Reference Jones2008, Reference Jones2009). Extensive wear facet structures point to a shearing action during occlusion, and the robust jaw structure indicates increased muscle attachment for a strengthened bite force (Throckmorton et al., Reference Throckmorton, Hopson and Parks1981; Jones, Reference Jones2008, Reference Jones2009).
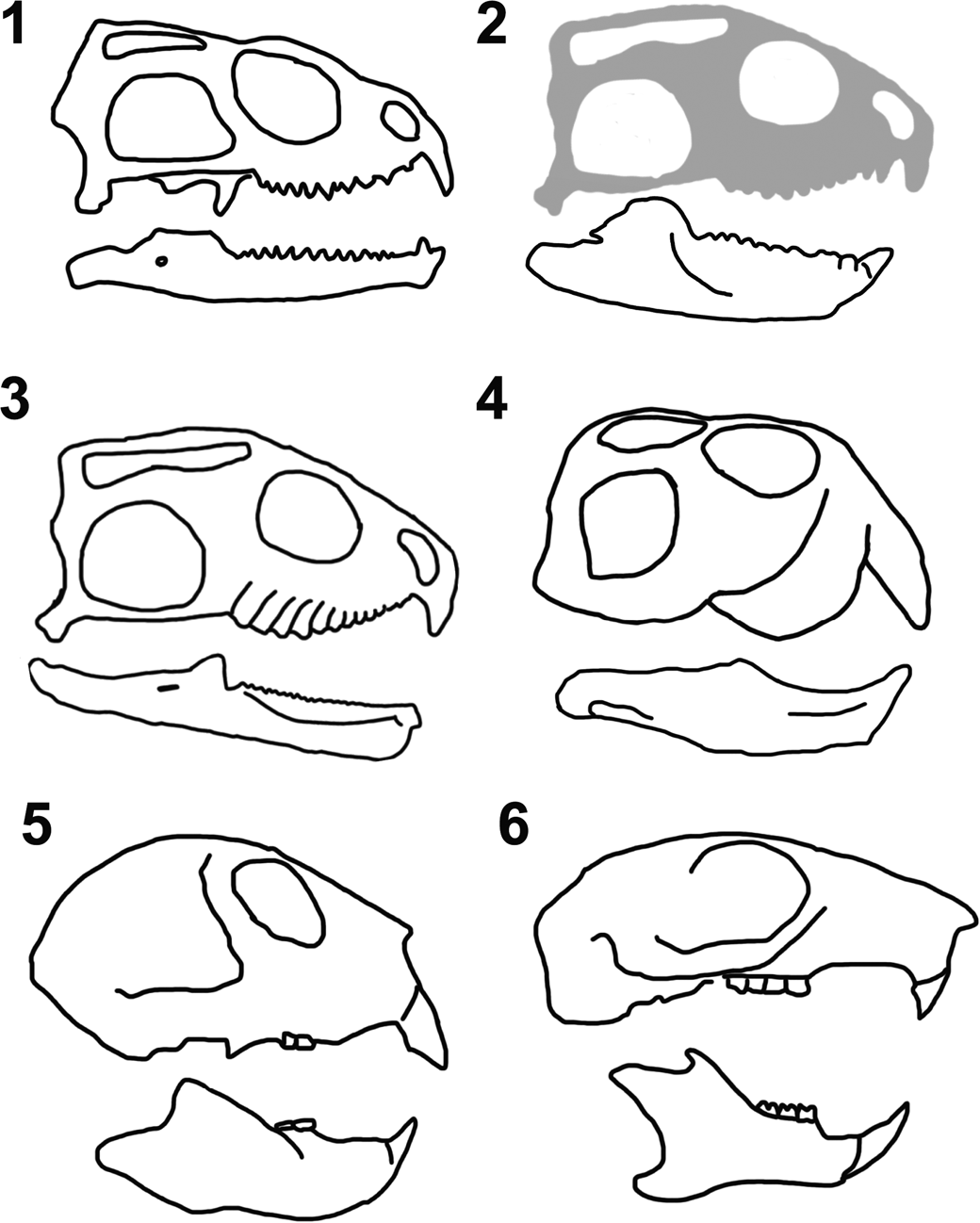
The ‘incisor-like’ tooth in the dentary of Trullidens purgatorii n. gen. n. sp. is unique among lepidosaurs, and comparisons must be made to non-lepidosaur amniotes with similar morphologies to infer its ecological function. Among lepidosaurs, caniniform teeth are a common feature in some taxa including Sphenodon punctatus (Fig. 7.1). But only Trullidens purgatorii n. gen. n. sp. (Fig. 7.2) possesses a hyperextended, enamel-bearing, ‘incisor-like’ tooth in the anterior-most tooth position of the dentary. Sphenotitan leyesi (Fig. 7.3) and Trullidens purgatorii n. gen. n. sp. share deep, robust lower jaws, but Sphenotitan leyesi lacks an ‘incisor-like’ tooth. The only other diapsids with deep, robust lower jaws and an anterodorsal projection of the anterior-most dentary are rhynchosaurs (Archosauromorpha), where an ‘incisor-like’ structure is formed by dense dentary bone (Fig. 7.4). Rhynchosaurs are widely regarded as herbivorous, with a precision-shear bite, and grinding action between their teeth (e.g., Benton, Reference Benton1983, Reference Benton1984). This suite of features is also seen in some mammals (Apesteguía, Reference Apesteguía2008; Apesteguía and Carballido, Reference Apesteguía and Carballido2014), including a robust and deep lower jaw with a tall coronoid process, expanded posterior teeth, and an anteriodorsally projected ‘incisor-like’ tooth in the lower jaw (Fig. 7.5, 7.6). In extant mammals, these features are often related to herbivory, as in rodents such as Sciurus carolinensis Gmelin, Reference Gmelin1788 (Fig. 7.6), but are not exclusive to herbivores and are present in the insectivorous Daubentonia madagascariensis (Gmelin, Reference Gmelin1788) (Fig. 7.5). Although Daubentonia madagascariensis is insectivorous, its incisors are used for biting into wood, exposing the insects inside, which are then extracted with their elongate third manual digit (Petter, Reference Petter, Ranier and Bourne1977). The structure of the ‘incisor-like’ tooth strongly resembles the ‘nipper-facet’ mode of incisor wear seen in rodents, lagomorphs, and Daubentonia madagascariensis (Koenigswald, Reference Koenigswald2018). Nipper-facets are formed on the paired lower incisors in the aforementioned mammals, where enamel is worn off the apical surface of the teeth leaving a dentine field, and enamel is only present on the labial and mesial surfaces. The apical wear surface is formed via abrasion with the upper incisors. Nipper-faceted incisors in lagomorphs and rodents are used for gnawing on various botanical substrates (Koenigswald, Reference Koenigswald2018). The morphological convergence of the ‘incisor-like’ tooth in Trullidens purgatorii n. gen. n. sp. to the nipper-faceted incisors in these mammals indicates its function in Trullidens purgatorii n. gen. n. sp. was for gnawing, and its ecological role was likely as a small-bodied herbivore with a specializing craniodental apparatus for gnawing plant material.

Figure 7. Comparative skull osteology of select amniotes. (1) Sphenodon punctatus (Gray, Reference Gray1842); (2) Trullidens purgatorii n. gen. n. sp. (cranium based on Sphenotitan leyesi Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013); (3) Sphenotitan leyesi (modified from Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013); (4) Teyumbaita sulcognathus Montefeltro, Langer, and Schultz, Reference Montefeltro, Langer and Schultz2010 (modified from Montefeltro et al., Reference Montefeltro, Langer and Schultz2010); (5) Daubentonia madagascariensis (Gmelin, Reference Gmelin1788; (6) Sciurus carolinensis Gmelin, Reference Gmelin1788 (5 and 6 modified from Morris et al., Reference Morris, Cobb and Cox2018).
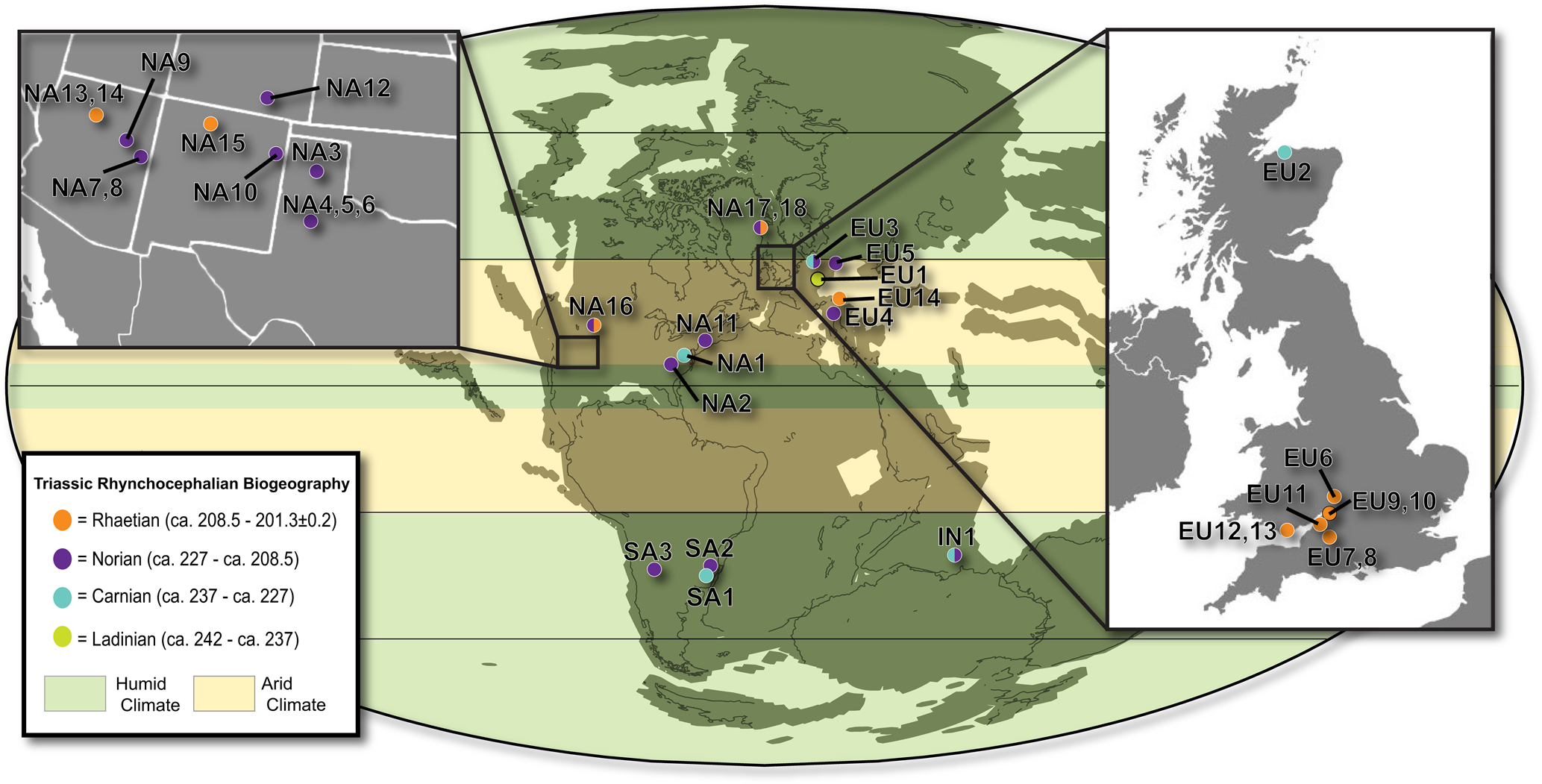
Biogeography
To analyze the biogeographic context of Trullidens purgatorii n. gen. n. sp., we assembled a novel dataset of all reported Triassic-aged rhynchocephalian occurrences (Table 1), visualized in Figure 8. The record of Triassic-aged rhynchocephalians from western North America is characterized by many occurrences of highly fragmentary specimens. Named taxa include Paleollanosaurus fraseri Heckert, Reference Heckert2004 and Planocephalosaurus lucasi Heckert, Reference Heckert2004 from the Tecovas Formation (Norian) in Kalgary County Texas (Heckert, Reference Heckert2004), and Whitakersaurus bermani Heckert et al., Reference Heckert, Lucas, Rinehart and Hunt2008 from the Chinle Formation (Rhaetian) in Rio Arriba County, New Mexico (Heckert et al., Reference Heckert, Lucas, Rinehart and Hunt2008). The evolutionary relationships of Paleollanosaurus fraseri, Planocephalosaurus lucasi, and Whitakersaurus bermani have yet to be analyzed in a phylogenetic analysis, but they resemble the early-diverging sphenodontian taxa Diphydontosaurus avonis and Planocephalosaurus robinsonae Fraser, Reference Fraser1982 from the Late Triassic of Europe due to the presence of dentary teeth with acrodont implantation posteriorly and pleurodont implantation anteriorly (e.g., Fraser, Reference Fraser1982; Whiteside, Reference Whiteside1986). Other records of rhynchocephalians from the Triassic of western North America are reported from Arizona, New Mexico, and Texas, but are too fragmentary to establish relationships beyond Rhynchocephalia (Murry Reference Murry1987a, Reference Murryb; Murry and Long, Reference Murry, Long, Lucas and Hunt1989; Kirby, Reference Kirby1991; Kaye and Padian, Reference Kaye, Padian, Fraser and Sues1994; Polcyn et al., Reference Polcyn, Winkler and Jacobs2002; Heckert, Reference Heckert2004; Martz et al., Reference Martz, Mueller, Nesbitt and Stocker2013; Britt et al., Reference Britt, Chure, Engelmann and Shumway2016; Lucas et al., Reference Lucas, Rinehart, Heckert and Hunt2016). An acrodont jaw fragment (MNA V3684; Kaye and Padian, Reference Kaye, Padian, Fraser and Sues1994) from the Placerias Quarry (MNA 207-2) was referred to the opisthodontian Clevosaurus latidens (renamed Fraserosphenodon latidens sensu Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018) by Fraser (Reference Fraser1993), but a taxonomic reassessment of this and other similar fragmentary jaws from the lower Chinle Formation places them in the enigmatic diapsid genus Palacrodon (Kligman et al., Reference Kligman, Parker and Marsh2017). The occurrence of Trullidens purgatorii n. gen. n. sp. indicates that rhynchocephalians were a common element of Late Triassic ecosystems in western North America, but prior to this study the fragmentary record obscured the importance of this region to the Triassic rhynchocephalian radiation. The discovery of Trullidens purgatorii n. gen. n. sp. is the first Triassic record of an opisthodontian rhynchocephalian from North America. Therefore, the Late Triassic rhynchocephalian biota of western North America includes both early diverging taxa, including Paleollanosaurus fraseri, Planocephalosaurus fraseri, and Whitakersaurus bermani, as well the well-nested taxon Trullidens purgatorii n. gen. n. sp. This assemblage is broadly similar to Late Triassic rhynchocephalian biotas from Europe and South America where assemblages include both early diverging and well-nested rhynchocephalian taxa, demonstrating the biogeographic and phylogenetic diversification of this clade by the Late Triassic in these continents (Table 1).

Figure 8. Paleogeographic map showing reported occurrences of rhynchocephalians during the Triassic. Locality, stratigraphic, and taxonomic information for each occurrence is included in Table 1. Paleoclimate data from Whiteside et al., Reference Whiteside, Grogan, Olsen and Kent2011. Paleogeographic map modified from a map produced using the online paleogeographic map generator (Alroy, Reference Alroy2013).
Trullidens purgatorii n. gen. n. sp. is one of three known Triassic-aged opisthodontian rhynchocephalians, revealing the wide geographic distribution of this clade by the Late Triassic. Sphenotitan leyesi, an opisthodontian notable for its large body size, is known from the Quebrada del Barro Formation (Norian) of Argentina (Martinez et al., Reference Martínez, Apaldetti, Colombi, Praderio, Fernandez, Malnis, Correa, Abelin and Alcober2013). Fraserosphenodon latidens is an opisthodontian known from fragmentary dentigerous material from the Rhaetian-aged Cromhall Quarry in England (Herrera-Flores et al., Reference Herrera-Flores, Stubbs, Elsler and Benton.2018). Trullidens purgatorii n. gen. n. sp. is the first opisthodontian reported from the Triassic of North America, demonstrating the widespread distribution of opisthodontians across Pangaea by the Late Triassic, with a range spanning the paleo-equator from high latitude southern Gondwana (Sphenotitan leyesi) to low latitude equatorial Pangaea (Trullidens purgatorii n. gen. n. sp.) to high latitude Laurasia (Fraserosphenodon latidens). This biogeographic pattern indicates that this clade either distributed across latitudinally arranged climate bands characteristic of the Late Triassic (Whiteside et al., Reference Whiteside, Grogan, Olsen and Kent2011; Kent et al., Reference Kent, Santi, Colombi, Alcober and Martínez2014) or had established their range earlier in the Triassic, prior to the aforementioned Late Triassic climate regime.
The post-Triassic biogeographic distribution of opisthodontians in North America includes two Jurassic occurrences, Opisthias rarus (Kimmeridgian–Tithonian, Wyoming, USA; Gilmore, Reference Gilmore1909), and Eilenodon robustus (Kimmeridgian–Tithonian, Colorado, USA; Rasmussen and Callison, Reference Rasmussen and Callison1981), as well as one from the Cretaceous, Toxolophosaurus cloudi (Aptian–Albian, Montana, USA; Throckmorton et al., Reference Throckmorton, Hopson and Parks1981). In South America, there are four Cretaceous taxa, Priosphenodon minimus Apesteguía and Carballido, Reference Apesteguía and Carballido2014 (Albian, Argentina; Apesteguía and Carballido, Reference Apesteguía and Carballido2014), Priosphenodon avelasi (Cenomanian–Turonian, Argentina; Apesteguía and Novas, Reference Apesteguía and Novas2003), Kaikaifilusarus calvoi (Cenomanian, Argentina; Simón and Kellner, Reference Simón and Kellner2003), Kawasphenodon expectatus (Campanian, Argentina; Apesteguía, Reference Apesteguía2005), and Kawasphenodon peligrensis (Paleogene, Argentina; Apesteguía, et al., Reference Apesteguía and Carballido2014). The distribution of this clade in the Triassic suggests that they became established in North and South America during the Late Triassic, where they persisted into the Cretaceous and Paleogene. Assuming that Fraserosphenodon latidens is an opisthodontian, this clade also occurred in Europe during the Late Triassic, but later went extinct in that area after the breakup of Pangaea (or are yet to be found in the Old-World record). If Fraserosphenodon latidens is not an opisthodontian, then this clade may be entirely endemic to the Americas.
Conclusions
Trullidens purgatorii n. gen. n. sp., a Norian-aged opisthodontian sphenodontian, reveals the presence of sphenodontians with derived mandibular morphology from Upper Triassic rocks in western North America. It is also the most complete and phylogenetically informative Triassic-aged lepidosaur known from western North America. The dental morphology of Trullidens purgatorii n. gen. n. sp. indicates an herbivorous diet, and the unique ‘incisor-like’ tooth suggests a novel ecological function within Lepidosauria. Extant herbivorous lepidosaurs are important elements of global ecosystems, particularly in the tropics, and the presence of Trullidens purgatorii n. gen. n. sp. in the Upper Triassic paleotropics demonstrates the antiquity of herbivorous lepidosaur ecologies. The addition of this taxon to the rhynchocephalian assemblage from upper Triassic strata of western North America shows similarity of this fauna to coeval rhynchocephalian assemblages of Europe and South America, where there is a mixture of early-diverging taxa and well-nested taxa with derived craniodental morphologies. The presence of a Norian-aged opisthodontian from North America shows that this clade achieved a near-global distribution by the Late Triassic, with a hidden biogeographic radiation earlier in the Triassic.
Our investigation of the geology and paleontology of Upper Triassic strata exposed by the Purgatoire River in southeastern Colorado reveals the diversity of this assemblage, with biostratigraphic links to the Chinle Formation, as well as unique elements endemic to this system. Further investigation of this geological system may reveal other unique aspects of the fauna, as well as more robust links to other Upper Triassic strata both regionally and globally.
Acknowledgments
M.R. Stocker and S.J. Nesbitt of the Virginia Tech Paleobiology Group provided funding for and conducted μCT scanning, which was conducted at the Duke Shared Instrumentation Facility and facilitated by J. Gladman. J. Autry, the Southeast Colorado Project Director at The Nature Conservancy, facilitated access to the J.E. Canyon Ranch and provided logistical support, supplies, and housing for the authors. B. Preston, S. Korbitz, B. Small, and Y. Huang assisted in field work. The Virginia Tech Paleobiology Research Group provided useful critiques and suggestions for figures in the manuscript. M. R. Stocker provided useful critiques and discussions on the manuscript. W.G. Parker of Petrified Forest National Park provided assistance in identifying aetosaur osteoderms. K. MacKenzie and J. Sertich of the DMNH facilitated curation of specimens used in this study. B. Creisler provided valuable etymological assistance. D. Whiteside and an anonymous reviewer provided valuable reviews of the manuscript.
Data availability statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.2jm63xsnw.
Supplemental data include: Supplemental data 1 (S1), re-coding of characters; Supplemental data 2 (S2), Nexus file containing character scorings used in phylogenetic analyses; Supplemental data 3 (S3), character list used in phylogenetic analysis; Supplemental data 4 (S4), complete list of unambiguous synapomorphies common to the seven MPTs obtained in the parsimony analysis; Supplemental data 5 (S5), complete list of unambiguous synapomorphies found in the consensus tree obtained in the parsimony analysis; Supplemental data 6 (S6), rhynchocephalian temporal range data set used in time-scaled analysis.