



Introduction and history of research
Little work has been done on the Early Devonian (late Emsian) placoderm fishes of Belarus despite these being relatively common in Belarus and their use in regional biostratigraphy elsewhere in the Old Red Sandston continent. Specimens from the orders Acanthothoraci, Ptyctodontida, Arthrodira, and Antiarcha are all recorded from the Lower Devonian deposits of Belarus (Lyarskaya, Reference Lyarskaya and Sorokin1978; Valiukevičius et al., Reference Valiukevičius, Karatajūtė-Talimaa and Kruchek1995; Mark-Kurik, Reference Mark-Kurik, Satkūnas and Lazauskienė2002; Plaksa, Reference Plaksa2008a, Reference Plaksa, Bogdanova and Krymgoltsb; Plax, Reference Plax2008, Reference Plax, Gubin, Zui, Lukashev and Sanko2015a, Reference Plaksa, Bogdanova and Krymgoltsb, Reference Plax2016, Reference Plax2017). Arthrodires are the most common placoderm group found in the upper Emsian deposits of Belarus. The first recorded find of an arthrodire in the Emsian of Belarus was in 1963 (personal communication, E. Mark-Kurik, 2015) from the Vilchitsy 1 borehole in the Obol and Lepel beds of the Vitebsk Regional Stage. These remains consisted of a skull roof, trunk shield, and a number of isolated plates from the trunk shield, as well as endocranium remains. Subsequently, Mark-Kurik considered the remains in a single species and referred to it as “Kartalaspis belarusica.” She considered it a phlyctaeniid placoderm. However, the species was not formerly described despite it being mentioned in many publications (Mark-Kurik, Reference Mark-Kurik, Satkūnas and Lazauskienė2002) and so it must be considered a nomen nudum. Even the species name of this fish in the literature has been spelled in several ways, e.g., belorussica, byelorussica, and belarussica (Mark-Kurik, Reference Mark-Kurik, Satkūnas and Lazauskienė2002), adding to the confusion. It is in need of being properly described.
Later, Lyarskaya (Reference Lyarskaya and Sorokin1978) recorded Ptyctodontida indet., Phyctaeniina indet., and Antiarcha indet. in the Vitebsk Formation of the Orsha Depression (in the Vilchitsy 1, Pochtari 1 and Liozno 1 boreholes).
Valiukevičius et al. (Reference Valiukevičius, Karatajūtė-Talimaa and Kruchek1995) recorded Millerosteus orvikui (Gross, Reference Gross1940) from the Mstislavl 1 borehole in the Lepel Beds of the Vitebsk Regional Stage. However, Gross (Reference Gross1940) erected this species from strata of Eifelian age (Kernavė Regional Substage) of the Baltic region. Furthermore, this species is now known considered a subjective junior synonym of Coccosteus cuspidatus Miller ex Agassiz, Reference Miller1841 (Mark-Kurik, Reference Mark-Kurik2000; Newman et al., Reference Newman, Mark-Kurik, den Blaauwen and Zupiņš2015). The genus Millerosteus Stensiö, Reference Stensiö1959 is known from the Givetian (Newman et al., Reference Newman, Mark-Kurik, den Blaauwen and Zupiņš2015) of both Estonia and Scotland; it thus seems unlikely that it is present in the Emsian of Belarus and the remains recorded by Valiukevičius et al. (Reference Valiukevičius, Karatajūtė-Talimaa and Kruchek1995) must be considered for now as indeterminate.
The first author of this paper started a study of the upper Emsian deposits of Belarus in 2002, and has reviewed and studied ~25 boreholes containing deposits of this age. Remains of late Emsian placoderms in various states of preservation were recovered from 13 boreholes, all within the Lepel Beds of the Vitebsk Regional Stage. Acanthothoracid remains were recovered from the Berdyzh 1 borehole (Plax, Reference Plax2015b). Ptyctodontida remains were recovered from the Chashniki 53, Lepel 1, and Latvishi 12 boreholes (Plax, Reference Plax2019). One undefinable plate fragment of a ptyctodontid was also recovered from the rocks of the Lepel Beds of the Vitebsk Regional Stage in the Rudnya 14 borehole. Arthrodiran remains are the most commonly recovered remains found in boreholes. They have been recovered from the Osipovichi 6, Bobruysk 691, Bobruysk 691/2, Lepel 1, Bykhov 151, Rogachev 736, Buda Dal'nyaya 35, Bykhov 1, and Korma 1 boreholes (unpublished data, Plax, 2018). Finally, a few antiarch remains have been recovered from the Osipovichi 6, Buda Dal'nyaya 35, Berdyzh 1, Bykhov 1, and Korma 1 boreholes.
In this work, we erect two new species of arthrodire and describe two other forms in open nomenclature.
Geological setting and stratigraphy
All of the specimens described in this work were collected from core samples extracted from the Lepel Beds. The Lepel Beds form the upper part of the Vitebsk Regional Stage, with the lower part being the Obol Beds. The Vitebsk Regional Stage constitutes the upper Emsian (Lower Devonian) in Belarus. These deposits are widespread throughout Belarus. The Eifelian Adrov Regional Stage lies above the Vitebsk Regional Stage. The Lepel Beds correspond to the upper part of the Rhabdosporites mirus-Gneudnaspora divellomedium Miospore Biozone and approximately to the upper part of the Polygnathus costatus patulus Conodont Biozone (Obukhovskaya et al., Reference Obukhovskaya, Kruchek, Pushkin, Nekryata, Plax, Sachenko, Obukhovskaya and Antipenko2010) (Table 1). This is approximately equivalent to the upper part of the Rezēkne Regional Stage in the Baltic region (Lyarskaya, Reference Lyarskaya and Sorokin1978) and the upper part of the Novobasovo Beds of the Ryazhsk Regional Stage in the central part of Russia (Valiukevičius and Kruchek, Reference Valiukevičius and Kruchek2000; Vorotnikova et al., Reference Vorotnikova, Gavryushova, Drutskoy, Zhuravlev, Kirikov, Lavrovich, Lavrovich, Myazina, Osipova, Papin, Petrov, Rusetskaya, Ryborak and Shokurova2011). Further details of the fauna found within the core samples are recorded below.
Table 1. Stratification of the Upper Emsian deposits of Belarus and their correlation with the synchronous deposits from the adjacent territories (Obukhovskaya et al., Reference Obukhovskaya, Kruchek, Pushkin, Nekryata, Plax, Sachenko, Obukhovskaya and Antipenko2010).

Identifiable remains of Stipatosteus svidunovitchi n. gen. n. sp. were collected from the Korma 1 borehole (Fig. 1) drilled in the territory of the Zhlobin Saddle at depths of 340.8 m, 340.2 m, 337.8 m, 337.9 m, and 337.7 m. Associated with these remains were isolated scales of Diplacanthus kleesmentae Valiukevičius in Valiukevičius and Karatajūtė-Talimaa, Reference Valiukevičius, Karatajūtė-Talimaa and Brangulis1986, Cheiracanthus brevicostatus Gross, Reference Gross1973, Cheiracanthus longicostatus Gross, Reference Gross1973, Cheiracanthus cf. C. splendens Gross, Reference Gross1973, plus indeterminable otoliths and remains of other acanthodians, chondrichthyans, and sarcopterygians. Rare, small fragments of lingulid shells and ostracodes were also found in these deposits.
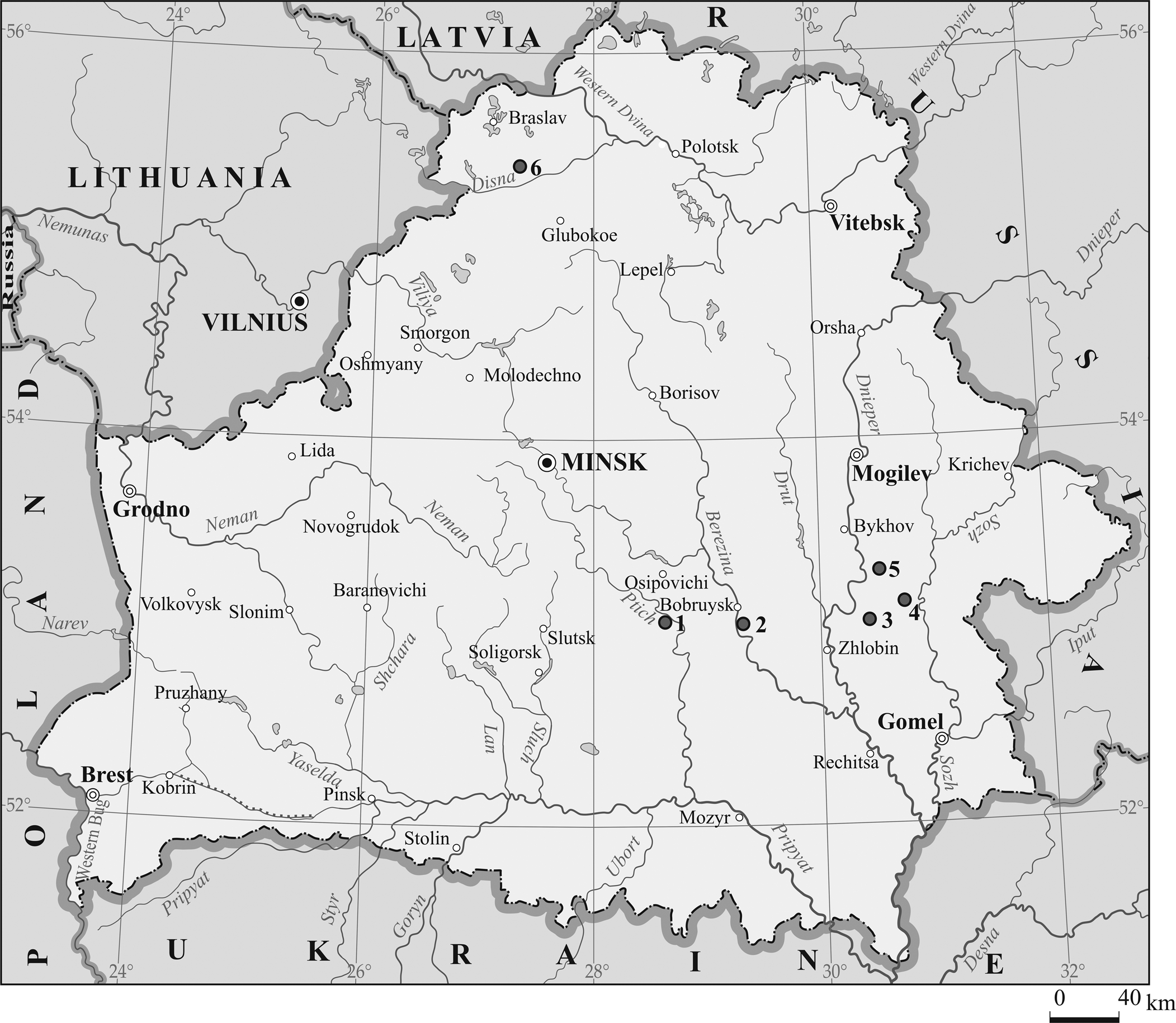
Figure 1. Map of Belarus showing the location of the borehole sections where the skeletal elements of the placoderm fishes were found. Boreholes: 1 = Osipovichi 1; 2 = Bobruysk 691; 3 = Rogachev 736; 4 = Korma 1; 5 = Bykhov 1; 6 = Buda Dal'nyaya 35.
Remains of Actinolepis zaikai new species were collected from the Osipovichi 6 borehole drilled within the Bobruysk Buried Ridge at depths of 118.8 m, 113.8 m, and 113.5 m. Associated with these remains were a plate fragment of Psammolepis cf. P. toriensis (Mark-Kurik in Obruchev and Mark-Kurik, Reference Obruchev and Mark-Kurik1968), isolated scales of Laliacanthus singularis Karatajūtė-Talimaa in Valiukevičius and Karatajūtė-Talimaa, Reference Valiukevičius, Karatajūtė-Talimaa and Brangulis1986, and remains of indeterminate arthrodires, acanthodians, sarcopterygians, and actinopterygians (Plax, Reference Plax2015b). Miospores were also found within these deposits. Several small indefinable plates with a similar ornamentation to Actinolepis zaikai n. sp. were collected from the Bobruysk 691 borehole also drilled within the Bobruysk Buried Ridge at a depth of 234.5 m. Associated with these remains were indeterminate remains of sarcopterygians and actinopterygians (Plax, Reference Plax2015b). A right anterior ventrolateral plate of Actinolepis zaikai n. sp. was collected from the Rogachev 736 borehole drilled in the region of the Zhlobin Saddle at a depth of 293.0 m. Several small plates with an ornamentation similar to Actinolepis zaikai n. sp. were collected from the Bykhov 1 borehole drilled in the Orsha Depression and Zhlobin Saddle junction zone at a depth of 307.3 m. Associated with these plates were fragments of lingulid shells, remains of P. cf. P. toriensis, isolated scales of Cheiracanthus brevicostatus, Cheiracanthus longicostatus, Cheiracanthus gibbosus Valiukevičius in Valiukevičius and Karatajūtė-Talimaa, Reference Valiukevičius, Karatajūtė-Talimaa and Brangulis1986, small isolated fragments of fin spines of Diplacanthus sp., and indeterminate remains of other acanthodians, antiarchs, sarcopterygians, and actinopterygians. Finally, several small plates with ornamentation similar to that of Actinolepis zaikai n. sp. were collected from the Korma 1 borehole at a depth of 322.3 m. Associated with these remains were nearly intact shells of lingulids, small plate fragments of Psammolepis cf. P. toriensis, isolated scales of Cheiracanthus longicostatus, Cheirolepis cf. C. gracilis Gross, Reference Gross1973, and indeterminate remains of other acanthodians, placoderms, sarcopterygians, and actinopterygians.
Remains of Coccosteidae gen. indet. sp. indet. were collected from the Buda Dal'nyaya 35 borehole drilled within the Vileyka Buried Ridge of the Belarusian Anticlise at a depth of 233.2 m. Associated with these remains were scales of Laliacanthus singularis, Cheiracanthus brevicostatus, Cheiracanthus longicostatus, Cheiracanthus cf. C. splendens, Rhadinacanthus primaris Valiukevičius in Valiukevičius and Karatajūtė-Talimaa, Reference Valiukevičius, Karatajūtė-Talimaa and Brangulis1986, Karksilepis cf. K. parva Märss, Kleesment, and Niit, Reference Märss, Kleesment and Niit2008, and indeterminable remains of psammosteids, acanthodians, antiarchs, sarcopterygians, actinopterygians, etc. (Plax, Reference Plax2016).
Remains of Placodermi indet. 1 were collected from the Bykhov 1 borehole at depths of 325.5 m, 324.3 m, 324.2 m, 321.3 m, 321.2 m, and 321.0 m. Associated with these remains were rare fragments of lingulids, numerous ostracodes, abundant conchostracans, isolated scales of Diplacanthus kleesmentae, and indeterminate acanthodians, chondrichthyans, and sarcopterygians. Remains of Placodermi indet. 1 were also collected in the Korma 1 borehole at depths of 343.5 m and 323.3 m. Associated with these remains were rare ostracodes, Cheiracanthus cf. C. splendens, and indeterminable otoliths and remains of placoderms and sarcopterygians.
Materials and methods
The material consists of disarticulated dermal plates in various states of completeness recovered from the rocks of six boreholes (Osipovichi 6, Bobruysk 691, Rogachev 736, Buda Dal'nyaya 35, Bykhov 1, and Korma 1). Most of the material was removed from the rock matrix using 9% acetic acid; a few others were mechanically removed. The preservation of the skeletal material is generally good, but the bone can be very brittle, with the edges often worn, thus not giving the true shape of the plate. The specimens have been prepared as far as is possible without damaging them.
Photography was done using a PowerShot SX130 IS camera and a Sony A58 camera with an Industar-50 lens. The electron microphotographs of the plate fragments were made with a JSM-5610 LV (JEOL, Japan) scanning electron microscope. The images were processed with Adobe Photoshop СS6; the figures were created with CorelDRAW X3. An MBS-1 microscope was also used during study of the skeletal material.
Repository and institutional abbreviation
All specimens are housed in the Belarusian National Technical University (BNTU), Department of Mining, Minsk.
Systematic paleontology
Class Placodermi M'Coy, Reference M'Coy1848
Order Arthrodira Woodward, Reference Woodward1891
Suborder Phlyctaenioidei Miles, Reference Miles1973
Family Phlyctaeniidae Fowler, Reference Fowler1947
Type species
Stipatosteus svidunovitchi n. gen. n. sp. by monotypy.
Diagnosis
As for Stipatosteus svidunovitchi, the type and only species.
Etymology
From the Latin, stipa, packed; and osteus, bone.
Remarks
Stipatosteus n. gen. differs from all other phlyctaeniid genera by its short ventral trunk armor. The anterior ventrolateral plates are shorter than the posterior ventrolateral plates. These are also very narrow compared with those of other genera and have a deep embayment along their mesial border.
Holotype
BNTU 121/38-1, a right anterior ventrolateral plate with attached right spinal plate; from the Korma 1 borehole, depth 337.7 m, from the Gomel region, Belarus; also from the Lepel Beds of the Vitebsk Regional Stage, which is dated upper Emsian of the Early Devonian.

Figure 2. Stipatosteus svidunovitchi n. gen. n. sp., dermal plate remains: (1–3) indefinable dermal plate fragments: (1) BNTU 121/44-23; (2) BNTU 121/44-18; (3) BNTU 121/44-16; (4) right anterior lateral plate (BNTU 121/38-2); (5–7) holotype, a right anterior ventrolateral plate with attached right spinal plate (BNTU 121/38-1): (5) ventral view with impression of visceral armor; (6) view of the visceral armor; (7) detail of right spinal plate (Fig. 2.5); (8, 9) incomplete right spinal plate (BNTU 121/46-1): (8) probable dorsal view; (9) probable ventral view; (10, 11) both sides of incomplete spinal plate, orientation unclear (BNTU 121/46-3); (12) left posterior ventrolateral plate (BNTU 121/40-1). gl = growth lines. Scale bars = 5 mm (4–6, 8–12), 1 mm (1, 7), 0.5 mm (2, 3).

Figure 3. Stipatosteus svidunovitchi n. gen. n. sp., reconstruction of ventral trunk armor. AMV = anterior median ventral plate; AVL = anterior ventrolateral plate; IL = interolateral plate; PMV = posterior median ventral plate; PVL = posterior ventrolateral plate; Sp = spinal plate. Dashed lines indicate the boundaries of unknown plates.
Diagnosis
Very small phlyctaeniid with an estimated total length of ~40–50 mm. Ventral trunk armor short with a breadth/length index of 1.0. Posterior ventrolateral plates slightly longer than the anterior ventrolateral plates. Fairly long, slender spinal plates with the free distal ends of approximately the same length as the part attached to the anterior ventrolateral plate. Posterior ventrolateral plate narrow, with a breadth/length index of 0.8. Ornamentation consisting of closely packed tubercles often forming rows extending subparallel to the lengths of the individual plates.
Occurrence
All specimens came from the Korma 1 borehole at depths of 340.2 m, 340.8 m, 337.7 m, 337.8 m, and 337.9 m, with the locality and horizon details specified above.
Description
Only four near complete identifiable plates are known, all from the trunk armor. These include a right anterior lateral plate, a right anterior ventrolateral plate with attached right spinal plate, and a right posterior ventrolateral plate. There are also fragments of spinal plates and other unidentifiable plate fragments. All plates are characterized by ornamentation of densely packed tubercles forming distinct rows (Fig. 2.1). The tubercles are star-shaped at their bases (Fig. 2.2). Some of the individual tubercles have an unusual crown ornament of sinuous, bifurcating ridges emanating from the center (Fig. 2.3). This is due to tubercles overgrowing older tubercles, indicating that these plates do not belong to juvenile individuals (see comparison below). The individual plates are described below.
BNTU 121/38-2 (Fig. 2.4) is a right anterior lateral plate (AL) with a broken posterior edge. The edge contacting the spinal plate is also slightly broken and irregular. This makes it difficult to estimate a breadth/length index. The ornamentation consists of tightly packed tubercles that extend in rows subparallel to the outline of the plate. The anterior ventral corner has a significant process pointing anteriorly from which extrudes a ridge. The anterior dorsal corner is rounded with the dorsal edge extending diagonally ventrally.
BNTU 121/38-1 (Fig. 2.5, 2.6) is a right anterior ventrolateral plate (AVL) attached to a right spinal plate (Sp) (Fig. 2.7) preserved in part and counterpart. The plate is worn at the edges, but its overall shape is discernible. The plate is quite long with a breadth/length index of ~0.8. The plate is quite flat-bottomed. The plate is very rounded mesially and anteriorly with no evidence for a contact face for an anterior ventral plate. Little or no ornamentation is preserved on the part (Fig. 2.5) although it is present on the attached Sp. The suture between the AVL and Sp is difficult to discern. The rest of the specimen has an impression of the visceral surface of the bone. The counterpart (Fig. 2.6) shows the visceral side of the plate. The bone is at full thickness at the center of the plate but is worn and delaminated toward the edges. Growth lines (gl) can be seen (Fig. 2.5), which indicate that the bone grew from the center of the plate. The visceral surface has a ridge that extends from the posterior contact of the spinal plate a short distance into the plate.
BNTU 121/46-1 (Fig. 2.8, 2.9) is the most complete of the known isolated spinal plates (Sp), and the species description is mostly based on this specimen. It is preserved in three dimensions, and oval in transverse section. It is probably a right Sp because one surface is smoother than the other. The anterior section of the ‘free’ part of the plate is preserved with little or no contact face to the anterior ventrolateral plate visible. The tip of the distal end is also missing. BNTU 121/46-1 is long and slender with a gentle mesial curve. Rows of rounded tubercles extend the length of the spinal plate. There are no spinelets on the inner edge of the plate. Similar ornamentation is seen in BNTU/46-3 (Fig. 2.10, 2.11), which shows BNTU 121/46-1 is neither worn nor aberrant. BNTU 121/38-1 (Fig. 2.5–2.7) is a third right Sp attached to an anterior ventrolateral plate (see below) and shows similar ornamentation (Fig. 2.7). However, it has been crushed flat and shows no curvature. Also, it is not clear how complete it is. Certainly, some of the free end is missing. It has a long contact with the anterior ventrolateral plate at about half of its preserved length.
BNTU 121/40-1 (Fig. 2.12) is a left posterior ventrolateral plate (PVL) with preserved lateral and ventral laminae. The plate is fairly complete with a breadth/length index of at least 0.5, although the posterior end of the plate is missing. The ventral lamina is approximately twice as wide as the lateral lamina and is as long as two thirds of the ventral lamina. The ventral lamina is flat-bottomed. The ventrolateral ridge is quite pronounced posteriorly. Where preserved, the plate is densely packed with tubercles. The visceral side is still buried within the matrix.
Etymology
The species is named after Igor P. Svidunovitch (Minsk, Belarus), who repeatedly actively participated in the sampling of paleoichthyological material from the cores of boreholes.
Materials
The holotype, a right anterior lateral plate, BNTU 121/38-2; a right posterior ventrolateral plate, BNTU 121/40-1; two spinal plates, BNTU 121/46-1 and 121/46-3; a small fragment of a spinal plate, BNTU 121/46-2; and indefinable plate fragments, BNTU 121/41-12, 121/44-16, 121/44-18, 121/44-21, 121/44-22, and 121/44-23.
Remarks
Because a number of the ventral trunk plates are known, we are able to reconstruct the ventral trunk armor (Fig. 7). It is clear the trunk armor is quite short with a breadth/length index of ~1.0 at the anterior ventral plate's widest points. The anterior ventrolateral plates are particularly short, and shorter than the posterior ventrolateral plates. Another distinctive character is the long spinal plates, which are only gently curved without mesial denticles. The free end of the spinal plate is approximately the same length as the spinal plate attached to the anterior ventrolateral plate. These characters are incompatible with Stipatosteus svidunovitchi n. gen. n. sp. being a petalichthyid. The ventral armor is most similar to that of the arthrodiran infraorder Actinolepina, but the lack of an anterior ventral plate clearly places it in the suborder Phlyctaenioidei. The long, thin spinal plates with long, thin, free distal ends are characteristic of the Phlyctaeniidae.
Approximately 18 genera are known in the Phlyctaeniidae with the highest diversity being known from the Early Devonian of Spitsbergen and Campbellton in New Brunswick, Canada. Stipatosteus svidunovitchi n. gen. n. sp. displays some similarity to species of Phlyctaenius Traquair, Reference Traquair1890. There are three species known: Phlyctaenius acadicus (Whiteaves, Reference Whiteaves1881), Phlyctaenius atholi (Pageau, Reference Pageau1969), and Phlyctaenius stenosus Young, Reference Young1983. The closely related form Gaspeaspis cassivii (Pageau, Reference Pageau1969) is probably a junior subjective synonym of Phlyctaenius atholi (see Young, Reference Young1983). Stipatosteus svidunovitchi n. gen. n. sp. differs from Phlyctaenius (see Pageau, Reference Pageau1969 and Young, Reference Young1983 for descriptions) in several ways. The anterior ventrolateral plates are shorter, but also wider in Stipatosteus svidunovitchi n. gen. n. sp., making the whole trunk shorter than in Phlyctaenius. The posterior ventrolateral plates form an embayment at their posterior ends, absent in Phlyctaenius stenosus. The spinal plates are also slenderer than in Phlyctaenius. The arrangement of the tubercles is uniform and close together in both Phlyctaenius acadicus and Phlyctaenius atholi (see Young, Reference Young1983). However, in Phlyctaenius stenosus, they are arranged parallel to the plate margins (Young, Reference Young1983), similar to those in Stipatosteus svidunovitchi n. gen. n. sp. Some of the tubercles of Phlyctaenius show a similar crown ornament of sinuous ridges (Young, Reference Young1983, fig. 11) due to tubercle overgrowth as seen in Stipatosteus svidunovitchi n. gen. n. sp. Stipatosteus svidunovitchi n. gen. n. sp. only slightly differs from Phlyctaenius in having a shorter body trunk with, therefore, shorter trunk plates. The spinal plates are also narrower.
Of the Spitsbergen species, some of the genus Heterogaspis Strand, Reference Strand1932—H. acuticornis (Heintz, Reference Heintz1929), H. minutus (Heintz, Reference Heintz1929), and H. hornsundi (Heintz, Reference Heintz1929)—are somewhat similar to Stipatosteus svidunovitchi n. gen. n. sp. Stipatosteus svidunovitchi n. gen. n. sp. differs from H. acuticornis in having a longer posterior ventrolateral plate. However, as in Stipatosteus svidunovitchi n. gen. n. sp., there is an embayment at the posterior median end of the posterior ventrolateral plates (Heintz, Reference Heintz1929, fig. 29). However, the spinal plate of H. acuticornis is strongly curved, making it quite different from Stipatosteus svidunovitchi n. gen. n. sp. In H. minutus, the ventral trunk armor is unknown (Heintz, Reference Heintz1929, fig. 30) but the spinal plate is strongly curved, unlike that of Stipatosteus svidunovitchi n. gen. n. sp. Heterogaspis hornsundi is poorly known (Heintz, Reference Heintz1929, fig. 31) with many missing characters. Like other Heterogaspis species, it has strongly curved spinal plates. Thus, whereas some Heterogaspis species are similar to Stipatosteus svidunovitchi n. gen. n. sp., they differ in having strongly curved spinal plates.
Suborder Actinolepidoidei Miles, Reference Miles1973
Family Actinolepidae Gross, Reference Gross1940
Genus Actinolepis Agassiz, Reference Agassiz1844
Actinolepis zaikai new species
Figure 4
- Reference Plaksa2007
Diadsomaspis cf. D. remscheidensis Gross, Reference Gross1937; Plaksa, p. 7.
- Reference Plaksa2008a
Diadsomaspis cf. D. remscheidensis; Plaksa, p. 84.
- Reference Plaksa, Bogdanova and Krymgolts2008b
Diadsomaspis cf. D. remscheidensis; Plaksa, p. 136.
- Reference Plax2008
Diadsomaspis cf. D. remscheidensis; Plax, p. 70, table 1.
- Reference Kruchek, Obukhovskaya and Plax2012
Diadsomaspis cf. D. remscheidensis; Kruchek et al., p. 100.
- Reference Plax2013a
Diadsomaspis; Plax, p. 96.
- Reference Plax2013b
Diadsomaspis; Plax, p. 85.
- Reference Plax2013b
Holonematidae gen. indet. sp. indet.; Plax, p. 85.
- Reference Plax, Gubin, Zui, Lukashev and Sanko2015a
cf. Diadsomaspis; Plax, p.171, pl., figs. 1–3.
- Reference Plax, Gubin, Zui, Lukashev and Sanko2015a
Holonematidae gen. indet. sp. indet.; Plax, p. 171, pl., fig. 7.
- Reference Plax2015b
cf. Diadsomaspis; Plax, p. 21, table, pl. 1, figs. 9, 11, text-fig. 2.
- Reference Plax2015b
Holonematidae gen. indet. sp. indet.; Plax, p. 21, table, pl. 2, fig. 1.

Figure 4. Actinolepis zaikai n. sp., dermal plate remains: (1–4) plate fragments: (1) BNTU 44/1-7f.2; (2) BNTU 44/1-6a; (3) BNTU 44/1-16; (4) BNTU 44/1-16a; (5, 6) fused left and right preorbital plate (BNTU 44/1-12): (5) dorsal view; (6) visceral view; (7, 8) right paranuchal plate (BNTU 44/1-21): (7) dorsal view; (8) visceral view; (9) left anterior dorsolateral plate (BNTU 44/1-13); (10) anterior lateral plate (BNTU 6/1-1); (11) indefinable plate fragment (BNTU 44/1-6). af = articular flange; lc = main lateral line groove; oa = overlap area; oa.C = area overlapped by central plate; oa.MD = area overlapped by median dorsal plate; occ = occipital cross commissure canal; pp = posterior pit line; r = ridges under supraorbital sensory line on visceral side; sl = sensory line; soc = supraorbital sensory line. Scale bars = 5 mm.
Holotype
BNTU 44/1-12, a fused left and right preorbital plate; from Osipovichi 6 borehole at a depth of 118.8 m; also from the Mogilev region of Belarus and the Lepel Beds of the Vitebsk Regional Stage, which is in the upper Emsian of the Lower Devonian.
Diagnosis
An actinolepid with ornamentation of ridges topped by tubercles. The ridges are parallel to the edges of the plates, often forming concentric circles. The fused preorbital plates are long, with deep embayments anteriorly for the pineal plates. A distinct posterior process on the posterior median edge sits between the central plates. The left and right branches of the supraorbital sensory lines meet at the center of the fused preorbital plates toward the posterior. Low ridges are present on the visceral side below the supraorbital sensory lines.
Occurrence
From the boreholes Osipovichi 6 at depths of 118.8 m, 113.8 m, and 113.5 m, Bobruysk 691 at a depth of 234.5 m, and Bykhov 1 at a depth of 307.3 m, all from the Mogilev region. Also, the boreholes Rogachev 736 at a depth of 293.0 m and Korma 1 at a depth of 322.3 m, both from the Gomel region. All occurrences are within the Lepel Beds of the Vitebsk Regional Stage, which is in the upper Emsian of the Lower Devonian.
Description
Seventeen of the 22 specimens are fragmentary and not identifiable as specific plates. However, they each have a similar external ornamentation allowing a fair degree of confidence that they all belong to the same species (Fig. 4.1–4.4). The ornamentation consists of narrow ridges topped by a single row of tubercles. Five specimens are identifiable plates:
BNTU 44/1-12 (Fig. 4.5, 4.6) consists of a fused left and right preorbital plate (PrO + PRO) and is the only specimen of this plate available to study. The plate is V-shaped and fairly well preserved. It has a maximum length of 24 mm with a maximum width of 16 mm. The width/length ratio is 0.67. The dorsal surface (Fig. 4.5) has an ornamentation of narrow ridges each topped by a single row of small tubercles. These tubercles are somewhat worn. The left and right branches of the supraorbital sensory line (soc) meet in the middle of the plate toward the posterior. On the visceral surface (Fig. 4.6) are two low longitudinal ridges under the soc forming the thickest part of the plate. The visceral surface is relatively smooth on both sides of these ridges.
BNTU 44/1-21 (Fig. 4.7, 4.8) is a right paranuchal plate (PNu), irregularly oval in shape with a rounded outer margin. The ornamentation of the dorsal surface (Fig. 4.7) is formed by concentrically located, narrow ridges, each topped by a single row of tubercles. The tubercles are somewhat smoothed. Three grooves of the lateral line system pass on the external surface of the plate: the posterior pit line (pp) extending approximately from the center of ossification of the plate upward to the central plate, the main lateral line canal (lc) extending to the marginal plate and anterior dorsolateral plate, and the occipital cross commissure canal (occ). The grooves connect with each other over the center of ossification at different angles: the angle between occ and lc is ~60°, the angle between occ and pp is ~70°, and the angle between pp and lc is ~100°. The area overlapped by the central plate (oa.C) is a depression on the anterior mesial margin. The rest of the mesial margin is broken. The visceral surface (Fig. 4.8) is worn and delaminated in places.
BNTU 44/1-13 (Fig. 4.9) is the most dorsal part of a left anterior dorsolateral plate (ADL). The preserved area is triangular, with only a small ornamented area. Most of the posterior portion of the preserved plate consists of the overlap area for the median dorsal plate (oa.MD), and the anterior portion for the articular flange (af). These two areas are separated by a low ridge. The ornamentation consists of narrow sinuous ridges with a single row of small, numerous tubercles on their tops, which for the most part are worn. Within these ridges in the center of the ornamented area is the anterior part of the main lateral line groove (lc). The visceral surface is uninformative. In some places, this plate is slightly rough on the margins.
BNTU 6/1-1 (Fig. 4.10) is a very incomplete anterior lateral plate (AL). It is flattened, rather large, with an ornamentation of concentrically orientated ridges topped by a single row of tubercles. The tubercles are quite worn. There is a pair of ridges that join to form a ‘V’ shape. This character is only seen on AL plates. However, because the plate is incomplete, we are unable to determine whether it is a right or left plate or even the original orientation. The visceral surface is obscured by matrix.
BNTU 44/1-6 (Fig. 4.11) is an incomplete unidentified plate. The ornamentation is well preserved consisting of narrow, concentrically orientated ridges topped by a single row of tubercles. Part of an overlap area (oa) and a sensory line (sl) are visible. However, due to the incomplete nature of the plate, we cannot determine what the sensory line was or what plate overlapped this plate. The visceral surface is uninformative.
Etymology
The species name is based on the surname of the Belarusian paleontologist Yury V. Zaika.
Materials
Holotype, a preorbital plate, BNTU 44/1-12; a right paranuchal plate, BNTU 44/1-21; an incomplete left anterior dorsolateral plate, BNTU 44/1-13; a left anterior ventral plate, BNTU 44/1-6; an incomplete right anterior ventral plate, BNTU 6/1-1; and 17 plate fragments, BNTU 44/1-7f, 19/1-2a, 19/1-2b, 44/1-6a, 44/1-6b, 44/1-6c, 44/1-16, 44/1-16a, 116/42-40, 116/42-41, 121/23-1, 121/23-2, 121/23-3, 121/23-4, 121/23-7, 121/23-9, and 121/23-11.
Remarks
Johnston et al. (Reference Johnson, Elliott and Wittke2000) thought that fused left and right preorbital plates were a character of the genus Actinolepis. However, Dupret et al. (Reference Dupret, Zhu and Wang2017) considered it a character of the family Actinolepididae, thereby confirming Actinolepis zaikai as a member of this family. Three genera belong within this family: Actinolepis, Bollandaspis Schultze, Reference Schultze1976, and Szelepis Liu, Reference Liu1981. Two Middle Devonian species of Actinolepis are known: Actinolepis tuberculata Agassiz, Reference Agassiz1844 from the Eifelian and Actinolepis magna Mark-Kurik, Reference Mark-Kurik1973 from the Givetian; both species are found in the Baltic region. A specimen from the Eifelian of Scotland described by Newman and Trewin (Reference Newman and Trewin2008) probably belongs to Actinolepis magna. Another species, Actinolepis spinosa Mark-Kurik, Reference Mark-Kurik1985, is known from the Early Devonian (Pragian) of Latvia. Bollandaspis wosschmidti Schultze, Reference Schultze1976 from the Early Devonian (Emsian) of Belgium is the only species of Bollandaspis. The final species identified to belong to the Actinolepididae is Szelepis yunnanensis (Liu, Reference Liu1979) from the Early Devonian (Pragian) of Yunnan, China.
The most recent cladistic analysis of the Actinolepididae was done by Dupret et al. (Reference Dupret, Zhu and Wang2017). In this analysis they stated that missing data (characters) did somewhat hinder the resolution of relationships among the species. In their analysis, Szelepis yunnanensis was recovered as the sister taxon to Actinolepis tuberculata and Actinolepis magna. One of the reasons for this was the character of the supraorbital sensory lines meeting posteriorly. However, they also stated that none of the specimens was preserved well enough to show whether the pineal and rostral plates were fused, as seen in all Actinolepis species. They also found Actinolepis spinosa as the sister taxon to Szelepis yunnanensis, Actinolepis tuberculata, and Actinolepis magna. However, they suggested this placement might be due to missing data in Actinolepis spinosa. It also must be remembered that Actinolepis spinosa is based on a single specimen recovered from a core and that the length of the armor is only 38 mm long, with the skull roof only 25 mm long. It could well be that the specimen is a juvenile and that some of the characters might be juvenile rather than adult features. However, Mark-Kurik (Reference Mark-Kurik1985) thought this unlikely because there was a second generation of tubercles visible on the specimen. The last member of the Actinolepididae, Bollandaspis wosschmidti, was placed by Dupret et al. (Reference Dupret, Zhu and Wang2017) as sister taxon to the Actinolepis spp. and Szelepis yunnanensis. However, care must also be taken here because B. wosschmidti is based on a single specimen recovered from a core. The armor is only ~17 mm long and therefore the specimen is likely a juvenile. Also, Dupret et al. (Reference Dupret, Zhu and Wang2017) stated that this species had the most missing data of all the actinolepid species. The species has a fused rostral and pineal plate, which could indicate a closer affinity to Actinolepis. In fact, Newman and Trewin (Reference Newman and Trewin2008) suggested that Bollandaspis might be a subjective junior synonym of Actinolepis. Finally, Dupret et al. (Reference Dupret, Zhu and Wang2017) proposed an alternative hypothesis in which Szelepis is more basal than the Actinolepis cluster (which the authors thought could include Bollandaspis).
Two unnamed taxa also have close affinities to Actinolepis. Mark-Kurik (Reference Mark-Kurik1985, Reference Mark-Kurik1991) considered that an undocumented specimen from Severnaya Zemlya probably belongs in Actinolepis. Due to the lack of a description, we cannot consider this form further. Skull roofs of another taxon from the Lower Devonian of Spitsbergen were figured by Ørvig (Reference Ørvig1957, fig. 6), Denison (Reference Denison1958, fig. 105J), and Mark-Kurik (Reference Mark-Kurik1973, fig. 8E). Ørvig (Reference Ørvig1975, fig. 4B) also figured the ventral armor of this taxon. This taxon has both fused left and right preorbital plates and a fused rostral and pineal plate. Denison (Reference Denison1958) and Mark-Kurik (Reference Mark-Kurik1973) thought that this taxon probably belongs in Actinolepis. Later, Denison (Reference Denison and Schultze1978) considered that it differs from Actinolepis by having less elongated preorbitals, a character that it shares with Bollandaspis. However, Mark-Kurik (Reference Mark-Kurik1985) considered it morphologically more similar to Actinolepis than to Bollandaspis. These variations hint at individual generic variation rather than different generic characters. It is somewhat surprising that the Spitsbergen form has not been formally described considering the quality and quantity of material available. Nonetheless, there is enough published material to allow comparison with Actinolepis zaikai n. sp.
Actinolepis zaikai n. sp. is a species based on disarticulated remains with many unknown plates, lacking many anatomical data. Dealing first with the trunk armor, none of the plates of Actinolepis zaikai n. sp. is complete enough to allow morphological comparison with other members of the Actinolepididae. The only real point of comparison is the ornamentation of the plates. Most Actinolepis spp. do not have prominent ridges on the head plates as seen in Actinolepis zaikai n. sp. Actinolepis spinosa is the most similar, in which the tubercles are on top of ridges albeit slightly less pronounced (Mark-Kurik, Reference Mark-Kurik1985, figs. 1, 4b). The nature of the trunk plate ornamentation in the closely related form Bollandaspis wosschmidti is more difficult to compare with Actinolepis zaikai n. sp. due to the small size of the only known specimen. Certainly, there are hints of ridges and tubercles (Schmidt, Reference Schultze1976, figs. 1, 2, pl. 3). However, it is the head plates that show the most differences in Actinolepis zaikai n. sp. compared with other members of the Actinolepididae, as discussed below.
The more complete plates in Actinolepis zaikai n. sp. are the fused preorbital plates (PrO + PrO) and the paranuchal plate (PNu). The PNu plate is very similar in shape to the PNu plate of Actinolepis tuberculata (see Mark-Kurik, Reference Mark-Kurik1973, fig. 1C, pl. 8, fig. 4) and Actinolepis magna (see Mark-Kurik, Reference Mark-Kurik1973, fig. 6, pl. 10, fig. 1). The arrangement of the sensory lines is also very similar. The PNu plates are not well preserved in Actinolepis spinosa and thus no comparison can be made. Like the trunk plates, the ornamentation of the PNu plates in Actinolepis zaikai n. sp. has a pattern of ridges with tubercles that follow the outlines of the plate. This differs from those of Actinolepis tuberculata and Actinolepis magna, which show no ridges. The PNu plates are unknown in Bollandaspis wosschmidti. The PNu is not very well preserved in Szelepis yunnanensis, making comparison with Actinolepis zaikai n. sp. difficult. One thing of note is that the ornamentation in Szelepis yunnanensis consists of widely spaced tubercles, which is quite different from Actinolepis zaikai n. sp.
The most distinctive plate of Actinolepis zaikai n. sp. is the fused preorbital plates (PrO + PrO). Like Actinolepis tuberculata (see Mark-Kurik, Reference Mark-Kurik1973, fig. 1A, B, pl. 8, figs. 1, 3) and Actinolepis magna (see Mark-Kurik, Reference Mark-Kurik1973, figs. 6, 8A, B, pl. 9, fig. 4, pl. 10, fig. 4), the left and right supraorbital lines (soc) meet toward the posterior of the plate and there is a deep embayment at the anterior end where the pineal (most likely fused with the rostral plate) attaches. Actinolepis zaikai n. sp. differs from Actinolepis tuberculata and Actinolepis magna in having a more pronounced long process (with a rounded end) at the posterior end that fits between the central plates. Actinolepis zaikai n. sp. differs from Actinolepis tuberculata (see Mark-Kurik, Reference Mark-Kurik1973, fig. 2C) in having much less robust ridges (r) on the visceral side under the soc. Actinolepis magna (see Mark-Kurik, Reference Mark-Kurik1973, fig. 7A, B) also has slightly more robust ridges than Actinolepis zaikai n. sp., but differs from it by having a prominently exposed fenestra hypophyseos. Also, the ornamentation of the plate, like all of the other plates, consists of pronounced ridges topped by tubercles that follow the outlines of the plates. These ridges are missing from both Actinolepis tuberculata and Actinolepis magna. The PrO + PrO of Actinolepis spinosa is similar in shape to the plate in Actinolepis tuberculata and Actinolepis magna, although it differs from them in that the soc do not meet. This is an obvious difference to Actinolepis zaikai n. sp. Also, the ornamentation is decidedly different, in that there is no indication of any ridges on any of the skull roof plates in Actinolepis spinosa. The PrO + PrO of Bollandaspis wosschmidti is much shorter and wider than any of the Actinolepis spp. including Actinolepis zaikai n. sp. (Schmidt, Reference Schultze1976, p. 11, figs. A, B). Also, the soc does not meet, marking a difference from Actinolepis zaikai n. sp.
The dorsal surface of the PrO + PrO of Szelepis yunnanensis is not well preserved (Dupret et al., Reference Dupret, Zhu and Wang2017, fig. 3C, D). Like on the paranuchal plate, the ornamentation consists of widely spaced tubercles, quite different from Actinolepis zaikai n. sp. A better-preserved specimen in visceral view (Dupret et al., Reference Dupret, Zhu and Wang2017, fig. 3A, B) shows that there is no deep embayment at the anterior end to receive the pineal plate. The PrO + PrO is also much shorter and wider. These two characters make it distinctive from the Actinolepis spp., including Actinolepis zaikai n. sp.
Actinolepis zaikai n. sp. differs from other members of the Actinolepididae by the nature of the ornamentation of the trunk and head plates and the long fused preorbital plates with a distinct, central posterior process.
Suborder Coccosteina Heintz, Reference Heintz and Gudger1932
Superfamily Coccosteoidea Denison, Reference Denison and Schultze1978
Family Coccosteidae Traquair, Reference Traquair1888
Coccosteidae genus indet. species indet.
Figure 5
- Reference Plax, Gubin, Zui, Lukashev and Sanko2015a
Coccosteidae gen. indet. sp. indet.; Plax, p. 171, pl. 1, figs. 8–9.
- Reference Plax2016
Coccosteidae gen. indet. sp. indet.; Plax, p. 16, table, pl. 1, figs. 4, 7–8.

Figure 5. Coccosteidae gen. indet. sp. indet., dermal plate remains: (1) right central plate (BNTU 51/2-2c); (2, 3) posterior part of a median dorsal plate (BNTU 51/2-2b); (2) dorsal view; (3) visceral view; (4, 5) left anterior dorsolateral plate (BNTU 51/2-2): (4) lateral view; (5) visceral view; (6, 7) right anterior dorsolateral plate (BNTU 51/2-2a): (6) lateral view; (7) visceral view; (8, 9) left posterior ventrolateral plate (BNTU 51/2-2e): (8) ventral view; (9) visceral view; (10, 11) probable left posterior lateral plate (BNTU 51/2-2g): (10) lateral view; (11) visceral view; (12, 13) spinal plate (BNTU 51/2-2f): (12) outer view; (13) visceral view; (14, 15) indefinable plate fragment (BNTU 51/2-2d): (14) external view; (15) visceral view. af.smd.pl = articular face for submedian dorsal plate; cf.PDL = area overlapping posterior dorsolateral plate; cf.PVL = area overlapping posterior ventrolateral plate; cr.pr = carinal process of median dorsal plate; csc = central sensory canal; k = keel; kd = glenoid condyle; lc = main lateral line canal; ld = dorsal branch of main lateral sensory line groove; mp = middle pit-line groove; oa.AL = area overlapped by anterior lateral plate; oa.AVL = overlap area for anterior ventrolateral plate; oa.MD = area overlapped by median dorsal plate; os = osteolepid scale; pr.p = cartilaginous prepectoral process of endoskeletal scapular belt; pr.sbgl = subglenoid condyle; soc = supraorbital sensory line. Scale bars = 5 mm (1–11, 14, 15), 1 mm (12, 13). Dashed lines indicate the estimated boundary of the missing parts of the plate.
Occurrence
From the Buda Dal'nyaya 35 borehole at a depth of 233.2 m; also from the Vitebsk region in the Lepel Beds of the Vitebsk Regional Stage, upper Emsian, Lower Devonian.
Description
Seven identifiable plates were found, one from the cranial roof and six from the trunk armor, as well as two unidentifiable plate fragments:
BNTU 51/2-2c (Fig. 5.1) is an incomplete right central plate (C) that is slightly convex. It is ~10 mm long and 8 mm wide. The supraorbital sensory line (soc), the central sensory canal (csc), and the middle pit-line groove (mp) are well developed on the dorsal side of the plate. The csc and mp branch off from one another on the left side of the plate and then extend parallel to one another to the right edge of the preserved plate. The soc curves anteriorly from the csc toward the right edge of the plate. The angle between the csc and soc at the right lateral edge is 30°. The tubercles are rounded and of approximately the same size. They are evenly distributed across the plate surface. These form weakly expressed rows near the sensory grooves.
BNTU 51/2-2b (Fig. 5.2, 5.3) is a fragment of the posterior part of a median dorsal plate (MD). The specimen is 21 mm long and 7 mm wide at its widest point. With extrapolation, it can be argued that the plate is relatively long and slightly concave in profile. The posterior part of the plate tapers to a point, although the tip is missing. The dorsal surface (Fig. 5.2) is covered with small round tubercles, which form rows parallel to the lateral edges. Toward the center of the plate, the tubercles are more random and much scarcer. The right side of the dorsal branch of the main lateral sensory line groove (ld) is present as a gentle curve. Although the left side of the plate is missing, the ld do not connect to the midline. The ld would have intersected the right and left posterior dorsolateral plates. The visceral surface (Fig. 5.3) is smooth, except for a slight depression for the area overlapping the posterior ventrolateral plate (cf.PVL). The median ventral keel (k) is sharp and well developed. It begins near the anterior margin of the missing part of the plate, and continues to the posterior, gradually deepening. It ends posteriorly with a carinal process (cr.pr). The posterior face of the carinal process flares out and is hollowed to make an articular surface for the submedian dorsal plate (af.smd.pl).
Two anterior dorsolateral plates (ADL) have been identified. BNTU 51/2-2 is a left ADL (Fig. 5.4, 5.5) and BNTU 51/2-2a a right ADL (Fig. 5.6, 5.7). BNTU 51/2-2 is the better-preserved specimen. It is convex on its lateral surface and is quadrangular overall. The plate is 10 mm long and 14 mm wide. The glenoid condyle and the subglenoid process are not preserved. Also, the most anterior ornamented part of the plate is not preserved. The lateral surface (Fig. 5.4) is covered by numerous, well developed, small, round tubercles arranged in concentric rows around a center of ossification. The areas overlapped by the median dorsal plate (oa.MD) and the left anterior lateral plate (oa.AL) are partially preserved. The dorsal branch of the main lateral line canal (ld) and main lateral line canal (lc) are clearly seen on the lateral surface. The lc originates where the glenoid condyle would be and extends posteriorly in a downward diagonal direction to where the posterior lateral plate would have been. The ld extends posteriorly from near the anterior end of the lc diagonally upward toward where the posterior dorsolateral plate would have been. The angle between the lc and ld is ~60°. The visceral surface (Fig. 5.5) is smooth. The area overlapping the posterior dorsolateral plate (cf.PDL) is visible on the visceral surface on the posterior margin.
BNTU 51/2-2a is a right anterior dorsolateral plate (ADL) with broken lateral edges. The plate is 12 mm long and 14 mm wide. Its lateral surface (Fig. 5.6) is somewhat obscured by matrix although the lateral lines are visible, particularly the dorsal branch of the main lateral line canal (ld). The plate is morphologically similar to BNTU 51/2-2 but has the glenoid condyle (kd) and the subglenoid process (pr.sbgl) preserved. These two structures are morphological similar to those of the better-known ones in Coccosteus cuspidatus as described by Miles and Westoll (Reference Miles and Westoll1968). The visceral surface (Fig. 5.7) is smooth and shows no diagnostic characters other than the area overlapping the posterior dorsolateral plate (cf.PDL).
BNTU 51/2-2e (Fig. 5.8, 5.9) is a left posterior ventrolateral plate (PVL) lacking the anterior lateral corner. It is elongated and ventrally convex in profile. The plate is 21 mm long and 11 mm wide at the widest point. At the anterior edge, part of the overlap area for the anterior ventrolateral plate (oa.AVL) is visible. The ventral surface (Fig. 5.8) is covered with small, rounded tubercles that are slightly larger and more prominent along the edges of the plate. An osteolepid scale (os) and matrix obscures part of the ventral surface of the plate, which is too delicate to allow removal. The visceral surface (Fig. 5.9) is mostly smooth. The area overlapping the right posterior ventrolateral plate (cf.PVL) is clearly seen on the mesial margin.
BNTU 51/2-2 g (Fig. 5.10, 5.11) is probably a left posterior lateral plate (PL) but it could be a posterior dorsolateral plate (PDL) because they are morphologically very similar. The plate is vaulted, triangular, and nearly complete. It is length is 8 mm long and 9 mm wide on the longest axis. Almost the entire lateral surface (Fig. 5.10) is covered with matrix, but there are small areas where several small, round tubercles can be observed. Overlap areas for other plates are visible on some of the edges but it is not certain which plates attached to these areas. The visceral surface (Fig. 5.11) is smooth and featureless.
BNTU 51/2-2f (Fig. 4.12, 4.13) is a relatively short spinal plate (Sp). It is 7 mm long and ~3.5 mm wide. Its external surface (Fig. 5.12) has small, rounded tubercles arranged in rows parallel to the long axis. Viscerally (Fig. 5.13), there is a conical cavity opening anteriorly that was filled with the cartilaginous prepectoral process of the endoskeletal scapular belt (pr.p).
BNTU 51/2-2d (Fig. 5.14, 5.15) is a small unidentifiable plate fragment. It is triangular is shape with a maximum length of 11 mm. The plate has a similar ornamentation on the external surface as the plates described above, showing that even in the smallest specimen, there is reasonable confidence in identifying it to species.
Materials
A right central plate, BNTU 51/2-2c; a fragment of the median dorsal plate, BNTU 51/2-2b; a right anterior dorsolateral plate, BNTU 51/2-2; a left anterior dorsolateral plate, BNTU 51/2-2а; a posterior lateral plate, BNTU 51/2-2g; a left posterior ventrolateral plate, BNTU 51/2-2e; a spinal plate, BNTU 51/2-2f; and unidentifiable plate fragments, BNTU 51/2-2d and 51/2-2h.
Remarks
We are confident that we are dealing with one coccosteid species because all specimens have a very similar ornamentation. However, it is not possible to assign the remains to a genus and species due to the uniformity of known plates when compared with other coccosteid genera such as Coccosteus, Dickosteus Miles and Westoll, Reference Miles and Westoll1963 (see Miles and Westoll, Reference Miles and Westoll1968), and Millerosteus (see Desmond, Reference Desmond1974).
Occurrence
From the Bykhov 1 borehole at depths of 325.5 m, 324.3 m, 324.2 m, 321.3 m, 321.2 m, and 321.0 m, Mogilev region; also from the Korma 1 borehole at depths of 343.5 m and 323.3 m, Gomel region. Both borehole specimens are from the Lepel Beds of the Vitebsk Regional Stage, which is in the upper Emsian of the Lower Devonian.

Figure 6. Placodermi indet. 1, dermal plate fragments: (1, 2) fragment of a spinal plate (BNTU 116/49-8): (1) external view; (2) visceral view; (3) fragment of spinal plate (BNTU 116/54-1); (4) fragment of a plate (BNTU 116/49-7); (5, 6) fragment of a plate (BNTU 116/53-4): (5) external view; (6) visceral view. Scale bars = 1 mm.
Description
From the localities described above, a large number of small (3–20 mm), thin, unidentifiable plate fragments (Figs. 6, 7). They have a characteristic ornamentation of variably sized, rounded tubercles, which are star-shaped at their bases. Often these tubercles form distinct rows; also larger tubercles are often present between smaller ones.
Several small fragments of spinal plates have also been collected (Figs. 6.1–6.3, 7.2–7.6). They are oval in cross section. The tubercles on these plates often fuse with each other to form distinct rows (e.g., Fig. 7.2). Along the mesial margin of the spinal plates are rows of large, well-developed, strongly curved, evenly spaced spinelets (Fig. 7.3–7.5).

Figure 7. Placodermi indet. 1, dermal plate fragments: (1) plate fragment (BNTU 116/49-1); (2–6) spinal plate fragments: (2) BNTU 121/48-1; (3) BNTU 116/49-2; (4) BNTU 116/48-3; (5) BNTU 116/48-5; (6) BNTU 116/56-1. spl.b = spinelet base. Scale bars = 1 mm (1, 3); 0.5 mm (2, 4); 0.2 mm (6); 0.1 mm (5).
Materials
Spinal plate fragments, BNTU 116/48-2, 116/48-3, 116/49-2, 116/49-3, 116/54-1, and 121/27-3; a denticle from a spinal plate, BNTU 116/48-5; small unidentifiable plate fragments, BNTU 116/48-1, 116/48-4, 116/48-6, 116/49-1, 116/49-7, 116/49-8, 116/50-2, 116/53-2, 116/53-6, 116/53-8, 116/56-1, 116/56-2, 116/56-3, 116/56-4, 116/56-5, and 121/48-1.
Remarks
Due to the incomplete nature of the specimens, it is not possible to assign them to a genus and species. The spinal plates are reminiscent of an actinolepid, phlyctaenine, or groenlandaspidid, but there are other possibilities, e.g., a petalichyiid, which makes assignment even to arthrodires impossible. They are different from the three other forms described in having spinelets on the spinal plates.
A brief taphonomic description of the upper Emsian placoderm remains
The Lepel Beds placoderms (quite rare) described in this article are found in the terrigenous and carbonate-terrigenous rocks. The lithology consists mostly of mudstones with occasional siltstones and sandstones. Associated with the placoderms are ostracode valves (less common), conchostracan shells (very rare), and near-complete valves of inarticulate brachiopods (less common). Vertebrate remains include dentine tubercles, tesserae, and small plate fragments of psammosteid heterostracans (quite rare); scales and fin spine fragments of acanthodians (scales very common, fin spines quite rare); scales of chondrichthyans (very rare); scales, teeth, jaws, and unidentifiable bone fragments of sarcopterygians (less common); scales of actinopterygians (very rare); and fish otoliths (very rare). Acritarchs (less common) and miospores (very common) are also present.
Most of the organic remains are found in massive or poorly laminated rock. The placoderm remains are usually isolated, but occasionally form small accumulations with other organic remains. The organic remains usually lie flat on the bedding planes. The placoderm remains are usually well preserved, but often fragmentary with only occasionally whole plates preserved. These remains range in size from a few to 40 mm. Although the plates are quite dense, they are often also brittle. The histological structure of the placoderm remains are well preserved. The placoderm plates are not deformed and are yellowish orange to dark brown (sometimes black). The plates have sharp edges with little rounding, indicating that they were deposited in a quiet environment. However, because the remains are fragmentary, they must have been transported for some distance. The presence of acritarchs indicates a marine influence on the deposit, but the lack of other true marine forms suggests that it was marginal. Conchostracans and inarticulate brachiopods often preferred brackish-water conditions during the Devonian (see e.g., Knox and Gordon, Reference Knox and Gordon1999) and thus the deposit could represent a shallow coastal basin with an input of freshwater, perhaps even an estuary or lagoon. However, it is difficult to be certain of the precise environment because there are no surface outcrops of the Lepel Beds.
Conclusions
Despite disarticulated material, it is clear that at least four placoderm species are present in the upper Emsian rocks of Belarus. There is sufficient material to erect two new taxa: Stipatosteus svidunovitchi n. gen. n. sp. and Actinolepis zaikai n. sp. The other two species were not well-enough preserved to allow species assignment and had to be left in open nomenclature: Coccosteidae gen. indet. sp. indet. and Placodermi indet. 1.
The presence of a species of Actinolepis indicates at least some connection during the Devonian between Belarus and the Baltic region where the genus also occurs. Actinolepis also occurs in Scotland and Spitsbergen, indicating a possible connection during the Devonian. Whether this connection was during the upper Emsian or earlier is impossible to say at present and further research by the authors and others is presently under way to ascertain this. Finally, Burrow et al. (Reference Burrow, den Blaauwen, Newman and Davidson2016) noted that the acanthodian Diplacanthus kleesmentae was very similar, if not a subjective junior synonym of the Scottish Middle Devonian species Diplacanthus crassisimus (Duff, Reference Duff1842), but that the Baltic material was not well-enough preserved to be certain. Further research is being undertaken by Burrow and her team on the Scottish acanthodians to establish the species diversity recorded within the Old Red Sandstone. The Belarus material is particularly important to the latter investigation because some of the Devonian strata includes a fully marine fauna, thus allowing a global correlation.
Based on lithological, palaeontological, and taphonomic data, it is thought that the placoderms described in this paper, with their associated vertebrates and invertebrates, lived in a shallow epicontinental sea basin, probably with freshwater input. These new data allow us to supplement the biostratigraphy of the subregional stratigraphic units of the most recently published stratigraphic chart of the Devonian deposits of Belarus (Obukhovskaya et al., Reference Obukhovskaya, Kruchek, Pushkin, Nekryata, Plax, Sachenko, Obukhovskaya and Antipenko2010).
Acknowledgments
The authors express deep gratitude to D.A. Stepanenko for the help rendered in photographing some studied placoderm skeletal elements. We would also like to thank R. Carr and two anonymous reviewers whose reviews greatly improved the quality of this paper.








