



Introduction
The paleobiogeography of the Central Andes during the Ordovician has been extensively investigated using benthic fossil groups such as brachiopods from Argentina (Benedetto et al., Reference Benedetto, Vaccari, Waisfeld, Sánchez, Foglia and Bassett2009 and references therein), Chile (Benedetto et al., Reference Benedetto, Niemeyer, González and Brussa2008), Bolivia (Havlíček and Branisa, Reference Havlíček and Branisa1980; Benedetto, Reference Benedetto2013), and Peru (Hughes et al., Reference Hughes, Rickards and Williams1980; Gutiérrez-Marco and Villas, Reference Gutiérrez-Marco and Villas2007; Villas et al., Reference Villas, Colmenar and Gutiérrez-Marco2015); trilobites from Argentina (Tortello and Esteban, Reference Tortello and Esteban2003; Waisfeld and Vaccari, Reference Waisfeld, Vaccari and Benedetto2003 and references therein), Bolivia (Přibyl and Vaněk, Reference Přibyl and Vaněk1980), and Peru (Hughes et al., Reference Hughes, Rickards and Williams1980; Gutiérrez-Marco et al., Reference Gutiérrez-Marco, Rábano, Azeñolaza and Chacaltana2015); bivalves and rostroconch mollusks from Argentina (Sánchez, Reference Sánchez and Benedetto2003, Reference Sánchez2008, and references therein) and Bolivia (Babin and Branisa, Reference Babin and Branisa1987; Sánchez and Babin, Reference Sánchez and Babin2005); sponges from Argentina (Carrera and Rigby, Reference Carrera and Rigby1999; Carrera, Reference Carrera and Benedetto2003); and ostracods from Argentina (Salas, Reference Salas2002a, Reference Salasb; Williams et al., Reference Williams, Floyd, Salas, Siveter, Stone and Vannier2003).
The Tremadocian–Floian brachiopod assemblages of the Cordillera Oriental, including its Argentinian, Bolivian, and Peruvian segments, show high endemism during this time, sharing only one genus with the western Puna Region of Argentina (Lipanorthis Benedetto in Benedetto and Carrasco, Reference Benedetto and Carrasco2002), suggesting proximity but a certain degree of isolation between these two regions. The faunal affinity in the Cordillera Oriental indicates a strong peri-Gondwanan influence (from both high and low latitudes) as demonstrated by the occurrence of Kvania Havlíček, Reference Havlíček1994, Gondwanorthis Benedetto and Muñoz, Reference Benedetto and Muñoz2017, Euorthisina Havlíček, Reference Havlíček1950, and Tarfaya Havlíček, Reference Havlíček1971. Celtic genera (Astraborthis Williams, Reference Williams1974 and Monorthis Bates, Reference Bates1968) are also present in the assemblages of the Cordillera Oriental, implying direct links with Avalonia during Tremadocian and Floian times.
The Argentinian western Puna and Famatina Lower Ordovician assemblages have several taxa in common such as the genera Hesperonomia Ulrich and Cooper, Reference Ulrich and Cooper1936, Tritoechia Ulrich and Cooper, Reference Ulrich and Cooper1936, and Rugostrophia Neuman, Reference Neuman and Dutro1971 and the species Monorthis transversa Benedetto, Reference Benedetto2003 and even share endemic genera such as Pinatotoechia Benedetto, Reference Benedetto2001 and Trigonostrophia Benedetto, Reference Benedetto2003. Faunal evidence is consistent with the existence of a continuous Puna–Famatinian arc likely connecting with Avalonia (Benedetto, Reference Benedetto, Pankhurst and Rapela1998; Ramos, Reference Ramos and Folguera2018) and retroarc foreland basin (Astini and Dávila, Reference Astini and Dávila2004) extending between these two regions along the proto-Andean margin of Gondwana (Benedetto, Reference Benedetto, Pankhurst and Rapela1998). Despite the strong endemicity of the Puna–Famatinian brachiopod assemblages, certain faunal exchange with other regions occurred during that time as suggested by associations of several Celtic and low-latitude genera (Schizambon Walcott, Reference Walcott1884, Rugostrophia, Ffynnonia Neuman and Bates, Reference Neuman and Bates1978, and Hesperonomia Ulrich and Cooper, Reference Ulrich and Cooper1936), minor representatives of high-latitude peri-Gondwana (Incorthis Havlíček and Branisa, Reference Havlíček and Branisa1980), and abundant cosmopolitan genera. The Celtic signature of the Puna–Famatinian assemblages was also supported by multivariate analysis (Benedetto, Reference Benedetto2004; Benedetto et al., Reference Benedetto, Vaccari, Waisfeld, Sánchez, Foglia and Bassett2009). Faunal exchange among the Central Andean Basin (comprising the Altiplano/Puna, the Cordillera Oriental, and the Subandean Ranges), the Famatina Basin, and the Cuyania Terrane (Precordillera) likely started during the late Floian–Dapingian, slightly before the estimated time for docking of the Cuyania Terrane against Gondwana (Benedetto, Reference Benedetto2004; Lavié and Benedetto, Reference Lavié and Benedetto2016, Reference Lavié and Benedetto2019). At that time, the separation of this terrane from the main continent could have been small enough to facilitate fluid faunal exchange (Benedetto, Reference Benedetto2004; Lavié and Benedetto, Reference Lavié and Benedetto2016, Reference Lavié and Benedetto2019).
As stated in the preceding, the faunal affinities of the Early Ordovician brachiopod assemblages of the Argentinian segment of the Central Andes are well known. However, the biogeographic relations of the Peruvian Altiplano, which is the northern continuation of the Bolivian Altiplano and Argentinian western Puna, have remained obscured until now. The assemblages of the Umachiri Formation described in this paper contain the only Early–?Middle Ordovician fossils known in the Peruvian Altiplano and, thus, could potentially shed light on the paleogeography of this region and its biogeographic affinity with other terranes of the southwestern margin of Gondwana during the Early–Middle Ordovician.
Geographical and geological setting
The Andean Altiplano (= high plains morphotectonic region) extends from western Bolivia to southern Peru, comprising base elevation ranging from 3,600 to 4,500 m (Fig. 1). Geologically, it is a structural block with a distinct tectonostratigraphic history from the Eastern and Western Cordilleras of the Central Andes (Carlotto, Reference Carlotto2013). The basement of the Altiplano is interpreted as the Arequipa Terrane (for extent and boundaries of this terrane, see Ramos, Reference Ramos2008). The partial detachment of this terrane from the Amazonian Craton led to the development of a large retroarc basin of Ordovician to Devonian strata known as the Central Andean Basin (Sempere, Reference Sempere, Tankard, Suárez and Welsink1995; Bahlburg et al., Reference Bahlburg, Carlotto and Cárdenas2006; Ramos, Reference Ramos and Folguera2018).
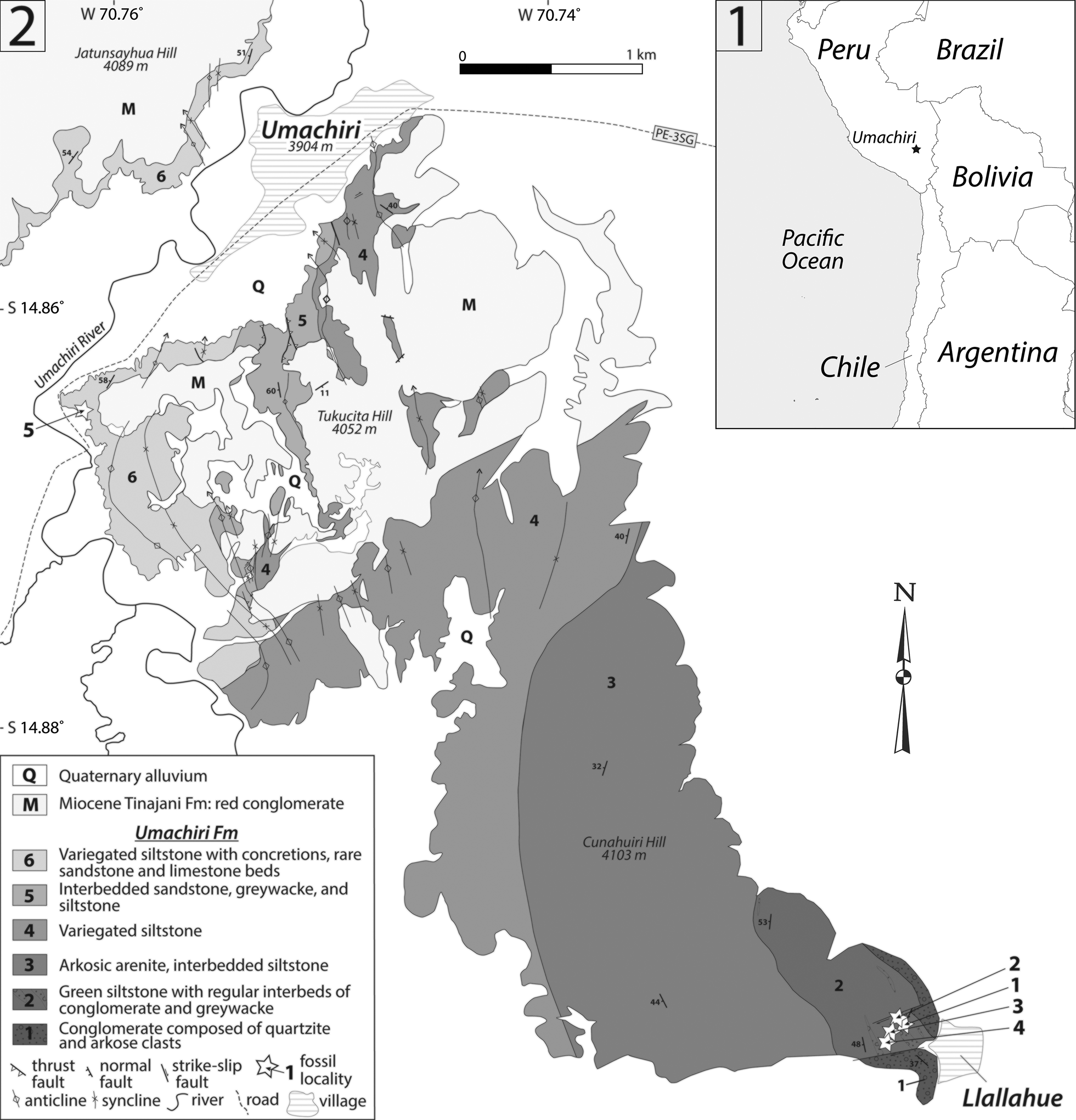
Figure 1. (1) Geographical location of Umachiri village. (2) Detailed geological map of the study area. Numbered white stars indicate fossiliferous horizons and localities: 1, samples B1799 and B1806; 2, samples K1708 and B1807; 3, sample B1808; 4, sample B1798; 5, graptolite locality from Cerrón and Chacaltana (Reference Cerrón and Chacaltana2002, Reference Cerrón and Chacaltana2003).
Upper Ordovician siliciclastic rocks have been identified near Lake Titicaca, an area known as the Calapuja Formation (Laubacher, Reference Laubacher1974, Reference Laubacher1977, Reference Laubacher1978; Laubacher et al., Reference Laubacher, Gray and Boucot1982). This unit has yielded abundant brachiopods, trilobites, gastropods, bivalves, echinoderms, bryozoans, cornulitids, cnidarians, and sponge spicules, suggesting a Sandbian age for these strata (Laubacher, Reference Laubacher1977; Villas et al., Reference Villas, Colmenar and Gutiérrez-Marco2015; Gutiérrez-Marco et al., Reference Gutiérrez-Marco, Carrera, Ghienne, Chacaltana, Gurdebeke, De Weirdt, Vandenbroucke and Cramer2016; Vinn and Gutiérrez-Marco, Reference Vinn and Gutiérrez-Marco2016; Ebbestad and Gutiérrez-Marco, Reference Ebbestad and Gutiérrez-Marco2019). Previously, outcrops identified as the Calapuja Formation north of Ayaviri have been tentatively correlated with the Sandia and San Gabán formations of the Peruvian Cordillera Oriental (Díaz-Martínez et al., Reference Díaz-Martínez, Acosta, Cárdenas, Carlotto and Rodríguez2001). Older strata crop out 15 km to the west of Ayaviri, near Umachiri village, in a succession that has been referred to as the Serie Umachiri (Flores and Rodríguez, Reference Flores and Rodríguez1999; Carlotto et al., Reference Carlotto, Cárdenas, Bahlburg, Westervoss, Flores, Cerpa, Dávila, Carlotto and Chalco2004; Ibarra et al., Reference Ibarra, Mamani, Rodriguez, Sempere, Carlotto and Carlier2004), also known as the Umachiri beds (Bahlburg et al., Reference Bahlburg, Carlotto and Cárdenas2006, Reference Bahlburg, Vervoort, DuFrane, Carlotto, Reimann and Cárdenas2011). These beds are currently being redefined as the Umachiri Formation, and it comprises approximately 2,400 m of conglomerate, sandstone, greywacke, siltstone, and rare limestone (Fig. 2). Preliminary results suggest a correlation of this unit with the Lower to Middle Ordovician Verónica and San José formations of southern Peru.

Figure 2. Composite stratigraphy of the Umachiri Formation at Umachiri. Numbered white stars indicate fossiliferous intervals: 1, samples B1799 and B1806; 2, samples K1708 and B1807; 3, sample B1808; 4, sample B1798; 5, graptolite sample from Cerrón and Chacaltana (Reference Cerrón and Chacaltana2002, Reference Cerrón and Chacaltana2003). si = silt; f = fine sand; m = medium sand; c = coarse sand; gr = gravel.
Materials and methods
The material for this study was collected by one of us (EBH) during field campaigns in 2017 and 2018. Five fossiliferous horizons were identified in the Umachiri Formation (Figs. 1, 2). Four horizons (localities 1–4) are situated 100–150 m from the base of the formation, yielding a brachiopod-dominated assemblage with scattered trilobite sclerites and a single columnal plate of pelmatozoan echinoderms; another fossiliferous horizon is located at about 1,700 m (locality 5), yielding poorly preserved graptolites (identified by Cerrón and Chacaltana, Reference Cerrón and Chacaltana2002). All fossil remains are preserved as internal and external molds. Brachiopods occur in localities 1–4 and will be described herein. Fossils have been prepared using mechanical methods in the labs of the Universidad Complutense of Madrid. Latex casts of the specimens were subsequently prepared for replicating the internal and external features. Fossils and latex casts were whitened using magnesium oxide smoke for photography and studied under binocular microscopes.
Repository and institutional abbreviation
Original specimens are deposited in the paleontological collection (prefixed CPI) of the Instituto Geológico, Minero y Metalúrgico (INGEMMET) in Lima, Peru.
Systematic paleontology
(by J. Colmenar)
The use of open nomenclature follows Bengtson (Reference Bengtson1988), and the synonymy lists follow recommendations by Matthews (Reference Matthews1973).
Phylum Brachiopoda Duméril, Reference Duméril1806
Subphylum Rhynchonelliformea Williams et al., Reference Williams, Carlson, Brunton, Holmer and Popov1996
Class Strophomenata Williams et al., Reference Williams, Carlson, Brunton, Holmer and Popov1996
Order Billingsellida Schuchert, Reference Schuchert1893
Suborder Billingsellidina Öpik, Reference Öpik1934
Superfamily Polytoechioidea Öpik, Reference Öpik1934
Family Polytoechiidae Öpik, Reference Öpik1934
Genus Enriquetoechia new genus
Type species
Enriquetoechia umachiriensis n. gen. n. sp., from the lower part of the Umachiri Formation, Peruvian Altiplano, Floian–?Dapingian (Lower–Middle Ordovician).
Diagnosis
As for the species by monotypy.
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Etymology
After Enrique Villas, brachiopod researcher, who described the first Ordovician brachiopod species of Peru.
Remarks
Phylogeny of Order Billingsellida has been recently revised by Topper et al. (Reference Topper, Harper and Brock2013) suggesting that polytoechioids are derived billingselloids, distinct from the clitambonitoids that are a sister group.
There have been extensive discussions about which are the key taxonomical characters for discriminating among the Billingsellida. Wright and Rubel (Reference Wright and Rubel1996) suggested the use of structures related with their vital functions as key features for superfamily discrimination within this order. These criteria were followed subsequently by several authors (Rubel and Wright in Williams et al., Reference Williams, Brunton, Carlson, Alvarez, Ansell and Baker2000; Popov et al., Reference Popov, Vinn and Nikitina2001; Vinn, Reference Vinn2001; Topper et al., Reference Topper, Harper and Brock2013, among others). Of special significance are the spondylium/pseudospondylium and related structures and the structures covering the delthyrium and notothyrium.
The delthyrium in the clitambonitine varies from open to partially covered by chilidial plates to completely covered by either a deltidium or a pseudodeltidium. However, the distinction between deltidium and pseudodeltidium is problematic since details, unfortunately, are not preserved in all genera (Wright and Rubel, Reference Wright and Rubel1996). The notothyrium in the Billingsellida varies from totally open to partially covered by chilidial plates to completely covered by the chilidium. The cladistic analysis performed by Topper et al. (Reference Topper, Harper and Brock2013) suggested that the combined presence of a well-developed pseudodeltidium and chilidial plates/chilidium are distinct features shared by most billingselloids and polytoechioids, while clitambonitoids are characterized by having a deltidium.
The spondylium has been described as a spoon-like elevated platform formed by dental plates that coalesce to a varying degree, supported by a median septum, accommodating the ventral muscle field (Williams et al., Reference Williams, Brunton, Carlson, Alvarez, Ansell and Baker2000). However, Vinn and Rubel (Reference Vinn and Rubel2000), studying this structure in several juvenile specimens of genera ascribed to the suborder Clitambonitidina, noted that the spondylium does not derive from the convergence of dental plates but develops from the free plate in early ontogenetic stages. In the clitambonitidine brachiopods, three kinds of spondylium are common: spondylium simplex, which is solely supported by the median ridge; spondylium triplex, which is freestanding and supported in early stages only by the median ridge and afterward additionally by auxiliary lateral ridges growing on both sides of the median ridge, on the undersurface of the spondylium that may or may not reach the valve floor; and pseudospondylium, a structure that mimics the spondylium but is developed instead by discrete, parallel dental plates that reach the valve floor early in the ontogeny, being the muscle field resting, between them, on the valve floor at that time. During the ontogeny, the muscle field is raised by shell thickening, forming a sessile spondylium. In some cases, this raised muscle field is undercut anterolaterally, in lesser or higher degree, becoming even free anteriorly and supported medially and laterally by dental plates simulating the spondylium triplex, as occurs in some genera assigned to the superfamily Polytoechioidea such as Polytoechia Hall and Clarke, Reference Hall and Clarke1892, Antigonambonites Öpik, Reference Öpik1934, and Raunites Öpik, Reference Öpik1939 (Wright and Rubel, Reference Wright and Rubel1996; Vinn and Rubel, Reference Vinn and Rubel2000; Topper et al., Reference Topper, Harper and Brock2013; Benedetto, Reference Benedetto2016). Wright and Rubel (Reference Wright and Rubel1996, p. 62) also stated that “the opposite is also true: a well-developed spondylium triplex, with its lateral septa reaching the valve floor, strongly simulates a pseudospondylium.” Finally, the spondylium simplex characterizes all clitambonitoids excepting the gonambonitids, which develop a spondylium triplex, whereas the presence of a pseudospondylium distinguishes the polytoechioids.
Thus, observations of the well-developed convex pseudodeltidium, the chilidial plates, and the pseudospondylium mimicking the spondylium triplex all suggest including the Peruvian material within the Polytoechioidea. Two families of polytoechioids are recognized: Polytoechiidae and Tritoechiidae. The former is characterized by having a free pseudospondylium, supported medially by a median ridge and laterally by dental plates resembling the spondylium triplex of gonambonitids (Vinn and Rubel, Reference Vinn and Rubel2000). However, the tritoechiids differ by displaying the ventral muscle field resting directly on the valve floor or raised by shell thickening on a pseudospondylium not undercut anteriorly (sessile). Thus, this material may be ascribed to the Polytoechiidae. Among the genera included in this family, the Peruvian taxon strongly resembles Antigonambonites by having a free-anteriorly pseudospondylium, with a small, transverse ridge separating posteriorly each socket in the dorsal valve, and radial grooves in the ventral interarea at both sides of the convex pseudodeltidium, which limit the external dental lamella with the interarea; it differs in lacking ventral and dorsal peripheral rims and a more raised pseudospondylium. Antigonambonites is considered in the Treatise (Williams et al., Reference Williams, Brunton, Carlson, Alvarez, Ansell and Baker2000) to be a gonambonitid, but since this genus has a pseudospondylium and not a spondylium triplex, as many authors suggested (Vinn and Rubel, Reference Vinn and Rubel2000; Popov et al., Reference Popov, Vinn and Nikitina2001; Vinn, Reference Vinn2001), it is now considered a polytoechiid (Topper et al., Reference Topper, Harper and Brock2013). In my opinion, another gonambonitid, Jaanussonites Neuman, Reference Neuman1976, also has a pseudospondylium, with the muscle field resting directly on the valve floor in early growth stages and becoming free and elevated anteriorly. In addition, this genus displays chilidial plates and radially arranged dorsal muscle scars, which are typical characters of the polytoechioids. This genus has not been included in the cladistic analysis of Topper et al. (Reference Topper, Harper and Brock2013), but it seems necessary to consider it a polytoechiid from now on. The Peruvian material strongly resembles Jaanussonites in its pseudospondylium with the lateral ridges/teeth growing toward the valve floor merging with two vascular ridges posteriorly, by having a convex pseudodeltidium, the same arrangement and shape of dorsal muscle field, and bisected sockets in the dorsal valve; it clearly differs from this genus by its multicostellate ornament without spines in the intercostal spaces and aditicules in the costellae and by lacking peripheral rims in both valves. The Peruvian taxon also strongly recalls Polytoechia but differs from it by its bigger size, ornamentation, more elongate outline, wider interarea, undifferentiated ventral adductor scars, and not deeply concave spondylium. Given all these differences, we propose a new genus, Enriquetoechia, to include the Peruvian material.
Enriquetoechia umachiriensis new species
Figure 3
Holotype
Internal and external mold of a ventral valve (CPI–9913) (Fig. 3.1–3.6) from the lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
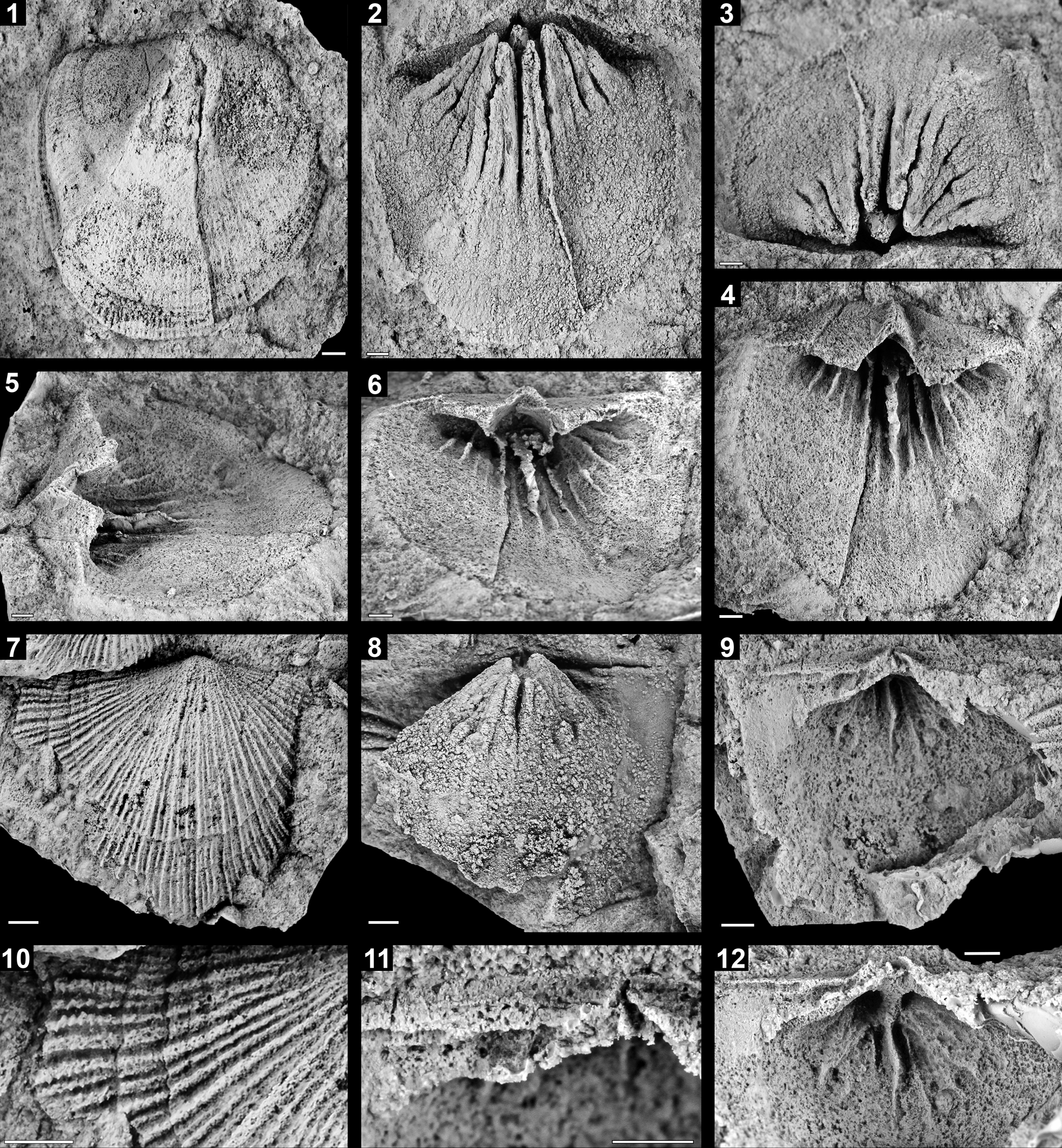
Figure 3. The polytoechioid brachiopod Enriquetoechia umachiriensis Colmenar n. gen. n. sp. from the Umachiri Formation at locality 2. (1–6) Holotype, external and internal molds of a ventral valve (CPI–9913); (1) latex cast of exterior, (2, 3) internal mold in (2) upper and (3) posterior views; (4–6) latex cast of interior in (4) ventral, (5) lateral oblique, and (6) anterior oblique views. (7–12) Paratype, external and internal molds of a dorsal valve (CPI–9923); (7) latex cast of exterior, (8) internal mold, (9) latex cast of interior, (10) detail of external ornamentation, (11) magnification of the cardinalia and dorsal interarea, (12) anterior oblique view of dorsal interior showing the adductor scars, median ridge, and median septum. Scale bars = 2 mm.
Paratypes
Internal and external mold of a dorsal valve (CPI–9923) (Fig. 3.7–3.12); internal mold of a ventral valve (CPI–9944); internal mold of a fragmented dorsal valve (CPI–9953); internal and external mold of a dorsal valve (CPI–9958).
Diagnosis
Polytoechioid of large size, biconvex, multicostellate with well-developed concentric irregular fila giving a zig-zag aspect to the ribs when eroded; pseudodeltidium convex; pseudospondylium formed very early in the ontogeny, elevated on the valve floor, supported posterolaterally by the two dental plates arising below the deltidiodont teeth, which grow toward the valve floor, fusing with two of the radially arranged ovarian vascular ridges and supported medially by a strong, high median ridge reaching its anterior edge and continuous anteriorly, decreasing in height until mid-valve length, where it disappears; ventral and dorsal peripheral rims absent; notothyrial platform small, restricted to the notothyrial cavity, elevated on the valve floor, strongly undercut in its frontal edge, continuous anteriorly in a median ridge; cardinal process ridge-like, swollen anteroventrally; chilidial plates leaving the notothyrium partially open medially; dorsal transmuscle septa and adductor scars radially arranged.
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Description
Shells of large size, up to 25 mm wide and 28 mm long; longitudinal profile equally biconvex; elongate in outline; cardinal extremities with obtuse angles; maximum shell width located slightly anteriorly of hinge line; rectimarginate anterior commissure.
Ventral valve about 110% as long as wide and 12% as deep as long; convex umbonal region, subplanar anteriorly. Ventral interarea slightly curved, apsacline, about 12% as long as valve length, with radial grooves marking the boundaries of the interarea with the dental lamella; deltiodont teeth strong, medially grooved; delthyrium covered by convex pseudodeltidium; pedicle foramen not visible. Dorsal valve about 85% as long as wide and 11% as deep as long; maximum depth near the umbo. Dorsal interarea planar, anacline, about 9% as long as valve length, with growth lines subparallel to hinge line; notothyrium partially covered by chilidial plates, diverging anteriorly, forming an angle of about 90°.
Radial ornament multicostellate with 12 ribs per 5 mm at 15 mm anteromedianly from umbo, all of them with rounded tops; costellae arise by intercalation at different growth stages; intercostal spaces as wide as ribs. Concentric ornament consisting of well-developed concentric irregular fila, when intersecting with the ribs generating a zig-zag pattern, more visible when the shell is abraded; strongly marked growth lines become more frequent anteriorly.
Ventral interior with a pseudospondylium formed very early in the ontogeny, as long as the interarea, elevated on the valve floor, supported posterolaterally by the dental plates, reaching the anterior edge of the spondylium, arising below the deltidiodont teeth, which grow toward the valve floor fusing with two of the radially arranged vascular ridges; spondylium also supported medially by a strong, high median ridge reaching its anterior edge and continuous anteriorly, decreasing in height until mid-valve length, where it disappears; ovarian vascular ridges radially arranged, evenly spaced, as long as median ridge, counting six at each side of this structure; subperipheral rim absent; mantle canal system pinnate; shell substance unknown.
Dorsal valve with short, anchor-shaped, highly elevated notothyrial platform, undercut in its frontal edge and restricted to the notothyrial cavity, it continues anteriorly into a high and thin median septum about 30% as long as valve; cardinal process ridge-like, swollen anteroventrally; dental sockets transversely elongated, bisected posteriorly by a transverse thin ridge originated in the interarea; sockets bounded anteriorly by strong socket ridges forming an angle of about 23° with hinge line; dorsal muscle field large, radially arranged, extending anteriorly slightly beyond median septum; adductor scars with rounded, strongly impressed anterior ends; peripheral rim absent; mantle canal system and shell substance unknown.
Etymology
After the Umachiri Inlier of the Peruvian Altiplano, where this species occurs.
Remarks
Enriquetoechia umachiriensis is the only species included so far within genus Enriquetoechia. See preceding remarks of Enriquetoechia n. gen.
Genus Altiplanotoechia new genus
Type species
Altiplanotoechia hodgini n. gen. n. sp. Colmenar in Colmenar and Hodgin, 2020, from the lower part of the Umachiri Formation, Floian–?Dapingian (Lower–Middle Ordovician).
Other species
Billingsella dice Walcott, Reference Walcott1905, from loose boulders at St. Albans, Vermont, USA, likely belonging to the Luke Hill Formation (Lower Ordovician) of the Philipsburg sequence; Antigonambonites sp. (in Neuman, Reference Neuman1976, pl. 4, fig. 24) from the Dapingian–lower Darriwilian Calcareous Tuff at Virgin Arm in the New World Island, Newfoundland; Jaanussonites? sp. (in Benedetto, Reference Benedetto2001, fig. 2V, W) from the upper Floian–lower Dapingian Aguada de la Perdiz Formation of the western Puna region of northwest Argentina.
Diagnosis
Ventribiconvex shells with hemipyramidal ventral valves and unisulcate commissure; ventral interior with sessile pseudospondylium during early stages, becoming slightly free, undercut anteriorly and supported laterally and medially by septa; radially arranged vascular septa usually present; dorsal interior with ridge-like cardinal process located in a small, triangular notothyrial platform, deeply undercut in its frontal edge, continuous anteriorly into a thin, long median ridge; dorsal adductor scars radially arranged.
Etymology
After the Altiplano (= high plains) morphotectonic region of Peru where the Umachiri Formation crops out.
Remarks
The presence of a pseudospondylium in the Peruvian material clearly relates it with the Polytoechioidea. Currently, two families are recognized within the Polytoechioidea: Tritoechiidae and Polytoechiidae (Popov et al., Reference Popov, Vinn and Nikitina2001, Reference Popov, Egerquist and Holmer2007; Topper et al., Reference Topper, Harper and Brock2013; Benedetto, Reference Benedetto2016). In the former, the entire ventral muscle field rests directly on the floor of the delthyrial cavity or is raised on a pseudospondylium, not undercut anteriorly. By contrast, in the Polytoechiidae, the pseudospondylium is free anteriorly and supported medially and laterally by septa resembling the spondylium triplex of gonambonitids (Vinn and Rubel, Reference Vinn and Rubel2000). The Peruvian material displays mixed characters between the Polytoechiidae and Tritoechiidae. The Peruvian taxon shows similar pseudospondylium development to the polytoechiid Pinatotoechia, which displays a sessile pseudospondylium until adult stages when it becomes slightly undercut and free anteriorly, but clearly it differs from that genus in its ornamentation consisting of hollow tubular spines developed on the ribs. It also resembles the genera Polytoechia Hall and Clarke, Reference Hall and Clarke1892 and Tritoechia Ulrich and Cooper, Reference Ulrich and Cooper1936. Hall and Clarke (Reference Hall and Clarke1892, p. 240) described the pseudospondylium of Polytoechia as “supported by a stout median septum, and two smaller lateral septa […], the former of these extends for the entire length of the plate, while the latter is free from the accessory septa near its anterior edge. The umbonal cavity of the valve is thus divided into five chambers […]”, which means that those three septa are discrete from early ontogenetic stages. Tritoechia, however, exhibits a sessile pseudospondylium that can be elevated onto the valve floor in higher or lesser extent by secondary shell thickening, leaving only three chambers in the umbonal cavity. Some gerontic specimens of few Tritoechia species, such as the type species Tritoechia typica (Schuchert and Cooper, Reference Schuchert and Cooper1932) and T. curvirostris Sando, Reference Sando1957, develop obscure ridges at the anterior margins of the ventral muscle field, which tend to elevate the muscular platform slightly above the floor of the valve (see Ulrich and Cooper, Reference Ulrich and Cooper1938, pl. 33B, fig. 19 and Sando, Reference Sando1957, pl. 14, fig. 3). The pseudospondylium of the Peruvian material is similar to the sessile spondylium of Tritoechia only in young stages, but it differs from Tritoechia since in the Peruvian specimens this structure becomes free, strongly undercut anteriorly, and supported laterally and medially by septa, in a similar way to Polytoechia, but only in adult stages. It also differs from Tritoechia in lacking radial lines in the interarea, which is another diagnostic character of that genus. Another character that differentiates this material from Polytoechia is the anterior commissure, unisulcate in the Peruvian specimens and uniplicate in Polytoechia. The Peruvian material also resembles Antigonambonites by having a sessile pseudospondylium becoming slightly raised anteriorly in gerontic specimens but differs from this genus in having a more convex ventral valve and a more raised pseudospondylium from early stages and in lacking a peripheral rim.
Material with pseudospondylium development similar to the material described here can be found in the literature, e.g., Tritoechia dice (Walcott, Reference Walcott1905) (see Ulrich and Cooper, Reference Ulrich and Cooper1938, pl. 31B) from the Floian Luke Hill Formation of Canada, Antigonambonites sp. (in Neuman, Reference Neuman1976, pl. 4, fig 24) from the Dapingian–lower Darriwilian Calcareous Tuff at Virgin Arm in Newfoundland, Jaanussonites? sp. (in Benedetto, Reference Benedetto2001, fig. 2V, W) from the upper Floian–lower Dapingian Aguada de la Perdiz Formation of the western Puna region of northwest Argentina, and Pomatotrema jiangsuense Wang and Xu (Reference Wang and Xu1966, pl. II, figs. 4–7) from the Floian–lower Darriwilian part of the Lunshan Formation at Tangshan, Nanjing. However, the latter clearly differ from the Peruvian material by having peripheral rims, which is a character only displayed by Pomatotrema within the Polytoechioidea. The mixed characters of the Chinese species suggest that it may belong to a new genus yet to be described. Dorsal valves attributable to this new genus display typical polytoechioid features such as chilidial plates, ridge-like cardinal process, small notothyrial platform continuous anteriorly in a median septum, widely divergent socket ridges, and radially arranged dorsal adductor scars. However, the dorsal valves studied here show a dorsal sulcus. This character is unique within the polytoechioids, given that the only folded genera in the superfamily are Polytoechia and Martellia Wirth, Reference Wirth1936, but both genera are uniplicate and not unisulcate as the Peruvian shells.
From the previous discussion, we propose here the establishment of a new genus, Altiplanotoechia, to embrace all the species with questionable generic determination mentioned in the preceding (except P. jiangsuense), with similar pseudospondylium development and without spinose ornamentation. The family assignment of the new genus is more complicated since it shares characteristics of both Polytoechiidae and Tritoechiidae, but we tentatively place it here within the Polytoechiidae until further investigations have been made in the suprageneric taxonomy of this superfamily.
Altiplanotoechia hodgini new species Figure 4.1–4.13
Holotype
Internal and external mold of a ventral valve (CPI–9929) (Fig. 4.6–4.10) from the lower part of Umachiri Formation at locality 2, Floian–?Dapingian (Lower–?Middle Ordovician).
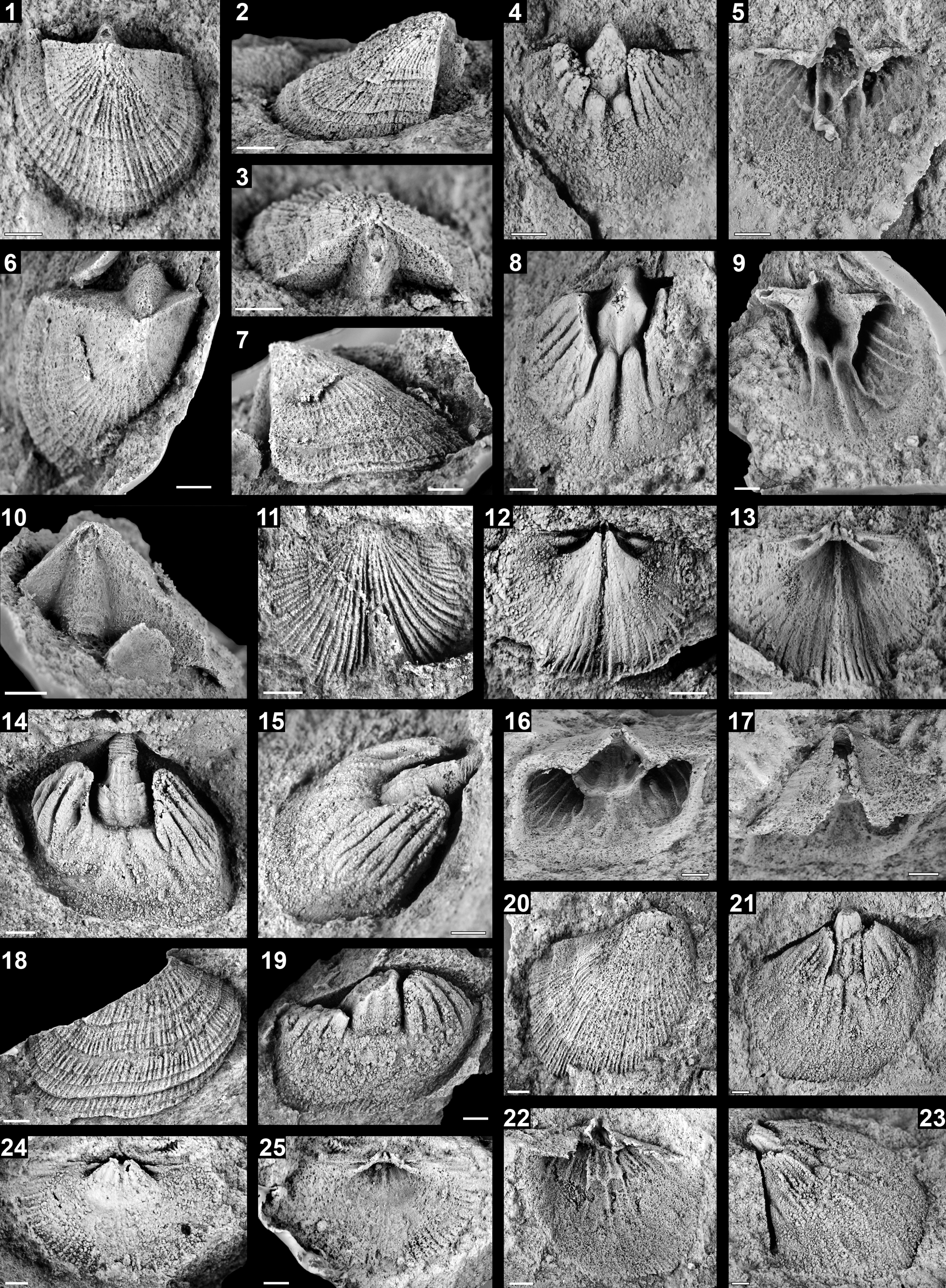
Figure 4. Polytoechioid brachiopods from the Umachiri Formation. (1–13) Altiplanotoechia hodgini n. gen. n. sp. Colmenar in Colmenar and Hodgin, 2020 (1–5) Paratype, external and internal molds of a ventral valve (CPI–9921); (1–3) latex cast of exterior in (1) upper, (2) lateral, and (3) posterior oblique views; (4) internal mold and (5) latex cast of interior. (6–10) Paratype, external and internal molds of a ventral valve (CPI–9929); (6, 7, 10) latex cast of exterior in (6) upper, (7) lateral, and (10) posterior views; (8) internal mold and (9) latex cast of interior. (11–13) Paratype, external and internal molds of a dorsal valve (CPI–9930); (11) latex cast of exterior, (12) internal mold and (13) latex cast of interior. (14–19) Pomatotrema laubacheri Colmenar n. sp. (14–17) Holotype, internal mold of a ventral valve (CPI–9916); (14, 15) internal mold in (14) upper and (15) lateral oblique views; (16, 17) latex cast of interior in (16) upper and (17) posterior oblique views; (18, 19) paratype, external and internal molds of a ventral valve (CPI–9914); (18) latex cast of exterior and (19) internal mold. (20–23) Tritoechia (Tritoechia) sp. (20–23) External and internal molds of a ventral valve (CPI–9915); (20) latex cast of exterior, (21–23) internal mold in (21) upper and (23) lateral oblique views, (22) latex cast of interior. (24, 25) Polytoechioidea indet., dorsal valve (CPI–9932); (24) internal mold and (25) latex cast of interior. Scale bars = 2 mm.
Paratypes
Internal and external mold of a ventral valve (CPI–9921) (Fig. 4.1–4.5); internal and external mold of a dorsal valve (CPI–9930) (Fig. 4.11–4.13).
Diagnosis
Altiplanotoechia species with hemipyramidal ventral valve, slightly unisulcate commissure and small notothyrial platform, extremely undercut in its frontal edge, highly elevated on the valve floor, and continuous anteriorly in a long and thin median ridge.
Occurrence
Lower part of Umachiri Formation at locality 2, Floian–?Dapingian (Lower–?Middle Ordovician).
Description
Shells of medium size up to 18 mm wide and 14 mm long; longitudinal profile ventribiconvex; subcircular in outline; cardinal extremities acute in early growth stages, right to obtuse in adult stages; maximum shell width approximately at mid-valve length; slightly unisulcate anterior commissure.
Ventral valve hemipyramidal, about 104% as long as wide and 50% as deep as long; maximum convexity at umbonal region. Ventral interarea slightly curved, apsacline, about 52% as long as valve length, with strong, triangular deltiodont teeth; delthyrium covered by strongly convex pseudodeltidium apically perforated by a large, ovate pedicle foramen.
Dorsal valve about 92% as long as wide and 22% as deep as long; maximum depth near the umbo; dorsal interarea planar, anacline, about 8% as long as valve length; notothyrium almost totally covered by chilidial plates, which nearly touch the posterior part of the cardinal process, diverging anteriorly forming an angle of about 99°.
Radial ornament multicostellate with 18 ribs per 5 mm at anterior commissure, all of them with rounded tops; costellae arise by branching at different growth stages; intercostal spaces narrower than ribs. Concentric ornament consisting of weakly marked fila and strong growth lines becoming more frequent anteriorly; in abraded parts of the shell, the intersection of costellae and fila is more visible (Fig 4.11).
Ventral interior with strong dental plates arising from the teeth and growing toward valve floor forming a pseudospondylium elevated on the valve floor by shell thickening, sessile during early stages, becoming free, undercut anteriorly and supported laterally and medially by septa in adult stages, medium septum extending anteriorly almost until the commissure; ventral muscle field elongate, with diductor scars separated from adductor scars by strong, subparallel ridges, adductor scars deeply impressed in the pseudospondylium; ovarian vascular ridges radially arranged, shorter than median ridge; mantle canal system pinnate; shell substance unknown.
Dorsal interior with ridge-like cardinal process, slightly thickened anteriorly, located on a small notothyrial platform, extremely undercut in its frontal edge, highly elevated on the valve floor, and continuous anteriorly in a long and thin median ridge reaching less than half of the valve length; divergent socket ridges forming an angle of about 28° with hinge line; sockets big, triangular in outline, fairly deep; adductor scars radially arranged, poorly impressed; shell substance and mantle canal system unknown.
Etymology
After Eben Blake Hodgin (Harvard University), who found the Umachiri Formation fossil localities.
Remarks
Some of the taxa included within this new genus are in open nomenclature; however, they will be included here for comparison with the Peruvian species. Altiplanotoechia hodgini is here proposed as type species since the type material of Tritoechia dice (Walcott, Reference Walcott1905) comes from loose boulders of uncertain origin within the Philipsburg sequence of St. Albans, Vermont, northeastern USA. A. hodgini differs from Tritoechia dice by having a more subcircular outline, slightly unisulcate anterior commissure, a more convex ventral valve, deeper impressed adductor scars, and longer ventral median and vascular septa. Antigonambonites sp. (in Neuman, Reference Neuman1976) differs from the new species in having a much shorter ventral median septum and a more convex dorsal valve with dorsal muscle scars strongly impressed anteriorly. Only one poorly preserved ventral interior of Jaanussonites? sp. (in Benedetto, Reference Benedetto2001, fig. 2V, W) is known. It displays internal characters very similar to those in A. hodgini, e.g., the deeply impressed ventral adductor scars and length of median and vascular septa. Additional and better-preserved material of the Argentinian taxon could perhaps confirm its synonymy with A. hodgini.
Family Tritoechiidae Ulrich and Cooper, Reference Ulrich and Cooper1936
Genus Pomatotrema Ulrich and Cooper in Schuchert and Cooper, Reference Schuchert and Cooper1932
Type species
Pomatotrema murale Ulrich and Cooper in Schuchert and Cooper, Reference Schuchert and Cooper1932 from the West Spring Creek Formation, Arbuckle Group, Oklahoma, Dapingian–lower Darriwilian (Middle Ordovician).
Other species
Pomatotrema semiconvexum (Poulsen, Reference Poulsen1927) (Nunatami Formation, northwest Greenland, Dapingian–lower Darriwilian), P. magnum Ulrich and Cooper, Reference Ulrich and Cooper1938 (Arbuckle Limestone, Oklahoma, Dapingian–lower Darriwilian), P. oklahomense Ulrich and Cooper, Reference Ulrich and Cooper1936 (West Spring Creek Formation, Oklahoma, Dapingian–lower Darriwilian), P. transversum Ulrich and Cooper, Reference Ulrich and Cooper1936 (West Spring Creek Formation, Oklahoma, Dapingian–lower Darriwilian), P. inconspicuum Williams and Curry, Reference Williams and Curry1985 (Tourmakeady Limestone, Ireland, Dapingian–lower Darriwilian), Pomatotrema sp. (upper Fenxiang Formation., China, Tremadocian–lower Floian, Lower Ordovician).
Pomatotrema laubacheri new species
Figure 4.14–4.19
Holotype
Internal mold of a ventral valve (CPI–9916) (Fig. 4.14–4.17) from the lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Diagnosis
Pomatotrema species with hemipyramidal ventral valve, extremely high interarea, and ventral muscle scars raised on the valve floor by shell thickening.
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Description
Hemipyramidal ventral valves of medium size, up to 15 mm wide and 13 mm long; transversely oval in outline; cardinal extremities rectangular; maximum shell width located at hinge line; rectimarginate anterior commissure.
Ventral valve about 86% as long as wide and 50% as deep as long; maximum convexity at umbonal region. Ventral interarea slightly curved, apsacline, about 50% as long as valve length, with horizontal, subparallel growth lines; deltiodont teeth strong; delthyrium covered by convex pseudodeltidium, longer than wide, apically perforated by a big, oval pedicle foramen.
Radial ornament multicostellate with 14 ribs per 5 mm at anterior commissure, all of them with rounded tops; costella arise by branching at different growth stages; intercostal spaces narrower than ribs. Concentric ornament consisting of strongly marked growth lines becoming more frequent and lamellose anteriorly.
Ventral interior with strong dental plates, slightly convergent toward valve floor forming a pseudospondylium and bounding laterally the muscle field that is resting on the thickened valve floor; ventral muscle field strongly impressed, diductor scars about 30% shorter than adductor scars, diductor scars elongated, subparallel, undercutting the pseudospondylium posteriorly, adductor scars lanceolate, separated from each other by a narrow, weak ridge and from the diductor scars by strong ridges; strongly marked pedicle callist present in the posterior part of the delthyrial cavity; radially arranged ovarian vascular ridges present posterolaterally; mantle canal system pinnate.
Etymology
After Gérard Laubacher, French geologist, who devised the first stratigraphic scheme for the Altiplano and the first Ordovician fossil findings in the region.
Remarks
The curved pseudodeltidium, apically perforated, and the sessile pseudospondylium, formed by the dental plates extending toward the valve floor, justify the inclusion of this material within the family Tritoechiidae. The presence of a peripheral rim and the absence of radial lines in the interarea are diagnostic characters of Pomatotrema Ulrich and Cooper in Schuchert and Cooper, Reference Schuchert and Cooper1932. Only two ventral internal molds and an associated external mold are available. However, the ventral muscle field raised on the valve floor by shell thickening allows differentiation from all species included in this genus. Among all described species, P. laubacheri is morphologically closest to the genotype P. murale Ulrich and Cooper in Schuchert and Cooper, Reference Schuchert and Cooper1932 by showing strong radial ovarian vascular ridges and strong dental plates, converging toward the valve floor, bounding laterally the ventral muscle field, but P. laubacheri clearly differs from the type species in having a more convex ventral valve and higher ventral interarea and lacking the median ridge in the ventral valve.
Genus Tritoechia Ulrich and Cooper, Reference Ulrich and Cooper1936
Subgenus Tritoechia (Tritoechia) Ulrich and Cooper, Reference Ulrich and Cooper1936
Type species
Deltatreta typica Schuchert and Cooper, Reference Schuchert and Cooper1932 from the Arbuckle Limestone of Oklahoma, USA, Dapingian–lower Darriwilian (Middle Ordovician).
Tritoechia (Tritoechia) sp.
Figure 4.20–4.23
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Description
Ventral valve of medium size about 20 mm wide and 19 mm long; subquadrate in outline; cardinal extremities rounded with obtuse angles; maximum shell width slightly anterior to hinge line; anterior commissure rectimarginate.
Ventral valve hemipyramidal, about 95% as long as wide and 60% as deep as long; maximum depth near the umbo; ventral interarea planar, apsacline; delthyrium covered by a convex pseudodeltidium.
Radial ornament multicostellate with 11 ribs per 5 mm at 10 mm anteromedianly, all of them with rounded tops; costellae arise by branching at different growth stages; intercostal spaces as wide as ribs.
Ventral interior with thin dental plates, almost subparallel, slightly convergent toward valve floor forming a sessile pseudospondylium and bounding laterally the muscle field, which is resting on the thickened valve floor; dental plates extending anteriorly beyond muscle field scars as low, narrow ridges; ventral muscle field elongated, about twice as long as wide, muscle scars strongly impressed, each diductor as wide as both adductor scars altogether, diductor scars separated from the adductors by strong, subparallel ridges, low and thin median ridge growing anteriorly from the adductor scars until approximately two-thirds of valve length; radially arranged ovarian vascular ridges present posterolaterally; mantle canal system pinnate.
Remarks
The curved pseudodeltidium and the sessile pseudospondylium formed by the dental plates extending toward the valve floor justify the inclusion of this material within the family Tritoechiidae. The extremely convex ventral valve, the non-undercut anteriorly ventral muscle field, and the median ridge extending in front of ventral muscle field are characters typically displayed by Tritoechia. More than 40 species have been included in this genus. Among the species with hemipyramidal ventral valve, the Peruvian species resembles T. delicatula Ulrich and Cooper, Reference Ulrich and Cooper1936 from the Tremadocian Kindblade Formation of Oklahoma in having narrow, almost subparallel ventral muscle scars with subparallel ridges separating diductor from adductor scars. However, the Peruvian species differs from T. delicatula in being almost double the size of the largest specimen attributed to that species and by having a much narrower median ridge. The Peruvian species also resembles the type species T. typica (Schuchert and Cooper, Reference Schuchert and Cooper1932) from the same formation in having the dental plates extending anteriorly beyond the muscle field as low ridges. However, the Peruvian material clearly differs from T. typica in lacking the aditicules/hollow costellae characteristic of the type species and by having a subparallel muscle field, anteriorly widely divergent in T. typica. The dental plates extending anteriorly beyond the elongated muscle field as low ridges is a character also shared with Tritoechia sp. from the upper member of the Suri Formation from the Famatina Range of northwest Argentina (Benedetto, Reference Benedetto2003), but the Peruvian species differs from the Argentinian species in having a slightly thicker ornamentation and having ridges separating diductor from adductor scars. Despite showing differences from all published Tritoechia species, it is preferable not to define a new species until additional and more abundant material of this taxon is available.
Subgenus Tritoechia (Parvitritoechia) Benedetto, Reference Benedetto2003
Type species
Tritoechia (Parvitritoechia) preandina Benedetto, Reference Benedetto2003 (in Benedetto et al., Reference Benedetto2003) from the lower part of the San Juan Formation, Precordillera, Argentina, upper Tremadocian–lower Floian (Lower Ordovician).
Tritoechia (Parvitritoechia) sp.
Figure 5.1–5.4
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).

Figure 5. Polytoechioid, orthoid, and porambonitoid brachiopods from Umachiri Formation. (1–4) Tritoechia (Parvitritoechia) sp., external and internal molds of a ventral valve (CPI–9920); (1) latex cast of exterior, (2) internal mold, (3) latex cast of exterior and (4) magnification of the cardinalia. (5–10) Paralenorthis cf. P. carlottoi. (5) Latex cast of exterior of a ventral valve (CPI–9917); (6, 7) ventral valve (CPI–9927), (6) internal mold and (7) latex cast of interior; (8) latex cast of exterior of a dorsal valve (CPI–9922); (9, 10) dorsal valve (CPI–9928), (9) internal mold, and (10) latex cast of interior. (11–13) Panderina? sp. (11) Latex cast of exterior of a dorsal valve (CPI–9926); (12, 13) ventral valve (CPI–9911); (12) internal mold and (13) latex cast of interior. (14–19) Mollesella cf. M. planidorsalis. (14, 15) Internal mold of a fragmented ventral valve (CPI–9925); (14) internal mold and (15) latex cast of interior; (16–19) dorsal valve (CPI–9924); (16) latex cast of exterior, (17) internal mold, (18) latex cast of interior, and (19) magnification of the cardinalia. (20–23) Rugostrophia cf. R. latireticulata. (20) Latex cast of exterior dorsal valve (CPI–9918); (21–23) dorsal valve (CPI–9912); (21) internal mold; (22, 23) latex cast of interior in (22) dorsal and (23) anterior oblique views. Scale bars = 2 mm.
Description
Dorsal valve of medium size up to 17 mm wide and 10 mm long; transversely oval in outline; cardinal extremities rounded with obtuse angles; maximum shell width located slightly before mid-valve length; slightly unisulcate anterior commissure.
Dorsal valve about 60% as long as wide and 11% as deep as long; maximum depth near the umbo; dorsal interarea planar, anacline, about 11% as long as valve length; notothyrium partially covered by chilidial plates, diverging anteriorly forming an angle of about 93°.
Radial ornament unequally parvicostellate with up to 12 prominent costae and up to six costellae arising by intercalation; some of the costellae become more marked near the anterior margin. Both costae and costellae with rounded tops. The only dorsal exterior available shows evidence of shell damage reflected as discontinuities in shell ornament (Fig. 5.1).
Dorsal interior with ridge-like cardinal process, located in a small notothyrial platform, undercut in its frontal edge, elevated on the valve floor, and continuous anteriorly in a short and thin median ridge restricted to notothyrial cavity; widely divergent socket ridges forming an angle of about 13° with hinge line; sockets triangular in outline, fairly shallow; dorsal muscle field not impressed; shell substance and mantle canal system unknowns.
Remarks
This material is assigned to Polytoechioidea on the basis of its typical cardinalia. The only genera in this superfamily with unequally parvicostellate ornament are Acanthotoechia Williams and Curry, Reference Williams and Curry1985, Raunites Öpik, Reference Öpik1939, and Tritoechia (Parvitritoechia) Benedetto, Reference Benedetto2003. The Peruvian material clearly differs from Acanthotoechia by not having rows of spines peripherally and by lacking peripheral rims in the interior. The Peruvian material differs from Raunites in lacking concentric fila and in having thicker ribs, rounded cardinal angles, better-marked and fewer costae over the disc, smaller, more elevated notothyrial platform, and thinner median ridge. Benedetto (in Benedetto et al., Reference Benedetto, Cech and Esbry2003) described Tritoechia (Parvitritoechia) to encompass all Tritoechia species with clearly unequal parvicostellate ornament. The Peruvian material perfectly fits within the diagnosis of this subgenus, so we propose to include our material within Tritoechia (Parvitritoechia). This material differs from the type species T. (Parvitritoechia) preandina from the lower part of the San Juan Formation of the Precordillera in having rounded cardinal extremities, a smaller size, a non-thickened anteriorly cardinal process, and a thinner median ridge. It differs from T. (P.) pyramidalis (Bates, Reference Bates1968) from the Treiorwerth Formation of Anglesey in having much thinner ribs and lacking growth lines. The Peruvian material is morphologically closer to the material assigned to this genus from the volcaniclastic rocks of the Aguada de la Perdiz Formation of the Argentinian western Puna, Tritoechia sp. A and Tritoechia sp. B (Benedetto, Reference Benedetto2001), later reassigned to T. (Parvitritoechia) by Benedetto (Reference Benedetto2003). The material from the western Puna also displays rounded cardinal extremities, a similar number of costae, and costellae between them, but differs from the Peruvian material in having a low median septum dividing the dorsal muscle field. Even though the material from the Umachiri Formation shows differences with all the known species of T. (Parvitritoechia), it is too scarce and fragmentary to define a new species.
Polytoechioidea indet.
Figure 4.24, 4.25
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Remarks
The dorsal valve available can clearly be ascribed to the Polytoechioidea on the basis of its cardinalia. However, it does not seem to correspond to any of the taxa belonging to that superfamily described in this paper in either external or internal features. Pomatotrema laubacheri and Tritoechia sp. lack dorsal valves in the collection, so this dorsal valve could potentially belong to any of those taxa. However, the absence of a peripheral rim precludes its attribution to Pomatotrema, and the subcircular outline does not match Tritoechia species described in the preceding. So it is plausible to think that other polytoechioids might be present in the Umachiri associations.
Order Orthida Schuchert and Cooper, Reference Schuchert and Cooper1932
Suborder Orthidina Schuchert and Cooper, Reference Schuchert and Cooper1932
Superfamily Orthoidea Woodward, Reference Woodward1852
Family Orthidae Woodward, Reference Woodward1852
Genus Paralenorthis Havlíček and Branisa, Reference Havlíček and Branisa1980
Type species
Paralenorthis immitatrix Havlíček and Branisa, Reference Havlíček and Branisa1980; Floian (Lower Ordovician) siltstones at San Lucas, Bolivia.
Paralenorthis cf. P. carlottoi Villas in Gutiérrez-Marco and Villas, Reference Gutiérrez-Marco and Villas2007
Figure 5.5–5.10
cf. Reference Gutiérrez-Marco and Villas2007 Paraleonorthis carlottoi Villas in Gutiérrez-Marco and Villas, p. 555, fig. 6A–6M2.
Holotype
External mold of a ventral valve (MGM 5973X−1) from the Horizon A of the San José Formation, Cordillera Oriental, Peru; upper Floian (uppermost Lower Ordovician).
Diagnosis
Paralenorthis species with rectangular cardinal angles, costate and capillate, with 18–20 costae in adult dorsal valves, only two median costae occupying median sulcus, ventral muscle field 26%–33% as long as valve, notothyrial platform restricted to notothyrial cavity and 14%–22% as long as valve, blade-like divergent brachiophores, and low median ridge.
Occurrence
Lower part of the Umachiri Formation at localities 1, 2, and 4, Floian–?Dapingian (Lower–Middle Ordovician).
Materials
Thirty-one specimens; figured: CPI–9917, 9922, 9927, 9928 (Fig. 5.5–5.10); non-figured: CPI–9931, 9933–9943, 9945–9952, 9954–9957, 9959–9961.
Remarks
The material from the Umachiri Formation strongly resembles, both externally and internally, Paralenorthis carlottoi Villas (in Gutiérrez-Marco and Villas, Reference Gutiérrez-Marco and Villas2007) from the middle–upper Floian San José Formation of the Peruvian Cordillera Oriental. The material studied here commonly displays rectangular cardinal angles, about 18 costae in adult dorsal valves, with only two median costae in the median sulcus, a ventral muscle field about 32% as long as valve, a notothyrial platform about 18% as long as valve, continuous anteriorly in a low median ridge. These values are within the variability range of the species P. carlottoi. However, this material displays differences in the dorsal muscle field, showing wider posterior adductor scars than the species described by Villas (in Gutiérrez-Marco and Villas, Reference Gutiérrez-Marco and Villas2007). Given this difference displayed by the population of the Umachiri Formation, it is assigned here only tentatively to this species.
Family Hesperonomiidae Ulrich and Cooper, Reference Ulrich and Cooper1936
Genus Mollesella Benedetto, Reference Benedetto2003
Type species
Mollesella planiventralis Benedetto, Reference Benedetto2003 from the uppermost Floian–lower Dapingian Molles Formation, Famatina, northwest Argentina.
Mollesella cf. M. planidorsalis Benedetto, Reference Benedetto2003
Figure 5.14–5.19
cf. Reference Benedetto2003 Mollesella planidorsalis Benedetto, p. 233, fig. 11.13–11.28.
Holotype
Internal mold of a ventral valve (CEGH-UNC 19649) from the Molles Formation, Famatina Range, Argentina; middle Floian-Dapingian (Lower–Middle Ordovician).
Diagnosis
Medium-sized to large shells with planar ventral valve and moderately convex sulcate dorsal valve. Ornamentation finely multicostellate. Ventral valve with triangular muscle field and proximal half of vascula media subparallel. Notothyrial platform strongly raised, continued anteriorly as high, rounded median ridge bearing simple, anteriorly thickened cardinal process, which becomes bulbous and prominent in full-grown specimens.
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Materials
One dorsal interior and exterior (CPI–9924) (Fig. 5.16–5.19) and one fragmentary ventral interior (CPI–9925) (Fig. 5.14, 5.15).
Remarks
The finely costellate ornament, the open delthyrium, the triangular and relatively short ventral muscle field impressed directly on the valve floor, the short dorsal interarea, the simple cardinal process, and the quadripartite dorsal muscle field allow assigning this material to the family Hesperonomiidae. The dental plates laterally and anterolaterally bounding the ventral muscle field, the cardinal process becoming bulbous anteriorly, the absence of chilidial plates, the relatively raised notothyrial platform, and the posterolateral, radially arranged ovarian vascular ridges warrant its inclusion within the monospecific genus Mollesella Benedetto, Reference Benedetto2003 from the Famatina Range of northwest Argentina. The Peruvian material shows some differences from M. planiventralis, such as having wider notothyrial platform and median ridge. Since these features seem to vary within the Argentinian species and given that the Peruvian material is fragmentary and poorly preserved, we decided to assign it tentatively to that species.
Family Productorthidae Schuchert and Cooper, Reference Schuchert and Cooper1931
Subfamily Productorthinae Schuchert and Cooper, Reference Schuchert and Cooper1931
Genus Panderina Schuchert and Cooper, Reference Schuchert and Cooper1931
Type species
Productus abscissus Pander, Reference Pander1830 from the Leetse Formation of northwest Russia, Dapingian (Middle Ordovician).
Panderina? sp.
Figure 5.11–5.13
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Materials
One ventral interior (CPI–9911) (Fig. 5.12–5.13) and one dorsal exterior (CPI–9926) (Fig. 5.11).
Remarks
Short and thick dental plates continued as a ridge around the front margin of the oval, small, ventral muscle field, forming a pseudospondylium, closely resembling the genus Panderina Schuchert and Cooper, Reference Schuchert and Cooper1931. However, in this genus the pseudospondylium usually continues anteriorly into a median ridge, absent in the Peruvian material. Benedetto (Reference Benedetto2003) described a new species, Panderina? ambigua from the Argentinian Famatina Range with similar features in the ventral valve. He also noted that this species showed mixed characters from other productorthid genera, making the generic classification problematic. Mixed characters include the strongly lamellose ornament in the whole shell surface, shared with the material described here. We agree with Benedetto (Reference Benedetto2003) that the generic assignment of the Argentinian species is doubtful, and we consider our material congeneric. Further investigation must be done to clarify the generic classification of the Argentinian and Peruvian taxa. Due to small differences in the ventral muscle field outline and the lack of dorsal interiors in the Peruvian collection, it is preferable to leave this material in open nomenclature.
Order Pentamerida Schuchert and Cooper, Reference Schuchert and Cooper1931
Suborder Syntrophiidina Ulrich and Cooper, Reference Ulrich and Cooper1936
Superfamily Porambonitoidea Davidson, Reference Davidson1853
Family Tetralobulidae Ulrich and Cooper, Reference Ulrich and Cooper1936
Subfamily Punctolirinae Andreeva, Reference Andreeva1982
Genus Rugostrophia Neuman, Reference Neuman and Dutro1971
Type species
Rugostrophia sylvestris Neuman, Reference Neuman and Dutro1971 from the Lower Birch Island Member of the Turnbull Mountain Formation. (Dapingian–lower Darriwilian) at Newfoundland, Canada.
Rugostrophia cf. R. latireticulata Neuman, Reference Neuman1976
Figure 5.20–5.23
cf. Reference Neuman1976 Rugostrophia latireticulata Neuman, p. 41, pl. 7, figs. 1–8.
Holotype
Dorsal valve, internal mold, and counterpart external mold (GSC 35085) from the ‘calcareous tuff’ at Virgin Arm, New World Island, Newfoundland; lower Dapingian–lower Darriwilian (Middle Ordovician).
Diagnosis
Large, transversely elliptical Rugostrophia having a coarse reticulate ornament, the costellae more widely spaced than the concentric laminae.
Occurrence
Lower part of the Umachiri Formation at locality 2, Floian–?Dapingian (Lower–Middle Ordovician).
Materials
One dorsal external (CPI–9918) (Fig. 5.20) and internal (CPI–9912) (Fig. 5.21–5.23) molds.
Remarks
The only ventral interior available is identical to R. latireticulata Neuman, Reference Neuman1976 (pl. 7, fig.7) from the Dapingian–lower Darriwilian of New World Island. Neuman (Reference Neuman1976) stated that differences between R. sylvestris and R. latireticulata lie essentially in the ornamentation. The Peruvian material differs from Rugostrophia sp. from the Dapingian–lower Darriwilian Treiorwerth Formation (Neuman and Bates, Reference Neuman and Bates1978, pl. 67, figs. 15–18) in displaying flattened posterolateral extremities. In addition, it clearly differs from the Argentinian Rugostrophia species, R. protoandina Benedetto, Reference Benedetto2003 and Rugostrophia sp. in having a much deeper notothyrial cavity and strongly raised anterior ends of adductor scars. Since no exteriors are available in the Peruvian collection that enable differentiating between the Canadian species, it is preferable to assign this material tentatively to R. latireticulata.
Paleobiogeographical remarks
Most of the brachiopod taxa occurring in the Umachiri Formation belong to the superfamilies Orthoidea and Polytoechioidea with the exception of a porambonitoid. Among the orthoids, the most abundant genus is Paralenorthis. Even though it is a cosmopolitan genus, the species occurring in the Umachiri Formation strongly resembles Paralenorthis carlottoi Villas (in Gutiérrez-Marco and Villas, Reference Gutiérrez-Marco and Villas2007) from the middle–upper Floian rocks of the San José Formation in the Carcel Puncco Canyon (Inambari River) section of the Peruvian Cordillera Oriental. Another orthoid brachiopod at Umachiri is very similar to the type species of Mollesella Benedetto, Reference Benedetto2003, previously known only in the uppermost Floian–lower Dapingian rocks of the Molles Formation from the Famatina Range of northwest Argentina (Benedetto, Reference Benedetto2003). The similarity of the Peruvian assemblage with those of the Famatina Range is also strengthened by the co-occurrence of specimens referable to the brachiopod genus Panderina. Although this genus is typical from Baltica and China, Benedetto (Reference Benedetto2003) described a new species from the upper volcaniclastic member (Loma del Kilómetro Mb) of the Suri Formation that he tentatively ascribed to Panderina. Panderina? ambigua Benedetto, Reference Benedetto2003 shows mixed characteristics from other productorthid genera, making the generic classification problematic. The material from the Umachiri Formation shows some of the same mixed characteristics, suggesting that the species are strongly related. All these occurrences highlight the resemblance of the Peruvian assemblage to those of the Argentinian Famatina Range.
The brachiopod-dominated assemblage described here contains numerous polytoechioids. One of them, Enriquetoechia, is endemic to the Peruvian Altiplano. The other new genus described here, Altiplanotoechia, is likely present in the Argentinian western Puna, the Ganderia and Monian composite (including Anglesey) terranes (both likely located west of Avalonia forming an island arc at intermediate latitudes between Gondwana and Laurentia), and the Dashwoods (New World Island) and Moretown (St. Albans, Vermont) terranes (both likely located near Laurentia) (see Domeier, Reference Domeier2016 and Liljeroth et al., Reference Liljeroth, Harper, Carlisle and Nielsen2017, for estimated position of these island arcs during the Cambrian–Ordovician times). Pomatotrema is a genus previously known only in the Laurentian realm and South China. Benedetto (Reference Benedetto1987) described two species of this genus from material of the San Juan Formation of the Precordillera; however, they were subsequently reassigned to Martellia (Popov et al., Reference Popov, Vinn and Nikitina2001). The occurrence of Pomatotrema in the Peruvian Altiplano represents the highest paleolatitudinal occurrence of this genus so far and indicates connections of this region with the Laurentian realm and South China during Floian times. The links with Laurentia, the peri-Laurentian terranes, and other low-latitude terranes during the Floian–Dapingian are strengthened by the presence of Tritoechia (Tritoechia), with several species described from localities of the Dashwoods Terrane, Laurentia, Siberia, Kazakhstan, and Tasmania (Schuchert and Cooper, Reference Schuchert and Cooper1932; Ulrich and Cooper, Reference Ulrich and Cooper1936, Reference Ulrich and Cooper1938; Sando, Reference Sando1957; Neuman, Reference Neuman1976; Laurie, Reference Laurie1980; Popov et al., Reference Popov, Vinn and Nikitina2001; Hansen and Harper, Reference Hansen and Harper2003). Species of Tritoechia (Tritoechia) have also been described from the Argentinian Precordillera, western Puna, and Famatina regions, as well as in the Ganderia Terrane. The subgenus Tritoechia (Parvitritoechia) is known from the Dapingian–lower Darriwilian assemblages of the Treiorwerth Formation of the Midland Valley–Mayo Terrane (Williams and Curry, Reference Williams and Curry1985), located in peri-Laurentian settings, from the San Juan Formation of the Precordillera (Benedetto, Reference Benedetto2003), and from the Aguada de la Perdiz Formation of the Argentinian western Puna (Benedetto, Reference Benedetto2001). The Peruvian material assigned to Tritoechia (Parvitritoechia) seems morphologically related to the species described from the upper Floian–lower Dapingian Aguada de la Perdiz Formation of the Argentinian western Puna.
The productorthid present in the Umachiri Formation, assigned here to the brachiopod genus Rugostrophia, strongly resembles Rugostrophia latireticulata Neuman, Reference Neuman1976 from the ‘calcareous tuff’ at Virgin Arm, New World Island, Newfoundland. This genus normally occurs in localities belonging to the Ganderia (e.g., Maine, New Brunswick) and Monian composite (Anglesey) terranes, located at intermediate latitudes between Gondwana and Laurentia, and on the Dashwoods Terrane (New World Island), located close to Laurentia, during early Dapingian–early Darriwilian times (Neuman, Reference Neuman1976; Neuman and Bates, Reference Neuman and Bates1978). However, species assigned to this genus also occur in the substantially older (uppermost Floian–lower Dapingian) volcaniclastic rocks of the Aguada de la Perdiz Formation of the Argentinian western Puna (Benedetto, Reference Benedetto2001, Reference Benedetto2003) as well as in the Darriwilian part of the San Juan Formation of the Precordillera (Herrera and Benedetto, Reference Herrera, Benedetto, Mackinnon, Lee and Campbell1991).
All these occurrences highlight the strong biogeographic affinity of the Peruvian Altiplano during the Early–?Middle Ordovician with the Famatina and western Puna regions of northwest Argentina. The brachiopod fauna of the Peruvian Altiplano could be integrated, together with their homologues from Famatina and western Puna, into a well-differentiated biogeographical subprovince (in the sense of Servais et al., Reference Servais, Cecca, Harper, Isozaki, Niocaill, Harper and Servais2013) during the Early–Middle Ordovician within the South American margin of Gondwana.
As discussed in the preceding, the Peruvian assemblage studied here also shows clear links with several terranes, of probable Gondwanan origin (Murphy et al., Reference Murphy, Keppie, Dostal, Nance, Ramos and Keppie1999; Zagorevski et al., Reference Zagorevski, McNicoll, Rogers and van Hees2015; Reusch et al., Reference Reusch, Holm-Denoma and Slack2018), located at intermediate (Ganderia and Monian composite terranes) and low (Moretown, Dashwoods, and Midland Valley–Mayo terranes) latitudes between Gondwana and Laurentia (see Johnson et al., Reference Johnson, Van Der Pluijm and Van der Voo1991 and Cutts et al., Reference Cutts, Zagorevski, McNicoll and Carr2011 for paleogeographic position of Dashwoods Terrane) during Floian-Dapingian times. Villas et al. (Reference Villas, Colmenar and Gutiérrez-Marco2015) proposed the existence during Sandbian times (early Late Ordovician) of intermediate islands in the Rheic Ocean between the Peruvian Altiplano and Avalonia, permitting dispersal following the Southern Westerlies currents (Pohl et al., Reference Pohl, Nardin, Vandenbroucke and Donnadieu2016) and island hopping of brachiopod species with low dispersal potential (short larval period) to overcome the presumably large distances between those terranes. Several of these islands/terranes (known as Ganderian terranes) likely detached from Gondwana and drifted toward Laurentia and Baltica during the late Cambrian–Early Ordovician, triggering the opening of the Rheic Ocean (van Staal et al., Reference van Staal, Sullivan and Whalen1996; Ayuso and Schulz, Reference Ayuso, Schulz, Goodfellow, McCutcheon and Peter2003; Ayuso et al., Reference Ayuso, Wooden, Foley, Slack, Sinha, Persing, Goodfellow, McCutcheon and Peter2003; Schulz and Ayuso, Reference Schulz, Ayuso, Goodfellow, McCutcheon and Peter2003; Wilson, Reference Wilson2003; Valverde-Vaquero et al., Reference Valverde-Vaquero, van Staal, McNicoll and Dunning2006; Zagorevski et al., Reference Zagorevski, van Staal, McNicoll and Rogers2007, Reference Zagorevski, van Staal, Rogers, McNicoll, Pollock, Tollo, Batholomew, Hibbard and Karabinos2010; Dennis et al., Reference Dennis, Miller, Hibbard, Tappa, Thunell, Murphy, Strachan and Quesada2020). It is also important to consider that these islands were likely connected to continuous, emergent arcs that may have been oblique, consuming oceanic crust and migrating physically across the greater Iapetus realm (see Domeier, Reference Domeier2016, for estimated position of these island arcs during the Cambrian–Ordovician times). These tectonic processes and the resultant island arcs would have facilitated faunal migration from Gondwana to Laurentia (Nowlan et al., Reference Nowlan, McCracken and McLeod1997; Fortey and Cocks, Reference Fortey and Cocks2003; Harper et al., Reference Harper, Owen, Bruton and Bassett2009). The same method of faunal migration is proposed here, during Floian to Dapingian times, facilitating faunal exchange between the Peruvian Altiplano and the Ganderia and Monian Composite terranes. In addition, Ganderian and Monian composite terranes could have acted as intermediate island arcs allowing subsequent faunal swapping of Gondwanan and Celtic faunas with peri-Gondwanan and peri-Laurentian terranes (Moretown, Dashwoods, and Midland Valley–Mayo terranes). These island arcs collided with Laurentia in the Taconic Orogeny and may have closed the main tract of the Iapetus Ocean in the Floian (Macdonald et al., Reference Macdonald, Ryan-Davis, Coish, Crowley and Karabinos2014, Reference Macdonald, Karabinos, Crowley, Hodgin, Crockford and Delano2017; Karabinos et al., Reference Karabinos, Macdonald and Crowley2017), much earlier than previous estimates (i.e., Late Ordovician, van Staal et al., Reference van Staal, Barr and Murphy2012 and references therein). After their docking with Laurentia, faunal transfer with the Scoto-Appalachian Province might have been facilitated (Neuman, Reference Neuman1984; Neuman and Harper, Reference Neuman, Harper, Webby and Laurie1992; Harper et al., Reference Harper, Mac Niocaill and Williams1996; Liljeroth et al., Reference Liljeroth, Harper, Carlisle and Nielsen2017). The faunal dispersal mechanisms discussed in the preceding may also explain the slight diachronism existing among the Peruvian, Celtic, and peri-Laurentian faunas, with younger occurrences in the latter.
Age of the fossil assemblages
Given the similarities of the Peruvian brachiopods recorded from the lower strata of the Umachiri Formation, especially with the volcaniclastic uppermost Loma del Kilómetro Mb of the Suri Formation and the Molles Formation of the Central Famatina Range and with those of the volcaniclastic Aguada de la Perdiz Formation from the western Puna of northwest Argentina, a similar Floian to ?middle Dapingian (475–468 Ma) age range has been inferred. The age of correlated strata is constrained by the occurrence of conodonts of the Prioniodus elegans, Oepikodus evae, and ‘Baltoniodus navis’ zones in the Famatinian strata (Albanesi and Vaccari, Reference Albanesi and Vaccari1994; Lehnert et al., Reference Lehnert, Bergström and Vaccari1997; Albanesi and Astini, Reference Albanesi and Astini2000) and by associated trilobite and graptolite occurrences in equivalent western Puna strata (Koukharsky et al., Reference Koukharsky, Torres Claro, Etcheverría, Vaccari and Waisfeld1996; Monteros et al., Reference Monteros, Moya and Monaldi1996; Brussa et al., Reference Brussa, Toro, Benedetto and Benedetto2003; Toro et al., Reference Toro, Meroi Arcerito, Muñoz, Waisfeld and de la Puente2015).
The youngest identifiable fossils in the Umachiri Formation are graptolites from the upper part of the Umachiri Formation at locality 5 (Figs. 1, 2), previously reported by Cerrón and Chacaltana (Reference Cerrón and Chacaltana2002). The preservation of these graptolites is poor, but preliminary data suggest a Middle–?Upper Ordovician age for this horizon (J.C. Gutiérrez-Marco, personal communication, 2020).
Acknowledgments
We thank J.L. Benedetto (CONICET) and an anonymous reviewer for their constructive review of the manuscript. We are grateful to J. Newmann (UC Santa Barbara), K. Butler (Western Washington University), and M. Huancca (Universidad Nacional San Antonio Abad del Cusco) for field assistance; to C. Chacaltana (INGEMMET, Lima) for information regarding a graptolite locality southwest of Umachiri; to J.C. Gutiérrez-Marco (Instituto de Geociencias CSIC-UCM, Madrid) for helping in the preparation of the samples, for fruitful discussions, and for critical reading of an earlier version of the manuscript; to C. Alonso (Universidad Complutense de Madrid) and A. Brenner (Harvard University) for fossil photography; to L. Tejada and A. Ticona (INGEMMET, Lima) for curatorial work of the specimens. EBH was funded by Harvard University and JC by the Spanish Ministry of Science and Innovation (Project CGL2017-87631-P). This is also a contribution to the International Geoscience Programme (IGCP) Project 653 ‘The Onset of the Great Biodiversification Event.’





