


Introduction
Noncalcified macroalgae with monopodial branching preserved as carbonaceous compressions were first reported from Ordovician strata by Whitfield (Reference Whitfield1894). The specimens described in that report had been collected from two localities, one in southwestern Wisconsin (Platteville area) and the other in eastern New York (Middleville area). The New York material was new to science and Whitfield (Reference Whitfield1894) assigned it to a new genus and species, Primicorallina trentonensis Whitfield, Reference Whitfield1894. The Wisconsin material, on the other hand, had been collected several decades earlier and included the type material of two taxa erected by James Hall, Buthograptus laxus Hall, Reference Hall1861 and Oldhamia fruticosa Hall, Reference Hall1865. With some reservation, Hall (Reference Hall1861, Reference Hall1865) had regarded these taxa as graptolites, but Whitfield interpreted them as having an algal affinity and transferred O. fruticosa to Callithamnopsis Whitfield, Reference Whitfield1894, a newly erected genus with a name derived from that of the living red alga Callithamnion. The slabs bearing the Wisconsin specimens also contained material assigned by Whitfield to an additional macroalgal genus and species, Chaetocladus plumula Whitfield, Reference Whitfield1894. For over 150 years, the only material of the three Wisconsin taxa was that available to Hall and Whitfield, which amounted to a handful of specimens. Here, new occurrences of Buthograptus laxus and Callithamnopsis fruticosa are reported from southern Wisconsin and two new Buthograptus species are described, the first recognized within the genus beyond the type species. The new occurrences, comprising six localities, provide the first detailed information on the paleoecologic and stratigraphic context of Buthograptus and Callithamnopsis and allow for further study of their phylogenetic affinity. None of the new Wisconsin localities has yielded additional examples of Chaetocladus plumula, but herein thalli with a similar form from the Ordovician Big Hill Formation of northern Michigan are formally assigned to Chaetocladus dubius (Spencer, Reference Spencer1884), known previously only from the Silurian of Ontario, and associated specimens are assigned to C. capitatus LoDuca, Melchin, and Verbruggen, Reference LoDuca, Melchin and Verbruggen2011, known previously only from the Silurian of Arctic Canada. Collectively, the Wisconsin and Michigan material provides new insights about macroalgae in the Ordovician biosphere and the early evolution of the Ulvophyceae (Chlorophyta).
Geologic setting and age
The new Wisconsin material described herein was recovered from six exposures of the Platteville Formation in the southern part of the state. These are situated, respectively, near the towns of Dickeyville, Platteville, New Glarus, Sun Prairie, and Columbus. Four of the localities are quarry exposures; the new material from the Platteville area was recovered from stream cuts along Blockhouse and Snowdon creeks, approximately 5 and 8 km south of Platteville. These six exposures trace an arc along the southern part of the Wisconsin Dome with a total length of ~160 km. The westernmost alga-bearing intervals, at Dickeyville and Platteville, are within gray to brown micrites of the Quimbys Mill Member, the uppermost member of the Platteville Formation (Choi and Simo, Reference Choi and Simo1998). The easternmost, at Columbus and Sun Prairie, are within chocolate-brown shale partings in the basal beds of the Pecatonica Member, the lowermost member of the Platteville Formation, and the New Glarus material is from chert nodules in the middle part of the Pecatonica Member. The Michigan specimens were all recovered from fine-grained dolostone beds of the Big Hill Formation at a quarry exposure within Delta County in the southwestern part of the Upper Peninsula (Lamsdell et al., Reference Lamsdell, LoDuca, Gunderson, Meyer and Briggs2017), ~330 km to the northeast of the easternmost Wisconsin locality. Material from the Platteville Formation is Sandbian (“Turinian”) in age, whereas that from the Big Hill Formation is late Katian (“Richmondian”). A summary of the taxonomic composition of the algal floras by locality for the new Wisconsin material is provided in Table 1.
Table 1. Taxa by locality for new material described herein from the Platteville Formation of southern Wisconsin. Localities as follows: 1, Dickeyville; 2, Snowdon Creek; 3, Blockhouse Creek; 4, New Glarus; 5, Sun Prairie; 6, Columbus. See text for detailed locality information.

Materials and methods
The Wisconsin and Michigan study material comprises ~50 small slabs in total, many of which bear multiple specimens. Much of this material was collected by amateur fossil collectors G. Gunderson and R. Meyer, who subsequently donated the specimens to the University of Wisconsin Geology Museum and brought the material to the attention of the author.
High-resolution morphometric data were collected from digital images using the open source program ImageJ. Scanning electron microscopic (SEM) study of the material was conducted using a Hitachi S-3200N and a JEOL JSM-7800 F at the Electron Microbeam Analysis Laboratory (EMAL), Department of Earth and Environmental Science, University of Michigan. All specimens were imaged uncoated, and backscattered electron (BSE) images were obtained using an accelerating voltage of 15 keV. Acid maceration was performed following the same method employed in Cardona-Correa et al. (Reference Cardona-Correa, Piotrowski, Knack, Graham, Kodner and Geary2016). For this, three small pieces of limestone, each with an approximate volume of 1 cm3 and with specimens of Buthograptus laxus visible on the surface, were utilized from the Quimbys Mill Member of the Platteville Formation at the Dickeyville locality.
Morphological terminology applied herein is that for algae. Bryopsidales terminology follows Taylor (Reference Taylor1960) and Dasycladales terminology follows Berger and Kaever (Reference Berger and Kaever1992).
Repositories and institutional abbreviations.—Types, figured, and other specimens examined in this study are deposited in the following institutions: University of Wisconsin Geology Museum (UWGM), Madison, USA; Smithsonian Institution National Museum of Natural History (PAL, P), Washington, D.C., USA; University of Michigan Herbarium (UMH), Ann Arbor, USA; Redpath Museum (RM), Montreal, Canada; Royal Ontario Museum (ROMIP), Toronto, Canada.
Paleoecology and taphonomy
The alga-bearing intervals within the Platteville Formation contain very little and, in some cases, nothing in the way of associated fauna. At the Dickeyville and New Glarus localities, associated macrofossils are limited to scarce strophomenid brachiopods, bryozoa, and crinoid columnals; scattered small orthid brachiopods and trilobite fragments accompany the material at the Platteville-area localities, and the alga-bearing intervals at the Columbus and Sun Prairie localities lack any associated shelly fauna. Because noncalcified macroalgae are the dominant macrofossils in each case, all of these intervals qualify as examples of algal-Lagerstätten (see LoDuca et al., Reference LoDuca, Melchin and Verbruggen2011, Reference LoDuca, Bykova, Wu, Xiao and Zhao2017). The unusual compositions of these biotas, as well as a complete absence of bioturbation, indicate a dysoxic depositional environment with poor bottom-water circulation.
Algal material from the Big Hill Formation is associated with a Lagerstätte in which medusae, lingulid brachiopods, and chasmataspidid arthropods are the most common faunal elements (Lamsdell et al., Reference Lamsdell, LoDuca, Gunderson, Meyer and Briggs2017). Other associated fossils include eurypterids, cephalopods, gastropods, and isoteline trilobites. In addition to the macroalgal material described herein, this deposit has yielded the dasycladalean alga Archaeobatophora typa Nitecki, Reference Nitecki1976. As with the Platteville occurrences, the unusual and limited taxonomic composition of the biota together with minimal bioturbation points to a dysoxic setting, but here a lack of rhynchonelliform brachiopods combined with an abundance of lingulids may indicate a salinity other than normal marine, although sedimentological evidence of hypersalinity, such as evaporite molds, is lacking. Further evidence of a stressed environment is provided by microbial mats.
All of the macroalgal taxa from the Wisconsin and Michigan deposits show thallus morphologies typical of the “Ordovician Flora” of LoDuca et al. (Reference LoDuca, Bykova, Wu, Xiao and Zhao2017). This distinctive flora, characterized by a preponderance of forms with monopodial branching, persisted through the Silurian and replaced the preceding Cambrian Flora, which comprised comparatively simple forms.
In most cases, the delicate algal thalli are largely intact, suggesting little transport prior to burial. For the Wisconsin material, no definitive holdfasts are known, which may indicate that thalli had been ripped from these structures immediately prior to burial. The alga-bearing intervals at the Columbus and Sun Prairie localities contain localized dense assemblages of detached Callithamnopsis fruticosa laterals, but these may have been shed during thallus ontogeny, as is known for both living and fossil dasycladalean algae (Berger and Kaever, Reference Berger and Kaever1992; LoDuca and Tetreault, Reference LoDuca and Tetreault2017). Specimens from the Michigan locality assigned herein to Chaetocladus capitatus appear to have been buried upright, in life position.
Buthograptus material from the Dickeyville, Platteville, and New Glarus localities shows the central axes with a degree of 3-D preservation, and the same applies in some cases to the pinnules, but individual thallus elements from the Columbus and Sun Prairies localities, including those belonging to Callithamnopsis, are completely flattened. Buthograptus fronds from all of these localities show some twisting, curling, and warping, so that thalli rarely form perfectly planar surfaces. In some cases, fronds show upwards of 30° of local axial twisting and/or several millimeters or more of gently undulation from one end to the other.
At the Dickeyville, Platteville, Columbus, and Sun Prairie localities, Buthograptus material is preserved as carbonaceous compressions, and the same applies to Callithamnopsis thalli from the latter. SEM images of Buthograptus thalli from the Dickeyville and Sun Prairie localities indicate that the carbonaceous material has a fractured and “mudcracked” texture. Thalli from the Dickeyville and Platteville localities (Quimbys Mill Member) locally lack the carbonaceous material, but in such cases the general form is still indicated by shallow external molds. Buthograptus material from the New Glarus locality (Pecatonica Member) is preserved in chert nodules. Here, too, some parts of thalli show a thin carbonaceous film, in this case with rare local overprinting by pyrite, and other areas are preserved only as external molds. Notably, thallus elements that show a degree of 3-D preservation from this deposit are composed of silica.
Systematic paleontology
Division Chlorophyta Reichenbach, Reference Reichenbach1828
Class Ulvophyceae Mattox and Stewart, Reference Mattox, Stewart, Irvine and John1984
Order Bryopsidales Schaffner, Reference Schaffner1922
Genus Buthograptus Hall, Reference Hall1861
Type species
Buthograptus laxus Hall, Reference Hall1861 from the Ordovician (Sandbian) Platteville Formation in the vicinity of Platteville, Wisconsin, by original monotypy.
Diagnosis
Erect nonmineralized complanate and plumose thallus consisting of an unbranched flexuous cylindrical central axis from which extend simple slender unbranched pinnules arranged in opposite to subopposite fashion, these emerging at nearly right angles to the central axis, becoming wider distally by a factor of two or more; pinnule endings typically acute or rounded, in some cases mucronate. External surface without pores or other openings.
Occurrence
Known only from the Ordovician (Sandbian) Platteville Formation in southern Wisconsin.
Remarks
The formal diagnosis presented above hews close to the description of the Buthograptus type species, B. laxus, provided by Whitfield (Reference Whitfield1894), including gradual distal widening of the pinnules. Whitfield (Reference Whitfield1894, p. 352), however, noted that “the lateral pinnules are not a part of the central stipe, but are separate organs articulated by a club-shaped end to the central stipe.” In the course of the present study, through examination of the holotype slab as well as many new specimens, some referred to B. laxus and others to two new species assigned to the genus, it was determined that articulation of this nature is not present (see below).
Several fossil genera are broadly similar to Buthograptus. The monospecific macroalgal taxon Menieria Wang, Jin, and Zhan, Reference Wang, Jin and Zhan2014, erected on the basis of material from the lower Silurian of Anticosti Island (Wang et al., Reference Wang, Jin and Zhan2014), as with Buthograptus, is characterized by distally expanded lateral appendages arrayed at close intervals along the length of a central supporting structure. The two taxa differ in that the lateral appendages of Menieria, as described by Wang et al. (Reference Wang, Jin and Zhan2014), are helically arranged around the supporting structure, as opposed to the pinnate arrangement characteristic of Buthograptus. A further difference is that the thallus of Menieria shows an additional level of branching relative to Buthograptus, such that lateral elements are attached to supporting axes, which in turn are arranged along the length of a roughly cylindrical primary axis (=central axis of Wang et al., Reference Wang, Jin and Zhan2014). Dawson (Reference Dawson1873, p. 26) noted a resemblance between Buthograptus and forms that were assigned by Hall (Reference Hall1858) to Plumalina Hall, Reference Hall1858, a hydroid known primarily from the Devonian but also reported from the Silurian (Muscente and Allmon, Reference Muscente and Allmon2013). Both genera are characterized by a plumose, centimeter-scale form comprising an erect central axis that gives rise at close intervals to undivided side branches. In this case, the key morphological distinction concerns the form of the side branches—those of Buthograptus widen distally whereas those of Plumalina do not. Additional differences between the Buthograptus type species and that of Plumalina (P. plumaria Hall, Reference Hall1858) include the angle formed between the proximal portions of the side branches and the central axis (~90° versus 50–60°), side branch stiffness and curvature (flexible and upcurved versus rigid and rectilinear), and central axis morphology (strictly unbranched versus occasionally branched). The distally expanded pinnule shape of Buthograptus also serves to distinguish it from the Ordovician hydroid Pennalina Cope, Reference Cope2005, known only from Wales, and Webbyites Kraft, Kraft, and Prokop, Reference Kraft, Kraft and Prokop2001, known only from Bohemia, but a further difference is that these taxa are characterized by a distinctly alternate rather than roughly opposite arrangement of the cylindrical side branches, although in Pennalina these structures can have an upcurved as opposed to rectilinear form. A plumose form also characterizes Dowlingia Fry, Reference Fry1983, erected as an algal taxon on the basis of material from the Ordovician of Manitoba (Fry, Reference Fry1983). Two of the three species assigned to the genus, D. penna Fry, Reference Fry1983 and D. gracillima (Whiteaves, Reference Whiteaves1895), have undivided side branches, which can be arranged in subopposite fashion. These species, however, as with Plumalina, differ from Buthograptus by lacking a distally expanded form for the laterals. Whiteavesia Fry, Reference Fry1983, which occurs with Dowlingia, differs in that the laterals are spirally arranged and occasionally branch dichotomously (Fry, Reference Fry1983).
With regard to phylogenetic affinity, the gross morphology and carbonaceous composition of Buthograptus invite comparison with graptolites, hydroids, and macroalgae. In describing the taxon, Hall (Reference Hall1861, p. 18) regarded it as allied with graptolites on the basis of the “appearance, color and texture” of the material and oval features interpreted as “cellules” observed on the central axis and, with less certainty, the branches. Whitfield (Reference Whitfield1894, p. 351–352), however, rejected such an affinity, noting: “There are no definite serratures or cells on either the midrib or on the lateral pinnules, but along the line of the midrib may sometimes be seen a series of dots or punctures which were supposed to represent the apertures of cells, analogous to those of Graptolites, but which now prove to be only depressions in its surface.” The present investigation did not detect evidence of theca on the central axes or pinnules of any of the study specimens, including in BSE images, although the local removal of patches of carbonaceous material by the splitting of laminae during collection may produce irregular areas that, under low magnification, might appear as pores. Nonetheless, a graptolite affinity for Buthograptus might be possible if each side branch, in its entirety, constituted a single large theca with a terminal aperture because tubaria comprising a central stipe with large conical thecae are known for some early graptolites, such as Tarnagraptus Sdzuy, Reference Sdzuy1974 (Maletz et al., Reference Maletz, Steiner and Fatka2005; LoDuca and Kramer, Reference LoDuca and Kramer2014; Maletz, Reference Maletz2017). Such an interpretation requires that the ends of the pinnules be open, but in this case good evidence exists to the contrary, including among material assigned herein to new species within the genus. A complete lack of fusellae in BSE images also argues against a graptolite affinity because fusellae have been observed in material assigned to Tarnagraptus as well as similar graptolite taxa (LoDuca and Kramer, Reference LoDuca and Kramer2014; Maletz, Reference Maletz2017). Among hydroids, examples of feather hydroids are known with a form roughly akin to that of Buthograptus, including the extant Aglaophenia. Nonetheless, a hydroid affinity is difficult to reconcile with several features of Buthograptus. First, none of the specimens shows evidence of polyps, despite the fact that many show a substantial carbonaceous film. Most Plumalina specimens also lack evidence of polyps, but box-like modules, each with a central pore or groove, have been documented on the hydrocladia of some P. plumaria and P. densa specimens, and these are comparable to the polyp bases of extant hydroids (Sass and Rock, Reference Sass and Rock1975; Muscente and Allmon, Reference Muscente and Allmon2013). Second, the hydrocladia of Plumalina and other feather hydroids do not show the distally expanded form characteristic of the pinnules of Buthograptus. Finally, the tips of feather hydroid colonies are typically triangular, a form that emerges because the upper hydrocladia progressively decrease in length, do not extend above the apex of the hydrocaulus, and maintain a consistent angle relative to the hydrocaulus. In contrast, the tips of Buthograptus thalli are characterized by a somewhat rounded form because the uppermost pinnules extend well above the apex of the central axis and curve more strongly upward than those below.
In light of the foregoing, a macroalgal affinity for Buthograptus emerges somewhat by default, but a good case can be made beyond negative evidence. In particular, such an affinity is supported by direct association with definitive macroalgal material and by an overall morphology resembling that of extant algae, particularly the bryopsidalean green alga Caulerpa. In this regard, the pinnules of B. laxus find a good analogue in Caulerpa sertularioides Howe, Reference Howe1905 in terms of size and spacing along the supporting axis, and those of the new Buthograptus species described herein from New Glarus bear a strong resemblance to the conspicuously stouter pinnules of Caulerpa taxifolia Agardh, Reference Agardh1817, including constricted bases. Pinnule arrangement on a given Buthograptus thallus also corresponds to that of the aforementioned Caulerpa species in that the fronds are oppositely pinnate, but small vertical offsets between pinnules of a particular pair are not uncommon (e.g., Taylor, Reference Taylor1960, pl. 11, fig. 4). In arguing for an algal affinity for Buthograptus, Whitfield (Reference Whitfield1894, p. 352) noted this strong resemblance to Caulerpa, stating “Among the living forms of marine Algae on the Florida coast and elsewhere we have a form known as Caulerpa plumaris [ =C. sertularioides] which to the naked eye is so exactly a counterpart of Buthograptus laxus that a figure of one would answer equally well for that of the other.” Ultimately, however, Whitfield rejected a close affinity between Buthograptus and Caulerpa because the pinnules of the former were interpreted as structures independent of, and articulated with, the main axis. Notably, Dawson (Reference Dawson1873, p. 25–26) had previously rejected a close affinity between Lycopodites vanuxemii (=Plumalina plumaria) and Caulerpa for precisely the same reason, and it is possible that this influenced Whitfield's interpretation in this regard. Evidence uncovered in the present study, however, points away from such a relationship between the pinnules and central axis and, at any rate, such a relationship would not necessarily preclude a Caulerpa affinity because it is now known that the pinnules in at least one species of Caulerpa can detach from the supporting structure to facilitate vegetative reproduction (Renoncourt and Meinesz, Reference Renoncourt and Meinesz2002). Buthograptus does show a substantial departure from Caulerpa in that no specimens were observed with a stolon. In addition, the oldest fossils assigned to Caulerpa are from the Permian (Gustavson and Delevoryas, Reference Gustavson and Delevoryas1992) and molecular studies suggest that Caulerpaceae split from the sister-clade Pseudochlorodesmis sometime in the late Paleozoic or early Mesozoic (Draisma et al., Reference Draisma, Prud'homme van Reine, Sauvage, Belton, Gurgel, Lim and Phang2014). For these reasons, Buthograptus is maintained as a distinct genus. Nonetheless, likely examples of calcareous bryopsidalean algae with complex thalli are known from the Ordovician, including Dimorphosiphon Høeg, Reference Høeg1927 and Palaeoporella Stolley, Reference Stolley1893. Therefore, given this fact and in consideration of all of the foregoing evidence, Buthograptus is interpreted herein as an early bryopsidalean alga with a thallus architecture resembling the fronds of Caulerpa.
As with all material regarded as the remains of noncalcified macroalgae in the fossil record that lacks reproductive structures and direct information about chlorophyll composition or cellular-level anatomy, it is not possible to unequivocally determine if Buthograptus is a red, green, or brown alga because convergent evolution of thallus form among these groups is not uncommon. Among living macroalgae, however, only the fronds of pinnate species of Caulerpa closely resemble material of Buthograptus, and there are no details known for Buthograptus thalli that would preclude a siphonous organization, a requirement for consideration as a bryopsidalean alga.
If Buthograptus is correctly interpreted as a bryopsidalean alga, an important implication is that plumose thalli evolved very early in the history of the group and among forms that did not produce a stolon. Whether such forms, then, gave rise to extant caulerpids via the evolution of a stolon, or instead correspond to an instance of convergent evolution, remains to be determined. The fact that a plumose thallus form is also characteristic of the extant bryopsidalean Bryopsis, a form that resembles Buthograptus in its lack of a pronounced stolon but differs by being much more flaccid and having substantially thinner thallus elements, provides some support for the convergent evolution hypothesis because molecular data indicate that Caulerpa is more closely related to several extant non-plumose forms, including Halimeda and Avrainvillea, than it is to Bryopsis (Verbruggen et al., Reference Verbruggen, Ashworth, LoDuca, Vlaeminck, Cocquyt, Sauvage, Zechman, Littler, Littler, Leliaert and De Clerk2009). Alternatively, it is conceivable that Menieria represents a transitional form between Buthograptus and Caulerpa, with the thallus element referred to by Wang et al. (Reference Wang, Jin and Zhan2014) as the “central axis” being anatomically, if not functionally, equivalent to the stolon.

Figure 1. Buthograptus laxus Hall, Reference Hall1861, Buthograptus gundersoni n. sp., and Buthograptus meyeri n. sp. from the Ordovician (Sandbian) Platteville Formation of southern Wisconsin. (1, 2) Slab bearing many B. laxus thalli, illustrated as a line drawing in Whitfield (Reference Whitfield1894), Platteville area, PAL 339997; (1) specimen at arrow in 2, proposed herein as the lectotype; (2) entire slab, photograph provided by M. Florence. (3–5) B. laxus from the Quimbys Mill Member at Platteville and Dickeyville; (3) two overlapping thalli, Dickeyville, UWGM 5083; (4) detail of area marked by arrow in 3 showing pinnules; (5) nearly complete thallus, Snowden Creek, Platteville, UWGM 2407. (6, 7) B. gundersoni n. sp. holotype, Pecatonica Member, New Glarus, UWGM 2384; (6) detail of area marked by arrow in 7 showing pinnules; (7) entire specimen. (8, 9) B. meyeri holotype slab, Pecatonica Member, Columbus, UWGM 2408; (8) detail of area marked by arrow in 9 showing pinnules; (9) two thalli, holotype at right, with scattered detached laterals of Callithamnopsis fruticosa evident in places. (10–14) B. laxus, Quimbys Mill Member at Snowden Creek, Platteville, UWGM 2407; (10) entire specimen, pinnules on right side missing owing to irregular matrix breakage; (11) detail of area marked by left arrow in 10 showing pinnules; (12) detail of area marked by right arrow in 10 showing pinnules with enlarged bases adjacent to main axis (arrows); (13) entire specimen, pinnules in places missing owing to irregular matrix breakage; (14) detail of area marked by arrow in 13 showing central axis and proximal parts of pinnules. Specimens in (1–4, 6, 7, 14) photographed dry; specimens in (5, 8–13) photographed immersed in alcohol. Scale bars are (1, 3, 5, 7, 9, 10, 13) 5 mm; (2) 10 mm; (4, 6, 8, 11, 12, 14) 1 mm.

Figure 2. Buthograptus Hall, Reference Hall1861 from the Ordovician (Sandbian) Platteville Formation (Pecatonica Member) of southern Wisconsin and modern Caulerpa from the Pacific. (1–12) Buthograptus gundersoni n. sp., New Glarus; (1) slab with many thalli, UWGM 2380; (2) UWGM 2383; (3) nearly complete thallus, pinnules on left missing owing to irregular matrix breakage, UWGM 2381; (4) UWGM 2378; (5) UWGM 2379; (6) counterpart of specimen in 5, UWGM 2383; (7) detail of thallus in 2 showing pinnules; (8) detail of thallus at arrow in 11 showing pinnules; (9) three thalli, UWGM 2381; (10) detail of upper part of thallus at right in 9 showing rounded thallus tip; (11) two thalli, UWGM 2381; (12) detail of central part of thallus in 5 showing basal parts of pinnules. (13) Caulerpa taxifolia, individual frond, pressed herbarium specimen, southern Japan, UMH 682355. (14) Buthograptus meyeri n. sp., Columbus, UWGM 2348. (15) Buthograptus laxus Hall, Reference Hall1861, Columbus, UWGM 2354. All specimens photographed dry. Scale bars are (1, 3–6, 9, 11, 13–15) 5 mm; (2) 3 mm; (7, 8, 10, 12) 1 mm.

Figure 3. Reflected light and BSE images of Buthograptus laxus Hall, Reference Hall1861 from the Ordovician (Sandbian) Platteville Formation of southern Wisconsin. (1–3) UWGM 2366, Pecatonica Member, Sun Prairie; (1) reflected light image of entire specimen, pinnules at left concealed by thin veneer of overlying matrix; (2) BSE image of the area indicated by the upper arrow in 1 showing the distal part of a pinnule; (3) BSE image of the area indicated by the lower arrow in 1 showing several pinnules emerging from the central axis and distinctive “mudcracked” appearance, arrow indicates pinnule with an enlarged basal area. (4–6) UWGM 5084, Quimbys Mill Member, Dickeyville; (4) reflected light image; (5) BSE image of the area indicated by the upper arrow in 4 showing several pinnules emerging from the central axis and fractured character of the material; (6) BSE image of the area indicated by the lower arrow in 4 showing part of a pinnule, light areas at center are patches of adhering matrix. Scale bars are (1, 4) 5 mm; (2, 3, 5) 0.2 mm; (6) 0.1 mm.
Buthograptus laxus Hall, Reference Hall1861
Figures 1.1–1.5, 1.10–1.14, 2.15, 3.1–3.6
- Reference Hall1861
Buthograptus laxus Hall, p. 19.
- Reference Hall1865
Buthograptus laxus; Hall, p. 17, text fig. 25.
- Reference Geinitz1866
Buthograptus laxus; Geinitz, p. 125.
- Reference Hall1868
Buthograptus laxus; Hall, p. 185, text fig. 27.
- Reference Dawson1873
Buthograptus laxus; Dawson, p. 26.
- Reference Whitfield1894
Buthograptus laxus; Whitfield, p. 351, pl. 11, figs. 1–3.
- Reference Whitfield1895
Buthograptus laxus; Whitfield, p. 40, pl. 4, figs. 1–3.
- Reference Ruedemann1909
Bythograptus laxus; Ruedemann, p. 195.
- Reference Teller1911
Buthograptus laxus; Teller, p. 185.
- Reference Ruedemann1947
Buthograptus laxus; Ruedemann, p. 140, 265.
- Reference Johnson1961
Buthograptus laxus; Johnson, p. 86.
- Reference Johnson1961
Chaetocladus plumula; Johnson, pl. 37, fig. 5.
- Reference Choi and Simo1998
Buthograptus laxus; Choi, p. 24.
- Reference Choi and Simo1998
Buthograptus laxus; Choi and Simo, p. 442.
Lectotype
PAL 339997 (Fig. 1.1) from the Ordovician (Sandbian) Platteville Formation in the vicinity of Platteville, Wisconsin, as designated herein.
Diagnosis
Buthograptus with slender pinnules typically 5–6 mm long and 0.3–0.4 mm in maximum width and with a length-to-width ratio of at least 12 for the longer pinnules on a given thallus; interval between adjacent pinnules typically less than two times proximal pinnule width.
Occurrence
Known from fine-grained limestone beds within the Quimbys Mill Member of the Platteville Formation at a small quarry on the west side of Church Road, ~5 km north of Dickeyville (42.667243°N, 90.571422°W) (locality “CN” in Choi and Simo, Reference Choi and Simo1998) and at stream exposures along Blockhouse and Snowdon creeks, 5 and 8 km south of Platteville, respectively (42.686364°N, 90.486215°W; 42.655638°N, 90.520264°W). Also known from chocolate-brown shale partings in the lowermost part of the Pecatonica Member of the Platteville Formation exposed in a quarry northeast of Sun Prairie on Highway 151 (43.207127°N, 89.174085°W) (Stop 2 of Simo et al., Reference Simo, Choi, Freiberg, Byers, Dott, Saylor and Mudrey1997) and a small pit (now filled) west of Columbus on County Highway K (43.327318°N, 89.147205°W).
Description
Thallus noncalcified, complanate, plumose, typically with a maximum width of 6–10 mm and reaching 30 mm in height (Table 2), comprising an undivided central axis with unbranched pinnules arranged in opposite to subopposite fashion, these separated vertically by intervals of 0.2–0.4 mm, emerging at nearly right angles and curving initially downward very slightly and then more strongly upward, typically 5–6 mm long at mid-thallus and somewhat shorter at lower and upper parts of thallus (Figs. 1.1–1.3, 1.5, 1.13, 2.15). Central axis of uniaxial construction, cylindrical, flexuous, 0.4–0.7 mm wide, apex rounded (Figs. 1.12, 1.14, 3.3, 3.5). Pinnules typically 0.15–0.2 mm wide where they emerge from the central axis, gradually expanding distally by a factor of two; faint longitudinal striations occasionally evident (Fig. 1.11); in rare cases, proximal portions show a bulbous form (Figs. 1.12, 3.1, 3.3); endings typically acute or rounded, in some cases mucronate (Figs. 1.4, 1.11, 3.2). Uppermost pinnules extend above top of central axis, curved more strongly upward than those below, conveying a somewhat rounded form to thallus tips (Fig. 1.5). No holdfast or reproductive structures observed.
Table 2. Buthograptus morphometric data. Pinnule L and W values represent the length and maximum width of the longest pinnule on a given thallus. All measurements in millimeters. Abbreviations: W, width; H, height; L, length; I, average interval between adjacent pinnules (N = 10); D, number of pinnules per 10 mm of central axis. Holotype and lectotype specimens indicated by asterisk.

Materials
UWGM 2349, 2350, 2353, 2354, 2355, 2356, 2357, 2358, 2359, 2363, 2364, 2366, 2377, 2407, 2409, 2410, 5083, 5084.
Remarks
Whitfield (Reference Whitfield1894, p. 352) described the pinnules of B. laxus as “articulated” with the central axis and regarded them as “separate organs” on this basis. Close examination of the type material and more than 25 new specimens, however, could not confirm such an articulation, and herein the pinnules and central axis are regarded to comprise a fully integrated, continuous entity. This revised interpretation is supported by two key observations. First, definitive evidence of a cross-wall at the junction between the pinnule and central axis is lacking, including in BSE images. Second, no examples of thalli were identified that show unequivocal evidence of pre-burial pinnule removal at the point of contact with the central axis, including among material examined by Whitfield. Some central axes do show localized intervals that lack pinnules. In many cases, however, this can be attributed to damage that occurred during collection, when slabs were split to reveal the specimens. Such instances are typically associated with areas of thalli that show abrupt twists or bends or intersect localized irregularities of the matrix surface. In other cases, pinnules only appear to be missing, but instead are locally concealed by overlying matrix. Here, too, such instances are typically associated with areas of thalli that show abrupt twists or bends or intersect localized irregularities of the matrix surface. No evidence could be found of punctures or other features on the central axis that could represent the attachment points of pinnules lost pre-burial, including in BSE images. Small depressions of irregular form are occasionally evident on the central axis, but in some cases these are superficial and consistent with artifacts arising from compaction of an originally cylindrical structure, and in the remainder result from localized flaking of some of the carbonized material during collection. Also important in this regard is that no definitive examples of isolated, detached pinnules could be identified. This is particularly notable because numerous detached laterals of Callithamnopsis fruticosa occur in direct association with Buthograptus thalli at the Sun Prairie and Columbus localities (see below) and, given their delicate construction, these would have been transported from the place of origin much more readily than Buthograptus pinnules. Whitfield (Reference Whitfield1894) did not mention observing examples of isolated detached pinnules, and his line drawing of an individual pinnule (Whitfield, Reference Whitfield1894, pl. 11, fig. 3) is not identified as a detached example. A few of the pinnules on the type slab and the new slabs do show somewhat expanded portions adjacent to the central axis, in keeping with the “club-like ends” mentioned by Whitfield (Reference Whitfield1894, p. 352) in support of his articulation interpretation. Most, however, do not, and such features are not present on any of the material assigned to the two new Buthograptus species described in this report. The significance of this expansion is not clear, but it is possible that it reflects distortion resulting from compaction.
With regard to the distal ends of the pinnules and the nature of their terminations, Hall (Reference Hall1861, p. 18) noted a “thickened lanceolate extremity” and Whitfield (Reference Whitfield1894, p. 352) described them as “rounded or obtusely pointed at the outer end”. In the present study, it was found that in most cases the distal parts of the pinnules show damage that occurred when slabs were split during collection or are concealed by overlying matrix, which proved difficult to remove without obliterating the underlying pinnule. Numerous pinnules with intact ends were observed, however, including among specimens assigned herein to new species of Buthograptus. These show that the pinnule endings were closed in life and have shapes matching the descriptions provided by Hall and Whitfield.
Acid maceration of small slabs bearing B. laxus thalli from the Dickeyville locality did not produce any material that could be definitively identified as belonging to this species. The resulting organic material was highly fragmentary, with nearly all fragments much less than 1 mm in maximum dimension, and did not include anything that resembled axes with pinnules. This outcome is not unexpected, given the highly fractured and “mudcracked” appearance of B. laxus specimens from this locality in BSE images (Fig. 3). All of the fragments lacked evidence of cellular structure. In this regard, they clearly stand apart from material recovered using acid maceration from the lower part of the Platteville Formation at a road cut ~25 km to the north of the New Glarus locality and interpreted by Cardona-Correa et al. (Reference Cardona-Correa, Piotrowski, Knack, Graham, Kodner and Geary2016) as remnants of bryophytes. Material recovered by acid maceration from the Ordovician (Darriwilian) Winneshiek Lagerstätte of Iowa and described as possible ulvophycean algae (Nowak et al., Reference Nowak, Harvey, Liu, McKay, Zippi, Campbell and Servais2017) shows marked differences from Buthograptus in terms of thallus architecture.
In a recent overview of early Paleozoic macroalgae, Buthograptus laxus was regarded as a possible member of the “stoloniform” morphogroup (LoDuca et al., Reference LoDuca, Bykova, Wu, Xiao and Zhao2017). This tentative assignment reflected Whitfield's (Reference Whitfield1894) remark that the form of Buthograptus closely resembled that of Caulerpa. Careful examination of the type material and many new specimens during the present study did not reveal any evidence of a stolon. On this basis, the thallus architecture of the species, and more broadly the genus, is instead regarded as monopodial and consistent with the “simple monopodial morphogroup” of LoDuca et al. (Reference LoDuca, Bykova, Wu, Xiao and Zhao2017).
The new material described herein indicates that B. laxus occurs at two different stratigraphic levels within the Platteville Formation of southern Wisconsin, one near the base of the formation, within the Pecatonica Member in the Sun Prairie area, the other in the uppermost part of the formation, within the Quimbys Mill Member in the Platteville area. Choi (Reference Choi1998) and Choi and Simo (Reference Choi and Simo1998) had previously reported observing B. laxus in the Pecatonica Member in Wisconsin, noting its abundance in organic-rich shale beds within the interbedded mudstone/organic-rich shale (M-OSh) lithofacies, but these reports did not tie the observations to particular localities.
In the original description of B. laxus, Hall (Reference Hall1861) did not provide illustrations and did not designate a holotype. Later, Hall (Reference Hall1865, fig. 25) illustrated a single specimen, and Whitfield (Reference Whitfield1894, Reference Whitfield1895) illustrated other material from among the specimens used by Hall to establish the species. For a time, it seemed that all of the material on which Hall and Whitfield had based their descriptions was lost (LoDuca et al., Reference LoDuca, Bykova, Wu, Xiao and Zhao2017) because a drawer search of the paleobotany collections at the Smithsonian Institution National Museum of Natural History in the 1990s failed to locate any specimens. During the present study, the slab illustrated by Whitfield (Reference Whitfield1894, pl. 11, fig. 1) was located in the graptolites type collection at the Smithsonian (M. Florence, personal communication, 2017) (Fig. 1.2). This slab bears the number PAL 339997 and contains many individual B. laxus thalli. Herein, the specimen on PAL 339997 illustrated in Figure 1.1 is proposed as the lectotype. Precise stratigraphic and locality information for this slab and other B. laxus specimens examined by Hall and Whitfield remains unknown. When describing the species, Hall mentioned that the material had been collected from “dark-colored shaly limestone of the age of the Trenton limestone; at Platteville and vicinity, Wisconsin” (Hall, Reference Hall1861, p. 19). Whitfield (Reference Whitfield1894, Reference Whitfield1895) provided no further details in this regard, and the labels with PAL 339997 do not provide detailed stratigraphic or locality information. The lithology of PAL 339997, however, is consistent with that of the Quimbys Mill Member of the Platteville Formation in the Platteville area.
Buthograptus gundersoni new species
Figures 1.6, 1.7, 2.1–2.12
Holotype
UWGM 2384 (Fig. 1.7) from the Ordovician (Sandbian) Platteville Formation (Pecatonica Member) in a quarry south of New Glarus, Wisconsin (42.780895°N, 89.623409°W).
Diagnosis
Buthograptus with stout pinnules typically 2–4 mm long, 0.5–0.7 mm in maximum width, and with a length-to-width ratio of no more than seven for the longer pinnules on a given thallus; interval between adjacent pinnules typically less than two times proximal pinnule width.
Occurrence
Known only from the type locality.
Description
Thallus of holotype (UWGM 2384) noncalcified, complanate, plumose, with a height of 27.1 mm and a maximum width of 8.3 mm, comprising an undivided central axis with unbranched pinnules arranged in opposite to subopposite fashion, these separated vertically by intervals of 0.11 mm, emerging from the central axis at nearly right angles and becoming very slightly upcurved, with a maximum length of 4.5 mm at mid-thallus, but somewhat shorter in lowermost and uppermost parts of thallus (Fig. 1.7; Table 2). Central axis of uniaxial construction, cylindrical, flexuous, 0.8 mm wide (Fig. 1.6, 1.7). Pinnules typically with a maximum width of 0.7 mm, constricted near the base, endings typically acute or rounded, in some cases mucronate (Fig. 1.6). Specimens on UWGM 2378, 2379, 2380, 2381, and 2383 (Fig. 2.1–2.12; Table 2) similar to holotype, including constrictions at basal parts of pinnules (Fig. 2.8, 2.12) and closed endings of the pinnules (Fig. 2.7, 2.8, 2.10). A thallus on UWGM 2381 with a particularly well-preserved distal end shows the central axis with a rounded apex and the uppermost pinnules extending well above the central axis, these curved more strongly upward than those below and thereby conveying a somewhat rounded form to the thallus tip (Fig. 2.10). No holdfast or reproductive structures observed.
Etymology
Named for the individual that collected the material upon which the species was erected, Gerald Gunderson.
Materials
UWGM 2378, 2379, 2380, 2381, 2382, 2383, 2384, 2385, 2386, 2395, 2396, 2398, 2405, 2406.
Remarks
Buthograptus gundersoni n. sp. is similar to B. laxus in most key respects, including closed endings for the pinnules, but differs in the stout form of the pinnules. In this regard, on a given thallus, the longer pinnules have a length-to-width ratio (L/W) of no more than seven and often less than five, whereas those of the latter are clearly more gracile, with a L/W value of more than 12 and often greater than 14 (Table 2). In addition, the basal parts of the pinnules of B. gundersoni n. sp. are strongly constricted and never show spherical inflations of the sort occasionally evident in specimens of B. laxus, and the pinnule surfaces do not show longitudinal striations.
This species occurs only at the New Glarus locality, where it is the sole buthograptid taxon. Extant species of Caulerpa are known to display considerable phenotypic plasticity, in some cases in response to differences in light level (Cevik et al., Reference Cevik, Yokes, Cavas, Erkol, Derici and Verlaque2007; Belton et al., Reference Belton, Reine, Huisman, Draisma, Gurgel and Frederico2014). Thus, it is possible that material assigned to B. gundersoni represents an ecophenotypic variant of B. laxus as opposed to a truly separate species. Nonetheless, because morphological disparity in the area of pinnule form between the New Glarus Buthograptus material and that from all other occurrences of the genus is particularly marked, and because intermediate forms between these morphotypes are unknown, it is regarded as best at the present time to place the New Glarus material in a separate species.
Buthograptus meyeri new species
Figures 1.8, 1.9, 2.14
Holotype
UWGM 2408 (Fig. 1.9, right specimen) from the lower part of the Pecatonica Member of the Ordovician (Sandbian) Platteville Formation exposed in a small pit (now filled) west of Columbus along County Highway K (43.327318°N, 89.147205°W).
Diagnosis
Buthograptus with slender pinnules widely spaced along the central axis, the interval between adjacent pinnules typically being more than three times proximal pinnule width. Pinnules at all levels of thallus strongly upcurved, with distal portions becoming nearly parallel to the central axis.
Occurrence
Known only from the type locality.
Description
Thallus of holotype noncalcified, complanate, plumose, with a height of 35.7 mm and a maximum width of 6.1 mm, comprising an undivided central axis with unbranched pinnules arranged in opposite to subopposite fashion, these separated vertically by intervals typically 1.6–1.8 mm (Table 2). Pinnules emerge from the central axis at nearly right angles, but after a short distance become strongly upcurved, so that distal parts are nearly parallel to the central axis (Fig. 1.9). Central axis of uniaxial construction, cylindrical, flexuous, ~0.6 mm wide, apex rounded. Pinnules mostly 6–7 mm in length, gently expanding in width from 0.3 mm at the base to 0.5 mm near the distal end, endings typically acute but occasionally rounded, surface smooth, without longitudinal striations (Fig. 1.8). Uppermost pinnules extend above top of central axis, conveying a somewhat rounded form to thallus tip. Thallus adjacent to holotype similar in all key respects, but upper part not preserved (Fig. 1.9; Table 2). Thallus of UWGM 2348 similar to aforementioned, but pinnules separated by intervals typically in the range of 0.7–1 mm (Table 2). No holdfast or reproductive structures observed.
Etymology
Named for Ron Meyer, an ardent collector of Wisconsin fossils.
Materials
UWGM 2408 (two thalli), 2348 (counterpart is 2351), 2352
Remarks
Buthograptus meyeri n. sp. occurs in direct association with B. laxus at the type and only known locality for the species (Table 1), and the two species are similar in most key respects, including pinnule form and size (Table 2). They differ chiefly in pinnule density along the central axis, with B. laxus thalli having three times as many pinnules per 10 mm of central axis length in comparison to the B. meyeri n. sp. holotype. UWGM 2348 represents a somewhat intermediate case in this regard, with pinnule density only about half that observed in typical specimens of B. laxus. Nonetheless, the distinction here is still marked. Buthograptus meyeri n. sp. also differs from B. laxus in that the pinnules curve more strongly upward, so that the distal parts of the pinnules in some cases run nearly parallel to the central axis, and pinnule surfaces have not been observed with longitudinal striations. Because the pinnules of this species are rather long and were evidently more flexuous than those of B. laxus, overlaps of pinnules from adjacent tiers are not uncommon. Such overlaps can give the appearance of a single pinnule with a distal dichotomy (e.g., right side of right thallus in Figure 1.9; two places along left side of thallus in Figure 2.14), but in all such cases careful examination reveals two pinnules in juxtaposition. As with B. gundersoni n. sp., it is possible that B. meyeri n. sp. represents an ecophenotypic variant of B. laxus rather than a truly separate species. In this case, however, the occurrence of both forms in direct association reduces the likelihood of this possibility. The separate species interpretation also finds a close modern analogue in the morphological distinction between the fronds of Caulerpa ashmeadii Harvey, Reference Harvey1858 and C. sertularioides.
Order Dasycladales Pascher, Reference Pascher1931
Family Triploporellaceae (Pia, Reference Pia1920) Granier and Bucur in Granier et al., Reference Granier, Dias-Brito, Bucur and Tibana2012
Genus Callithamnopsis Whitfield, Reference Whitfield1894
Type species
Oldhamia fruticosa Hall, Reference Hall1865 from the Ordovician (Sandbian) Platteville Formation in the vicinity of Platteville, Wisconsin, by original monotypy.
Diagnosis
Noncalcified thallus with monopodial branching comprising a narrow main axis bearing branched laterals arranged in verticils spaced at intervals of 1–2 mm; main axis uniaxial, cylindrical, without segmentation; laterals branched to the sixth order, each division giving rise to three or four laterals of the next level, lateral segments cylindrical; endosporate.
Occurrence
Ordovician of southern Wisconsin and eastern New York; Silurian of western New York.
Remarks
The specimens assigned herein to Callithamnopsis fruticosa are the first for the Callithamnopsis type species from an area outside the type locality. The new material is for the most part in keeping with the detailed description of the taxon provided by Ruedemann (Reference Ruedemann1909), particularly with regard to the monopodial branching of the thallus and the characteristics of the laterals. Notably, the new specimens show the primary laterals arranged in distinct verticils (euspondyl). This is important because details of this nature cannot be clearly discerned for the type material, in many cases owing to overlapping laterals. Ruedemann (Reference Ruedemann1909, p. 202) described the primary laterals as “always arranged in pairs,” but this is consistent with a whorled arrangement in which all of the laterals in a verticil cannot be clearly observed. Notably in this regard, a verticillate arrangement is evident on a specimen on the same slab as the type material, illustrated in line drawing and assigned to C. fruticosa by Ruedemann (Reference Ruedemann1909, pl. 1, fig. 4) (Fig. 4.6), and C. delicatula Ruedemann, Reference Ruedemann1909, a form very similar to the Wisconsin species from the Ordovician of eastern New York, is characterized by a verticillate arrangement of the primary laterals (Ruedemann, Reference Ruedemann1909).
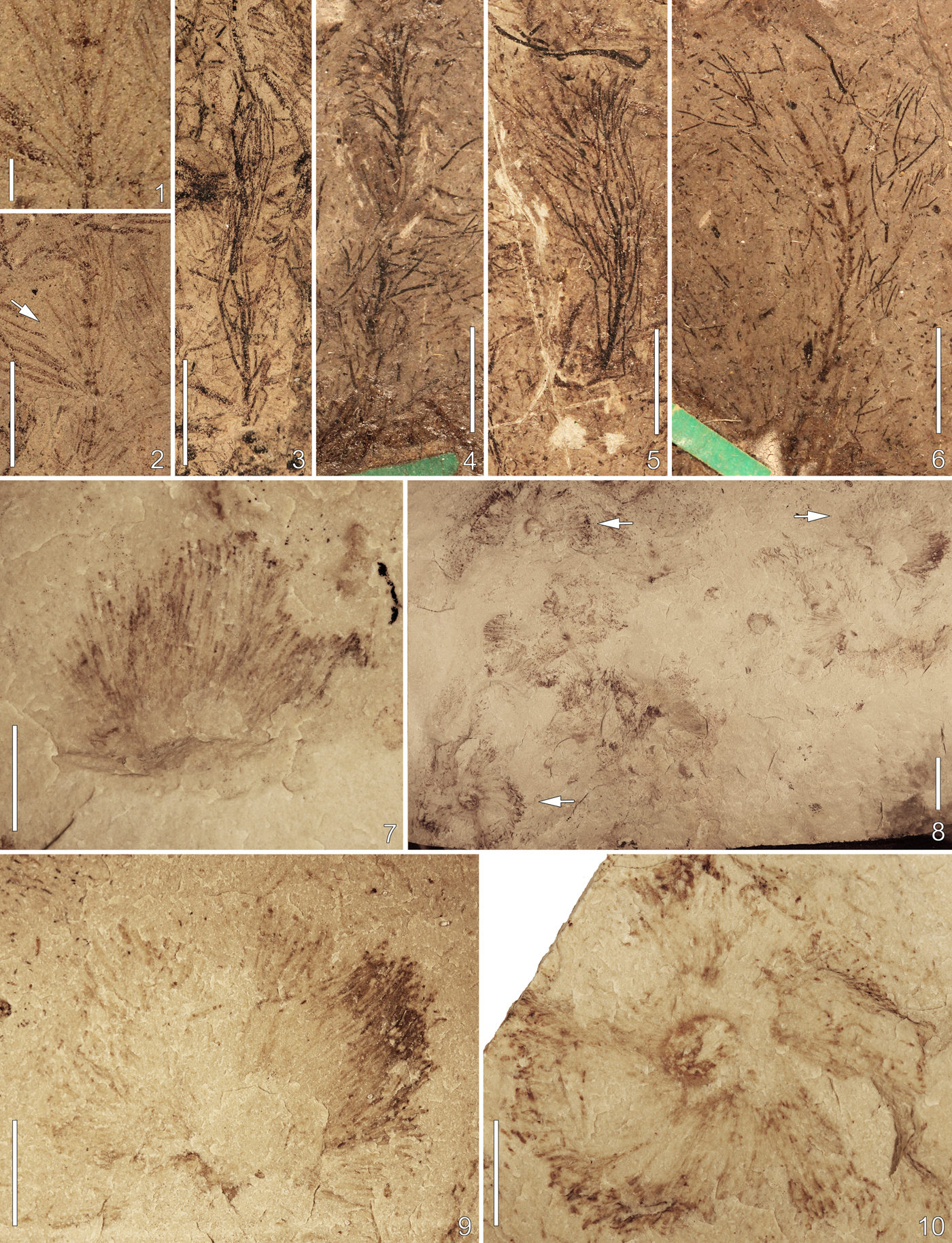
Figure 4. Callithamnopsis fruticosa (Hall, Reference Hall1865) from the Ordovician (Sandbian) of Wisconsin and Chaetocladus capitatus LoDuca, Melchin, and Verbruggen, Reference LoDuca, Melchin and Verbruggen2011 from the Ordovician (Katian) of Michigan. (1–3) C. fruticosa from the Pecatonica Member of the Platteville Formation at Sun Prairie and Columbus; (1) detail of thallus at level of arrow in 2 showing primary laterals arranged in verticils; (2) nearly complete thallus, scattered detached laterals at right, Sun Prairie, UWGM 2364; (3) nearly complete thallus, Columbus, UWGM 2411. (4–6) C. fruticosa type specimen slab, Platteville Formation, Platteville area, USNM P 41089; (4) lectotype, as proposed herein; (5) paratype; (6) thallus showing primary laterals arranged in verticils. (7–10) C. capitatus from the Big Hill Formation, Delta County, Michigan; (7) capitulum of a thallus in lateral view, UWGM 2312; (8) small patch of thalli buried upright, specimens marked by arrows at left show the main axis as a ring at center of capitulum, specimen marked by arrow at upper right shows capitulum folded over to one side, UWGM 2311; (9) enlargement of thallus marked by upper right arrow in 8 showing numerous elongate laterals that comprise the capitulum; (10) enlargement of thallus marked by lower left arrow in 8 showing the capitulum in overhead view with main axis forming ring at center. Specimens in (4–6) photographed dry; specimens in (1–3, 7–10) photographed immersed in alcohol. Scale bars are (1) 1 mm; (2–7, 9, 10) 5 mm; (8) 10 mm.
In referring to the main axis of C. fruticosa, Whitfield (Reference Whitfield1894, p. 354) described this structure as “jointed” and Ruedemann (Reference Ruedemann1909, p. 202) noted that “distinct transverse lines are visible which suggest a segmentation.” Neither report provided additional details in this regard, and such features are not shown in the associated line drawings. In examining the specimens described by Whitfield and Ruedemann during the present study, no definitive evidence was observed of original segmentation of the main axis. Some faint transverse fractures are evident, but these have a highly irregular spacing and are interpreted to have arisen from post-burial compaction. No transverse features of any kind were observed on the main axes of the two new specimens. Consequently, the main axes of C. fruticosa are regarded as continuous structures, without segmentation or articulation.
Among fossil genera, Callithamnopsis is most similar to the monotypic taxon Eocladus LoDuca et al., Reference LoDuca, Melchin and Verbruggen2011, from the upper Silurian of Arctic Canada (LoDuca et al., Reference LoDuca, Melchin and Verbruggen2011). The principal distinction between these taxa pertains to the spacing of the verticils along the main axis, these being set at intervals of ~1–2 mm in C. fruticosa, but only ~0.6 mm in E. xiaoi LoDuca, Melchin, and Verbruggen, Reference LoDuca, Melchin and Verbruggen2011. Consequently, thalli of the former are conspicuously less bushy than those of the latter. An additional difference is evident in terms of overall size, with thalli of C. fruticosa being about half the size of a typical specimen of E. xiaoi. Callithamnopsis also bears a resemblance to Inopinatella Elliott, Reference Elliott1971, Dasycladophycus Elliott, Reference Elliott1982, Archaeobatophora Nitecki, Reference Nitecki1976, Primicorallina Whitfield, Reference Whitfield1894, Heterocladus LoDuca et al., Reference LoDuca, Kluessendorf and Mikulic2003, and Uncatoella Li and Cai, Reference Li and Cai1978, but differs from each in one or more key respects. Inopinatella and Dasycladophycus, from the upper Silurian and lower Carboniferous of England, respectively, differ mainly in the branching details of the laterals because these taxa are characterized by laterals that branch respectively only to the second and third order. Inopinatella further differs in that the primary lateral segments have distinctly bulbous proximal ends. Archaeobatophora, known exclusively from the same locality in the Ordovician Big Hill Formation of Michigan as the Chaetocladus material described herein, differs in that the main axis is wider by a factor of three and shows distinct constrictions between the verticils. Heterocladus and Uncatoella, from the Silurian of Wisconsin and Devonian of China, respectively, show notable differences in that these taxa develop distinctive reproductive structures (Kenrick and Li, Reference Kenrick and Li1998; LoDuca et al., Reference LoDuca, Kluessendorf and Mikulic2003). Primicorallina differs in that the laterals are not arranged in verticils, although lateral arrangement in this taxon clearly closely approaches such a configuration. Further differences are that the laterals of Primicorallina are more robust and only branch to the third order. Callithamnopsis would also appear to be similar to Kinwowia Fry, Reference Fry1983, known from the Ordovician of Manitoba. At present, however, the type material is poorly known and a detailed comparison must await a thorough restudy of this material.
Hall (Reference Hall1865, p. 50) tentatively regarded this taxon as having a graptolite affinity on the basis of a perceived outward similarity to Dendrograptus, but also noted that “there are no visible cellules.” Whitfield (Reference Whitfield1894) regarded C. fruticosa as an alga because the specimens lacked theca and considered it allied with the extant red alga Callithamnion, placing it in the new genus Callithamnopsis Whitfield, Reference Whitfield1894. Ruedemann (Reference Ruedemann1909), however, noted that Callithamnopsis lacked the bifurcating pattern of branching characteristic of Callithamnion and indicated that lateral branching and other thallus characteristics indicated a close affinity with Primicorallina, which he regarded as a dasycladalean alga. Pia (Reference Pia and Hirmer1927, p. 67) included the genus in his overview of fossil dasycladalean algae, but expressed some uncertainty in this regard, perhaps owing to the paired arrangement of the primary laterals mentioned in the description of C. fruticosa by Ruedemann (Reference Ruedemann1909). Seward (Reference Seward1933) regarded the genus as a dasycladalean alga, and a number of classification schemes for Dasycladales (e.g., Bassoullet et al., Reference Bassoullet, Bernier, Deloffre, Génot, Jaffrezo and Vachard1979; Deloffre, Reference Deloffre1988; Berger and Kaever, Reference Berger and Kaever1992), following the lead of Pia (Reference Pia and Hirmer1927), tentatively included the genus within the order, regarding it as a possible member of the Seletonellaceae, a family characterized by a non-verticillate arrangement of the primary laterals. The morphology revealed by the new specimens removes any obstacle to inclusion of C. fruticosa and, by extension, Callithamnopsis within Dasycladales. Specifically, the verticillate (euspondyl) arrangement of the primary laterals is characteristic of extant dasycladalean algae, as is the branching pattern of the laterals. In addition, the apparently hollow nature of the thallus interior, as indicated by longitudinal creasing related to compaction (Ruedemann, Reference Ruedemann1909, p. 201), is consistent with the siphonous organization characteristic of dasycladalean thalli, and some living dasyclads show bifurcation of the main axis, which may be present in one of the specimens (Fig. 4.6), although it is possible that two separate thalli are preserved in close association. Because all specimens lack external reproductive structures, the genus appears to have been endosporate and, on this basis, together with the verticillate arrangement of the primary laterals, is assigned to the Family Triploporellaceae. It should be noted, however, that because nothing is known for this taxon with regard to chlorophyll composition or cellular-level organization, the possibility remains that Callithamnopsis belongs to a group of algae apart from Dasycladales for which there are no living descendants.

Figure 5. Chaetocladus dubius (Spencer, Reference Spencer1884) from the Ordovician (Katian) of Michigan and the Silurian of Ontario, and Chaetocladus plumula Whitfield, Reference Whitfield1894 from the Ordovician (Sandbian) of Wisconsin. (1–4) C. dubius from the Big Hill Formation, Delta County, Michigan; (1) detail of thallus at level of arrow in 2 showing laterals arranged in close-spaced sets of two or three verticils; (2) complete thallus, UWGM 2310; (3) upper part of thallus, UWGM 2314; (4) detail of thallus at level of arrow in 3 showing laterals arranged in close-spaced sets of two or three verticils. (5, 6) C. dubius from the Eramosa Formation of southern Ontario; (5) holotype (arrow), as identified herein, Hamilton, RM 20.4778, photograph provided by A. Howell; (6) complete thallus, Vinemount, ROMIP 64753. (7) Detail of main axis of C. plumula holotype, Platteville Formation, Platteville area, USNM P41137. Specimens in (5, 7) photographed dry; specimens in (1–4, 6) photographed immersed in alcohol. Scale bars are (1, 4) 1 mm; (2, 3, 6, 7) 5 mm; (5) 10 mm.
Callithamnopsis fruticosa (Hall, Reference Hall1865)
Figure 4.1–4.6
- Reference Hall1865
Oldhamia fruticosa Hall, p. 50.
- Reference Walcott1894
Oldhamia fruticosa; Walcott, p. 313.
- Reference Whitfield1894
Callithamnopsis fruticosa; Whitfield, p. 354, pl. 11, figs. 4–8.
- Reference Whitfield1894
Chaetomorpha? prima; Whitfield, p. 355, pl. 11, figs. 9, 10.
- Reference Whitfield1895
Callithamnopsis fruticosa; Whitfield, p. 42, pl. 4, figs. 4–8.
- Reference Whitfield1895
Chaetomorpha? prima; Whitfield, pl. 4, figs. 9, 10.
- Reference Ruedemann1909
Callithamnopsis fruticosa; Ruedemann, p. 201, text figs. 4–10, pl. 1, figs. 3, 4.
- Reference Teller1911
Callithamnopsis fruticosa; Teller, p. 186.
- Reference Pia and Hirmer1927
Callithamniopsis fruticosa; Pia, p. 67.
- Reference Johnson1961
Callithamniopsis fruticosa; Johnson, p. 39, pl. 15, figs. 1, 2.
Lectotype
USNM P 41089 (Fig. 4.4) from the Ordovician (Sandbian) Platteville Formation in the vicinity of Platteville, Wisconsin, as designated herein.
Diagnosis
Callithamnopsis with primary lateral segments 0.1–0.15 mm in width.
Occurrence
In addition to type locality, known from chocolate-brown shale partings in the lowermost part of the Pecatonica Member of the Platteville Formation at a quarry northeast of Sun Prairie along Highway 151 (43.207127°N, 89.174085°W) (Stop 2 of Simo et al., Reference Simo, Choi, Freiberg, Byers, Dott, Saylor and Mudrey1997) and a small pit (now filled) west of Columbus along County Highway K (43.327318°N, 89.147205°W).
Description
Thallus of UWGM 2364 noncalcified, comprising an undivided uniaxial main axis surrounded by numerous branched lateral appendages arranged in verticils (euspondyl) (Fig. 4.2). Main axis cylindrical, 0.2 mm wide and 10 mm long, maintaining a nearly uniform diameter, without appreciable change at or between verticils. Verticils spaced at intervals of ~1 mm, each comprising four to six primary laterals, the precise number difficult to determine owing to the flattened condition of the material (Fig. 4.1). Laterals branched to at least the third order; lateral segments cylindrical; primary lateral segments with a width of 0.10 mm, slightly expanded at base. Thallus of UWGM 2411 similar to that of 2364, but main axis 0.4 mm wide, primary lateral segments 0.15 mm wide, and verticils set at intervals of ~2 mm (Fig. 4.3). Neither specimen shows evidence of a holdfast or reproductive structures.
Materials
UWGM 2352, 2364, 2411; also, many detached laterals on slabs UWGM 2411 and 2408.
Remarks
In addition to the thalli assigned herein to C. fruticosa, slabs UWGM 2411 and 2408 are covered with detached laterals attributable to this species (Fig. 1.9), and branched filaments observed in association with Buthograptus gundersoni n. sp. at the New Glarus locality (UWGM 2396, 2400, 2404) might also represent detached laterals from this species. Ruedemann (Reference Ruedemann1909, Reference Ruedemann1925) assigned two additional species to Callithamnopsis, C. delicatula Ruedemann, Reference Ruedemann1909, from the Ordovician of eastern New York, and C. silurica Ruedemann, Reference Ruedemann1925, from the Silurian (Pridolian) of western New York. These species are poorly known and are in need of restudy, but would appear to differ significantly from C. fruticosa in terms of the widths of the primary lateral segments. Specifically, those of C. delicatula are narrower (~0.05 mm on the basis of line drawings provided in Ruedemann, Reference Ruedemann1909), whereas those of C. silurica are wider (~0.3 mm on the basis of line drawings provided in Ruedemann, Reference Ruedemann1925). The species diagnosis provided herein for C. fruticosa conveys this distinction.
In the original description of the species, Hall (Reference Hall1865) did not provide illustrations and did not designate a holotype. Later, Whitfield (Reference Whitfield1894, Reference Whitfield1895) illustrated in line drawing material used by Hall to establish the species, including two specimens among at least four thalli belonging to this species on slab P 41089, currently housed in the Smithsonian type collection. Herein, the specimen on P 41089 shown in Figure 4.4, which was illustrated by Whitfield (Reference Whitfield1894, pl. 11, fig. 6), is proposed as the lectotype.
Genus Chaetocladus Whitfield, Reference Whitfield1894
Type species
Chaetocladus plumula Whitfield, Reference Whitfield1894 from the Ordovician (Sandbian) Platteville Formation in the vicinity of Platteville, Wisconsin, by original monotypy.
Remarks
A comprehensive overview of the genus, including an examination of its phylogentic affinity and updated descriptions of all constituent species, was provided by LoDuca (Reference LoDuca1997). In the time since that report, two additional species have been assigned to Chaetocladus, C. gracilis Kenrick and Vinther, Reference Kenrick and Vinther2006, from the Ludlovian of Sweden, and C. capitatus LoDuca et al., Reference LoDuca, Melchin and Verbruggen2011, from the Ludlovian of Cornwallis Island, Nunavut, Canada. An interesting aspect of Big Hill specimens UWGM 2310 and 2314, which are formally assigned herein to Chaetocladus, is that they show the laterals arranged in “verticil sets” comprising two or three close-spaced whorls, with each set separated from the next vertically along the main axis by a gap of ~1 mm (Fig. 5.1, 5.4). Such a configuration is also known for C. capillatus and C. ruedemanni (LoDuca, Reference LoDuca1997). Notably, this arrangement of the laterals in verticil sets resembles the “biserial” arrangement of laterals known for some Mesozoic species of dasycladalean algae (Pia, Reference Pia1912), including Oligoporella serripora Pia, Reference Pia1912 and Physoporella dissita (Gümbel, Reference Gümbel1872).
Chaetocladus dubius (Spencer, Reference Spencer1884)
Figure 5.1–5.6
Holotype
RM 20.4778 from the base of the Silurian Eramosa Formation (“Barton Beds”) along Redhill Creek, 2 km west of Albion Falls, Hamilton, Ontario (Fig. 5.5).
Occurrence
Ordovician (Katian) Big Hill Formation, Delta County, Michigan, USA; Silurian (Wenlockian) Eramosa Formation in the vicinity of Guelph, Vinemount, and Hamilton, Ontario, Canada.
Description
Thallus of UWGM 2310 noncalcified, cylindrical, 53 mm long and 4.5 mm wide, comprising an undivided uniaxial main axis surrounded by numerous unbranched lateral appendages arranged in verticils (euspondyl) (Fig. 5.2). Main axis cylindrical, 0.75 mm wide and 53 mm long. Laterals cylindrical, emerging at nearly 90° to the main axis and then gently curving upward; most with a width of 0.1 mm and a length of 3 mm, but those along lowermost 8 mm of main axis conspicuously shorter; terminations abrupt, slightly rounded. Verticils comprise ~12 laterals, arranged in close-spaced sets of two or three whorls, successive verticil sets separated by intervals of ~1 mm (Fig. 5.1). Lower part of thallus with distinct right angle bend, the portion below this likely comprising the holdfast. Reproductive structures not evident. UWGM 2314 missing lower part of thallus, comparable to UWGM 2310 in all key respects, but axis slightly wider (1.0 mm) and laterals in lower part of specimen somewhat longer (up to 6 mm) (Fig. 5.3, 5.4).
Materials
UWGM 2310, 2314.
Remarks
UWGM 2310 was illustrated in Lamsdell et al. (Reference Lamsdell, LoDuca, Gunderson, Meyer and Briggs2017) and LoDuca et al. (Reference LoDuca, Bykova, Wu, Xiao and Zhao2017) as Chaetocladus sp. In having unbranched cylindrical lateral appendages arranged in verticils around an undivided uniaxial central axis, this specimen and UWGM 2314 accord in all key respects with the emended diagnosis of Chaetocladus provided by LoDuca (Reference LoDuca1997). Among the various species assigned to the genus, only C. dubius, from the Silurian of Ontario (see LoDuca, Reference LoDuca1997), is comparable to the Big Hill specimens in terms of both lateral width (0.07 mm versus 0.1 mm) and main axis width (between-specimen range of 0.7–1.5 mm versus 0.75–1.0 mm, respectively; compare Fig. 5.2 and 5.6). The main axes of all other species currently recognized within the genus, including that of the genotype, C. plumula (Fig. 5.7), are wider by a factor of two or more, and C. ruedemanni LoDuca, Reference LoDuca1997 and C. capitatus also differ by having markedly wider laterals (0.2 mm and 0.25 mm, respectively).
Both UWGM 2310 and UWGM 2314 show the laterals arranged in verticil sets comprising two or three close-spaced whorls with each set separated from the next vertically along the main axis by a gap of ~1 mm. This configuration has not been observed in specimens of C. dubius from the Eramosa Formation of Ontario, but, as noted by LoDuca (Reference LoDuca1997), central parts of these thalli are obscured by overlapping laterals and/or are poorly preserved, and it is possible that such an arrangement is present. Indeed, such an arrangement would explain the discrepancy in the number of laterals per verticil between the Big Hill specimens and specimens of C. dubius from Ontario (~12 versus a reported value of ~40). Specifically, if the latter had verticil sets comprising three close-spaced whorls, then the number of laterals per individual verticil would fall to 13, a number comparable to that observed in the Big Hill material. In addition, the presence of verticil sets in the Ontario material would account for distinct horizontal “banding” present along the main axes (LoDuca, Reference LoDuca1997) (Fig. 5.6) because the height of each dark band corresponds to the distance between the lower and upper whorl in each verticil set in the Big Hill specimens (Fig. 5.2, 5.3).
Chaetocladus dubius has been documented previously only from the Eramosa Formation of southern Ontario. Consideration of the Big Hill material as C. dubius results in a small increase in the geographic distribution of this taxon, but markedly extends its stratigraphic range, from the mid-Silurian into the Upper Ordovician (uppermost Katian).
During this study, an effort was made to locate the holotype of C. dubius. A previous attempt that relied upon catalogs of type specimens did not prove fruitful (LoDuca, Reference LoDuca1997). Spencer did not explicitly designate a holotype, but the sole illustrated specimen (in line drawing) that accompanied the original species description (Spencer, Reference Spencer1884, pl. 1, fig. 1) would qualify as such. Unfortunately, the caption for this illustration does not provide a museum number or repository. At the Redpath Museum, two slabs with the number RM 20.4778 from the C. dubius type locality bear numerous C. dubius specimens. One of these specimens appears to have been the model for Spencer's line drawing and is regarded herein as the holotype (Fig. 5.5). Although this material is housed with the type collection at the Redpath Museum, this specimen did not carry a holotype designation (A. Howell, personal communication, 2018). Notably in this regard, the Redpath Museum houses the holotype of Thallograptus grantii (Dawson, Reference Dawson1890) (see Hewitt and Birker, Reference Hewitt and Birker1986), which was indicated by Grant (Reference Grant1892) to have been collected from the same locality and unit as the C. dubius material described by Spencer (Reference Spencer1884). It stands to reason, therefore, that material used by Spencer to erect C. dubius, including the specimen illustrated with the species description, would be reposited at the Redpath Museum as well. Slabs at the Royal Ontario Museum that carry the numbers 613N and 1560N (now cataloged as ROMIP 20923 and 21731, respectively) are covered with C. dubius material and bear labels written in the same hand as RM 20.4778, but none of these specimens, which are annotated as “referred specimens” in the museum catalog (M. Akrami, personal communication, 2018), is a close match for Spencer's line drawing. The same applies to C. dubius specimens on six slabs in the Smithsonian collection that carry the number PAL 102335, one of which has a label written in the same hand as RM 20.4778, along with a second label stating “Grant Collection.” The Smithsonian database lists the specimens on these slabs as hypotype material, and figure captions that accompany examples of these specimens illustrated in Ruedemann (Reference Ruedemann1947) describe the specimens as topotypes.
Chaetocladus capitatus LoDuca, Melchin, and Verbruggen, Reference LoDuca, Melchin and Verbruggen2011
Figure 4.7–4.10
Holotype
ROM 59925A, from the Silurian (Ludfordian) Cape Phillips Formation along an unnamed stream ~5 km southeast of Cape Manning on northeastern Cornwallis Island, Nunavut, Canada (LoDuca et al., Reference LoDuca, Melchin and Verbruggen2011, fig. 4.1).
Occurrence
Ordovician (Katian) Big Hill Formation, Delta County, Michigan, USA; Silurian (Ludlovian) Cape Phillips Formation, Cornwallis Island, Nunavut, Canada.
Description
Thallus of UWGM 2312 noncalcified, comprising numerous elongate undivided cylindrical elements ~0.2 mm wide and up to 12 mm long, these converging at the base and arranged to form a hemispherical distribution (Fig. 4.7). Thalli on UWGM 2311 similar (Fig. 4.8, 4.9), but some show a ring-shaped structure at the center, ~3 mm in diameter, from which the elongate cylindrical elements arise and are arranged in a radial pattern (Fig. 4.10). Reproductive structures and holdfast not evident.
Materials
UWGM 2311 with at least five specimens; UWGM 2312 with one specimen.
Remarks
Specimens on UWGM 2311 provide the key to understanding the affinity of this interesting alga because some show the elongate cylindrical elements originating from a ring-shaped structure at the center. Such a form, including the central structure, is comparable to that observed in specimens of Medusaegraptus mirabilis Ruedemann, Reference Ruedemann1925 and M. graminiformis (Pohlman, Reference Pohlman1886) buried upright, in-situ, in the Silurian of New York (LoDuca, Reference LoDuca1990, fig. 3.7, 3.10; Ruedemann, Reference Ruedemann1925, pl. 8, fig. 3), where the central structure is the main axis in cross section. The main axes of Chaetocladus ruedemanni are also known to give ring-shaped cross-sections (LoDuca, Reference LoDuca1997, fig. 5.12). Thus, these circular tufts are interpreted as the capitulum of an alga with an architecture similar to Medusaegraptus or Chaetocladus, for which the thallus was buried erect, in life position, and the globose capitulum was flattened by a rapid influx of sediment. In such instances, exposure of upper parts of thalli during collection results from selective breakage of the otherwise homogeneous fine-grained matrix at the level of the large flattened capitulum. Other specimens on UWGM 2311, together with UWGM 2312, are interpreted to have been preserved in similar fashion, but with the capitulum flopped to one side during the burial event, and so preserved in lateral view. Interpreted in this way, slab UWGM 2311 shows a small stand of thalli buried upright, in life position.
Among the handful of available specimens, none is known that shows the main axis in lateral view. This is significant because the principal distinction between Chaetocladus and Medusaegraptus concerns the manner in which the laterals are arranged around the main axis (euspondyl and aspondyl, respectively). Nonetheless, a good argument can be made for assigning the Big Hill material to Chaetocladus capitatus on the basis of capitulum morphology. In this regard, the overall form of the capitulum in lateral view and the dimensions of the constituent laterals are indistinguishable from C. capitatus, but differ markedly from material of Medusaegraptus. The diameter of the main axis, which is wider by a factor of three than that of associated specimens of C. dubius, also corresponds to that of C. capitatus.
In Lamsdell et al. (Reference Lamsdell, LoDuca, Gunderson, Meyer and Briggs2017) one of the specimens on UWGM 2311 was illustrated as a “tuft-like alga” and in LoDuca et al. (Reference LoDuca, Bykova, Wu, Xiao and Zhao2017) this specimen was regarded as a member of the delicately dichotomously branched morphogroup. On the basis of the new observations and interpretations presented above, all specimens on UWGM 2311 as well as UWGM 2312 are better regarded as belonging to the simple monopodial morphogroup of LoDuca et al. (Reference LoDuca, Bykova, Wu, Xiao and Zhao2017).
Chaetocladus capitatus previously had been documented only from the type locality. Consideration of the Big Hill material as C. capitatus extends the stratigraphic range of this taxon from the Upper Silurian (Ludfordian) into the Upper Ordovician and extends the geographic distribution from the type locality in Arctic Canada (Cornwallis Island, Nunavut) into northern Michigan.
Acknowledgments
G. Gunderson and R. Meyer collected much of the material upon which this study is based and their keen observations in the field proved invaluable when constructing the manuscript. B. Datillo and R. Plotnick contributed Buthograptus material from the Dickeyville locality. Assistance with museum collections was provided by C. Eaton at the University of Wisconsin Geology Museum, by M. Florence, J. Wingerath, and G. Chavez at the Smithsonian Institution, by V. Millien and A. Howell at the Redpath Museum, and by J.-B. Caron and M. Akrami at the Royal Ontario Museum. G. Moore, W. Bender, and O. Neill provided assistance with the SEM. The final manuscript was improved by comments provided by reviewers I. Bucur and H. Nowak, the associate editor, R. Lupia, and the editor, E. Currano. Research was supported in part by National Science Foundation grant EAR 1250756, awarded to the author.







