



Introduction
Ceratopsids are a clade of large-bodied herbivorous dinosaurs that rapidly diversified in the latest Cretaceous (Campanian–Maastrichtian) of North America, and have a well-sampled fossil record (Dodson et al., Reference Dodson, Forster and Sampson2004). Recent discoveries from southern Alberta (Ryan, Reference Ryan2007; Ryan et al., Reference Ryan, Russell and Hartman2010, Reference Ryan, Evans and Shepherd2012; Evans and Ryan, Reference Evans and Ryan2015), Montana (Longrich, Reference Longrich2013; Ryan et al., Reference Ryan, Evans, Currie and Loewen2014), and Utah (Loewen et al., Reference Loewen, Farke, Sampson, Getty, Lund and O’Connor2013; Sampson et al., Reference Sampson, Lund, Loewen, Farke and Clayton2013; Lund et al., Reference Lund, O’Connor, Loewen and Jinnah2016a) have revealed a diversity of early ceratopsids that have significantly increased our knowledge of the plesiomorphic anatomy of the group. However, the fossil record of the early radiation of ceratopsids remains poor, and in light of new anatomical information that obscures the morphological distinction between the two subfamilies, Centrosaurinae and Chasmosaurinae, conventional wisdom about the systematic position of taxa known from fragmentary material may require re-evaluation (Mallon et al., Reference Mallon, Ott, Larson, Iuliano and Evans2016).
Medusaceratops lokii Ryan, Russell, and Hartman, Reference Ryan, Russell and Hartman2010 from the lowermost strata of the Judith River Formation is one such ceratopsid taxon that has a complicated taxonomic history, with the hypodigm having been previously assigned to both subfamilies. The material of the taxon was collected from a middle Campanian bonebed, called the Mansfield bonebed (ca. 79 Ma; Roberts et al., Reference Roberts, Sampson, Deino, Bowring and Buchwaldt2013), making it one of the oldest known members of Ceratopsidae (Kirkland and DeBlieux, Reference Kirkland and DeBlieux2010; Longrich, Reference Longrich2013; Evans and Ryan, Reference Evans and Ryan2015; Campbell et al., Reference Campbell, Ryan, Holmes and Schröder-Adams2016). The Mansfield bonebed ceratopsid material was initially assigned to Albertaceratops nesmoi Ryan, Reference Ryan2007, which is unequivocally a basal member of Centrosaurinae, based on superficial resemblances of some morphologies to the A. nesmoi holotype, such as the large supraorbital horncores and the large pachyostotic parietal hook (Ryan, Reference Ryan2007). Subsequently, the two most complete parietals from the bonebed were described as a new chasmosaurine species, Medusaceratops lokii, based on an interpretation of the specimens exhibiting only three epiparietals on each side of the midline, which is typical of Chasmosaurinae (Ryan et al., Reference Ryan, Russell and Hartman2010). The remaining material from the bonebed was left as indeterminate Centrosaurinae (Ryan et al., Reference Ryan, Russell and Hartman2010). However, Longrich (Reference Longrich2013) retained the original centrosaurine designation, noting that the number of epiparietals is variable in Chasmosaurinae, and the parietosquamosal contact more closely resembles that of Centrosaurinae.
Here, we describe newly collected centrosaurine material from the Mansfield bonebed, re-examine the type series of Medusaceratops, reassess the known specimens from the bonebed, and conduct a systematic revision of the taxon. New data allow Medusaceratops to be included in a numerical cladistic analysis for the first time. We confirm the subfamilial identity of Medusaceratops as an early centrosaurine, which has implications for understanding the early diversification of ceratopsids.
Geological context
The Mansfield bonebed is located in the badlands of Kennedy Coulee, north of Rudyard, Hill County, Montana, USA (Fig. 1.1). Here, the middle Campanian alluvial deposits, commonly referred to the lower Judith River Formation (Goodwin and Deino, Reference Goodwin and Deino1989), crop out extensively along the drainage systems flowing toward the Milk River Valley in the north (Fig. 1.2, 1.3). Following recent stratigraphic revision of the Judith River Formation by Rogers et al. (Reference Rogers, Kidwell, Deino, Mitchell, Nelson and Thole2016), the beds exposed at Kennedy Coulee correlate to the McClelland Ferry Member to the south, but also with the upper Foremost and overlying lower Oldman formations of southern Alberta to the north, including the Taber Coal Zone (TCZ) and the Herronton Sandstone Zone (HSZ; Ogunyomi and Hills, Reference Ogunyomi and Hills1977; Eberth and Hamblin, Reference Eberth and Hamblin.1993; Cullen et al., Reference Cullen, Fanti, Capobianco, Ryan and Evans2016). The TCZ, which represents the top of the Foremost Formation in Alberta and correlative coal deposits exposed to the south, represents a datum for calibrating stratigraphic sections and associated fossil taxa (Eberth and Hamblin, Reference Eberth and Hamblin.1993; Brinkman et al., Reference Brinkman, Russell, Eberth and Peng2004; Ryan, Reference Ryan2007; Evans and Ryan, Reference Evans and Ryan2015; Freedman Fowler and Horner, Reference Freedman Fowler and Horner.2015; Cullen et al., Reference Cullen, Fanti, Capobianco, Ryan and Evans2016; Ryan et al., Reference Ryan, Holmes, Mallon, Loewen and Evans2017). The Mansfield bonebed occurs ~10 m above the top of the Marker A Coal layer, which is equivalent to the top of the TCZ (sensu Goodwin and Deino, Reference Goodwin and Deino1989) based on multiple sections measured in the Kennedy Coulee and at the Probrachylophosaurus bergei Freedman, Fowler, and Horner, 2015 locality (MOR locality JR-518), which is ~8 km to the north east. There are two bentonite layers occurring ~10 m below and ~25 m above the TCZ, which provides radiometric ages of 79.02 and 78.01 Ma, respectively (Roberts et al., Reference Roberts, Sampson, Deino, Bowring and Buchwaldt2013, but see Freedman Fowler and Horner, Reference Freedman Fowler and Horner.2015 and Fowler, Reference Fowler2016 for discussion), that bracket the Mansfield bonebed and constrain its chronostratigraphic age. The bonebed host beds sit on top of a 10 m thick interval of interbedded organic-rich mudstones with discontinuous carbonaceous seams, siltstone, and sandstones (Fig. 1.4). Fossils occur at the base of a 3 m thick, fining-upward sandy channel deposit that is rich in ironstone nodules; paleocurrent measurements suggest predominant flows toward the ESE.
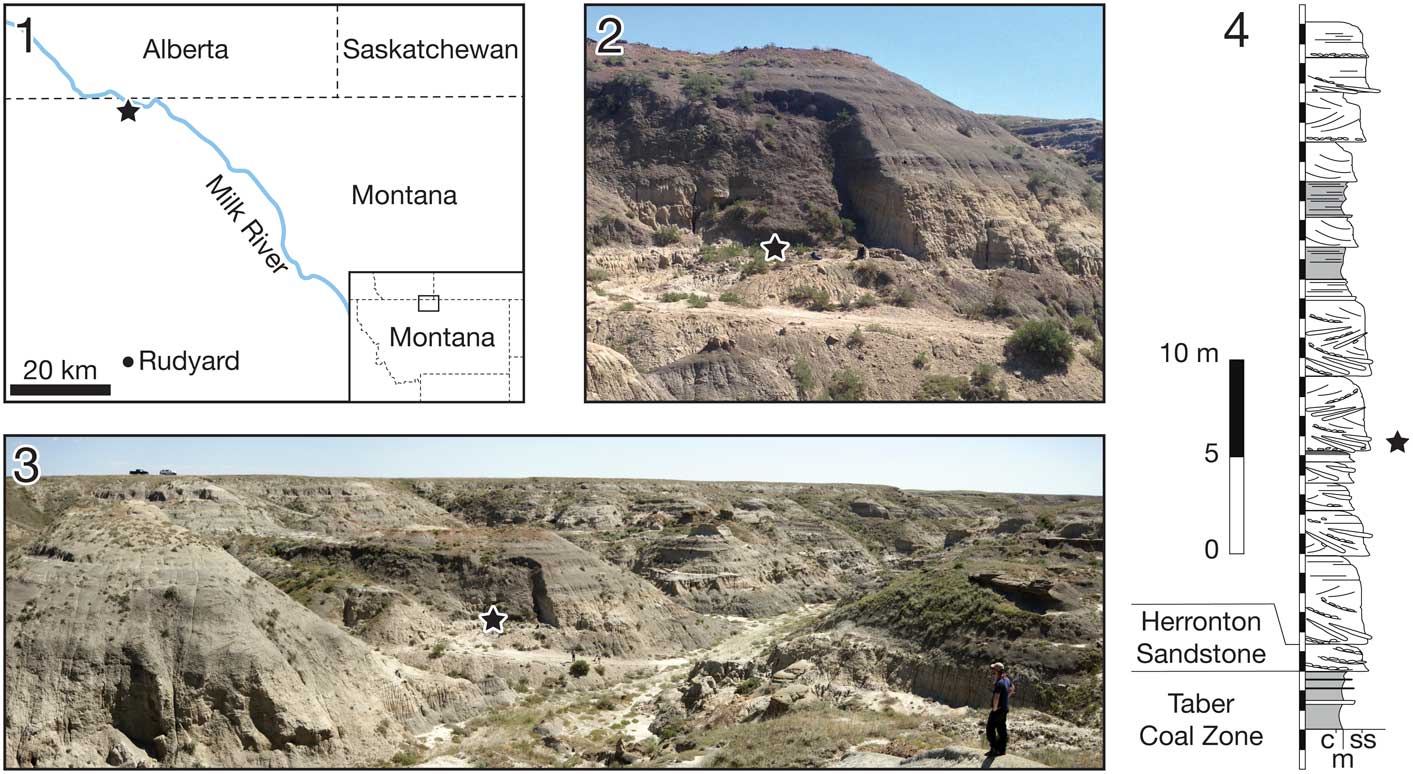
Figure 1 (1) Locality map of the Mansfield bonebed. (2, 3) Overview photos of the locality area. (4) Stratigraphic column of the locality area. The bonebed is indicated by a black star.
The stratigraphic occurrence of Medusaceratops places the taxon above the Herronton Sandstone Zone in the same stratigraphic interval where Albertaceratops and Wendiceratops were recovered ~25 km to the northwest in southern Alberta. Correlation to the top of the TCZ places Albertaceratops slightly lower in the section (8 m above the TCZ) with respect to Medusaceratops (10 m above the TCZ) and Wendiceratops (12 m above the TCZ), making them virtually indistinguishable stratigraphically. Due to the discontinuous nature of the beds forming the HSZ and the channelized deposits in which the Medusaceratops bonebed occurs, the reciprocal occurrence of these taxa may be slightly inaccurate.
Materials and methods
The Mansfield bonebed has been commercially excavated by Canada Fossils Ltd since 1994. The bonebed specimens previously described in Ryan (Reference Ryan2007) and Ryan et al. (Reference Ryan, Russell and Hartman2010) are housed in the Royal Tyrrell Museum of Palaeontology (TMP) and the Wyoming Dinosaur Center (WDC). The new material described here was collected in the summer of 2011 and 2012 by a crew led by D. Trexler of the Two Medicine Dinosaur Center in Bynum, Montana, and subsequently acquired by the Royal Ontario Museum. The quarry was visited by DCE and FF on July 16, 2016, where a detailed stratigraphic section was measured and sedimentological observations were made at the quarry site.
Photogrammetric 3D models of selected specimens (Figs. 3, 6) were created using Agisoft PhotoScan Standard Edition, ver. 1.1 and 1.2 to help visualize certain morphologies that can be obscured by the dark color of the specimens. The models were exported as STL files and scaled to the original size in mm using MeshLab, ver. 1.3.3 (SourceForge). The models were then oriented and screen-captured on Avizo, ver. 6.1.1. (FEI). The 3D models were uploaded to Dryad (http://doi.org/10.5061/dryad.8h067).
For the phylogenetic analysis, Medusaceratops was coded into the data matrix of Ryan et al. (Reference Ryan, Holmes, Mallon, Loewen and Evans2017) based on the hypodigm listed in the supplemental material (Text S1 and Table S1). Based on our interpretation of the available material, there is no indication of any other ceratopsid species in the Mansfield bonebed collections, and it is therefore assumed to be a monodominant bonebed, which is common for centrosaurines (Ryan et al., Reference Ryan, Russell, Eberth and Currie2001; Eberth and Getty, Reference Eberth and Getty2005; Ralrick and Tanke, Reference Ralrick and Tanke2008; Chiba et al., Reference Chiba, Ryan, Braman, Eberth, Scott, Brown, Kobayashi and Evans2015; Eberth, Reference Eberth2015). Hence, all ceratopsid material from the bonebed is interpreted to represent a single species, and coded as a single operational taxonomic unit, following common practice in centrosaurine studies (e.g., Sampson et al., 1995; Ryan and Russell, Reference Ryan and Russell2005; Currie et al., Reference Currie, Langston and Tanke2008; Farke et al., Reference Farke, Ryan, Barrett, Tanke, Braman, Loewen and Graham2011; Fiorillo and Tykoski, Reference Fiorillo and Tykoski2012; Ryan et al., Reference Ryan, Evans and Shepherd2012; Evans and Ryan, Reference Evans and Ryan2015). We added Medusaceratops lokii and the recently described centrosaurine, Machairoceratops cronusi Lund et al., Reference Lund, O’Connor, Loewen and Jinnah2016a into the original data matrix, which includes 28 ceratopsian operational taxonomic units.
The final matrix is composed of 101 characters, which includes 97 characters used first in Farke et al. (Reference Farke, Ryan, Barrett, Tanke, Braman, Loewen and Graham2011) and subsequently used in Sampson et al. (Reference Sampson, Lund, Loewen, Farke and Clayton2013), and four characters added by Evans and Ryan (Reference Evans and Ryan2015). All characters were equally weighted, but Character 20 was treated as an ordered (additive) character following previous analyses (e.g., Farke et al., Reference Farke, Ryan, Barrett, Tanke, Braman, Loewen and Graham2011; Sampson et al., Reference Sampson, Lund, Loewen, Farke and Clayton2013; Evans and Ryan, Reference Evans and Ryan2015). Codings of two characters are modified from Ryan et al. (Reference Ryan, Holmes, Mallon, Loewen and Evans2017). Reassignment of the Mansfield bonebed material to Medusaceratops in this paper results in the coding change for Character 30 (curvature of supraorbital horncore in rostral view) for Albertaceratops from ‘2’ (lateral) to ‘?’. Character 100 (shape of epiparietal 1) was modified for two taxa as follows: from 1 to 2 (elongate flattened process or spike, greater than twice as long as wide) in Coronosaurus brinkmani (Ryan and Russell, Reference Ryan and Russell2005) based on TMP 2002.068.0001; from 1 to (0 and 1) in Styracosaurus albertensis Lambe, Reference Lambe1913 reflecting polymorphism based on CMN 344 and ROM 1436 (Ryan et al., Reference Ryan, Holmes and Russell2007). We follow the epiparietal homology argument of Evans and Ryan (Reference Evans and Ryan2015), which counts the most medially positioned paired epiparietals as ep 1 in non-eucentrosauran (see following paragraph) centrosaurines. The phylogenetic analysis was performed using Traditional Search with the Tree Bisection Reconnection algorithm in TNT ver. 1.5 (Goloboff and Catalano, Reference Goloboff and Catalano2016) with Leptoceratops gracilis Brown, Reference Brown1914b designated as the outgroup taxon, following the previous analysis (Sampson et al., Reference Sampson, Lund, Loewen, Farke and Clayton2013). The analysis was run with 1,000 replicates, and up to 1,000 trees were saved in each replication. Branches are collapsed if there is no possible support (“Rule 3”). To assess the robustness of the attained topology, standard bootstrap resampling (sampling with replacement) was conducted with 10,000 replications of traditional tree search, and Bremer support was computed with retaining trees suboptimal by ten steps. In order to further test the subfamilial phylogenetic position of Medusaceratops lokii as a centrosaurine rather than a chasmosaurine, a constraint analysis was performed in which the position of M. lokii was constrained as the sister taxon of Chasmosaurus and Pentaceratops in a monophyletic Chasmosaurinae, using this option in TNT.
We define the new node-based centrosaurine clade Eucentrosaura as the least inclusive clade containing Centrosaurus apertus and Pachyrhinosaurus canadensis in order to facilitate the morphological comparisons of the Medusaceratops material with other centrosaurines in this study. Eucentrosaura includes the members of Centrosaurini and Pachyrhinosaurini recovered in the strict consensus tree topology described in the phylogenetic analysis section of this paper, which is consistent with other recent phylogenetic analyses of centrosaurines (e.g., Farke et al., Reference Farke, Ryan, Barrett, Tanke, Braman, Loewen and Graham2011; Evans and Ryan, Reference Evans and Ryan2015; Ryan et al., Reference Ryan, Holmes, Mallon, Loewen and Evans2017).
A histological thin section of the diaphysis of a large tibia (ROM 67873) was made using standard techniques (Lamm, Reference Lamm2013) in the palaeohistology lab at the Royal Ontario Museum. The original thin section images made using plane and cross-polarized light are available at http://doi.org/10.5061/dryad.8h067.
Repositories and institutional abbreviations
Canadian Museum of Nature (CMN), Ottawa, Canada; Fukui Prefectural Dinosaur Museum (FPDM), Fukui, Japan; Museum of the Rockies (MOR), Bozeman, Montana, USA; Royal Ontario Museum (ROM), Toronto, Ontario, Canada; Royal Tyrrell Museum of Palaeontology (TMP), Drumheller, Alberta, Canada; Wyoming Dinosaur Center (WDC), Thermopolis, Wyoming, USA.
Systematic paleontology
Dinosauria Owen, Reference Owen1842
Ornithischia Seeley, Reference Seeley1887
Ceratopsia Marsh, Reference Marsh1890
Neoceratopsia Sereno, Reference Sereno1986
Ceratopsidae Marsh, Reference Marsh1888
Centrosaurinae Lambe, Reference Lambe1915
Medusaceratops Ryan, Russell, and Hartman, Reference Ryan, Russell and Hartman2010
Medusaceratops lokii Ryan, Russell, and Hartman, Reference Ryan, Russell and Hartman2010
Holotype
WDC-DJR-001, an incomplete parietal comprised of a largely complete left lateral bar with loci for epiparietals (ep) 1–5 (Fig. 2.1–2.4).

Figure 2 Previously described parietals from the Mansfield bonebed: (1) ventral view, (2) close-up of ep 1, (3) dorsal view, and (4) close-up of lateral view of lateral ramus of WDC-DJR-001; (5) dorsal and (6) ventral views of WDC-DJR-002; (7) ventral and (8) lateral view of TMP 2002.069.0005. Upper and lower scale bars are for (1, 3) and (5–8), respectively. Abbreviation: ep, epiparietal.
Referred material
All centrosaurine ceratopsid material from the Mansfield bonebed. See supplementary table (Table S1) for the complete list of the referred material examined in this paper.
Locality and horizon
The Mansfield bonebed is located in the outcrop along the Kennedy Coulee near Rudyard, Montana, USA (Fig. 1.1). Detailed locality data is on file at the ROM, TMP, and WDC. The bonebed layer is located ~10 m above the top of the Marker A coal (Goodwin and Deino, Reference Goodwin and Deino1989) in the McClelland Ferry Member of the Judith River Formation (Rogers et al., Reference Rogers, Kidwell, Deino, Mitchell, Nelson and Thole2016). The age of the bonebed is constrained between 79.0 Ma and 78.7 Ma by radiometric dates derived from bentonites that occur 5 m below and 27 m above the top of the Marker A Coal, respectively, within Kennedy Coulee (Roberts et al., Reference Roberts, Sampson, Deino, Bowring and Buchwaldt2013, but see Freedman Fowler and Horner, Reference Freedman Fowler and Horner.2015 and Fowler, Reference Fowler2016 for discussion).
Emended diagnosis
Centrosaurine ceratopsid with five epiparietals on posterolateral parietal ramus, from medial to lateral: ep 1, a small, sometimes procurved epiparietal; ep 2, comparatively massive, broad-based pachyostotic process that is strongly curved and projects anterolaterally; ep 3, small pachyostotic process curved anterolaterally, with a similar shape to ep 2; ep 4 and ep 5, unmodified, small triangular-shaped epiparietals. Medusaceratops differs from Albertaceratops in possession of a low, elongate ep 1 as the medialmost epiparietal, and differs from Xenoceratops in the strong anterior curvature and projection of ep 2.
Remarks
The specimen figured as the paratype in the original description of Medusaceratops (Fig. 12.2 in Ryan et al., Reference Ryan, Russell and Hartman2010) is not WDC-DJR-002, but TMP 2002.069.0005. Here, we retain WDC-DJR-002 as a paratype since this specimen preserves the diagnostic combination of parietal ornamentation, notably part of the small ep 1 and the pachyostotic ep 2, rather than TMP 2002.069.0005. Casts of the holotype and paratype specimens are reposited at the ROM and at the TMP.
WDCB-MC-001, is a large, intact region of the skull roof that includes nasal and postorbital ornamentation, that was reported to have been collected from the Mansfield bonebed by Ryan (Reference Ryan2007). However, new information suggests that, although it was collected in Kennedy Coulee, it probably did not originate in the bonebed (D. Trexler, personal communication, 2017). For this reason, and the fact that the postorbital horncores are much shorter than any known from the bonebed, we do not include it in the hypodigm of Medusaceratops lokii at this time.
Description
The frill ornamentation, composed of co-ossified epiossifications, and the facial horns are generally the most diagnostic suite of traits for ceratopsid dinosaurs, especially for centrosaurines (Dodson et al., Reference Dodson, Forster and Sampson2004), thus we describe only the parietal, squamosal, postorbital, and nasal below, following previous work (e.g., Ryan and Russell, Reference Ryan and Russell2005). Due to the incomplete nature of the material, and to provide clarity with respect to the new interpretations presented here, detailed descriptions of individual specimens are provided in the appropriate sections, following a general description of each element. A comprehensive list of specimens comprising the hypodigm of Medusaceratops referred to in this study is provided in the supplemental data (Table S1).
Parietal
Re-examination of WDC-DJR-001 (holotype; Fig. 2.1–2.4), with reference to ROM 73832 (Fig. 3.1–3.5) and ROM 73836 (Fig. 3.8–3.10) indicates that Medusaceratops lokii has five, not three, epiparietals (Fig. 4.3). These are the newly recognized diminutive epiparietal (ep 1) and the small triangular-shaped epiparietal (ep 5) that are positioned medially and laterally, respectively, to the three epiparietals that were originally recognized by Ryan et al. (Reference Ryan, Russell and Hartman2010). From medial to lateral, these epiparietals have the following morphology: one small epiparietal (ep 1) sits on the posterior margin of the posterior ramus adjacent to the midline (i.e., ROM 73832, ROM 73837, WDC-DJR-001, and WDC-DJR-002); a large pachyostotic hook (ep 2) curves anterolaterally (i.e., WDC-DJR-001, WDC-DJR-002, and TMP 2002.069.0005), a smaller pachyostotic hook (ep 3) curves subtly anterolaterally (i.e., WDC-DJR-001, TMP 2002.069.0005), and two smaller, triangular-shaped epiparietals (ep 4 and 5; i.e., ROM 73836 and WDC-DJR-001).

Figure 3 Newly described parietals from the Mansfield bonebed: (1) oblique view (direction is figured by arrows above) of 3D model, (2) posterior view, (3) dorsal view, (4) anterior view, and (5) left lateral view of ROM 73832; (6) dorsal and (7) oblique view (direction figured by arrows above) of 3D model of ROM 73837; (8) lateral view, (9) dorsal view, and (10) close-up of ventral surface of ROM 73836. Abbreviations: ep, epiparietal; mb, bumps on midline ramus; mr, midline ramus; pr, posterior ramus; sqc, squamosal contact.
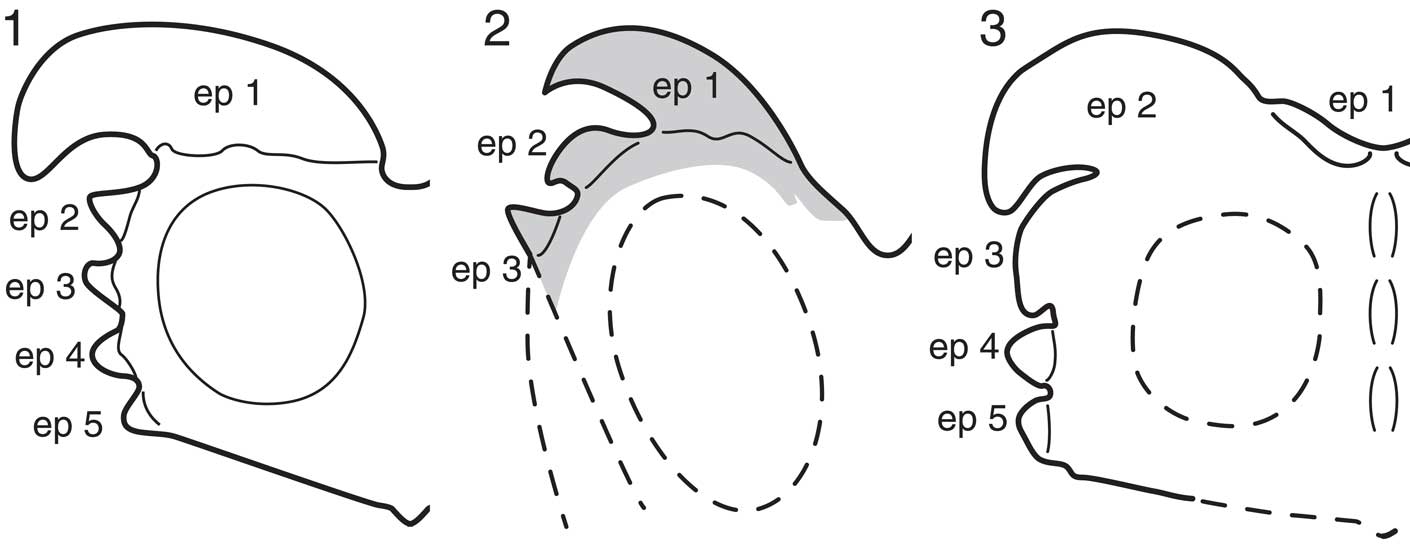
Figure 4 Comparison of frill ornamentations of (1) Albertaceratops nesmoi in Ryan (Reference Ryan2007), (2) previous reconstruction of Medusaceratops lokii frill in Ryan et al. (Reference Ryan, Russell and Hartman2010) (gray part represents where the holotype [WDC-DJR-001] corresponds), and (3) new reconstruction of M. lokii frill in this study, mainly based on ROM 73832, ROM 73836, and WDC-DJR-001. Abbreviation: ep, epiparietal.
The number of parietal processes of Medusaceratops and other morphological features seen on parietals from the bonebed do not reveal any definitive chasmosaurine traits. However, they exhibit several key centrosaurine characters, such as a broad midline ramus (ROM 73832), imbricated epiparietals (ROM 73836; also seen on the previously described holotype WDC-DJR-001, as well as TMP 2002.069.0005), and a convex, interdigitating squamosal contact (ROM 73836) (Longrich, Reference Longrich2013). Longrich (Reference Longrich2013) noted that the preserved epiparietals are different between WDC-DJR-001 and TMP 2002.069.0005 due to the size discrepancies of ep 3 (P2 in Ryan et al., Reference Ryan, Russell and Hartman2010). However, the ep 3 of TMP 2002.069.0005 is lobe-shaped rather than being an unmodified, triangular shape. Therefore, we agree with the interpretation of Ryan et al. (Reference Ryan, Russell and Hartman2010) that the preserved epiparietals on TMP 2002.069.0005 are ep 2–4. We interpret the size difference of ep 3 on TMP 2002.069.0005 and WDC-DJR-001 as an individual and/or ontogenetic variation.
ROM 73832 (Fig. 3.1–3.5).—ROM 73832 preserves the posterior half of a midline ramus and the medial part of the left posterior ramus with a distinct yet diminutive epiparietal (ep 1) adjacent to the midline. The ventral surface is relatively smooth, whereas the dorsal side is incised by numerous vascular grooves, especially on the tori on the midline ramus and the epiparietal on the posterior ramus, and has adult bone texture (sensu Sampson et al., Reference Sampson, Ryan and Tanke1997; Brown et al., Reference Brown, Russell and Ryan2009). Ep1 is well fused to the posterior ramus, and its contact boundary is indistinguishable (Fig. 3.3). Taken together, the bone surface texture, the epiparietal fusion, and the thickened posterior margin of the midline ramus (31.6 mm), suggest that this specimen is derived from a skeletally mature, adult individual (Sampson et al., Reference Sampson, Ryan and Tanke1997). In cross-section, the midline ramus tapers laterally on either side of the midline, making it transversely broad with a triangular cross-sectional shape (Fig. 3.4), which is a synapomorphy for Centrosaurinae (Farke et al., Reference Farke, Ryan, Barrett, Tanke, Braman, Loewen and Graham2011; Sampson et al., Reference Sampson, Lund, Loewen, Farke and Clayton2013; Evans and Ryan, Reference Evans and Ryan2015). The midline ramus has two low tori (‘bumps’) on the dorsal side (Fig. 3.1 and 3.5). An embayment of the posterior ramus at the midline is present, but it is wide and relatively shallow (Fig. 3.3), as in Xenoceratops (Ryan et al., Reference Ryan, Evans and Shepherd2012).
Ep 0 is definitively absent in this specimen. The base of the preserved epiparietal, ep 1, is unusual compared to other centrosaurines in that the body exhibits a slight torsion such that its medial portion is located on the dorsal surface of the posterior ramus, but the lateral portion is on the posterior margin (Fig. 3.1–3.3). This epiparietal is proportionally broad and low (114.2 by 15.7 mm), but projects forward (Fig. 3.1). Although smaller, it is most similar in morphology and position to the ep 1 of Xenoceratops foremostensis Ryan, Evans, and Shepherd, Reference Ryan, Evans and Shepherd2012 (P2 in Ryan et al., Reference Ryan, Evans and Shepherd2012) among known centrosaurines.
ROM 73836 (Fig. 3.8–3.10).—ROM 73836 is an anterolateral segment of the parietal that includes the three lateralmost epiparietals (ep 3–5, also see description of WDC-DJR-001 below) and the squamosal contact of the right lateral ramus. Although all three epiparietals are completely fused, the boundary between the epiparietals and the underlying parietal ramus is clearly delineated (Fig. 3.9). On both dorsal and ventral surfaces, the specimen exhibits the combination of a long-grained texture on the lateral side and adult bone texture on the medial side of the lateral parietal ramus (Fig. 3.10). Note that the long-grained texture is always proximally associated with the mottled texture but not the adult texture in centrosaurines (e.g., Avaceratops, “Brachyceratops,” “Monoclonius,” Coronosaurus brinkmani, Centrosaurus, Styracosaurus, Pachyrhinosaurus) that were examined by Brown et al. (Reference Brown, Russell and Ryan2009). The intriguing mixture of textures on ROM 73836 may suggest a unique ontogenetic textual transition in Medusaceratops. The squamosal contact is rugose and convex, which is typical of centrosaurines but not known in chasmosaurines (Dodson et al., Reference Dodson, Forster and Sampson2004).
The two epiparietals closest to the squamosal contact (ep 4 and 5) have a low and rounded triangular morphology when viewed dorsally, similar to unmodified epiparietals in other centrosaurines. Comparatively, the epiparietal (ep 3) posterior to these has a shorter base and a tall, triangular profile with an acute apex (Fig. 3.9). Ep 3 closely resembles the morphology and proportions of the lateral epiparietals on the holotype of Albertaceratops (TMP 2001.026.0001). All of the epiparietals on ROM 73836 are strongly imbricated (Fig. 3.8), and the squamosal contact is convex, both of which support a centrosaurine assignment for the Mansfield bonebed material (Dodson et al., Reference Dodson, Forster and Sampson2004).
WDC-DJR-001 (holotype, Fig. 2.1–2.4).—Part of the posterior ramus and all of the lateral ramus of the parietal are preserved on WDC-DJR-001, the holotype of M. lokii. Long-grained bone texture occurs on the lateral side of the lateral ramus, as seen on ROM 73836. The posterior ramus thickness (28.1 mm) is also similar to that of ROM 73832, the midline bar. Ryan et al. (Reference Ryan, Russell and Hartman2010) recognized only three epiparietals on this specimen: from medial to lateral these are; a large, wide-based, and pachyostotic hook; a smaller lobe-shaped process; and a round triangular-shaped epiparietal (Ryan et al., Reference Ryan, Russell and Hartman2010). However, comparisons to ROM 73832 and ROM 73836 suggest that WDC-DJR-001 actually possesses two previously unidentified epiparietal loci, for a total of five.
The additional epiparietals include a diminutive epiparietal medial to the large pachyostotic hook, which we interpret as ep 1, and an open sutural locus for ep 5. The medialmost epiparietal (ep 1) is represented by a small eminence with a porous texture on the posterodorsal margin of the posterior ramus (Fig. 2.2, 2.3). Although the size of the epiparietal differs from the ep 1 on ROM 73832, the incipient nature and the location of these epiparietals on the holotype and ROM 73832 suggests that they are homologous. Other parietals exhibit the presence of a variably sized epiparietal (ep 1) medial to the enlarged pachyostotic hook (ep 2) (see below for detailed description of WDC-DJR-002, ROM 73837 [Fig. 3.6, 3.7] and TMP 2002.069.0006).
The smaller pachyostotic process (ep 3) and the round triangular-shaped epiparietal (ep 4) are imbricated (Fig. 2.4). The pattern of imbrication indicates that the holotype preserves the left side of the frill, instead of the right side as Ryan et al. (Reference Ryan, Russell and Hartman2010) originally described. According to the re-orientation, the large pachyostotic hook (ep 2) slightly flexes ventrally at its base and has a distinct margin of the overgrowth on the ramus ventrally, which is similar to the condition of P1 and 2 of Coronosaurus brinkmani parietals from the Milk River Ridge Reservoir bonebed (Ryan and Russell, Reference Ryan and Russell2005).
The lateral parietal ramus of WDC-DJR-001 has a wide gap (96.3 mm) between the squamosal contact and its anteriormost fused epiparietal (ep 4), which is unusual in centrosaurines. However, ROM 73834, which lacks this gap, has an epiparietal very close to the squamosal contact. The corresponding area of the holotype shows an open, asymmetrical, interdigiting suture on the lateral surface (Fig. 2.4), indicative of the prior presence of an unfused epiparietal (ep 5) that was lost post-mortem.
TMP 2002.069.0005 (Fig. 2.7, 2.8).—TMP 2002.069.0005 is composed of a right lateral ramus with a large pachyostotic ep 2, a lobe-like ep 3, and an undulating lateral surface representing the locus for unattached ep 4. The ep 3 and the ep 4 loci are imbricated. The ep 2 of this specimen differs in the lack of overgrowth at the base on the ventral side of the parietal ramus. The ep 3 is also smaller than that of WDC-DJR-001, but the morphology of this epiparietal is more similar to the anteriorly curved ep 3 on WDC-DJR-001 than to an unmodified epiparietal.
WDC-DJR-002 (paratype; Fig. 2.5, 2.6).—WDC-DJR-002 is a right half of the posterior ramus of the parietal. There is a large, laterally oriented, pachyostotic hook, which is comparable to ep 2 on WDC-DJR-001 and TMP 2002.069.0005. The ventral overgrowth of the ep 2 does not appear on this specimen, similar to TMP 2002.069.0005, but unlike WDC-DJR-001 and ROM 73837. Medial to the ep 2, this specimen has a lateral edge of the ep 1 on the posterior margin of the posterior ramus. Lateral to the ep 2, there is a partially preserved undulating surface representing a locus for ep 3, but the morphology of this ep 3 cannot be inferred due to the broken nature of the specimen.
FPDM-V-10 (not figured).—This specimen is a composite skeleton composed of the cranial and postcranial material from the Mansfield bonebed. Since the bonebed material was thought to represent a chasmosaurine, the frill reconstruction is incorrect. Three original frill specimens (not figured), including one partial midline ramus, one partial right ramus, and one partial left lateral ramus, are incorporated into the frill. The midline ramus has long-grained bone texture and a thin posterior margin (22.7 mm), suggesting this specimen is derived from a juvenile individual. There are three bumps along the dorsal midline, and the cross-section is triangular, as in centrosaurines. From posterior to anterior, the right lateral parietal ramus preserves a partial lobe-shaped epiparietal and two unmodified triangular epiparietals. The morphology and arrangement of the epiparietals on this specimen are congruent with the morphology of the ep 3-5 of other Mansfield bonebed parietal specimens (WDC-DJR-001 and TMP 2002.069.0005). The less complete left ramus has two unmodified, triangular epiparietals that likely represent ep 4 and ep 5.
ROM 77214 (not figured).—This specimen is a lateral bar of a right parietal preserving the squamosal contact, three epiparietals, and another epimarginal locus. This parietal ramus exhibits long-grained bone textures on both ventral and dorsal surfaces, suggesting it represents a relatively young individual. The anterolateral half of the pachyostotic ep 2 epiparietal locus is preserved. It is thickened (28.5 mm) compared to the lateral ramus (17.7 mm, measured lateral to the base of the ep 2) and protruding by at least ~60 mm from the lateral margin of the ramus (the tip of the epiparietal is slightly damaged). As indicated by the bone surface texture, we refer this epiparietal to an undeveloped ep 2. The ep 3 is a low (32.3 mm), broad-based (103.6 mm), and rounded epiparietal. This ep 3 is not pachyostotic, and its tip is not projecting anterolaterally, unlike the ep 3 of WDC-DJR-001. The ep 4 is low and triangular-shaped, and fused to the ramus. Ep 3 and 4 are imbricated. Anterolateral to the ep 3, the lateral margin of the ramus has a rugose texture continuing to the squamosal contact, suggesting this area represents an open suture for either a distinct ep 5 or an epiparietosquamosal. Although the epiparietals are not fully developed, the configuration of the epiparietals on this specimen is congruent with other parietals described above.
Squamosal
None of the available squamosals are complete, but all show centrosaurine features. The preserved posterior flanges that form the lateral sides of frills are round rather than rectangular (ROM 73833, Fig. 5.4; TMP 2002.069.0003, Fig. 5.3), which is similar to taxa in the newly defined Eucentrosaura rather than basal centrosaurines (Maiorino et al., Reference Maiorino, Farke, Piras, Ryan, Terris and Kotsakis2013). The maximum number of preserved episquamosals (or loci) on a single specimen is three (ROM 73833, Fig. 5.4; TMP 2002.069.0003, Fig 5.3). The profiles of the episquamosals are wide and low crescent-shaped (TMP 2002.069.0003), or subrectangular (ROM 73833) when viewed dorsally, except the anteriormost episquamosal on TMP 2002.069.0002, which is large, completely fused, and semicircular in outline. A ridge or a series of dorsal tori are often developed on the lateral side of centrosaurine squamosal, which are particularly prominent in early centrosaurines (Penkalski and Dodson, Reference Penkalski and Dodson1999; Evans and Ryan, Reference Evans and Ryan2015; Rivera-Sylva et al., Reference Rivera-Sylva, Hedrick and Dodson2016). These bumps are only weakly developed on TMP 2002.069.0002 (Fig. 5.1, 5.2), except for a larger eminence preserved on the anterodorsal part of the squamosal near the broken postorbital contact. The shape of the parietal contact of the squamosal is uncertain because this region is only preserved on the pathological WDC-DJR-017 (Ryan, Reference Ryan2007), however, the convex squamosal contacts of the parietals (e.g., ROM 73834) indicate that the squamosal has the typical centrosaurine concave parietal contact.

Figure 5 Squamosals from the Mansfield bonebed: (1) lateral and (2) medial views of TMP 2002.069.0002; (3) lateral view of TMP 2002.069.0003; (4) lateral view of ROM 73833; (5) lateral view of WDC-DJR-017. Abbreviations: esq, episquamosal; esql episquamosal locus; itf, infratemporal fenestra; jn, jugal notch; jc, jugal contact; lb, lateral bump; lr, lateral ridge; pog, paroccipital groove; qc, quadrate contact.
TMP 2002.069.0002 (Fig. 5.1, 5.2).—TMP 2002.069.0002 is a partial left squamosal. The anterior part is relatively intact and the jugal contact is almost completely preserved on the ventral side of the lateral surface, although the postorbital contact is damaged. Most of the posterior flange is missing except for the posteroventral corner, therefore the overall shape of the flange cannot be assessed. The corner is represented by a large, completely fused semicircular episquamosal. A ridge associated with a series of bumps is weakly developed on the dorsal side of the squamosal, except for a more prominent bump preserved on the anterodorsal part of the squamosal near the broken postorbital contact. The jugal notch is deeply embayed compared to the dorsal margin of lateral temporal fenestra.
TMP 2002.069.0003 (Fig. 5.3).—This specimen is a partial right squamosal lacking the anterior portion and the dorsal margin of the posterior flange. Three crescentic episquamosals are preserved and two of them are intact. Both of them are wide (97.1 and 110 mm for anteroventral and posterodorsal ones, respectively) and low when viewed dorsally.
ROM 73833 (Fig. 5.4).—This specimen only preserves the posterior margin of the posterior flange with two episquamosals and an open, interdigitating suture at an episquamosal locus posterior to the two. The two episquamosals are wide (64.3 and 82.6 mm for anterior and posterior ones, respectively) and more rectangular than those of TMP 2002.069.0003. The inferred outline of the posterior flange is rounded, similar to TMP 2002.069.0003.
WDC-DJR-017 (Fig. 5.5).—This specimen is a partial left squamosal with a pathologically modified parietal contact (Ryan, Reference Ryan2007). There is an open suture for an episquamosal or epimarginal at the posterodorsal corner of the posterior flange. Two episquamosal loci are anterior to the open suture. The two preserved episquamosals are present completely fused and lack any evidence of their original sutural contact. A notch in the posterolateral squamosal margin represents the locus for an unfused epimarginal that straddled the parieto-squamosal suture.
Postorbital
Several well-preserved postorbital horncores have been recovered from the Mansfield bonebed (Fig. 6). These span a large ontogenetic range of horncore morphology from small, triangular, centrosaurine-like horncores, to elongated and robust chasmosaurine-like horncores. The following detailed specimen descriptions are ordered by size, which presumably reflects the relative maturity of each individual at the time of death. The reported horncore lengths record the rectilinear length from the postorbital margin to the tip of the horncore.
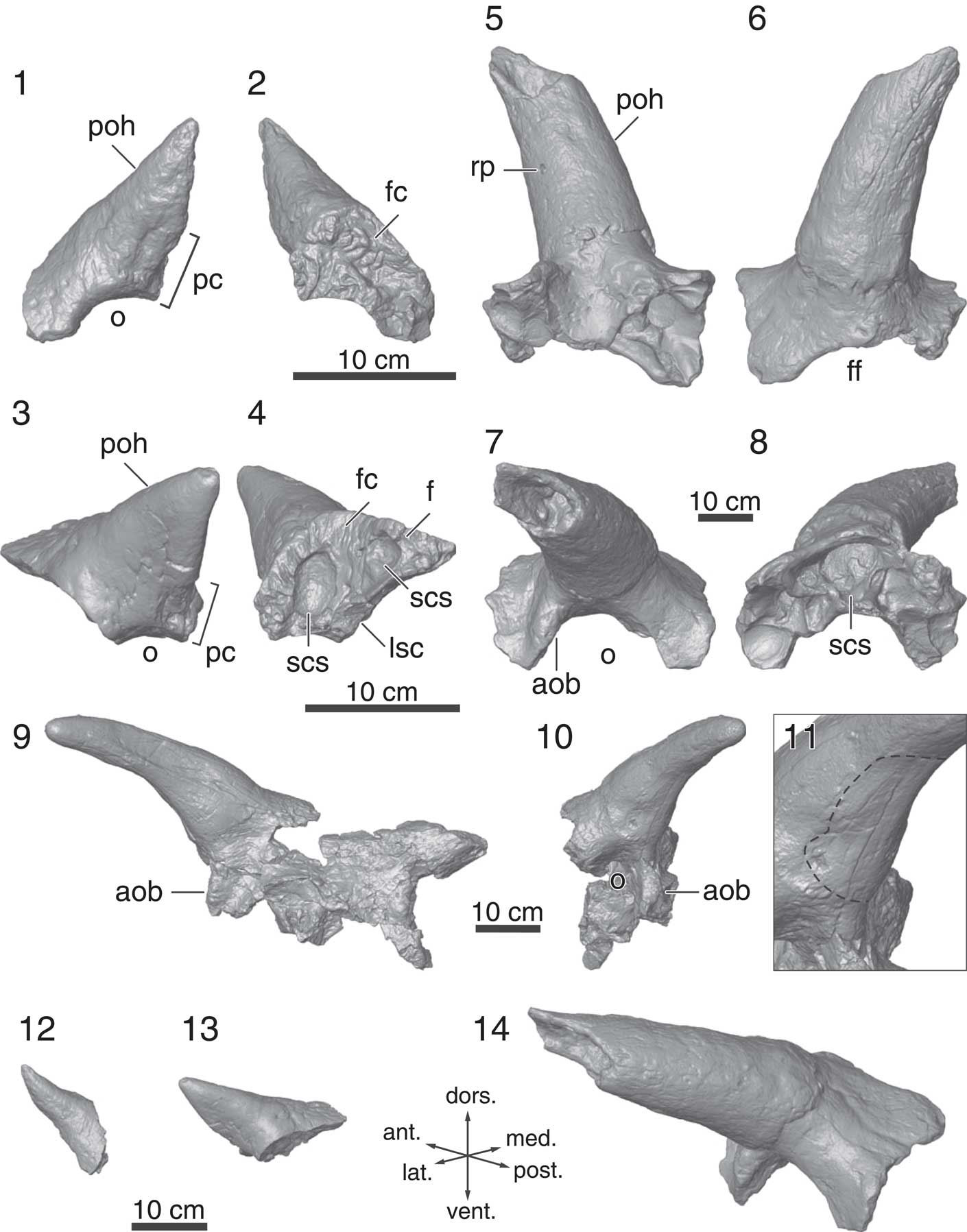
Figure 6 Postorbitals from the Mansfield bonebed; all of the specimens in this figure are represented by 3D models: (1) lateral and (2) medial views of TMP 2002.069.0010; (3) lateral and (4) medial views of ROM 73834; (5) ventral, (6) dorsal, (7) lateral, and (8) medial views of ROM 73831; (9) anterior, (10) right lateral views, and (11) the close-up of WDC-DJR-003; dotted line in (11) indicates shallow depression on the horncore; oblique views (direction is figured by arrows) of (12) TMP 2002.069.0010, (13) ROM 73834, and (14) ROM 73831, demonstrating ontogenetic change of postorbital horncores in Medusaceratops lokii. Abbreviations: aob, antorbital buttress; f, frontal; fc, frontal contact; ff, frontal fontanelle; lsc, laterosphenoid contact; o, orbit; pc, palpebral contact; poh, postorbital horncore; rp, resorption pit; scs, supracranial sinus.
TMP 2002.069.0010 (Fig. 6.1, 6.2, 6.12).—TMP 2002.069.0010 (Ryan, Reference Ryan2007; Fig. 10.4, 10.5) preserves a right postorbital with a complete horncore, but it is missing its posterior region. The short horncore (117.6 mm in length from the orbital margin to the tip of the horncore) is the smallest in the sample and the palpebral and frontal sutures are completely open (Fig. 6.1 and 6.2, respectively), suggesting it is from a juvenile individual. There is no evidence of a supracranial sinus system on this specimen. The horncore projects dorsally with slight anterolateral inclination (Fig. 6.1, 6.12). The gross morphology of the horncore is tall and pyramidal, which is similar to that of some juvenile centrosaurines (Ryan et al., Reference Ryan, Russell, Eberth and Currie2001; Ryan and Russell, Reference Ryan and Russell2005). However, the round cross-sectional horncore base resembles those of juvenile chasmosaurines (Currie et al., Reference Currie, Holmes, Ryan and Coy2016; Lehman et al., Reference Lehman, Wick and Barnes2016) and Zuniceratops (Wolfe et al., Reference Wolfe, Kirkland, Smith, Poole, Chinnery-Allgeier and McDonald2010).
ROM 73834 (Fig. 6.3, 6.4, 6.13).—This specimen only preserves the horncore portion of a right postorbital. The palpebral contact is completely open (Fig. 6.4). The frontal suture is only open anterior to the horncore and the frontal is indistinguishably fused posteriorly. The supracranial sinus system is represented by two depressions on the medioventral surface of the specimen (Fig. 6.4) that do not invade the shaft of the horncore, as is the typical condition for Centrosaurini (Farke, Reference Farke2010). The length of the horncore (139.7 mm) is ~20% longer than that of TMP 2002.069.0010 (Fig. 6.12). The cross section of the horncore is more flattened and triangular than that of TMP 2002.069.0010, with its flat surface projecting posterolaterally rather than laterally as for juvenile Centrosaurus apertus (Ryan et al., Reference Ryan, Russell, Eberth and Currie2001, figs. 10C–10F) and Coronosaurus brinkmani (Ryan and Russell, Reference Ryan and Russell2005).
WDC-DJR-003 (Fig. 6.9–6.11).—This specimen preserves the supraorbital region (fused lacrimals, palpebrals, postorbitals, and frontals) with the almost complete right (the tip is reconstructed in the figure) and incomplete left horncores. The preserved horncore length is 296 mm with a basal circumference of 383 mm. Unlike TMP 2002.069.0010 and ROM 73834, the cross-section of these horncores is circular (although they have been slightly distorted taphonomically). The horncore is strongly laterally oriented, but this is exaggerated due to the deformation (Fig. 6.9, 6.10). The basal overgrowth of the contact surface for the keratinous sheath, which is prominent on ROM 73831, is only poorly developed on the dorsoposterior two-thirds of the basal circumference of this specimen. A unique shallow, broad depression occurs on the anterior and ventral side of the horncore base, which is different from small round deep pit on the ROM 73831 (Fig. 6.11).
The frontal fontanelle margin is intact anteriorly, and it seems to have been open at the time of death. The supracranial sinus system is restricted to the base of the horncores. The antorbital buttress is swollen, making an oval-shaped eminence in lateral view (Fig. 6.10).
ROM 73831 (Fig. 6.5–6.8, 6.14).—ROM 73831 is a large (the basal circumference is 500 mm) left postorbital fused with its complementary lacrimal, palpebral, and frontal. The massive horncore is similar to those of Albertaceratops. The anterior region of the orbital rim is thickened, creating a prominent antorbital buttress (Fig. 6.7). The medial margin of the frontal does not reach the midline, indicating an open frontal fontanelle. The supracranial sinus (Fig. 6.8) partially invades the horncore at its base, similar to that seen in WDC-DJR-003. The tip of the horncore is broken and missing, and therefore the total length of the horncore is unknown, but the circumference of the horncore at its base is large (500 mm) and comparable to the largest horncore size of the non-Triceratopsini chasmosaurines (510 mm in Pentaceratops sternbergii, Wiman, Reference Wiman1930; Fig. 7; Table S2). The cross-section of the horncore is circular throughout the entirety of its preserved length. The horncore has a strong lateral inclination (~80˚ from the sagittal plane in the dorsal and ventral view), with only a limited projection dorsally (~20˚ from the horizontal plane in the anterior and posterior view). The horncore also has a slight curvature in dorsal and ventral views, but far less than that of Nasutoceratops (Sampson et al., Reference Sampson, Lund, Loewen, Farke and Clayton2013; Lund et al., Reference Lund, Sampson and Loewen2016b).

Figure 7 Comparison of postorbital horncore basal circumference between Medusaceratops, Centrosaurinae, non-Triceratopsini Chasmosaurinae, and Triceratopsini. In the box plots, mean values are represented by lines in the boxes, lower and upper bounds of the boxes represent the first and third quartiles, and the ends of the dashed lines indicate minimum and maximum values of the data. Data used for this plot are provided in Table S2.
The horncore has an unusual overgrowth that creates the step at the base of the horncore (Fig. 6.5, 6.6), which is also seen in the postorbital horncores of Albertaceratops nesmoi Ryan, Reference Ryan2007. Spiclypeus shipporum Mallon et al., Reference Mallon, Ott, Larson, Iuliano and Evans2016 also has a step at the base of the right postorbital horncore, but the step is restricted to the posterior side of the horncore, and it appears to be formed by extensive pitting rather than overgrowth. The holotype horncores of ‘Ceratops montanus’ (Marsh, Reference Marsh1888) from the Judith River Formation (Mallon et al., Reference Mallon, Ott, Larson, Iuliano and Evans2016) do not have this overgrowth. On the ventral side of the horncore, there is a round foramen that is suggestive of resorption (Fig. 6.5), which is known to occur in postorbital horncores of various eucentrosauran centrosaurines, such as Coronosaurus (Ryan and Russell, Reference Ryan and Russell2005), Spinops (Farke et al., Reference Farke, Ryan, Barrett, Tanke, Braman, Loewen and Graham2011), Centrosaurus (Ryan et al., Reference Ryan, Russell, Eberth and Currie2001; Tanke and Farke, Reference Tanke and Farke2006), Styracosaurus (Ryan et al., Reference Ryan, Holmes and Russell2007), and Einiosaurus (Sampson, Reference Sampson1995), but not in non-Eucentrosaura centrosaurines.
Nasal
The nasals of Medusaceratops lack a distinct vertical horncore but instead have ornamentation that is in the form of a low, elongate, rugose ridge. No new nasal material can be added to the sample described by Ryan (Reference Ryan2007). We re-examined the available nasals from the bonebed, but have no significant morphological observations beyond those in Ryan (Reference Ryan2007); we therefore direct the reader to the appropriate section of that publication.
Osteohistological ontogenetic assessment
In order to assess the potential that the material from the Mansfield bonebed represents the full ontogenetic sequence of this taxon, the distal end of a left tibia from the bonebed (ROM 67873) was histologically sampled at its minimum diaphyseal circumference. This specimen represents one of the largest individuals among the bonebed material (minimum circumference=300 mm). The mid-diaphysis is very robust and the lateral half of the distal end, where the calcaneum articulated, is extended distally compared to the medial half, which is characteristic of ceratopsids. Although it cannot be unequivocally referred to Medusaceratops, it is unequivocally ceratopsid, and, given that no other ceratopsids can be confirmed from the locality, we infer that this specimen represents the tibia of Medusaceratops.
The mid-diaphyseal thin section of ROM 67873 is mainly composed of densely vascularized secondary bone (Fig. 8). A lenticular medullary cavity (~3 cm × 2.5 cm) is decentered posteriorly. A 0.5–1 cm layer of trabecular bone consistently surrounds the medullary cavity, except on the lateral side where the trabecular bone spans 3 cm. The cortical bone is distributed relatively consistently on the posterior half of the shaft (2 cm), but gradually thickens towards the anteriormost part where cortical bone thickness reaches ~3 cm. The cortical bone is composed of heavily remodeled Haversian bone with occasional Volkmann’s canals. Therefore, primary bone tissue is severely obliterated, except in a thin peripheral region ~5 mm from the outer periosteal margin. The primary bone tissue along the outer periphery of the bone is comprised of poorly vascularized, parallel-fibered bone tissue with multiple growth lines that likely represent closely packed lines of arrested growth (LAGs), although any given growth mark cannot be traced around the entire periphery of the bone. Spacing between these growth lines rapidly narrows towards the outer periosteal margin, and forms an external fundamental system (EFS; Fig. 8.3). Extensive remodeling, parallel-fibered bone, and the presence of an EFS indicate that this individual had reached its asymptotic body size. This indicates that the largest individuals in the Mansfield bonebed were likely mature individuals, and allows comparisons of body size with other later ceratopsids (see Discussion).

Figure 8 Thin-section photographs of mid diaphyseal cross-section of a tibia from the Mansfield bonebed (ROM 67873). Whole cross section images under (1) plane polarized and (2) cross-polarized light; (3) close-up images under plane polarized (left half) and cross-polarized light (right half).
Phylogenetic analysis
Medusaceratops could be coded for 64 out of 101 characters in the phylogenetic analysis (Supplemental data). The analysis recovered 630 most parsimonious trees, each with 181 steps. The Consistency Index and Retention Index of each most parsimonious tree is 0.624 and 0.791, respectively. In the strict consensus tree (Fig. 9), Medusaceratops lokii is supported by three autapomorphies (character 59[2], elongate flattened ep 2; character 60[1], laterally curved ep 2; and, character 62[1] laterally curved ep 3). Medusaceratops is nested within Centrosaurinae with four unambiguous synapomorphies (character 1[0], a rostral with short dorsal and ventral processes; character 2[1], a semicircular premaxillary septum; character 8[1], caudoventral expansion of a premaxilla, and character 51[1], a relatively wide parietal midline bar) and one ambiguous character (character 3[1], the septum composed of premaxilla and nasal).

Figure 9 Strict consensus tree of the phylogenetic analysis in this paper. Numbers on nodes are decay indices (above) and bootstrap supports (below). Additional information available in supplemental data.
Medusaceratops is recovered in a polytomy with Albertaceratops, Wendiceratops + Sinoceratops, and the newly defined clade Eucentrosaura. In our analysis, this new clade includes Centrosaurini (sensu Maiorino et al., Reference Maiorino, Farke, Kotsakis, Teresi and Piras2015 and Ryan et al., Reference Ryan, Holmes, Mallon, Loewen and Evans2017; Rubeosaurus, Styracosaurus, Spinops, Centrosaurus, Coronosaurus), Einiosaurus, and members of Pachyrostora with nasal bosses (sensu Fiorillo and Tykoski, Reference Fiorillo and Tykoski2012). Eucentrosaura does not include Xenoceratops, Wendiceratops, or Sinoceratops, unlike Sampson et al. (Reference Sampson, Lund, Loewen, Farke and Clayton2013) and Lund et al. (Reference Lund, O’Connor, Loewen and Jinnah2016a, Reference Lund, Sampson and Loewen2016b), where these taxa were recovered in Pachyrhinosaurini. The members of Eucentrosaurua here have five synapomorphies: 17[1], maxillary tooth row at the same level as the rostral edentulous portion of the maxilla; 20[1], presence of a distinct nasal horncore; 28[0], short postorbital horncore; 66[1], presence of ep 6; 68[1], presence of ep 7. Xenoceratops is recovered as the sister taxon to the least inclusive clade containing Medusaceratops and Pachyrostra in the strict consensus tree. The overall morphology of the strict consensus tree is similar to that of Evans and Ryan (Reference Evans and Ryan2015) and Ryan et al. (Ryan et al., Reference Ryan, Holmes, Mallon, Loewen and Evans2017), except for the loss of resolution in the strict consensus tree within the Centrosaurini.
The strict consensus topology recovered in this analysis is generally weakly supported, as is typical of most recent centrosaurine phylogenetic analyses (e.g., Evans and Ryan, Reference Evans and Ryan2015). Within Ceratopsidae, the Bremer Decay Index is only high (>2) at the bases of Chasmosaurinae, Centrosaurinae, Pachyrhinosaurini, and the genus Pachyrhinosaurus, with most nodes within Centrosaurinae having a Bremer Decay value of 1. Bootstrap values shows similar patterns of relative support (Fig. 9).
The constrained analysis, in which Medusaceratops lokii is inferred to be a chasmosaurine, resulted in eight extra steps in the most parsimonious trees compared to the original analysis where it is recovered nested within Centrosaurinae.
Discussion
The reassignment of Medusaceratops to Centrosaurinae resolves the unusual occurrence of pachyostotic frill ornamentation in the early chasmosaurines, to which it previously had been referred (Ryan, Reference Ryan2007), and supports the observation of Longrich (Reference Longrich2013), who suggested that Medusaceratops was a member of Centrosaurinae based on the architecture of the parietal-squamosal contact.
In the original description of Medusaceratops (Ryan et al., Reference Ryan, Russell and Hartman2010), the number of the epiparietals on the holotype and the paratype parietals was counted as three (Fig. 4.2). Because many chasmosaurines have three epiparietals on each side of the midline (Dodson et al., Reference Dodson, Forster and Sampson2004), and all known centrosaurines have at least five epiparietals, Medusaceratops was described as a member of Chasmosaurinae (Ryan et al., Reference Ryan, Russell and Hartman2010). However, it was an unusual member of this clade due to the presence of hypertrophied epiparietal hooks, which are otherwise typical of centrosaurines. Longrich (Reference Longrich2013) argued that the utility of the number of epiparietals for distinguishing the two ceratopsid subfamilies is debatable, and cited the morphology of the parietosquamosal joint to argue for a centrosaurine affinity for Medusaceratops, but did not conclude if Medusaceratops was conspecific with Albertaceratops, or if it was a distinct centrosaurine species. The lack of a posterior midline ramus associated with either type specimen of Medusaceratops made it unclear up to that point whether the whole array of epiparietals was unknown for this taxon.
Description of the newly acquired material, including the first midline parietal ramus, and subsequent reexamination of the type specimens, provides new information that indicates Medusaceratops is a distinct species of centrosaurine ceratopsid. The holotype is now reinterpreted to have five epiparietals (Character 57[1]), a character changing the state at the base node of Centrosaurinae in the strict consensus tree. In addition, the new parietals exhibit diagnostic centrosaurine traits, including a broad midline ramus on ROM 73832, imbricated epiparietals on ROM 73836 (also seen on the previously described specimens, WDC-DJR-001 and TMP 2002.069.0005), and a convex squamosal contact on ROM 73836 (also seen on WDC-DJR-001). The constrained analysis, in which Medusaceratops lokii is inferred to be a chasmosaurine, resulted in a hypothesis of eight extra steps compared to the original analysis, strongly supporting Medusaceratops as a member of Centrosaurinae.
Our new reconstruction of the parietal ornamentation of Medusaceratops allows for more detailed comparison to Albertaceratops and other ceratopsids. Medusaceratops is most similar to Xenoceratops in terms of having a small dorsally projecting ep 1 and a large pachyostotic and laterally projecting ep 2; however, the ep 2 of Xenoceratops is straight in the two known specimens, unlike the strongly curved ep 2 of Medusaceratops. Although we assigned epiparietal numbers from the midline laterally, following the character coding methodology of Evans and Ryan (Reference Evans and Ryan2015), the laterally oriented ep 2 of Medusaceratops is morphologically similar to the massive ep 1 of Albertaceratops. Therefore, it is possible that these two epiparietals could be homologous (i.e., the ep 1 is not developed in Albertaceratops as has been inferred for Pachyrhinosaurus), which is congruent with the interpretation of Albertaceratops parietal ornamentation in Farke et al. (Reference Farke, Ryan, Barrett, Tanke, Braman, Loewen and Graham2011), who assigned the large pachyostotic epiparietal of this taxon to ep 2. More work on the homology of ceratopsid frill ornamentation is needed to resolve these issues (e.g., Farke et al., Reference Farke, Ryan, Barrett, Tanke, Braman, Loewen and Graham2011).
We also note that the ep 1 is expressed differently on ROM 73832, where it occurs on the dorsal surface of the parietal ramus medially and extends to the posterior surface laterally, than on the holotype, where it is so much smaller that it was not recognized as an epiparietal by Ryan et al. (Reference Ryan, Russell and Hartman2010). The basal centrosaurines Wendiceratops and Xenoceratops also exhibit a high degree of intraspecific plasticity in epiparietal size and morphology. For example, in the holotype of Xenoceratops, the morphology of ep 1 is asymmetrical. On the left side, it is dorsally curved and larger than on the right side, where it does not appear to curve dorsally. In Wendiceratops, the size of the lateral epiparietals varies considerably from the subadult specimen (TMP 2011.051.0019) compared to the holotype (TMP 2011.051.0009). Similarly, eucentrosaurans can display a large degree of intraspecific variability in mature size, shape, and symmetry of their most-prominent epiparietals. For example, the ep 1 of Centrosaurus apertus (e.g., AMNH 5239, Brown, Reference Brown1914a; CMN 971, Frederickson and Tumarkin-Deratzian, Reference Frederickson and Tumarkin-Deratzian2014), Styracosaurus albertensis (Ryan et al., Reference Ryan, Holmes and Russell2007, fig. 14B), and ep 3 of Pachyrhinosaurus lakustai Currie, Langston, and Tanke, 2007 (Currie et al., 2007, figs. 32, 33) exhibits considerable variation, but this clade is generally very conservative within the other epiparietals. Thus, it is possible that a high degree of plasticity across all epiparietals may be plesiomorphic for Centrosaurinae, with this variability being primarily limited to only the prominently modified, more medially positioned epiparietals in eucentrosaurans. A future quantitative study of intraspecific frill variability may lead to refinement of ceratopsid systematics and phylogenetics studies.
The large size of the thin-sectioned tibia and the postorbital horncores of Medusaceratops are notable for an early ceratopsid. The size range of Medusaceratops limb elements, as well as the humeral size of the penecontemporaneous centrosaurine Wendiceratops (Evans and Ryan, Reference Evans and Ryan2015), are comparable to the late Campanian centrosaurines such as Centrosaurus and Styracosaurus, as well as the non-Triceratopsini chasmosaurines from the Belly River Group and correlative strata of the Judith River Formation (Fig. 10; Table S3). Centrosaurus apertus Lambe, Reference Lambe1904 and Styracosaurus albertensis have adult basal skull lengths that range in size from 666 mm to 786 mm (Table S3), with associated estimated body masses between 2,500 and 4,800 kg (body mass estimation based on sum of humerus and femur circumference using an interspecific limb scaling equation provided in Campione and Evans, Reference Campione and Evans2012; Table S3). Earlier ceratopsids, notably the middle Campanian (~79 Ma) Diabloceratops (basal skull length=620 mm; Kirkland and DeBlieux, Reference Kirkland and DeBlieux2010) and Machairoceratops, the holotype of which is interpreted as being approximately the same size as Diabloceratops (Lund et al., Reference Lund, O’Connor, Loewen and Jinnah2016a), are smaller than Centrosaurus and Styracosaurus. The large body size of Medusaceratops and Wendiceratops extends the fossil record of large-bodied ceratopsids into the middle Campanian and may have implications for the paleobiology of these taxa.

Figure 10 Comparison of humerus and tibia circumference of ceratopsids from the Belly River Group and the Judith River Formation. In the box plots, mean values are represented by lines in the boxes, lower and upper bounds of the boxes represent the first and third quartiles, and the ends of the dashed lines indicate minimum and maximum values of the data. Data used for this plot are provided in Table S3. Open circles and triangles represent Medusaceratops and Wendiceratops, respectively.
After examination of all available material, no unequivocal chasmosaurine bones, or diagnostic material from any other ceratopsid, could be identified in the Mansfield bonebed collections, suggesting that it represents a monodominant accumulation of a single centrosaurine taxon, Medusaceratops. The stratigraphic positions of the Mansfield Medusaceratops bonebed within the lower half of the Judith River Formation at Kennedy Coulee, Montana, the chronostratigraphically equivalent Wendiceratops bonebed from the lower Oldman Formation of Alberta (Evans and Ryan, Reference Evans and Ryan2015), and a low-density bonebed of Xenoceratops from the slightly chronostratigraphically older Foremost Formation (Ryan et al., Reference Ryan, Evans and Shepherd2012) mark some of the oldest occurrences of centrosaurine bonebeds. Monodominant centrosaurine bonebeds become more abundant in slightly higher stratigraphic units (e.g., Coronosaurus brinkmani from younger strata of the Oldman Formation; Ryan and Russell, Reference Ryan and Russell2005), and are common in the Dinosaur Park Formation and younger strata in Alberta, Montana, and Alaska. The detailed taphonomic studies of the Centrosaurus apertus bonebeds in the Belly River Group indicate that these deposits represent mass death assemblages of massive herds of centrosaurs (Ryan et al., Reference Ryan, Russell, Eberth and Currie2001; Eberth and Getty, Reference Eberth and Getty2005; Eberth et al., Reference Eberth, Brinkman and Barkas2010; Chiba et al., Reference Chiba, Ryan, Braman, Eberth, Scott, Brown, Kobayashi and Evans2015). To date, detailed taphonomic analyses have yet to be carried out on the Medusaceratops and Wendiceratops bonebeds, but such studies could lead to important insights into the palaeobiology of early ceratopsids. Previous taphonomic work on more derived taxa, and examples of basal neoceratopsian and Zuniceratops bonebeds (Hunt and Farke, Reference Hunt and Farke2010; Wolfe et al., Reference Wolfe, Kirkland, Smith, Poole, Chinnery-Allgeier and McDonald2010) suggest that large-scale gregarious behavior has deep evolutionary roots in Ceratopsia and may be plesiomorphic for Centrosaurinae.
Acknowledgments
We thank D. Brinkman, B. Strilisky, G. Housego, B. Bavington-Sanchez, T. Courtenay, and R. Russell for specimen access in Royal Tyrrell Museum of Palaeontology. A. Guyon and L. Shinkle provided specimen access in the Wyoming Dinosaur Center, and C. Levitt-Bussian for specimen access at the Natural History Museum of Utah. M. Shibata, T. Sekiya, T. Sonoda, S. Kawabe, H. Yukawa facilitated specimen access in the Fukui Prefectural Dinosaur Museum. We thank A. Dzindic (Canada Fossils Ltd.) for his care in preparing many of the specimens, and I. Morrison for preparation at the ROM, as well as J. Issa (Canada Fossils Ltd.) for specimen loans and facilitating specimen acquisition. We thank D. Trexler (Two Medicine Dinosaur Center) for providing the information on WDCB-MC-001. We are grateful to D. and L. Redding, and M. Goodwin (University of California Museum of Paleontology) for fieldwork logistics, and T. Cullen (University of Toronto) for assistance in the field. Thanks to K. Seymour and B. Iwama for logistical support at the ROM. We thank A. Farke for a constructive review, and K. Brink and D. Eberth (Royal Tyrrell Museum of Palaeontology) for discussions and editorial assistance.
The Dinosaur Research Institute for provided a travel grant to KFC to present this study at the Society of Vertebrate Paleontology annual meeting in 2016. The Louise Hawley Stone Charitable Trust and Royal Ontario Museum provided funding to acquire specimens for this project. DCE was supported by a Natural Sciences and Engineering Research Council of Canada Discovery Grant (NSERC Grant File Number: RGPIN 355845). Fieldwork associated with this research was supported by the Royal Ontario Museum Reproductions Fund and the ROM Collections & Research Fieldwork Fund. The Willi Henning Society provided access to TNT.
Accessibility of supplemental data
Data available from the Dryad Digital Repository: http://doi.org/10.5061/dryad.8h067
Supplemental data include the following:
Text S1. List of the Mansfield bonebed specimens used for character coding.
Table S1. Specimen list of the Mansfield bonebed material housed in museum collections.
Table S2. Compiled postorbital horncore measurements of Centrosaurinae, non-Triceratopsini Chasmosaurinae, and Triceratopsini.
Table S3. Compiled humerus and tibia circumference measurements of ceratopsids from Belly River Group and Judith River Formation (Campanian).
Supplemental data 1. Data matrix used in phylogenetic analysis in this study (TNT file).
Supplemental data 2. 3D data of a Medusaceratops parietal (ROM 73832, STL file).
Supplemental data 3. 3D data of a Medusaceratops parietal (ROM 73837, STL file).
Supplemental data 4. 3D data of a Medusaceratops postorbital (TMP 2002.069.0010, STL file).
Supplemental data 5. 3D data of a Medusaceratops postorbital (ROM 73834, STL file).
Supplemental data 6. 3D data of a Medusaceratops postorbital (ROM 73831, STL file).
Supplemental data 7. 3D data of a Medusaceratops postorbital (WDC-DJR-003, STL file).
Supplemental data 8. Thin section photograph of mid diaphyseal cross-section of a Medusaceratops tibia (ROM 67873) with plane polarized light (JPG file).
Supplemental data 9. Thin section photograph of mid diaphyseal cross-section of a Medusaceratops tibia (ROM 67873) with cross polarized light (JPG file).










