Introduction
The marine upper Permian strata occur widely not only in South China but also in Tibet, Southwest China. Diener (Reference Diener1897) first reported Chinese Lopingian rugose corals from southern Tibet. Grabau (Reference Grabau1922, Reference Grabau1928) and Huang (Reference Huang1932) reported and simply described the Lopingian corals in stratigraphical and paleontological investigation papers. Tseng (Reference Tseng1949) published the first taxonomic paper on the Lopingian corals. Subsequently, Wu (Reference Wu1957) and Tseng (Reference Tseng1959) provided important taxonomic information on fasciculate rugose faunas. During the past three decades, important data on Lopingian rugose corals has been widely scattered in Chinese research journals and books such as paleontological atlases issued by the local geological surveys (Zhao, Reference Zhao1976, Reference Zhao1981, Reference Zhao1984; Wu et al., Reference Wu, Liao and Zhao1982; Wu and Zhao, Reference Wu and Zhao1983; Xu, Reference Xu1984a, b; Ding et al., Reference Ding, Jin and Sun1989; Fang and Fan, Reference Fang and Fan1994; Xu and Sando, Reference Xu, Sando, Xu, Luo, Wang, Zhou and Xiao1997; Wang and Wang, Reference Wang and Wang2007).
Although Diener reported Lopingian rugose corals from Tibet as early as 1897, it was not until the past three decades that some coral faunas were described in detail from the late Permian of Tibet (Zhao, Reference Zhao1984; He, Reference He, Yang and Nie1990; Cheng et al., Reference Cheng, Wu and Zhang2002; Fan et al., Reference Fan, Yu, He, Pan and Li2003). The Gyanyima section contains abundant rugose coral faunas. Owing to the very remote location, the rugose faunas in the Gyanyima section have not been studied systematically until now. Only four species of the genus Waagenophyllum Hayasaka, Reference Hayasaka1924 were reported in previous works (Wang and Xu, Reference Wang and Xu1988; He, Reference He, Yang and Nie1990), but the materials were not adequately thin-sectioned, and the descriptions are far too simple.
The majority of specimens described in this paper were collected by Cao Changqun and Li Wenzhong in 2002, and some specimens were collected by Li Wenzhong in 2006 from the Gyanyima section in southwestern Tibet. We propose a new genus and species, Gyanyimaphyllum crassiseptatum n. gen. n. sp., describe seven new species, including Waagenophyllum (Liangshanophyllum) clisicolumellum n. sp., W. (Waagenophyllum) intermedium n. sp., W. (W.) gyanyimaense n. sp., Ipciphyllum naoticum n. sp., I. floricolumellum n. sp., and I. zandaense n. sp., and provide more complete descriptions and discussions of some previously named species, including W. (W.) ngariense He, Reference He, Yang and Nie1990 and W. (W.) elegantulum He in Luo et al., Reference Luo, He, Wang, Wang, He and Chen1989.
Geological setting and biostratigraphy
The Gyanyima section is located in Burang County in the Ngari region of southwestern Tibet. It is about 30 km north of the Chinese-Indian border and about 50 km northwest of Burang County Town (Fig. 1). The starting point of the section was measured as 30°43′13.5″N, 80°41′46.4″E with an altitude of 4,534 m above sea level (within the Great Himalaya Range), and the Permian-Triassic boundary of the section is at 30°42′38.4″N, 80°41′42.4″E.

Figure 1. Sketch map showing the location of the Gyanyima section in southern Tibet (base map modified after Zhang et al., Reference Zhang, Wang and Shen2009).
In the Gyanyima Section, the Permian Gyanyima Formation is overlain by the Triassic Lanchengqu Formation. The total thickness of the measured section is 269.18 m (Fig. 2). The Gyanyima Formation is subdivided into two parts: the lower part is 97.7 m in thickness and comprises beds 1–5, including basalt at the top (bed 5). This part mainly consists of reddish, thick-bedded to massive limestone in the lower part and gray, medium-bedded limestone in the upper part containing volcanic fragments. The upper part includes beds 6–10 and is gray and reddish gray, medium- to thick-bedded bioclastic limestone and partly contains thin-bedded limestone intercalations with several siliceous bands.
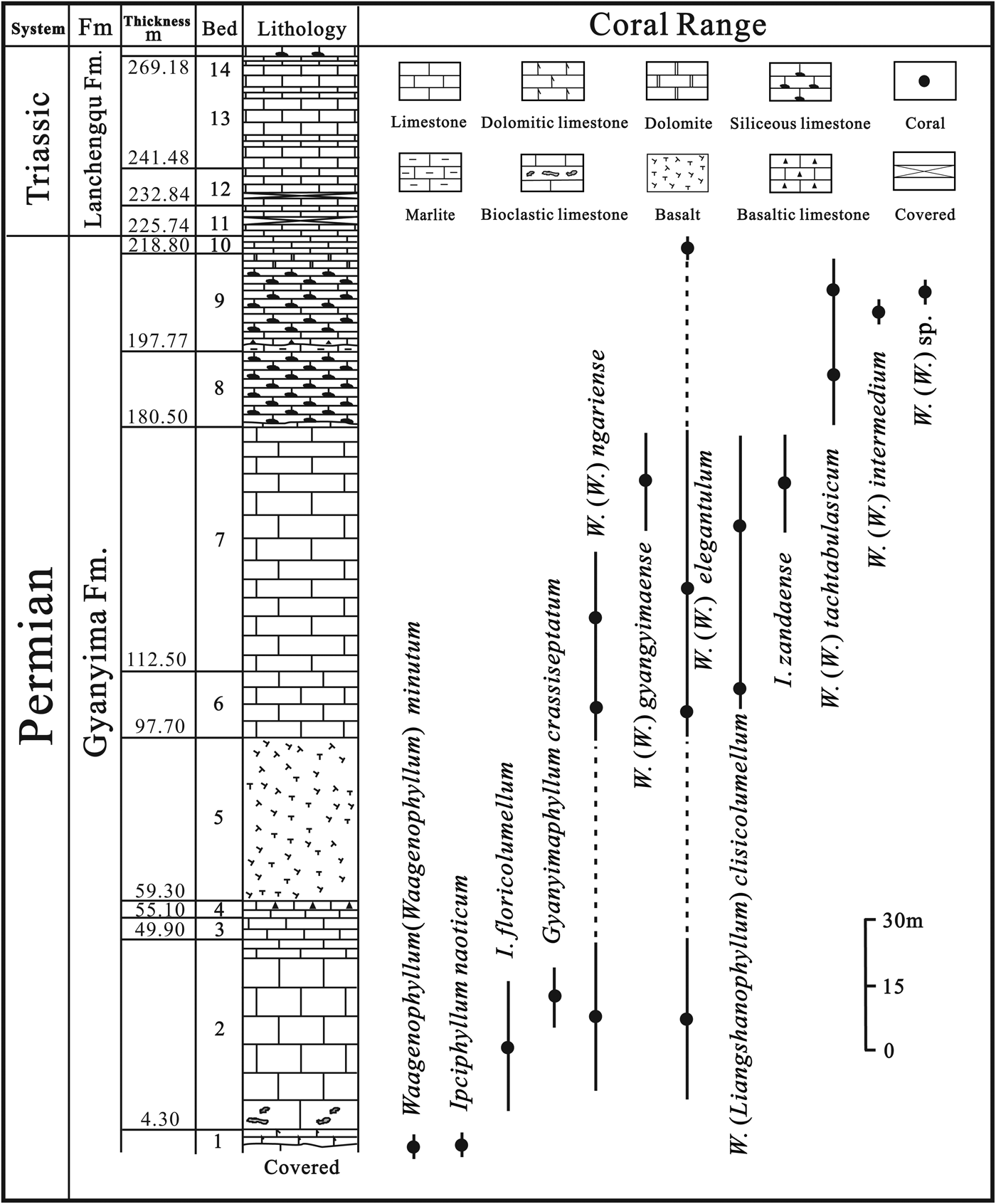
Figure 2. Stratigraphic distribution of corals in the Gyanyima section.
Corals were mainly collected from beds 2, 6, 7, and 9 (Fig. 2). Well-preserved rugose corals are accompanied by a highly diversified foraminiferal fauna that includes Reichelina cribroseptata Erk, Reference Erk1941, Reichelina changhsingensis Sheng and Zhang, Reference Sheng and Zhang1958, Colaniella fusiformis Song, Reference Song, Yang and Nie1990, Colaniella parva (Colani, Reference Colani1924), and others. These foraminiferids indicate a possible Changhsingian (late Permian) age (Wang et al., Reference Wang, Ueno, Zhang and Cao2010). A coral reef, with Waagenophyllum as the major skeletal reef builder, occurs in the uppermost part of the sequence (bed 10). This is possibly one of the latest Permian rugose coral reefs in the world (Wang and Wang, Reference Wang and Wang2007). To the best of our knowledge, Gyanyimaphyllum crassiseptatum n. gen. n. sp. and Ipciphyllum naoticum n. sp. are the latest representatives, respectively, of fasciculate and massive rugose corals with naotic structure.
Paleobiogeographical implications
Paleogeographically, the section represents an isolated carbonate build-up or a seamount deposited in the Neotethys between the Himalayan Tethys Zone and the Lhasa Block (Shen et al., Reference Shen, Cao, Li and Shi2010; Wang et al., Reference Wang, Ueno, Zhang and Cao2010; Fig. 3). The Gyanyima Formation occurs as an exotic limestone block within the Yarlung-Tsangpo Suture Zone (Shen et al., Reference Shen, Cao, Li and Shi2010; Zhang et al., Reference Zhang, Shi and Shen2013). Elsewhere in this suture zone, these limestone blocks were widely reported (e.g., Wang and Mu, Reference Wang and Mu1980; Robertson, Reference Robertson1998; Shen et al., Reference Shen, Cao, Shi, Wang and Mei2003a, b). These limestone blocks were mostly preserved in Late Triassic or Jurassic–Cretaceous mélanges along the ophiolite belt (Shen et al., Reference Shen, Sun and Shi2003c; Zhu et al., Reference Zhu, Du, Liu, Feng and Tian2005; Cai et al., Reference Cai, Ding, Leary, Wang and Xu2012). Their formation was considered to be closely related to the subduction of the Neotethys plate beneath the Lhasa Block or an intraoceanic arc (Ali and Aitchison, Reference Ali and Aitchison2008; Cai et al., Reference Cai, Ding, Leary, Wang and Xu2012). It is also inferred to be the sequence of the accretionary complex that the Permian exotic limestone blocks were transferred northward from the Gondwana margin (Cai et al., Reference Cai, Ding, Leary, Wang and Xu2012). Therefore, these exotic limestone blocks should locate at the northern margin of Gondwana before their northward drift together with the Tethys Himalaya area during the Early Cretaceous (van Hinsbergen et al., Reference Van Hinsbergen, Lippert, Dupont-Nivet, McQuarrie and Doubrovine2012). In addition, the absence of Palaeofusulina Deprat, Reference Deprat1912 representing warm-water fusulinids and the presence of cool-water brachiopod elements may indicate that the Gyanyima area was not far from the Gondwana margin and easily affected by cool-water currents or cooling events (Shen et al., Reference Shen, Cao, Li and Shi2010).
The Changhsingian corals were also sporadically reported from other regions in the Tethys Himalaya area. In the Drongba area, the coral Tachylasma sp. was discovered in the Gaqoi limestone block (Wen et al., Reference Wen, Zhang, Wang, Sun and Dong1984). It is associated with Changhsingian foraminiferal genera Reichelina Erk, Reference Erk1941, Paracolaniella Wang, Reference Wang1966, and Colaniella Licharev, Reference Licharev1939. In the Nyalam area, the Lopingian Selong Group is dominated by cold-water brachiopod fauna (Shen et al., Reference Shen, Archbold, Shi and Chen2000, Reference Shen, Archbold, Shi and Chen2001). The associating coral fauna are all small solitary types such as Lytvolasma Soshkina, Reference Soshkina1925, Wannerophyllum Schouppe and Stacul, Reference Schouppe and Stacul1955, Tachylasma Grabau, Reference Grabau1922, and Lophyphyllidium Grabau, Reference Grabau1928 (Wu, Reference Wu1975). The absence of compound corals in the Gaqoi limestone block and the Selong Group is pronounced compared to their abundance in the Gyanyima limestone block. It is suggested here that the Gyanyima coral assemblage may represent the southernmost occurrence of compound corals and coral reefs during the Changhsingian. More significantly, the presence of Lopingian compound corals in the Gyanyima limestone blocks indicates that the northern Tethys Himalaya margin was warm enough to allow the coral reefs to dwell there. It is noteworthy that the climate in the Gyanyima area may have improved since the late Guadalupian because the underlying Xilanta Formation yields abundant compound corals of the Iranophyllum-Ipciphyllum assemblage (Wang and Xu, Reference Wang and Xu1988) and diverse fusulinid fauna (Zhang et al., Reference Zhang, Wang and Shen2009). The presence of coral reefs in this region further suggests that a clear climatic boundary may have existed in the northern Gondwana margin during the late Guadalupian and Lopingian epochs. The faunas in the north of this boundary are marked by the presence of diverse compound corals, warm-water brachiopods, and fusulinids, whereas the areas south of this boundary are dominated by cold-water brachiopods and small solitary corals (Fig. 3).
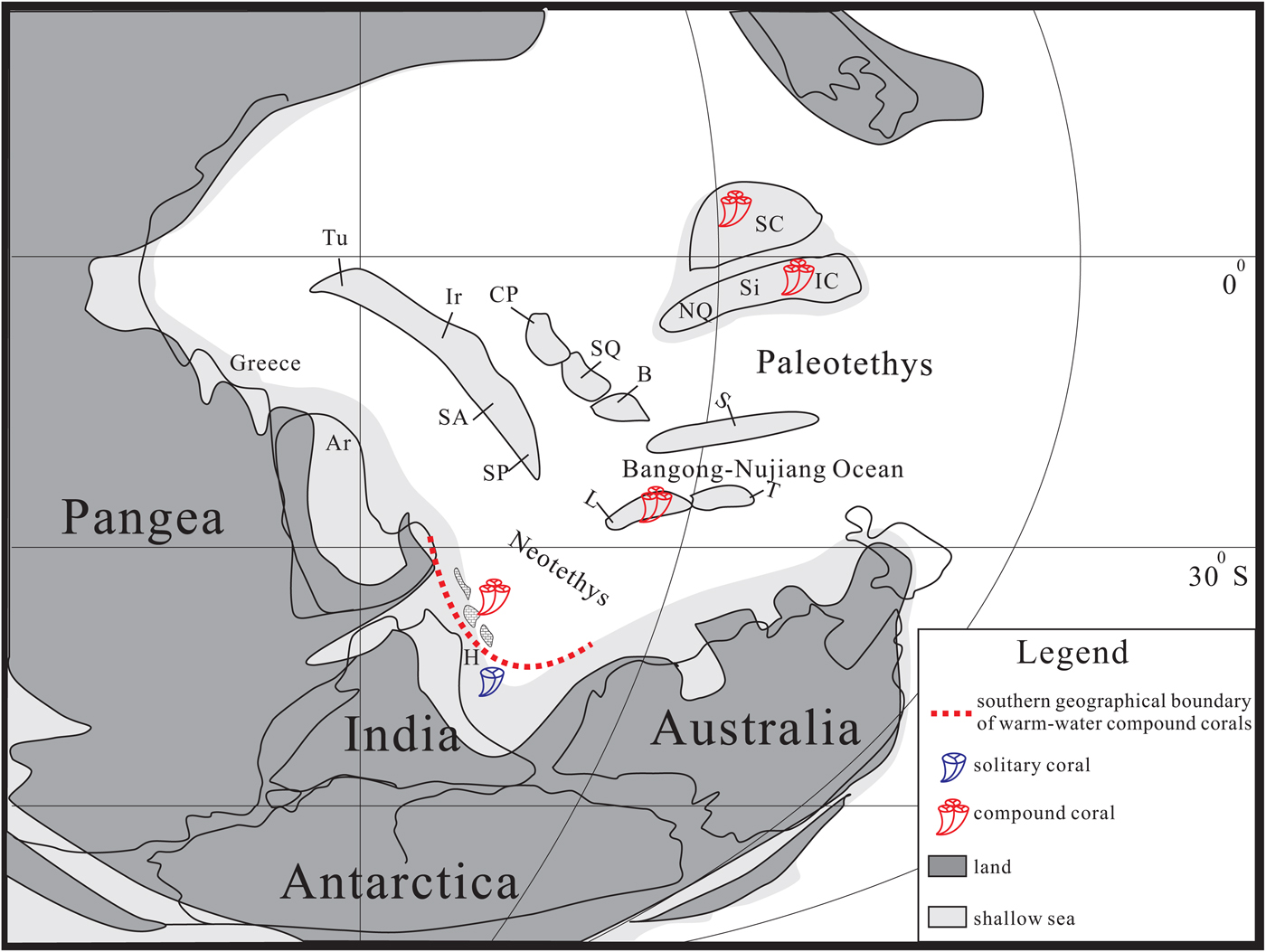
Figure 3. Schematic paleogeographic map of the Tethyan area during the Changhsingian showing the distribution of compound and solitary corals. Base map modified after Zhang et al. (Reference Zhang, Shen, Zhang, Zhu and An2019). Ar, = Arabia; B = Baoshan Block; CP = central Pamir; H = Himalaya; IC = Indochina Block; Ir = central Iran (including Armenia); L = Lhasa Block; NQ = North Qiangtang Block; S = Sibumasu Block; SC = South China; SA = South Afghanistan; SQ = South Qiangtang Block; Si = Simao Block; SP = south Pamir (including Karakorum); T = Tengchong Block; Tu = Turkey.
Materials and methods
A total of 41 specimens were collected mainly from beds 2, 6, 7, and 9 in the Gyanyima section in southwestern Tibet. From these, 298 thin sections were made for observation, among which 61 thin sections from 23 specimens were photographed and illustrated in this paper. Drawing on these thin sections and specimens, 12 compound rugose species belonging to four genera or subgenera are described and discussed. The morphologic terminology used here follows that of Hill (Reference Hill and Teichert1981).
The diameter of corallites was measured by Image-Pro Plus 5.0 under a Nikon E800 microscope. The mean diameter of corallites was determined in transverse section by taking the average of the measured corallite diameters. The greatest and least diameters of Waagenophyllum and Gyanyimaphyllum n. gen. were measured. For measurement of the diameter of Ipciphyllum Hudson, Reference Hudson1958, we take the average of the distance between the midpoint of each wall and opposite side through the central point of the axial structure. The ratio of septal number to corallite diameter (n/d) has been calculated because of its importance in species recognition (Fedorowski, Reference Fedorowski, Jell and Pickett1989).
Repositories and institutional abbreviations
All figured specimens are registered with the prefix NIGP and housed in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Science, Nanjing, China. Other holotypes mentioned in this paper were registered and housed as follows: prefix SEM, the museum of Fuzhou University, Fuzhou, China; prefix TF, Tibetan fossil collection of China University of Geosciences (Beijing), Beijing, China; postfix PIN, Palaeontological Institute, Russian Academy of Science, Moscow, Russia.
Systematic paleontology
Subclass Rugosa Milne-Edwards and Haime, Reference Milne-Edwards and Haime1850
Order Stauriida Verrill, Reference Verrill1865
Suborder Lonsdaleiina Spassky, Reference Spassky1974
Family Waagenophyllidae Wang, Reference Wang1950
Subfamily Waagenophyllinae Wang, Reference Wang1950
Genus Waagenophyllum Hayasaka, Reference Hayasaka1924
Subgenus Waagenophyllum Hayasaka, Reference Hayasaka1924
Type species
Lonsdaleia indica Waagen and Wentzel, Reference Waagen and Wentzel1886, p. 897, pl. 101, figs. 1–3. Middle Productus limestone, Salt Range, Pakistan.
Diagnosis
See Minato and Kato, Reference Minato and Kato1965, p. 97.
Remarks
The taxonomic relationship between Waagenophyllum Hayasaka, Reference Hayasaka1924 and Liangshanophyllum Tseng, Reference Tseng1949 has being controversial since Liangshanophyllum was first described and established to be a subgenus of Waagenophyllym by Tseng, in 1949. Tseng mentioned that the axial structure of Liangshanophyllum is much simpler than that of Waagenophyllum, which is the major difference between the two genera. Yu et al. (Reference Yu, Lin and Fan1962) revised Liangshanophyllum as a genus and emphasized the difference in the development of tabulae between Waagenophyllum and Liangshanophyllum. The former has very steep clinotabulae, whereas the latter has commonly horizontal tabulae and a relatively wider tabularium (Yu et al., Reference Yu, Lin and Fan1962). Most Chinese researchers have accepted this taxonomic recognition (Wu and Wang, Reference Wu and Wang1974; Wang, Reference Wang1978; Li and Liao, Reference Li and Liao1979; Jiang, Reference Jiang1982; Cao et al., Reference Cao, Ouyang, Jin and Cai1983; He and Weng, Reference He and Weng1983; Ding, Reference Ding1995; Xu and Sando, Reference Xu, Sando, Xu, Luo, Wang, Zhou and Xiao1997). However, the majority of authors still treat Liangshanophyllum as a subgenus of Waagenophyllum (Yu et al., Reference Yu, Wu, Zhao and Zhang1963; Minato and Kato, Reference Minato and Kato1965; Yamagiwa and Hattori, Reference Yamagiwa and Hattori1970; Chen and Xiong, Reference Chen and Xiong1978; Flügel, Reference Flügel1972; Hill, Reference Hill and Teichert1981; Stevens et al., Reference Stevens, Miller and Nestell1987; Fontaine and Tien, Reference Fontaine and Tien1988; Ezaki, Reference Ezaki1991; Shen et al., Reference Shen, Kawamura and Yang1998). A few authors have pointed out that the shape and arrangement of tabulae are variable in different specimens of the same species, even in different corallites in the same colony (Xu in Jia et al., Reference Jia, Xu, Kuang, Zhang and Zuo1977; Shen et al., Reference Shen, Kawamura and Yang1998), so that Liangshanophyllum should be treated as a synonym of Waagenophyllum (Xu in Jia et al., Reference Jia, Xu, Kuang, Zhang and Zuo1977). In this paper, we treat Liangshanophyllum as a subgenus of Waagenophyllum.
Waagenophyllum minutum Zhao, Reference Zhao1981
Figure 4.1–4.3
- 1981
Waagenophyllum minutum Zhao, p. 252, pl. 7, fig. 7a, b.

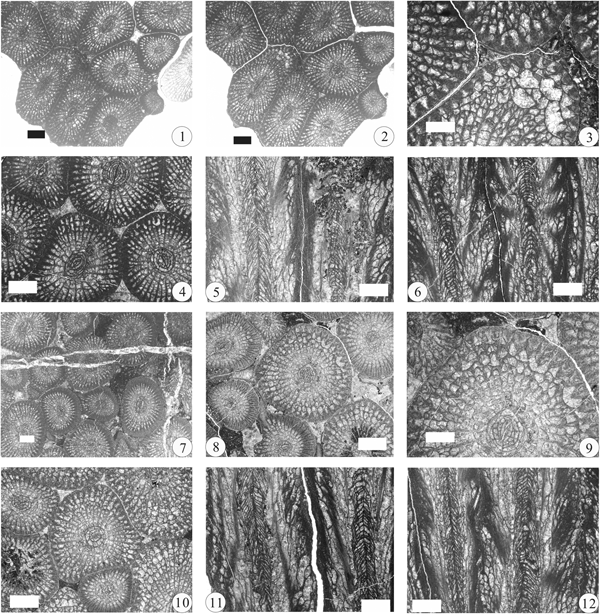
Figure 4. (1–3) Waagenophyllum (Waagenophyllum) minutum Zhao, Reference Zhao1981, Gyanyima, transverse sections and longitudinal section of NIGP 148070 from bed 1. (4–6) Waagenophyllum (W.) intermedium n. sp., Gyanyima, holotype, NIGP 148071 from bed 9, transverse sections and longitudinal section. (7–13) Waagenophyllum (W.) elegantulum He in Luo et al., Reference Luo, He, Wang, Wang, He and Chen1989, Gyanyima: (7–9) transverse sections and longitudinal section of NIGP 148072 from bed 2; (10, 11) transverse section and longitudinal section of NIGP 148073 from bed 6; (12, 13) transverse section and longitudinal section of NIGP 148074 from bed 10. (14, 15) Waagenophyllum (W.) gyanyimaense n. sp., Gyanyima, holotype, NIGP 148075 from bed 7, transverse section and longitudinal section. Scale bars = 2 mm.
Holotype
NIGP 47434 from the Changhsing Formation, Liangshan, Hanzhong, Shaanxi, China (Zhao, Reference Zhao1981, p. 252, pl. 7, fig. 7a, b).
Occurrence
This species occurs in the Gyanyima Formation, bed 1 of the Gyanyima Section, Drhada, South Tibet, the Changhsingian Stage of the upper Permian.
Description
Corallum fasciculate. Corallites generally small and cylindrical. Mean diameter of mature corallites 3.0–3.5 mm. Major septa number 15–17 (Fig. 5). Ratio of septal number to corallite diameter (n/d) 4.7–5.2. Septa gently curved, obviously thickened in the peripheral area, and gradually thinning toward axis. Major septa almost reach to axial column and mostly connect with axial column. Minor septa as long as two-thirds to three-fourths length of major septa. Axial column ovate and loosely constructed, 0.8–1.1 mm in diameter, occupying about one-fourth of diameter; median plate generally detectable; concentric axial tabellae in two to three rows; septal lamellae relatively dense and short. Dissepiments arranged concentrically in transverse section.

Figure 5. Relationship between mean corallite diameter and number of major septa in Waagenophyllum (Waagenophyllum) minutum.
In longitudinal section, dissepimentarium well developed, two to three rows of elongated dissepiments. Tabularium narrow; clinotabulae poorly developed, steeply inclined, and look like elongated dissepiments; horizontal tabulae short and partly dense. Axial column composed of long, steeply ascending, and loosely arranged axial tabellae, a few sinuous septal lamellae, and sinuous and conspicuous median plate.
Material
One specimen (NIGP 148070) with two transverse thin sections and two longitudinal thin sections.
Remarks
This species is characterized by small corallite size and a simple or complex axial column (Zhao, Reference Zhao1981). The present specimens slightly differ from the type in having dense horizontal tabulae and a relatively variable axial column. In addition, the present species differs from other Waagenophyllum species in its smaller number of septa, only larger than Waagenophyllum domarense Yu in Fan et al., Reference Fan, Yu, He, Pan and Li2003, which has 12–14.
Waagenophyllum elegantulum He in Luo et al., Reference Luo, He, Wang, Wang, He and Chen1989
Figure 4.7–4.13
- 1989
Waagenophyllum elegantulum He in Luo et al., p. 123, pl. 8, fig. 6; pl. 59, figs. 7–12.
Holotype
SEM-054 from the Gyanyima Formation, upper Permian, Drhada, South Tibet, China (Luo et al., Reference Luo, He, Wang, Wang, He and Chen1989, p. 123, pl. 8, fig. 6).
Occurrence
This species occurs in the Gyanyima Formation, beds 2, 6, 7, and 10 of the Gyanyima Section, Drhada, South Tibet, the Changhsingian Stage of the upper Permian.
Description
Corallum fasciculate. Corallites cylindrical, sometimes in contact with each other, while in most cases relatively apart. Mean diameter of mature corallites 4.1–6.5 mm (commonly 4.9–6.2 mm). Major septa number 22–28 (commonly 22–24; Fig. 6). Ratio of septal number to corallite diameter 3.6–5.2 (mean n/d 4.0–4.5). Septa commonly wedge shaped, strongly thickened in the peripheral area, forming a stereozone. Major septa gradually thinning toward axis and extend nearly to axial column. Minor septa generally less than half the length of major septa. Wall quite thin, but sometimes thickened by stereozone on peripheral parts. Third order of septa present in the stereozone. Axial column mostly circular to somewhat elliptical, 1.4–2.1 mm in diameter, occupying nearly one-third of diameter of corallites; median plate mostly distinct and thick, straight or sinuous; four to six rows of irregularly shaped axial tabellae; septal lamellae generally discontinuous and variously developed in different corallites. In transverse section, dissepiments arranged concentrically or angulo-concentrically.

Figure 6. Relationship between mean corallite diameter and number of major septa in Waagenophyllum (Waagenophyllum) elegantulum. Specimen A: NIGP148072; Specimen B: NIGP148073; Specimen C: NIGP148074.
In longitudinal section, dissepimentarium well developed; peripherally elongated dissepiments in one to three rows; inner dissepiments largely elongated and look somewhat like clinotabulae. Tabularium narrow; arrangement of clinotabulae variable even in different parts of the same corallite; horizontal tabulae well developed where clinotabulae poorly developed or loosely arranged. Axial column composed of straight and conspicuous median plate, densely or loosely overlapped axial tabellae, and a few sinuous and discontinuous septal lamellae.
Materials
Eight specimens (NIGP 148072–148074, 148093–148097) with 15 transverse thin sections and 16 longitudinal thin sections.
Ontogeny and intraspecific variation
Mean diameter of the immature corallites with 15–18 major septa is less than 3.5 mm. Median plate, axial tabellae, and septal lamellae appear. The range of diameters of the corallites with 20 major septa is relatively wide. Axial column is still small and simple. Mature corallites have commonly 22–24 major septa, some up to 28.
The dissepiments in the inner parts of the dissepimentarium are largely elongated and somewhat like clinotabulae. The clinotabulae always developed where the clinotabulae-shaped dissepiments are not developed.
There are slight differences among the specimens from different horizons. In most corallites from bed 2, the axial column has irregularly shaped axial tabellae and poorly developed septal lamellae. By contrast, most of the axial columns in the corallites from beds 6 and 10 are cobweb shaped and have regularly shaped axial tabellae and developed septal lamellae.
Remarks
Luo et al. (Reference Luo, He, Wang, Wang, He and Chen1989) described W. elegantulum as follows: “Corallum phaceloid, possess connective tubules. Peripheral stereozone is 0.5 mm in width. Septa are 20×2 in number. Dissepiments are 1-2 rows. Clinotabulae are developed. Horizontal tabulae are short.” Although the connective tubules are not shown in the present specimens, the forms described here are very similar to those specimens described by Luo et al. (Reference Luo, He, Wang, Wang, He and Chen1989). The present specimens are fasciculate rather than phaceloid. The growth pattern is changeable even in certain coral species; for example, Disphyllum goldfussi Geinitz, Reference Geinitz1846 alternated from fasciculate in quiet water to phaceloid form in high-energy water (Tsien, Reference Tsien1968), and Siphonodendron martini (Milne-Edwards and Haime, Reference Milne-Edwards and Haime1851) developed connective structure in agitated environments (Poty, Reference Poty1981). Thus, though the present specimens show fasciculate form, we treat them as W. elegantulum.
Corallites that more or less touch each other and form a peripheral stereozone in some specimens were collected from bed 2. These are most similar to the type specimen. This character may be related to a higher-energy environment (Tsien, Reference Tsien1968; Xu and Sando, Reference Xu, Sando, Xu, Luo, Wang, Zhou and Xiao1997).
Waagenophyllum intermedium new species
Figure 4.4–4.6
Holotype
NIGP 148071, upper part of the Gyanyima Formation (Changhsingian Stage), Drhada, South Tibet.
Diagnosis
Waagenophyllum having18–22 (commonly 20 or 22) major septa at mean diameter 3.4–4.7 mm; septa obviously thick in the peripheral area; axial column loosely constructed; dissepimentarium broad, subglobose; elongate dissepiments well developed.
Occurrence
This species occurs in the upper part of the Gyanyima Formation, bed 9, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum fasciculate. Corallites generally cylindrical. In transverse section, corallites approximately circular. Major septa number 18–22 (Fig. 7). Mean diameter of mature corallites 3.4–4.7 mm (commonly 3.8–4.2 mm). Ratio of septal number to corallite diameter 4.3–6.0. Septa slightly curved, obviously thickened in the peripheral area, and gradually thinning toward axis. Major septa rarely connect with axial column except the cardinal septum. Minor septa about one-half to two-thirds the length of major septa. Axial column variable in size and loosely constructed, 0.9–1.7 mm in diameter, occupying about one-third the diameter of mature corallites; median plate mostly sinuous; axial tabellae in one to three rows and generally irregular; septal lamellae relatively loose and generally discontinuous.

Figure 7. Relationship between mean corallite diameter and number of major septa in Waagenophyllum (Waagenophyllum) intermedium.
In longitudinal section, dissepimentarium well developed, dissepiments arranged in two to three rows, peripheral dissepiments subglobose. No distinct boundary between inner elongate dissepiments and clinotabulae. Tabularium narrow; horizontal tabulae loosely developed. Axial column composed of various and loosely arranged axial tabellae, sinuous and discontinuous median plate, and septal lamellae.
Etymology
A reference to intermediate characters in septal number, diameter of corallite, size of axial column, and length of minor septa between W. elegantulum and W. minutum.
Material
One specimen, the holotype NIGP 148071, with two transverse thin sections and two longitudinal thin sections.
Remarks
The present species is closely similar to Waagenophyllum virgalense (Waagen and Wentzel, Reference Waagen and Wentzel1886) and W. klamathensis Stevens, Reference Stevens, Miller and Nestell1987 in septal number and corallite size. However, W. virgalense exhibits a small axial column and well-developed clinotabulae. Waagenophyllum klamathensis is different from the present species in having a large and dense axial column, poorly developed dissepiments, and developed elongate clinotabulae.
The present form also resembles W. elegantulum and W. minutum but exhibits differences and is intermediate in septal number, diameter of corallite, size of axial column, and length of minor septa (Table 1).
Table 1. Comparison of characters among six species of Waagenophyllum at the Gyanyima section.

Waagenophyllum ngariense He, Reference He, Yang and Nie1990
Figure 8.1–8.9
- 1990
Waagenophyllum ngariense He, p. 77, pl. 14, fig. 7a, b.
- 2002
?Waagenophyllum megacolumetum Cheng et al., p. 25, pl. 1, fig. 2, 3.

Figure 8. (1–9) Waagenophyllum (Waagenophyllum) ngariense He, Reference He, Yang and Nie1990, Gyanyima: (1–3, 6) transverse sections and longitudinal sections of NIGP 148076 from bed 2; (4, 5) transverse section and longitudinal section of NIGP 148077 from bed 6; (7–9) transverse sections and longitudinal section of NIGP 148078 from bed 7. (10–15) Waagenophyllum (Liangshanophyllum) clisicolumellum n. sp., Gyanyima: (10–12, 15) holotype, NIGP 148079 from bed 6, transverse sections and longitudinal sections; (13, 14) paratype, NIGP 148080 from bed 7, transverse section and longitudinal section. Scale bars = 2 mm.
Holotype
TF 647 from the Gyanyima Formation, upper Permian, Drhada, South Tibet, China (He, Reference He, Yang and Nie1990, p. 77, pl. 14, fig.7a, b).
Occurrence
This species occurs in the upper part of the Gyanyima Formation, beds 2, 6, and 7, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum fasciculate. Corallites cylindrical. Mature corallites measuring 7.0–9.8 mm (commonly 7.3–9.1 mm) in mean diameter (Fig. 9). Major septa number 25–29 (commonly 26–29). Ratio of septal number to corallite diameter 2.8–3.7 (mean n/d 3.1–3.4). Septa wedge shaped, straight or slightly curved, thickened in the peripheral area, gradually thinning toward axis. Major septa long, extending nearly to axial column. Minor septa about one-third to one-half the length of major septa. Axial column large, 3.1–3.7 mm in diameter in mature corallite, occupying about two-fifths of diameter; median plate straight or curved, sometimes inconspicuous or absent; axial tabellae in six to eight rows, circularly arranged in most corallites; septal lamellae well developed, slightly sinuous, and discontinuous, generally more than 10 radially arranged on both sides of median plate. Dissepiments arranged concentrically, inconspicuous within minor septa area because of the thick septa.

Figure 9. Relationship between mean corallite diameter and number of major septa in Waagenophyllum (Waagenophyllum) ngariense. Specimen A: NIGP148076; Specimen B: NIGP 148077; Specimen C: NIGP 148078.
In longitudinal section, dissepimentarium variable in width; one to two rows of globose dissepiments locally developed in peripheral part; large and elongate dissepiments well developed. Width of tabularium unstable; clinotabulae well developed and difficult to distinguish from the large and elongate dissepiments; high variation in shape and arrangement of clinotabulae; horizontal tabulae short and loosely developed. Axial column composed of steeply inclined and tightly overlapped axial tabellae, sinuous and conspicuous median plate, and sinuous and dense septal lamellae.
Materials
Three specimens (NIGP 148076–148078) with seven transverse thin sections and seven longitudinal thin sections.
Intraspecific variation
The large and elongate dissepiments are well developed and look like clinotabulae in longitudinal section, so it is difficult to distinguish one from another. Clinotabulae-shaped elongate dissepiments and clinotabulae alternately developed in some corallites. For example, clinotabulae-shaped dissepiments well developed, clinotabulae poorly developed in left lateral, largely elongate dissepiments not developed, clinotabulae well developed in right lateral (Fig. 8.3).
Remarks
The present species resembles Waagenophyllum wengchengense Huang, Reference Huang1932 in some characters of septa and columella, but differs from the latter in having larger corallites and more globose dissepiments.
The present species is also similar to Waagenophyllum megacolumetum Cheng et al., Reference Cheng, Wu and Zhang2002 but slightly different from the latter in having a smaller axial column. The latter has a diameter of corallites 7–11 mm, major septa number 24–30, and axial column occupies about half of the diameter.
Waagenophyllum ngariense is different from most species of this genus in forming a large and complex axial column in the early stage and a cardinal septum joined to the axial column in the early stage (Fig. 8.1).
Waagenophyllum tachtabulasicum Ilyina, Reference Ilyina1997
Figure 10.1–10.9
- 1997
Waagenophyllum tachtabulasicum Ilyina, p. 136, pl. 2, fig. 1.

Figure 10. (1–9) Waagenophyllum (Waagenophyllum) tachtabulasicum Ilyina, Reference Ilyina1997, Gyanyima: (1–3) transverse sections and longitudinal section of NIGP 148081 from bed 9; (4–6) transverse sections and longitudinal section of NIGP 148082 from bed 9; (7) transverse section of NIGP 148083 from bed 8; (8, 9) transverse section and longitudinal section of NIGP 148084 from bed 9. (10, 11) Waagenophyllum (W.?) sp. transverse section and longitudinal section of NIGP 148085 from bed 9, Gyanyima. Scale bars = 2 mm.
Holotype
2376/344 PIN from the middle part of Taktabulak Formation, upper Permian, middle reaches of Kur-Istyk River, Southern Pamir.
Occurrence
This species occurs in the upper part of the Gyanyima Formation, beds 8 and 9, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum fasciculate. Corallites more or less round or ovate in transverse section. Mean diameter of mature corallites 7.8–9.3 mm. Major septa number 25–30 (Fig. 11). Ratio of septal number to corallite diameter 3.0–3.8 (mean n/d 3.2–3.5). Septa slightly curved, prominently thickened in basal part, forming peripheral stereozone and gradually thinning toward axis. Major septa long, extending nearly to axial column. Minor septa also long, more than two-thirds the length of the major septa. Axial column variable in both size and structure; median plate is generally sinuous and sometimes inconspicuous or absent; axial tabellae in one to six rows, smoothly circular or irregularly arranged; septal lamellae sinuous and densely arranged. Dissepiments concentrically arranged in transverse section.

Figure 11. Relationship between mean corallite diameter and number of major septa in Waagenophyllum (Waagenophyllum) tachtabulasicum. Specimen A: NIGP 148081; Specimen B: NIGP 148082.
In longitudinal section, dissepimentarium well developed, composed of three to five rows of small globose or subglobose dissepiments in outer part; inner dissepiments one to two rows, elongated or clinotabulae-like elongated, rarely bubble-arc shaped. Tabularium relatively narrow; clinotabulae well developed, slightly inclined where tabularium relatively wide; horizontal tabulae poorly developed. Axial column composed of slightly inclined, loosely or tightly overlapped axial tabellae, sinuous and conspicuous median plate, and relatively sinuous and discontinuous septal lamellae.
Materials
Five specimens (NIGP 148081–148084, 148098) with 14 transverse thin sections and 13 longitudinal thin sections.
Intraspecific variation
The axial column is variable in size, structure, and shape. The mean diameter of larger axial column ranges from 2.3 to 3.5 mm, occupying nearly one-third of corallite diameter. Some axial columns are regularly shaped. Median plate is generally sinuous and inconspicuous or absent, rarely conspicuous and straight. Axial tabellae are arranged in three to six rows. Septal lamellae are sinuous, about 7–15 rows arranged on both sides of the median plate.
Remarks
The present specimen is very similar to type specimens of W. tachtabulasicum from the upper Permian Dorashanmian, middle part of Kur-Istyk River, although only two corallites were collected there. Waagenophyllum tachtabulasicum is similar to W. ngariense in corallite size and septa number but differs from the latter in having a smaller axial column, broad dissepimentarium, and developed globose dissepiments.
Waagenophyllum gyanyimaense new species
Figure 4.14–4.15
Holotype
NIGP148075, Gyanyima Formation (Changhsingian Stage), Drhada, South Tibet.
Diagnosis
Waagenophyllum having about 32 major septa at mean diameter 15 mm; the cardinal septum connected to the axial structure; minor septa about half the length of the major septa; axial column small; elongated dissepiments well developed; clinotabulae steeply inclined.
Occurrence
This species occurs in the Gyanyima Formation, bed 7, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum fasciculate. Corallites ovate in transverse section. Mean diameter of mature corallite 15 mm. Major septa number 32. Septa thickened in basal part, forming narrow peripheral stereozone. Major septa long, partly reach axial column; cardinal septum connected with the outer axial tabellae. Minor septa relatively short, about half the length of the major septa. Axial column occupies rather narrow space, 3.5 mm in diameter, ovate or somewhat round, compactly constructed; median plate short and inconspicuous; axial tabellae in five to six rows, tightly arranged; relatively numerous septal lamellae radially disposed, commonly sinuous and discontinuous. Dissepiments angulo-concentric in transverse section.
In longitudinal section, dissepimentarium broad; globose dissepiments poorly developed, occasionally one to two rows in peripheral part; elongated dissepiments two to four rows. Tabularium narrow; steeply inclined clinotabulae well developed; horizontal tabulae short, loosely arranged. Median plate sinuous; axial tabellae slightly inclined and tightly overlapped, partly variable in shape; septal lamellae sinuous and discontinuous, sometimes inconspicuous.
Etymology
A reference to the location name Gyanyima, where specimens were collected.
Material
One specimen, the holotype NIGP 148075, with two transverse thin sections and one longitudinal thin section.
Remarks
The present species is similar to Waagenophyllum polyseptatum Minato, Reference Minato1955, W. compactum Minato and Kato, Reference Minato and Kato1965, W. ganhaiziense Fan, Reference Fan1978, and W. zandaense He, Reference He, Yang and Nie1990 in septal number and corallite size. The detailed distinction of them is listed in Table 2.
Table 2. Distinctive characters of Waagenophyllum with about 30–34 major septa.

Waagenophyllum? sp.
Figure 10.10–10.11
Occurrence
This species occurs in the Gyanyima Formation, bed 9, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallites nearly round in transverse section. Major septa number no more than 18. Minor septa short, less than one-third of major septa. Cardinal septum reaches the center of corallite. In longitudinal section, one to two rows of small globose or subglobose dissepiments in outer part; inner elongated dissepiments three to six rows. Horizontal tabulae short and loose.
Material
One specimen (NIGP 148085) with one transverse thin section and two longitudinal thin sections.
Remarks
The present specimen is poorly preserved. An axial column does not appear in either transverse or longitudinal sections. It is possible that the present specimen is a species of Waagenophyllum (Huayunophyllum). Waagenophyllum (Huayunophyllum) differs from Waagenophyllum (Waagenophyllum) in having a simpler axial column, composed of a median plate and axial tabellae.
Subgenus Liangshanophyllum Tseng, Reference Tseng1949
Type species
Liangshanophyllum lui Tseng, Reference Tseng1949, p. 101–102, pl. 1, figs. 2a, b, upper Permian, Liangshan, Shaanxi, southern China.
Liangshanophyllum clisicolumellum new species
Figure 8.10–8.15
Type specimens
Holotype NIGP 148079 and paratype NIGP 148080, Gyanyima Formation (Changhsingian Stage), Drhada, South Tibet.
Diagnosis
Liangshanophyllum having 22–25 major septa at mean diameter 5.8–7.9 mm; major septa long; minor septa about one-half to two-thirds the length of major septa; axial column mostly cobweb shaped; tabularium broad; clinotabulae loosely arranged.
Occurrence
This species occurs in the Gyanyima Formation, beds 6 and 7, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum fasciculate. In transverse section, corallites approximately circular. Mean diameter of mature corallites 5.8–7.9 mm (commonly 6.1–7.4 mm). Major septa number is 22–25 (Fig. 12). Ratio of septal number to corallite diameter 3.2–3.8 (mean n/d 3.6). Septa slightly curved, weakly thickened in the peripheral area, forming narrow stereozone, and thinning toward axis. Major septa long, extending nearly to axial column. Cardinal septum commonly connected with the outer axial tabellae. Minor septa about one-half to two-thirds the length of major septa. Axial column mostly cobweb-like appearance, occupying nearly one-third of diameter; median plate straight or curved, short, and commonly extends into the inner axial tabellae; septal lamellae discontinuous, five to eight radially arranged on both sides of median plate; axial tabellae in four to five rows, regularly arranged in most corallites. Dissepiments concentrically arranged.

Figure 12. Relationship between mean corallite diameter and number of major septa in Waagenophyllum (Liangshanophyllum) clisicolumellum. Specimen A: NIGP 148079 (holotype); Specimen B: NIGP 148080 (paratype).
In longitudinal section, dissepimentarium narrow, one to three rows globose and elongate dissepiments. Tabularium broad; clinotabulae slightly inclined and loosely arranged; horizontal tabulae poor developed. Median plate sinuous and conspicuous; septal lamellae thin and sinuous; axial tabellae steeply inclined and tightly overlapped.
Etymology
A reference to a cobweb-shaped axial column, similar to that of Clisiophyllum.
Materials
Two specimens, the holotype NIGP 148079, with two transverse thin sections and two longitudinal thin sections, and the paratype NIGP 148080, with two transverse thin sections and two longitudinal sections.
Remarks
The present species is closely similar to Liangshanophyllum huishuiense major Wu and Zhao, Reference Wu and Zhao1983 in septal number, corallite size, tabulae shape, and axial column size. However, L. huishuiense major is different from the present species in having shorter septa and poorly developed dissepiments.
Liangshanophyllum cylindricum Ding, Reference Ding1995 is similar to the present species in corallite size but exhibits marked differences in having a small and simple axial column and relatively short minor septa.
Genus Ipciphyllum Hudson, Reference Hudson1958
Type species
Ipciphyllum ipci Hudson, Reference Hudson1958, p. 179, pl. 33, figs. 1, 2, 3, 7, 10, pl. 35, fig. 4. Wentzelella Limestone, middle Permian, Geli Khana, Ora, northern Iraq.
Ipciphyllum naoticum new species
Figure 13.1–13.3
Holotype
NIGP 148086, Gyanyima Formation (Changhsingian Stage), Drhada, South Tibet.

Figure 13. (1–3) Ipciphyllum naoticum n. sp., Gyanyima: (1, 3) holotype, transverse section and longitudinal section of NIGP 148086 from bed 1; (2) enlarged view of (1) showing weakly naotic structure of septa. (4–8) Ipciphyllum floricolumellum n. sp., Gyanyima: (4, 5, 7) holotype, NIGP 148087 from bed 2, transverse section and longitudinal section; (6, 8) paratype, NIGP 148088 from bed 2, transverse sections and longitudinal section. (9–14) Ipciphyllum zandaense n. sp., Gyanyima: (9–11, 14) holotype, NIGP 148089 from bed 7, transverse sections and longitudinal sections; (12, 13) paratype, NIGP 148090 from bed 7, transverse sections. Scale bars = 2 mm.
Diagnosis
Ipciphyllum having 18–24 (commonly 19–21) major septa at mean diameter 5.4–7.3 mm (commonly 6–7 mm); septa weakly naotic; dissepiments three to five rows; tabularium narrow, clinotabulae well developed.
Occurrence
This species occurs in the Gyanyima Formation, bed 1, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum cerioid. In transverse section, corallites polygonal generally with five to six sides, rarely seven, even nine, sides. Mean diameter (measured through the axis from the approximate midpoints of opposite sides) of mature corallites 5.4–7.3 mm (commonly 6–7 mm). Major septa number 18–24 (commonly19–21; Fig. 14). Ratio of septal number to corallite diameter 2.7–3.6. Wall thin, slightly undulatory. Septa wedge shaped, strongly thickened in peripheral part but becoming thin where naotic and lateral dissepiments developed. Major septa long, closely approach or (rarely) join axial structure. Minor septa thinner than the major septa and about two-thirds to four-fifths the length of the major septa. Axial column ovate or somewhat round, occupying about one-fourth of diameter; median plate straight and slightly thick; axial tabellae three to five rows; septal lamellae 5–10, radially arranged on both sides of median plate. Dissepiments commonly angulo-concentric or concentric.

Figure 14. Relationship between mean corallite diameter and number of major septa in Ipciphyllum naoticum.
In longitudinal section, dissepiments three to five rows, globose or subglobose and mostly small, some elongated dissepiments present. Tabularium narrow; tightly arranged clinotabulae well developed, slightly inclined where tabularium relatively wide; horizontal tabulae appear in part where elongated dissepiments well developed and tabularium quite narrow. Axial tabellae irregularly shaped and overlapped tightly or loosely; septal lamellae dense and conspicuous.
Etymology
A reference to weakly developed naotic septa.
Material
One specimen, the holotype NIGP 148086, with four transverse thin sections and one longitudinal thin section.
Ontogeny
Ontogeny of the new species cannot be discussed in detail without serial sections, but the corallite character from the neanic stage can be described according to the available sections. Mean diameter of the corallites with 15–17 major septa number is variable, commonly less than 5.5 mm. The naotic structure and lateral dissepiments partly appear. The septa still strongly thickened in peripheral part of this stage. Major septa number of mature corallites is commonly more than 19–21. The naotic structure and lateral dissepiments well developed in this later stage. Septa become thin in the peripheral part and obviously thinner than septa of the central part within the tabularium.
Remarks
The present species is similar to I. timoricum Gerth, Reference Gerth1921 except for developing naotic structure and lateral dissepiments. Ipciphyllum naoticum n. sp. differs from other species of this genus in having weakly naotic septa.
Ten late Permian Ipciphyllum species have been reported in China: I. anshunense (Zhao, Reference Zhao1976), I. exilum Xu, Reference Xu, Feng, Xu, Lin and Yang1984b, I. heshanense Wu and Zhao, Reference Wu and Zhao1983, I. markaense (Zhao, Reference Zhao1976), I. minor (Wu and Zhao, Reference Wu and Zhao1983), I. shiqianense (Zhao and Wang in Wang, Reference Wang1978), I. simplex Wu, Reference Wu1963, I. stereoseptatum Xu, Reference Xu, Feng, Xu, Lin and Yang1984b, I. subtimoricum var. kwangsiense Wu, Reference Wu1963, and I. ziyunense Wang, Reference Wang1978 (Wu, Reference Wu1963; Zhao, Reference Zhao1976, Reference Zhao1984; Jia et al., Reference Jia, Xu, Kuang, Zhang and Zuo1977; Wang, Reference Wang1978; Wu and Zhao, Reference Wu and Zhao1983; Xu, Reference Xu, Feng, Xu, Lin and Yang1984b). Two of them (I. minor and I. shiqianense) have developed lateral dissepiments (Wang, Reference Wang1978; Wu and Zhao, Reference Wu and Zhao1983).
Ipciphyllum floricolumellum new species
Figure 13.4–13.8
Type specimens
Holotype NIGP 148087 and paratype NIGP 148088, Gyanyima Formation (Changhsingian Stage), Drhada, South Tibet.
Diagnosis
Ipciphyllum having 17–19 major septa at mean diameter 4.2–5.7 mm; septa wedge shaped; minor septa about two-thirds the length of the major septa; axial column large and mostly flower shaped; slightly inclined clinotabulae well developed.
Occurrence
This species occurs in the Gyanyima Formation, bed 2, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum cerioid. In transverse section, corallites polygonal generally with five to six sides, rarely 7 or 8 sides. Mean diameter of mature corallites 4.2–5.7 mm (commonly 4.5–5.5 mm). Major septa number 16–21 (commonly 17–19; Fig. 15). Ratio of septal number to corallite diameter 3.3–4.3 (mean n/d 3.6–3.9). Wall thin, slightly undulatory. Septa wedge shaped, strongly thickened in peripheral part and formed stereozone. Major septa relatively straight, become thinner gradually toward axis, partly reach axial column. Minor septa abruptly thin in peripheral part, about two-thirds the length of the major septa. Axial column ovate or somewhat round, occupying one-fourth to one-third of diameter, flower shaped; median plate straight and thick; axial tabellae five to seven rows, arranged tightly in outer part; septal lamellae poorly developed. Dissepiments commonly concentric in transverse section.

Figure 15. Relationship between mean corallite diameter and number of major septa in Ipciphyllum floricolumellum. Specimen A: NIGP 148087 (holotype); Specimen B: NIGP 148088 (paratype).
In longitudinal section, complete aspect of dissepimentarium hardly exhibited because of the thick septa; globose and subglobose dissepiments three to five rows, mostly small; large elongated dissepiments partly developed. Tabularium broad, occupying approximately one-third of diameter; slightly inclined clinotabulae well developed, commonly tightly arranged; horizontal tabulae poorly developed. Median plate straight and conspicuous; axial tabellae nearly perpendicular to median plate in axial part, abruptly inclined in distal part and tightly overlapped. Septal lamellae inconspicuous.
Etymology
A reference to a character of flower-shaped axial column.
Materials
Two specimens, the holotype NIGP 148087, with three transverse thin sections and one longitudinal thin section, and the paratype NIGP 148088, with four transverse thin sections and two longitudinal thin sections.
Remarks
Ipciphyllum floricolumellum differs from other species of this genus in having thick septa in its largely flower-shaped axial column. The present species is also distinct from I. timoricum in having a smaller corallite and fewer septa.
It is important to note that the mean diameters of corallites were determined in transverse section by taking the average of the side-corner corallite diameters, measured through the axis from the approximate midpoints of all sides. The diagonal distance cannot be used to measure odd polygonal corallites; the distance between the corners and the midpoints of opposing sides is unsuitable for such polygonal corallites. Different measuring standards will result in artificial error. Thus, measurements from the corners of corallites were avoided.
Ipciphyllum zandaense new species
Figure 13.9–13.14
Type specimens
Holotype NIGP 148089 and paratype NIGP 148090, Gyanyima Formation (Changhsingian Stage), Drhada, South Tibet.
Diagnosis
Ipciphyllum having 17–22 major septa at mean diameter 3.2–5.3 mm; minor septa about three-fourths to four-fifths the length of the major septa; axial column about one-fourth of diameter.
Occurrence
This species occurs in the Gyanyima Formation, bed 7, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum cerioid, or partly subastraeoid. Corallites polygonal, generally with five to six sides, rarely 7 sides. Mean diameter of mature corallites 3.2–5.3 mm (commonly 4–5 mm). Wall thin, slightly undulatory. Major septa number 17–22 (commonly 18–19; Fig. 16). Ratio of septal number to corallite diameter 4.0–5.7 (mean n/d 4.7–5.2). Septa slightly thickened in basal part, become thinner gradually toward axis. Major septa reach or nearly reach axial column. Minor septa long, about three-fourths to four-fifths the length of the major septa. Axial column ovate or fusiform, occupying one-fourth of diameter; median plate mostly short, straight or curved, few inconspicuous; axial tabellae circular, generally three rows; septal lamellae three to five, radially arranged on both sides of median plate. Dissepiments commonly concentric or angulo-concentric in transverse section.

Figure 16. Relationship between mean corallite diameter and number of major septa in Ipciphyllum zandaense. Specimen A: NIGP 148089 (holotype); Specimen B: NIGP 148090 (paratype).
In longitudinal section, dissepimentarium broad; dissepiments two to four rows in outer part, globose or subglobose with different size; inner elongated dissepiments two to three rows, gradually larger toward inside. Tabularium partly very narrow; slightly inclined clinotabulae loosely arranged; horizontal tabulae developed where clinotabulae poorly developed. Median plate straight and conspicuous; axial tabellae abruptly inclined and regularly overlapped; septal lamellae sinuous and discontinuous.
Etymology
A reference to a location name of Drhada, South Tibet.
Materials
Two specimens, the holotype NIGP 148089, with three transverse thin sections and three longitudinal thin sections, and the paratype NIGP 148090, with four transverse thin sections and one longitudinal thin section.
Remarks
The present species is similar to Ipciphyllum timoricum in the number of septa but differs in the thinner shape of septa. In addition, the latter species shows distinction in having a larger corallite size and more tightly arranged axial tabellae.
Genus Gyanyimaphyllum new genus
Type species
Gyanyimaphyllum crassiseptatum n. gen. n. sp.
Diagnosis
Corallum fasciculate, generally tightly gathered and cerioid shaped; corallites cylindrical or somewhat polygonal; laterally offset; septa two orders; naotic structure developed in basal part of septa; axial column generally circular, composed of median plate, axial tabellae, and some radially arranged septal lamellae; dissepimentarium broad, composed of concentric globose, elongated dissepiments; cystosepiments partly developed; clinotabulae and horizontal tabulae well developed.
Occurrence
Changhsingian Stage of the upper Permian, Gyanyima of Drhada, South Tibet.
Etymology
A reference to the location name of Gyanyima, which was where all specimens were collected.
Remarks
The new genus differs from other waagenophyllid corals by its fasciculate corallum, generally tightly gathered and cerioid shaped, and thick septa, some of which are replaced by a column of naotic structure of dissepiments in their peripheral parts.
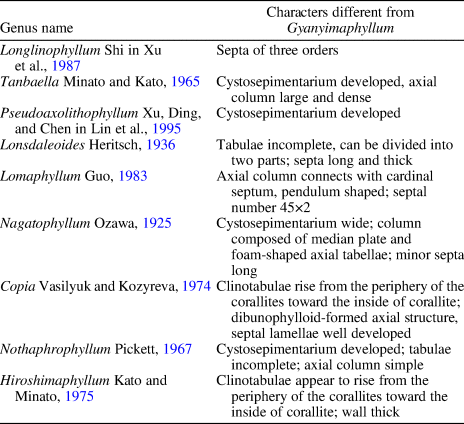
The major differences between Gyanyimaphyllum and other fasciculate genera with naotic structure are presented in Table 3.
Table 3. Major differences between Gyanyimaphyllum and other fasciculate genera that have naotic structure.
Gyanyimaphyllum crassiseptatum new species
Figure 17

Figure 17. Gyanyimaphyllum crassiseptatum n. gen. n. sp., Gyanyima: (1, 2, 4–6) holotype, NIGP 148091 from bed 2, transverse sections and longitudinal sections; (3) enlarged view of (2) showing naotic structure of septa. (7, 8, 10–12) paratype, NIGP 148092 from bed 2, transverse sections and longitudinal sections; (9) enlarged view of (8) showing naotic structure of septa. Scale bars = 2 mm.
Type specimens
Holotype NIGP 148091 and paratype NIGP 148092, Gyanyima Formation (Changhsingian Stage), Drhada, South Tibet.
Diagnosis
Corallum fasciculate, generally tightly gathered and cerioid shaped; corallites are cylindrical or polygonal; naotic structure developed in basal part of dilated septa; dissepimentarium broad, cystosepiments partly developed; both clinotabulae and horizontal tabulae well developed.
Occurrence
This species occurs in the Gyanyima Formation, bed 2, Gyanyima Section, Drhada, South Tibet; the Changhsingian Stage of the upper Permian.
Description
Corallum fasciculate to subcerioid. In transverse section, some corallites ovate or approximately circular, others out of shape because corallites gathered too tightly. Mean diameter of mature corallites 5.0–10.4 mm (commonly 6.0–8.3 mm; Fig. 18). Major septa number 20–26 (commonly 22–24). Ratio of septal number to corallite diameter is 2.9–3.9 (mean n/d 3.2–3.3). Septa prominently thickened in peripheral part and formed stereozone, gradually thinner to axis. Naotic structure developed in basal part of septa in mature stage. Major septa long, almost connect with axial column. Minor septa about two-thirds to three-fourths the length of major septa. Axial column small, round or elliptical, 1.1 to 2.2 mm (commonly 1.5–2.1 mm) in diameter; median plate straight or conspicuous, generally connects with the cardinal septum; circular axial tabellae four to six rows; four to seven discontinuous septal lamellae disposed on both sides of median plate. Dissepiments concentrically arranged, inconspicuous in peripheral part; cystosepiments partly developed, especially conspicuous in the ephebic and gerontic stages, naotic structure corresponding well developed.

Figure 18. Relationship between mean corallite diameter and number of major septa in Gyanyimaphyllum crassiseptatum. Specimen A: NIGP 148091 (holotype); Specimen B: NIGP 148092 (paratype).
In longitudinal section, dissepimentarium broad; 2–4 rows globose or subglobose dissepiments in outer zone, variable in size; inner largely elongated dissepiments look like clinotabulae. Tabularium narrow; clinotabulae elongated and loosely arranged; horizontal tabulae short and tightly arranged. Median plate straight and thick; axial tabellae steeply inclined and tightly overlapped; septal lamellae inconspicuous.
Etymology
A reference to a character of naotic structure developed in basal part of dilated septa.
Materials
Two specimens, the holotype NIGP 148091, with five transverse thin sections and two longitudinal thin sections, and the paratype NIGP 148092, with five transverse thin sections and four longitudinal thin sections.
Ontogeny
Although no serial thin sections were especially prepared for study of ontogeny, some corallites presented different stages that can be observed in abundant transverse sections, so the ontogeny can be analyzed to some extent.
In the first stage, mean diameter of corallites less than 2.5 mm. Major septa number is fewer than 12. Minor septa poorly developed. In the late phase of this stage, septa still thin in the basal part, dissepiments conspicuous in transverse section, embryonic axial column appears.
In the second stage, mean diameter of corallites no more than 4.0 mm. Major septa number is about 14–16. Minor septa developed. Septa obviously thicken and stereozone formed. Dissepiments are inconspicuous in transverse section. Axial column is small.
In the third stage, mean diameter of corallites is abruptly increased. Major septa number is 18–21. The positive relationship between septa number and corallite size is evident. The septa partly become thin in peripheral part; stereozone becomes narrow. Naotic structure weakly developed in basal part of septa. Axial column structure clearer than in the former stage, slightly increased in column size.
In the fourth stage, major septa number is 22–26. Mean diameter of corallites ranges from 6.0 to 8.5 mm, a few more than 10 mm, but the diameter of corallites is not obviously increased compared to the rapidly increasing septal number. Naotic structure obviously developed in basal part of septa. Cystosepiments are strongly developed and interrupt both major and minor septa. The size range of axial column relatively steady, mostly 1.5–2.2 mm in mean diameter. The axial tabellae form more rows than those in the former stage.
It is worth mentioning that Gyanyimaphyllum crassiseptatum n. gen. n. sp. and Ipciphyllum naoticum n. sp. are possibly the latest occurrences of fasciculate and massive rugose corals with naotic structure, respectively, in the world. By contrast, Lobatophyllum Cheng et al., Reference Cheng, Wu and Zhang2002 is the last representative of solitary naotic rugose corals, which occurs in the late Permian in north Tibet (Cheng et al., Reference Cheng, Wu and Zhang2002).
Remarks
The tightly gathered character of corallites in the present specimen resembles some species of Praewentzelella Minato and Kato, Reference Minato and Kato1965, but the latter show marked difference from the former in development of tertiary or more orders of septa.
Acknowledgments
We are grateful to S.-Z. Shen and W.-Z. Li for providing specimens. I. Somerville of University College Dublin, Ireland, critically read the manuscript and made many helpful improvements. This research is financially supported by the Second Tibetan Plateau Scientific Expedition and Research (2019QZKK0706), the Strategic Priority Research Program (B) of the Chinese Academy of Sciences (XDB18030400, 26000000), the Ministry of Science and Technology Foundation Project (2013FY111000), National Science Foundation of China (91855205), and the international academic exchange program of Andong National University.