Introduction
Millipedes are a widely distributed group of terrestrial arthropods (Shelley and Golovatch, Reference Shelley and Golovach2011; Enghoff, Reference Enghoff2015; Minelli, Reference Minelli2015). But, because of their terrestrial habitus, millipedes are rarely preserved as fossils. Fossil millipedes are mainly documented from the Paleozoic because they are relatively abundant in the extensive coal-forest deposits of the upper Carboniferous; they are also well documented in amber deposits, particularly those of the Cenozoic. Only rarely have they been described from the Mesozoic (Dzik, Reference Dzik1981; Jell, Reference Jell1983; Duy-Jacquemin and Azar, Reference Duy-Jacquemin and Azar2004; Rasnitsyn and Golovatch, Reference Rasnitsyn and Golovatch2004; Shear et al., Reference Shear, Selden and Gall2009; Shear and Edgecombe, Reference Shear and Edgecombe2010; Edgecombe, Reference Edgecombe2015; Liu et al., Reference Liu, Rühr and Wesener.2017). Mesozoic occurrences include two described genera from the Triassic (France and central Siberia), one genus from the Jurassic of Australia, and six genera from the Cretaceous (Mexico, Europe, Lebanon, Mongolia, and Myanmar). Additional undescribed or poorly preserved millipede material is known from the Triassic of Africa and the Cretaceous of Europe.
Asian Mesozoic occurrences are from the Cretaceous of Mongolia (Dzik, Reference Dzik1975), Myanmar (Cockerell, Reference Cockerell1917; Grimaldi et al., Reference Grimaldi, Engel and Nascimbene2002; Rasnitsyn and Golovatch, Reference Rasnitsyn and Golovatch2004; Carlson, Reference Carlson2007; Liu et al., Reference Liu, Rühr and Wesener.2017), China (Huang, Reference Huang2015, Reference Huang2016), and the Triassic of Siberia (Dzik, Reference Dzik1981) and China (Hu et al., Reference Hu, Zhang, Chen, Zhou, Lu, Xie, Wen, Huang and Benton2011). Fossil millipedes have been noted from three Mesozoic localities in China: Yanliao District (the famous fossil localities of Daohugou Biota) in Liaoning Province, Inner Mongolia, and Luoping County in Yunnan Province. Jurassic myriapods (including millipedes) have been noted from Daohugou (Huang, Reference Huang2015, Reference Huang2016), but they have been neither illustrated nor described, also a mid-Jurassic millipede is known from Inner Mongolia (D.Y. Huang, personal communication, 2017), but is yet to be illustrated or described. Hu et al. (Reference Hu, Zhang, Chen, Zhou, Lu, Xie, Wen, Huang and Benton2011, p. 2279, fig. 5j) have noted the presence of millipedes in the Luoping fauna, illustrating one of those specimens.
The purpose of this paper is to describe the specimen illustrated by Hu et al. (Reference Hu, Zhang, Chen, Zhou, Lu, Xie, Wen, Huang and Benton2011, p. 2279, fig. 5), to briefly document another specimen with possible millipede affinity in the Luoping biota, and to provide brief comments on other Mesozoic millipedes.
The biota and its geological setting
The Luoping fossil Lagerstätte is located in Luoping County, Yunnan Province, southwest China (Fig. 1). This is in the southwestern part of the Yangtze Platform between the Nanpanjiang Basin and the Yangtze Platform (Enos et al., Reference Enos, Lehrmann, Wei, Yu, Xiao, Chaikin, Minzoni, Berry and Miontgomery2006). The Luoping biota is part of Member II of the Guanling Formation, of the Anisian stage (Middle Triassic), based upon conodont zonation (Huang et al., Reference Huang, Zhang, Zhang, Lu, Zhou and Bai2009; Zhang et al., Reference Zhang, Zhou, Lu, Xie, Lou, Liu, Sun, Huang and Zhao2009). The biota comprises a mixture of marine and terrestrial organisms (Hu et al., Reference Hu, Zhang, Chen, Zhou, Lu, Xie, Wen, Huang and Benton2011) deposited in a marine environment. It includes typical marine reptiles, fishes, echinoderms (crinoids, sea urchins, sea cucumbers, and sea stars), bivalves, gastropods, belemnoids, ammonoids, brachiopods, conodonts, foraminiferans, and marine and nearshore arthropods. Among the thousands of fossils from the Luoping biota, arthropods, including lobsters, shrimp, mysidaceans, isopods, cycloids, conchostracans, ostracods, horseshoe crabs, and millipedes, comprise more than 90% of the biota (Hu et al., Reference Hu, Zhang, Chen, Zhou, Lu, Xie, Wen, Huang and Benton2011, p. 2278, fig. 4).

Figure 1 Location map and stratigraphic occurrence of the millipede horizon and other fossils in Luoping County of east Yunnan Province, southwest China. Occurrence and range of the conodont Nicoraella kockeli (Tatge, Reference Tatge1956a, Reference Tatgeb) is indicated by the black boxes and line. Numbered fossils, from 1 to 17, are: 1, marine vertebrates; 2, ichthyosaurs; 3, fishes; 4, crustaceans and limulids; 5, sea stars; 6, holothurian sclerites; 7, crinoids; 8, bivalves; 9, gastropods; 10, brachiopods; 11, ammonites; 12, other cephalopods; 13, foraminiferans; 14, ostracodes; 15, conodonts; 16, plant fragments; 17, marine vertebrate fragments.
Sediments of the Luoping biota comprise five units (Fig. 1). The lower thick-bedded unit (bed 0) consists of muddy and dolomitic limestone, mainly containing bivalves and gastropods. This is overlain by the lower thin-bedded unit (beds 1–68), which consists of laminar micritic limestone, containing abundant fossil fish, marine reptiles, arthropods, echinoderms, plants, etc., in rocks bearing siliceous concretions and bentonite layers. Above this lies the middle thick-bedded unit (beds 69–74), consisting of strongly bioturbated bioclastic limestone, with marine reptiles, bivalves, and gastropods. The upper thin-bedded sequence (beds 75–145) consists of laminar argillaceous limestones, containing abundant fossil fish, marine reptiles, and arthropods (Hu et al., Reference Hu, Zhang, Chen, Zhou, Lu, Xie, Wen, Huang and Benton2011). These beds typically exhibit sedimentary characters of slump structures, indicating soft-sediment deformation. The upper thick-bedded sequence (bed 146) consists of thick massive limestone beds with bivalves and gastropod fragments, and rare marine reptile remains.
Materials and methods
Materials
The main specimen described (LPI-61593) is preserved in mostly dorsal view, somewhat rotated to the left, providing lateral views of sternites and legs on some sections of the right side of the millipede. This has also resulted in the left-hand side of the millipede being folded over to a small degree. The body is arrayed in a loose, sinuous, relaxed pattern. The specimen has been compressed, with lines of breakage differing on different portions of the body. Some segments are slightly offset at breaks. The mostly dorsal attitude of preservation may be due to the millipede being flattened, or simply due to the lack of strong coiling. Strong coiling, even if partial, results in preservation of fossil millipedes in lateral view. Specimen LPI-63009 has 21 segments, most of which are similar in length. It lacks diagnostic features, but is described here for the sake of completeness.
Methods
The specimens were photographed with a Leica M125 stereo-microscope using a Leica DFC295 digital camera in the Chengdu Center of China Geological Survey (CDCGS), Chengdu, China, and photographed in more detail by a VH-Z20R digital camera with the Keyence three-dimensional imaging system in the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) of Beijing. Micro-ornamentation was scanned by scanning electron microscope (SEM) using the Zeiss EVO-MA25 in IVPP.
Repository and institutional abbreviation
Specimens are deposited at the Chengdu Center of China Geological Survey (CDCGS), (LPI=Luoping invertebrates).
Systematic paleontology
Class Diplopoda Blainville in Gervais, Reference Gervais1844
Subclass Helminthomorpha Pocock, Reference Pocock1887
Superorder, Order, and Family incertae sedis
Remarks
Characters present do not allow confident assignment to any extant or extinct order of millipedes, but the segment number (39), ring structure (sternites not fused to the pleurotergites), large head, simple collum that does not cover the head, and presence of a pair of pits on the lateral swellings of the metazonites that may have been insertion points for setae, suggest a nematophoran affinity, although a colobognath affinity cannot be ruled out. A distinct midline is lacking however; therefore this taxon is not assigned to the Nematophora. A nematophoran, specifically calipodidan, affinity has been suggested for Hannibaliulus wilsoniae Shear, Selden, and Gall, Reference Shear, Selden and Gall2009 (Shear et al., Reference Shear, Selden and Gall2009; Edgecombe, Reference Edgecombe2015, p. 347), which is a form that bears some resemblance to Sinosoma n. gen.
Genus Sinosoma Huang and Hannibal new genus
Type species
Sinosoma luopingense new genus new species, by original designation and monotypy.
Diagnosis
Millipedes with 39 body segments, including a small collum. Metazonites divided by grooves into three transverse parts that are flanked laterally by narrow, suboval swellings bearing a pair of posterolateral pits (?spine bases). Sternites free. Legs of medium length, stout.
Etymology
The generic name is derived from the stem Sino-, meaning “pertaining to China,” in reference to the country where the specimen was collected. The ending -soma (Greek for body [soma, neuter]) is a common ending for millipedes that has the advantage of not implying membership in a particular millipede order.
Occurrence
The upper thin-bedded unit of the Luoping biota, Member II of the Guanling Formation, Anisian, Middle Triassic. Luoping County, Yunnan Province, southwest China.
Remarks
Sinosoma n. gen. is similar to Hannibaliulus Shear, Selden, and Gall, 2009, which also has an elongate body, a large number of the body segments, lateral features that might be interpreted as swellings, and a small collum that does not cover its head. Segment number differs (39 versus up to 44) as well as the number of transverse grooves on the metazonites (two versus one). Sinosoma n. gen. also bears a pair of distinctive posterolateral pits on its swellings, a feature that Hannibaliulus lacks.
Sinosoma luopingense Huang and Hannibal new species
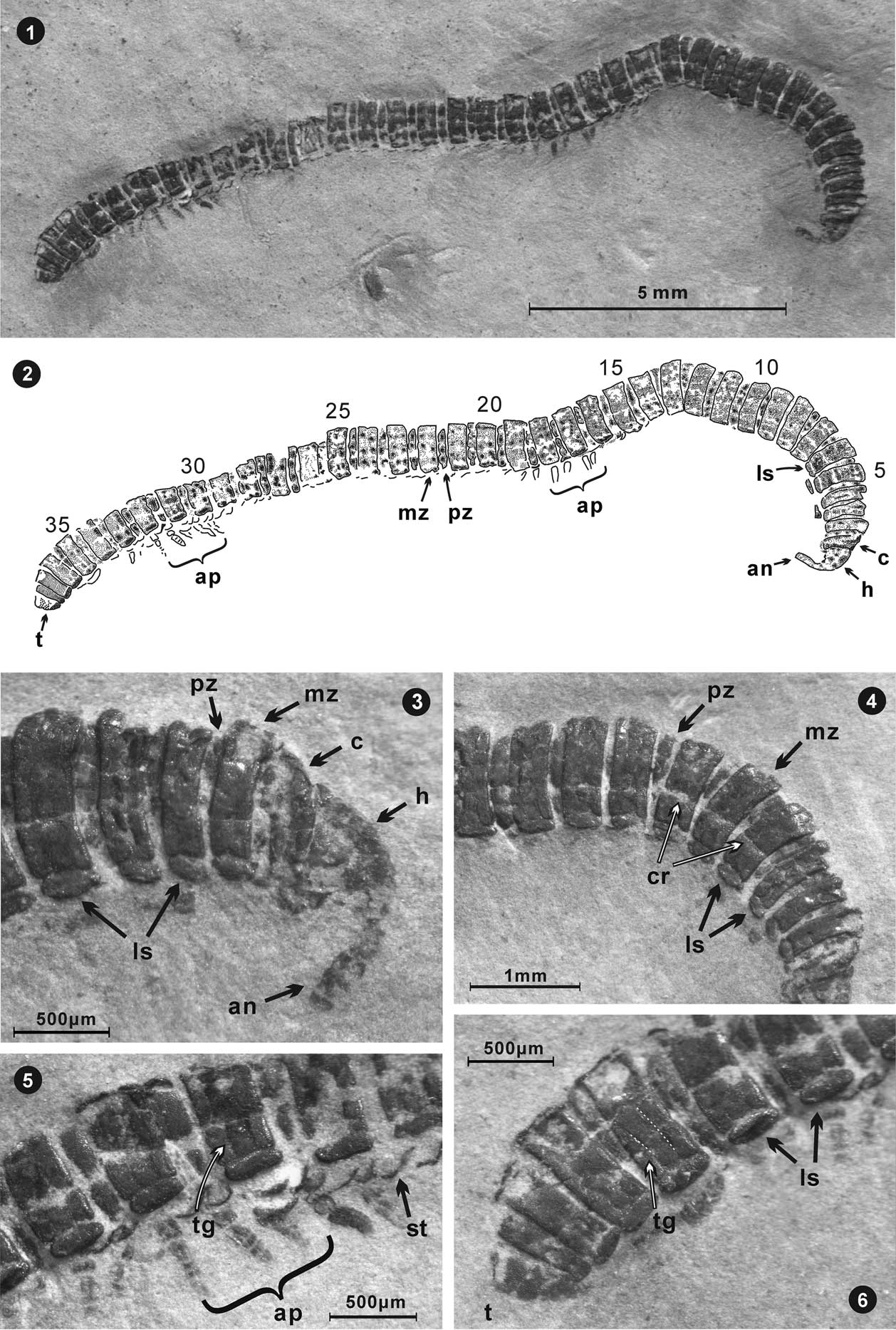
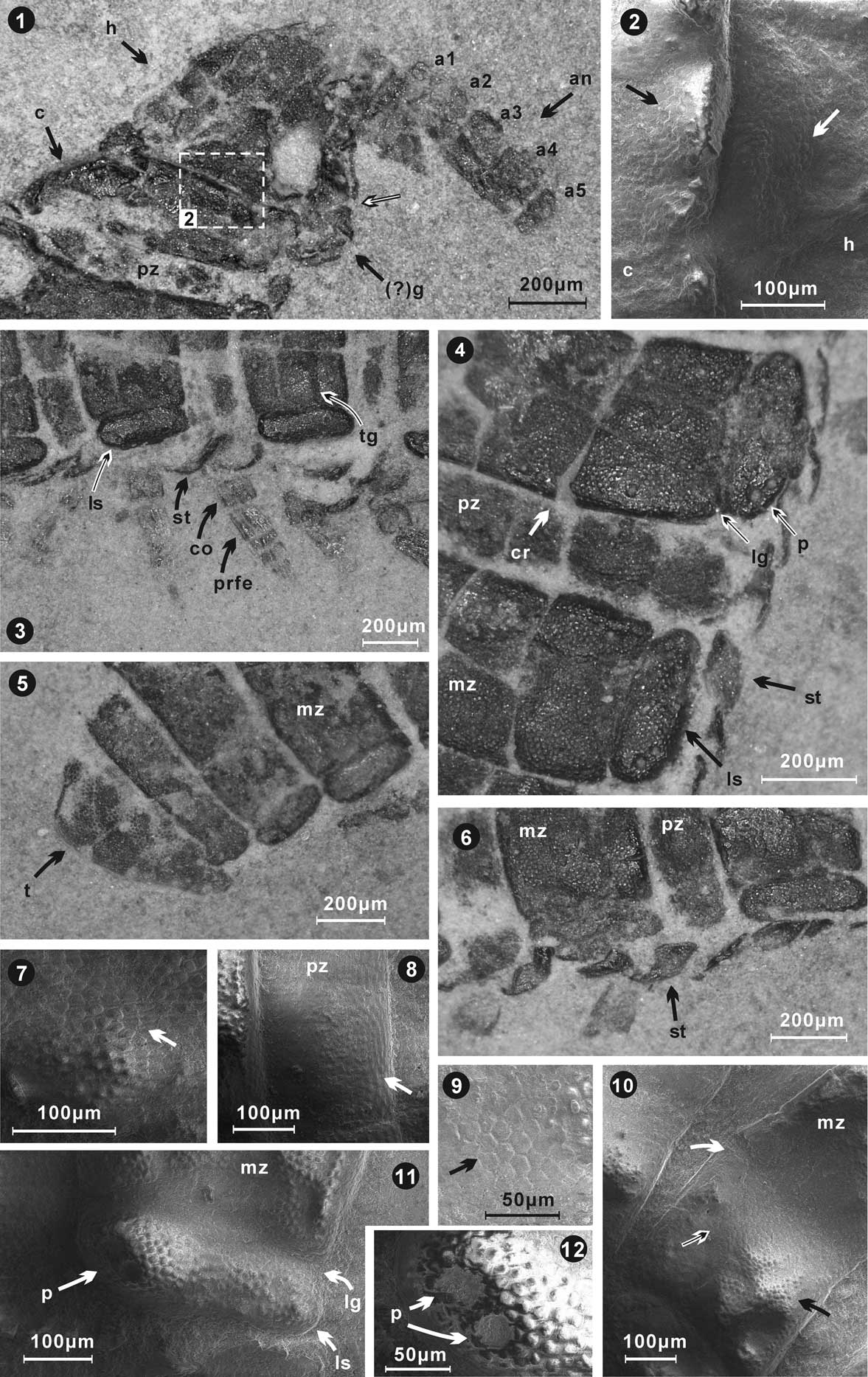
Figure 2 Sinosoma luopingense n. gen. n. sp., holotype, from the Middle Triassic Guanling Formation. Specimen LPI-61593, mostly in dorsal view. (1) Entire millipede after preparation; (2) explanatory drawing of holotype; (3) anterior, showing the antenna, head, collum, metazonite, prozonite, and lateral swellings (?paranota); (4) segments toward anterior, including what may be sutures, but are more likely cracks; (5) segments toward posterior with preserved appendages, free sternites below lateral swellings (?paranota), and a transverse groove identified; (6) posterior of millipede, with telson, lateral swellings (?paranota), and transverse grooves on a pleurotergite identified. (an=antenna, ap=appendages, c=collum, cr=crack, ls=lateral swelling (?paranotum), h=head, mz=metazonite, pz=prozonite, st=sternite, t=telson, tg=transverse groove).

Figure 3 Details of anatomical structures, micro-ornamentation, and appendages of the millipede Sinosoma luopingense n. gen. n. sp., LPI-61593, holotype. (1, 2) Anterior, showing head, collum (whose upper margin is flexed cephalad), short prozonite of the segment following the collum, antennae with five visible antennomeres, (?)gnathocliarium, numerous fine granules on collum, and head (enlarged in 2; black and white arrows point to area delineated by box in 1); (3) lateral view of pleurotergites and sternites (not fused to pleurotergites), and legs; (4, 6) lateral view of pleurotergites and sternites, including (4) two posterolateral pits on lateral swellings and small granules on the prozonite and metazonite surfaces, prominent cracks, lateral swellings (?paranota) separated from remainder of paranota by prominent longitudinal grooves; (5, 7) posterior, showing relatively larger granules on the telson than on other trunk rings; (7) enlarged granular ornament on preanal ring surface; (8–10) enlarged views of the metazonite and prozonite, granules on prozonite are relatively smaller than those on the metazonite; cytoscutes (cuticular platelets) (9) polygonal, five-to-six sided, ~10 µ wide; 9 is enlarged from area pointed out by white/black arrows in 10; (10) cracks and depression (white arrow pointed in 4 and 10); (11, 12) enlarged view of lateral swelling (?paranotum) on the metazonite, showing two pits (?spine bases) and granular ornamentation. a1=antennomere1, an=antenna, c=collum, co=coxa, cr=crack, (?)g=(?)gnathocliarium, h=head, lg=longitudinal groove, ls=lateral swelling (?lateral paranotum/paranota), mz=metazonite, p=pits, prfe=prefemur, pz=prozonite, st=sternite, t=telson, tg=transverse groove. 2, 7–12 are scanning electron microscopy images.
Holotype
Specimen LPI-61593.
Diagnosis
As for genus.
Occurrence
The upper thin-bedded unit of the Luoping biota, Member II of the Guanling Formation, Anisian, Middle Triassic. Luoping County, Yunnan Province, southwest China.
Description
Medium-sized millipede, ~19 mm long, elongate with 39 segments, inclusive of collum and telson (epiproct). Most segments more-or-less uniform in width, widest midbody segments ~1 mm wide as preserved, reconstructed width slightly more than 1 mm; first few body segments at anterior tapering anteriad, last few segments tapering posteriad. Prozonites well exposed, exposed length up to 0.18 mm, metazonites up to 0.38 mm long. Ratio of exposed length of prozonites (excluding anteriormost and posteriormost segments) to length of metazonites ranging from ~34% to 50%, indicating that prozonites were probably well exposed in life (although not quite as exposed in the fossil, since the specimen is relaxed). Penultimate segment reduced in length.
Head medium sized, triangular in shape as preserved, but difficult to interpret because of twisting from dorsal position and crushing (oval missing area on right side is the result of preparation). Parts of gnathocliarium(?) exposed below head, slightly displaced. Antennae swollen, club-shaped, at least five antennomers preserved on ?right antenna (Fig. 3.1), the third and fourth of which are the largest. Collum small, not covering head, quadrangular, slightly smaller than metazonite of following segment. Prozonites well exposed, depressed, with granular ornamentation. Metazonites raised, divided into three subequal parts by two transverse grooves.
Indication of midline lacking (longitudinal cracks on dorsal surface are not aligned along midline).
Longitudinal groove divides short lateral swellings (?paranota) from remainder of pleurotergite dorsally. Anteriormost and posteriormost part of articulation of swellings with remainder of pleurotergite somewhat invaginated. Lateral swellings extend short distance from sides of pleurotergites. Swellings elongately suboval, covered with small tubercles. Pair of prominent pits (?insertion points for setae), located on posterolateral corner of dorsal surface of swellings. Pits present at least on segments 7, 8, 15, 18, 19, 20, 21, 24, 28, 29, and 31. Pits in each pair obliquely oriented on an irregular raised area. Ozopores apparently lacking. Epiproct (telson) small, longer than segment preceding it, but subequal in length to segment anterior of that segment. Posterior termination of epiproct smoothly rounded, lacking any projections. Surface coarsely granulate.
Anal valves not seen (covered by epiproct). Surface of prozonites and metazonites bearing very small granules, with relatively larger granules on preanal segment.
Sternites incompletely seen, but disarticulated from pleurotergites, indicating that they were separate in life.
Legs stout, length subequal to the length of body segments (prozonite plus metazonite). Coxae and second segment (presumed prefemur) wide, prefemur longest, with remainder of segments decreasing in length. Two segments preceding epiproct apodous.
Etymology
The trivial species name is an adjective denoting the location from which the specimen was collected. The gender is neutral.
Remarks
The head area is difficult to interpret (Fig. 3.1). It is compressed and there has been some displacement due to compression. Preparation has also removed parts of the integument. There is a possibility that the two antennae may overlap somewhat.
The lateral swellings described here as possible paranota could also be conceivably interpreted as ventral portions of pleurotergites, or even as pleurites. Such an interpretation would be unlikely, however, because pleurites and ventrolateral parts of pleurotergites of fossil millipedes are typically seen only when specimens are preserved in ventral or lateral view (e.g., Wilson and Hannibal, Reference Wilson and Hannibal2005, figs. 2, 3, 5, 6). Furthermore, there is no breakage and displacement along the groove that divides the swellings from the pleurotergites in Sinosoma n. gen. as there is in Pleurojulus. Such breakage has contributed to a longstanding discussion of the nature of the pleurites of Pleurojulus, although Wilson and Hannibal (Reference Wilson and Hannibal2005) have provided detailed evidence for there being separate pleurites in that taxon. To complicate this picture, paranota have also been confused with pleurites, as with the paranota of “Pleurojulus steuri” Schneider and Werneberg, Reference Schneider and Werneburg1998 described as pleurites (Wilson and Hannibal, Reference Wilson and Hannibal2005, p. 1106).
Paranota as interpreted here also could be interpreted as the ventral portion of the pleurotergites because what appears to be a border is present on the lateral side of some metazonites. The inflation of the lateral part of the tergites (that is what are described as swellings here), however, argues against such an interpretation.
The paired pits on the posterolateral corners of the lateral swellings of the metazonites are here interpreted as spine bases, in part because of being paired. They do not show any internal structure suggestive of being ozopores. There is a history of confusion between ozopores and spine bases in fossil millipedes, and ozopores have been incorrectly described as spine bases in both fossil euphoberiids and palaeosomatids (Hannibal and Krzemiński, Reference Hannibal and Krzemiński2005, p. 209). Still, it would be unusual for ozopores to be paired, as are the pits of Sinosoma n. gen.
Comparisons to other taxa
This specimen is similar in a number of ways to the Triassic form Hannibaliulus wilsonae, from the Grès à Voltzia (Anisian) of eastern France, which Shear et al. (Reference Shear, Selden and Gall2009) assigned tentatively to the Callipodida. Edgecombe (Reference Edgecombe2015, p. 347) subsequently noted the lack of features that would confirm assignment to that order with certainty. Hannibaliulus wilsonae has up to 44 segments, a small collum, and probably free sternites. The epiproct of H. wilsonae is also similar in shape to, but shorter than, that of Sinosoma n. gen. Shear et al. (Reference Shear, Selden and Gall2009) also noted a general resemblance of H. wilsonae to the Paleozoic genus Pleurojulus. This is in great part due to the presence of ?crushed ventral flanges on the former, which resemble the free pleurites of the latter.
Division of the dorsal surface of the pleurotergites into transverse sections is not unusual and is found in a number of unrelated taxa, including the archipolypod Palaeodesmus tuberculata Wilson and Anderson, Reference Wilson and Anderson2004, and extant polydesmid Polydesmus, which have transverse furrows. In both of those taxa, however, the surface of the metazonite is also subdivided by longitudinal furrows. A single transverse dorsal furrow crosses the pleurotergite of H. wilsonae.
The body ring of the Luoping millipede is most like that of callipodids and chordeumatids in that the sterna are not fused to the pleurotergite. Segment number of the Luoping millipede (39) is within the range of extant callipodids (which have 36 to 89 segments including the telson) and is somewhat greater than that of chordeumatids (which have ~26–32 segments including the telson). Tapering of the anterior and, especially, posterior is like that of chordeumatids. In the presence of spines on the swellings (as interpreted here), Sinosoma luopingense n. gen. n. sp. is more like a chordeumatid (see for example Shear, Reference Shear1977, fig. 1). The relatively large head, especially compared to the collum, is also like a chordeumatid. The new species differs from chordeumatids, however, in its lack of a prominent midline.
Segment number of the new millipede is very close to that of palaeosomatid archipolypods, which have up to 38 segments (Hannibal and Krzemiński, Reference Hannibal and Krzemiński2005, p. 208). The pleurotergites of palaeosomatids have ozopores born on lateral bosses, however.
Class ?Diplopoda
Order, family, genus, and species incertae sedis

Description
Segmented organism with 21 segments preserved (Specimen LPI-63009). Segments ~1 mm long. Margin(s) rimmed. Larger segment at one end.
Remarks
This specimen can conceivably be a fossil millipede because it consists of a series of similar segments. Details of its segments are difficult to interpret, however, and it lacks sufficient features to be sure of an assignment to the Diplopoda. These segments are in the same size range, but are simpler, than those of Sinosoma n. gen.
Mesozoic millipedes
Mesozoic millipedes have been described, noted, and/or illustrated in a number of publications, beginning in the mid 1850s. These reported occurrences are reviewed below to put the occurrence of Sinosoma n. gen. into context. This brief review updates the reviews of Mesozoic millipedes in Shear et al. (Reference Shear, Selden and Gall2009), Shear and Edgecombe (Reference Shear and Edgecombe2010), and Edgecombe (Reference Edgecombe2015).
A number of Triassic millipedes have been previously noted, although only two have been named and described in detail. These are Tomiulus angulatus Martynov, Reference Martynov1936, and Hannibaliulus wilsonae Shear et al., Reference Shear, Selden and Gall2009. Tomiulus angulatus, found in Siberia, is a julimorph-like millipede. It was diagnosed by Sharov (Reference Sharov1962), based on Martynov (Reference Martynov1936), and subsequently redescribed by Dzik (Reference Dzik1981), who assigned it to the family Xyloiulidae Cook, Reference Cook1895. This specimen is preserved in lateral view, with only the pleurotergites preserved. Reexamination of the holotype at the Paleontological Institute Moscow shows that its pleurotergites are terminated ventrally by a distinct, rounded rim (JTH, personal observation, 1998), a feature indicated in Sharov’s (Reference Sharov1962, p. 23) diagnosis (see translation by Dzik, Reference Dzik1981, p. 397). This implies that the sternites were not fused to the pleurotergites. Ozopores appear to be lacking. The lack of preserved sternites in the holotype of Tomiulus also indicates that the sternites and pleurotergites were unfused. Furthermore, it is likely that the sternites were narrow. The Xyloiulidea, as diagnosed by Wilson (Reference Wilson2006, p. 639), include elongate, cylindrical millipedes with ozopores, short legs, and ornamentation composed of longitudinal striations ranging from parallel to slightly sinusoidal in orientation. As Shear et al. (Reference Shear, Selden and Gall2009) have pointed out, only the lattermost character is present in Tomiulus, so its identity as a xyloiulid cannot be confirmed, although it is certainly likely that this form was also elongate, cylindrical, and juliform-like in general configuration.
Additional millipedes have, however, been noted and illustrated, but not formally described or named, from Triassic rocks. These include juliform millipedes from the Lower Triassic of Bethulie and Bergville, South Africa, noted several decades ago by Kitching (Reference Kitching1977, p. 9) and Lawrence (Reference Lawrence1984, p. 134). Lawrence described them as gregarious juvenile juliform millipedes. Reisz and Laurin (Reference Reisz and Laurin1991, fig. 1) identified similar forms found in association with the skeletons of the procolophonid Owenetta as millipede-like arthropods, and interpreted these arthropods as scavengers. Subsequently, Groenewald and Kitching (Reference Groenewald and Kitching1995, p. 37) listed millipedes comparable to extant Gymnostreptus Brölemann, Reference Brölemann1902 (a spirostreptid in the superorder Juliformia) as occurring in the Lystrosaurus Assemblage Zone of the Beaufort beds (earliest Triassic) of South Africa. More recent mentions include those of Reisz and Scott (Reference Reisz and Scott2002, fig. 1), who again illustrated what appear to be juliform millipedes. These same millipedes were subsequently illustrated in color by MacRae (Reference MacRae1999, p. 195). Retallack et al. (Reference Retallack, Smith and Ward2003, p. 1142) noted that the specimens previously reported from Bethulie were comparable to extant Gymnostreptus, a spirostreptid. More recently, Abdala et al. (Reference Abdala, Cisneros and Smith2006, p. 507, fig. 3D) noted and illustrated what they identified as a probable juliform millipede preserved with tetrapods in a carbonate concretion (collected by B.J. Kitching) from the Bethulie area. They did not observe any limbs on the millipede, which they interpreted as sharing a burrow with Owenetta (Abdala et al., Reference Abdala, Cisneros and Smith2006, p. 511). Based on the published comments and the published illustrations of these forms, it is probably safe to state that these are juliform-like millipedes.
A supposed millipede body-fossil impression has been described from the Triassic of Utah (Mickelson et al., Reference Mickelson, Huntoon and Kvale2006), but Lerner et al. (Reference Lerner, Lucas and Spielmann2007) have disputed its identity as a millipede body fossil.
This leaves two groups of Triassic millipedes: juliform-like millipedes and millipedes that are at least similar to nematophorans.
The Jurassic record consists of the enigmatic form Decorotergum warrenae Jell, Reference Jell1983. This form, which is rounded in cross-section with clear prozonites and metazonites, has been accepted by recent authors as a chilognath millipede, although without support of an original assignment as an oniscomorph or a polydesmid (Shear et al., Reference Shear, Selden and Gall2009; Edgecombe, Reference Edgecombe2015). Even its identity as a millipede is in need of confirmation; some of the ventral terminations of the pleurotergites appear to be arched dorsally. There are a number of examples of misidentification of millipedes as other taxa, and vice versa (see Hannibal, Reference Hannibal2001).
Cretaceous forms, however, have a decidedly modern look to them. These include polyxenids (Duy-Jacquemin and Azar, Reference Duy-Jacquemin and Azar2004; Rasnitsyn and Golovatch, Reference Rasnitsyn and Golovatch2004), colobognaths (Wilson, Reference Wilson2006, fig. 4; Carlson, Reference Carlson2007), spirobolids (Dzik, Reference Dzik1975), siphoniulids (Liu et al., Reference Liu, Rühr and Wesener.2017), and a polydesmid (Wilson, Reference Wilson2006, fig. 4). Of these, only the polyxenids, spirobolids, and siphoniulids have been well studied. The polyxenids (three genera) have been referred to families that include extant forms. The colobognaths, one noted as a polyzoniid with siphonophorid characters, have not been described in detail. Gobiulus Dzik, Reference Dzik1975, is a spirobolid that Shelley and Floyd (Reference Shelley and Floyd2014, p. 24–25; but see also Shear et al., Reference Shear, Selden and Gall2009, p. 10–11) assigned to an extant subfamily. Polydesmids were first noted as occurring in the Cretaceous by Wilson (Reference Wilson2006), and subsequently illustrated without descriptions from Myanmar (Xia et al., Reference Xia, Yang, Zhang, Shi and Wang2015, p. 151). The siphoniulids, including two new species of genus Siphoniulus from Myanmar (Liu et al., Reference Liu, Rühr and Wesener.2017), have morphology similar to that of recent species in Central America, and this discovery satisfies the viewpoint of the Siphoniulida as a “declining” order (Golovatch, Reference Golovatch2015) or living relic (Shelley and Golovatch, Reference Shelley and Golovach2011). Recently, however, a new Cretaceous millipede fauna has been noted by Poyato-Ariza and Buscalioni (Reference Poyato-Ariza and Buscalioni2016), and Selden and Shear (Reference Selden and Shear2016) assigned the millipedes from this fauna tentatively to the superfamily Xyloiuloidea, which does not include extant taxa.
Fritsch (Reference Fritsch1910, p. 6–7, pl. 4, figs. 9–13) also described Cretaceous myriapods from Bohemia, including specimens he identified as a (?)glomerid and a julid. He did not name these taxa, and these specimens are poorly preserved and difficult to interpret. Of these, the specimen of the supposed glomerid is the most millipede-like. These Cretaceous specimens are more poorly preserved than the specimens he described from the Gaskohle of Bohemia, and Fritsch’s illustrations of the Czech Cretaceous material are not as accurate as are those of the material of the Gaskohle, which, in turn, are not very accurate (JTH, personal observation, 1984, 1993). Based on the original description and illustrations, the species ?Xylobius mexicanus Mullerried, Reference Mullerried1942 from the Upper Cretaceous of Mexico, may be an authentic millipede. The specimen is missing however, so its identity cannot be confirmed. Excluded from consideration here is Julopsois cretacea Heer, Reference Heer1874, a Cretaceous form interpreted as a julid by Scudder (Reference Scudder1886, p. 18), which Hoffman (Reference Hoffman1969, p. R605) excluded from the Diplopoda. Also excluded is Calciphilus, a Cenozoic millipede that a number of authors, including Hoffman (Reference Hoffman1969, p. R604) correctly included in the Diplopoda, but erroneously listed as being Cretaceous (see McKee [Reference McKee1946] for a description of the deposit).
Based on the sparse fossil record of the Mesozoic, it appears that is was only in the Cretaceous that the Diplopoda took on a modern aspect.
Paleoecological implications
Millipedes, along with conifers, are clear terrestrial components of the predominantly marine Luoping biota. Based on the preservation of conifer remains, Hu et al. (Reference Hu, Zhang, Chen, Zhou, Lu, Xie, Wen, Huang and Benton2011, p. 2278) hypothesized that conifers were transported ~10 km into the Luoping Basin. Comparisons can be made with other biotas that have mixed marine and terrestrial components. Fossil millipedes are a small component, for instance, of the Essex fauna of Mazon Creek (Baird and Anderson, Reference Baird and Anderson1997; Hannibal, Reference Hannibal1997, p. 173; Hannibal, Reference Hannibal2000, p. 30), which has been interpreted as being marginal marine, consisting of predominantly marine organisms capable of tolerating changes in salinity (Baird et al., Reference Baird, Shabica, Anderson and Richardson1985; Baird, Reference Baird1997). And the Triassic Hannibaliulus wilsonae is from a brackish-water facies that has yielded marine or marginal marine organisms including lingulid brachiopods and limulids (Shear et al., Reference Shear, Selden and Gall2009, p. 2), two groups that are also found in the Luoping biota. The Luoping biota has a stronger marine influence than have these other facies, however. The assumed transportation distances to the basin are not extreme for millipedes. Distribution of extant millipedes shows that millipedes have been able to colonize land across long distances of open ocean. A premier example is the colonization of the Hawaiian Islands by various millipedes (Shelley and Golovatch, Reference Shelley and Golovach2011, p. 33, 68), presumably by rafting. The paucity of millipedes in the Luoping biota contrasts with the more common occurrence of millipedes in some assemblages with a nonmarine origin, for instance in the Cretaceous site of Las Hoyas, Spain, where millipedes are more numerous, but still only a small proportion of the fauna (Selden and Shear, Reference Selden and Shear2016).
Acknowledgments
This work was supported by the National Natural Science Foundation of China (nos. 41502013, 41772022 and 41661134047), Chinese Geological Survey projects (Grant nos. DD20160020, 12120114068001, 1212011140051, 1212011120621, and 1212010610211). JTH thanks colleagues at the Paleontological Institute, Moscow, and the National Museum of the Czech Republic, for allowing access to material from Russia and Bohemia. We also thank two anonymous reviewers for their insightful comments.