



Introduction
Mesozoic stalked crinoids have been scarcely recorded from the Southern Hemisphere and most findings consist of mentions of isolated ossicles (mostly columnals) and columnal noditaxis of doubtful taxonomic affinities. In particular, from South America, there are records of isolated ossicles from the Late Triassic–Early Jurassic of the Pucará Basin of Perú (Rosas et al., Reference Rosas, Fontboté and Tankard2007), Early and Late Jurassic of the Neuquén Basin (see details below), Early Jurassic of northern Chile (Aberhan, Reference Aberhan1993), Late Jurassic of the Austral Basin of Argentina (Ramos, Reference Ramos1978, Reference Ramos1981), Early–Late Cretaceous of the Sergipe Basin of Brazil (Ferre et al., Reference Ferre, Walter and Bengston2005), Early Cretaceous of the Neuquén Basin (see details below), and Late Cretaceous of southern Chile (Darwin, Reference Darwin1846; Olivero et al., Reference Olivero, Medina and López C.2009).
Cretaceous stalked crinoids (including Isocrinida, Bourgueticrinida, and Millericrinida) are mostly known from the Northern Hemisphere, and are especially abundant in the European Tethys (e.g., Rasmussen, Reference Rasmussen1961; Klikushin, Reference Klikushin1982; Jagt, Reference Jagt1999; Žítt et al., Reference Žítt, Vodrážka, Hradecká, Svobodá and Zágoršek2006; Salamon, Reference Salamon2009, Reference Salamon2018; Hess and Gale, Reference Hess and Gale2010; Jäger, Reference Jäger2010; Salamon and Gorzelak, Reference Salamon and Gorzelak2010; Lach, Reference Lach2016; Lach and Salamon, Reference Lach and Salamon2016; Salamon et al., Reference Salamon, Ferré and Gorzelak2019), and less so from North America (e.g., Clark and Twitchell, Reference Clark and Twitchell1915; Moore, Reference Moore1967; Ausich et al., Reference Ausich, Buckley and Plint2011; Hunter et al., Reference Hunter, Larson, Landman and Oji2016) and Japan (e.g., Oji, Reference Oji1985; Oji et al., Reference Oji, Kanoh, Toshimitsu and Tashiro1996; Hunter et al., Reference Hunter, Oji, Ewin and Kitazawa2011). At the same time, they may well be considered a rarity in the Southern Hemisphere. Records of articulated specimens are only reported from Antarctica and Australia (Moore, Reference Moore1870; Howchin, Reference Howchin1921; Taylor, Reference Taylor1966). Stalked crinoids are also recorded globally after the Cretaceous/Paleogene boundary from deep- and shallow-water deposits, including from Australia, Antarctica, New Zealand, and South America (see Whittle et al., Reference Whittle, Hunter, Cantrill and McNamara2018). Previously, there have been no descriptions of Mesozoic stalked crinoids from South America. Therefore, the present finding of crinoids, which includes complete specimens, from the Lower Cretaceous Agrio Formation in the Neuquén Basin is potentially important in terms of Cretaceous crinoid diversity, evolutionary paleoecology, and paleobiogeography. At the same time the finding is notable in terms of regional paleoecological implications because most of the previously recorded Early Cretaceous benthic assemblages in the basin are dominated by mollusks, corals, or serpulids, with echinoderms much less abundant (see e.g., Weaver, Reference Weaver1931; Lazo et al., Reference Lazo, Cichowolski, Rodríguez, Aguirre-Urreta, Veiga, Spalletti, Howell and Schwarz2005).
In the Neuquén Basin, there are mentions of dispersed crinoid ossicles and noditaxis from the Jurassic and Lower Cretaceous. Most of these mentions correspond to those characteristic pentalobate or pentastellate columnals and noditaxis, sometimes identified as Pentacrinites sp., which are commonly found loose in calcareous sandstones, but also found dispersed in mollusk-rich shell beds. These records suggest that stalked crinoids were present at least from the Early Jurassic and continued into the Early Cretaceous in the Neuquén Basin. A significant taphonomic bias, mainly by means of disarticulation, dispersal, and eventually disintegration of ossicles, resulted in poor and isolated preservation of crinoids and hampered a clear comprehension of crinoid-dominated benthic associations in the basin.
So far, this was the local picture of Jurassic–Lower Cretaceous crinoids, until the discovery of crinoid beds containing several articulated specimens in the Pilmatué Member of the Agrio Formation, plus another record of an articulated specimen contained in a calcareous concretion from the Agua de la Mula Member of the same formation. Both findings were unexpected, taking into account that we have visited Agrio Formation sections for more than 20 years, but enhanced the study in detail of these crinoids. Therefore, the main objective of the present contribution is to report the description of two new isocrinid species from the Lower Cretaceous of the Neuquén Basin (Argentina) that represent the first record of articulated stalked crinoids from the Mesozoic of South America.
Crinoid bed is here used as a synonym of crinoid skeletal concentration, regardless of the degrees of articulation, ossicle-packing, orientation, size-selection, and geometry. The term encrinite is avoided because it refers to those concentrations where ossicles are densely packed only (e.g., Hunter and Zonneveld, Reference Hunter and Zonneveld2008).
Previous crinoid records from the Neuquén Basin
Mentions of crinoids are scarce in the literature, probably as a consequence of a record that until now has been known by isolated ossicles, even though the presence of pentalobate and pentastellate columnals and noditaxis typical of the Isocrinina made it clear that stalked crinoids were present in the basin from Early Jurassic times, reaching the Early Cretaceous.
In the Lower Jurassic, there are records of columnals from sandstones and fine-grained conglomerates at Cañada Colorada locality (Malargüe, Mendoza) that were reported and figured by Weaver (Reference Weaver1931, pl. 12, fig. 16) as Pentacrinus sp., and a mention of ossicles assigned to Pentacrinites Blumenbach from the Lower Jurassic (Pliensbachian?) of Piedra Pintada, Southern Neuquén (Damborenea et al., Reference Damborenea, Manceñido and Riccardi1975). All of them seem to correspond to columnals and noditaxis. In addition, there are records of Tithonian radials and brachials of pelagic stemless forms assigned to the family Saccocomidae from carbonates of the Vaca Muerta Formation that were identified in thin sections (Kietzmann and Palma, Reference Kietzmann and Palma2009).
From the Lower Cretaceous, there are mentions of ossicles in the Vaca Muerta, Chachao, Mulichinco, and Agrio formations. Crinoid debris was identified in thin sections of carbonates from the lower Valanginian Chachao Formation, Barda Blanca section near Malargüe in Mendoza (Palma, Reference Palma1996). Isolated columnals and brachials were recorded from late Berriasian to early Valanginian thin shell beds of the Vaca Muerta Formation in front of Las Lajas city (D.G.L., personal observation, 2017), and similar columnals were mentioned from shell beds of the Mulichinco Formation at Vega de Escalone by Rodriguez et al. (Reference Rodriguez, Pazos and Aguirre-Urreta2007) in lower Valanginian beds containing the ammonoid Lissonia riveroi (Lisson, Reference Lisson1907). In the Agrio Formation, most of the mentions are restricted to the basal third of the Pilmatué Member (see below), but there is also one new important record from the upper third of the Agua de la Mula Member of a partially articulated crown plus column (this paper). Finally, columnals were recorded in late Hauterivian mollusk-dominated shell beds in the Crioceratites diamantensis Zone of the Agua de la Mula Member of the Agrio Formation at Agua de la Mula (D.G.L., personal observation, 2018).
Geological setting and fossil localities
The Neuquén Basin and Agrio Formation
The studied crinoids come from the Lower Cretaceous Agrio Formation in the Neuquén Basin, west-central Argentina. This basin is one of the most important oil sources in Argentina, and includes one of the most complete and spectacular Jurassic and Cretaceous fossil records in the southern hemisphere. It extends through most of the Neuquén and Mendoza provinces along the Andean foothills, between 32°S and 40°S (Fig. 1). During most of the Jurassic and up to the end of the Early Cretaceous it developed as a back-arc setting under regional thermal subsidence regime due to the initiation of subduction along the western margin of South America (Howell et al., Reference Howell, Schwarz, Spalletti, Veiga, Veiga, Spalletti, Howell and Schwarz2005). The marine connection was towards the Pacific Ocean through an Andean magmatic arc, but at times the arc partially isolated the basin from the open ocean, especially during moments of uplifts and relative sea-level falls (Howell et al., Reference Howell, Schwarz, Spalletti, Veiga, Veiga, Spalletti, Howell and Schwarz2005). During the Early Cretaceous it had a mid-latitudinal location not far from its current position, approximately between 28°S and 36° S, based on reliable palaeomagnetic data (Somoza, Reference Somoza2011).
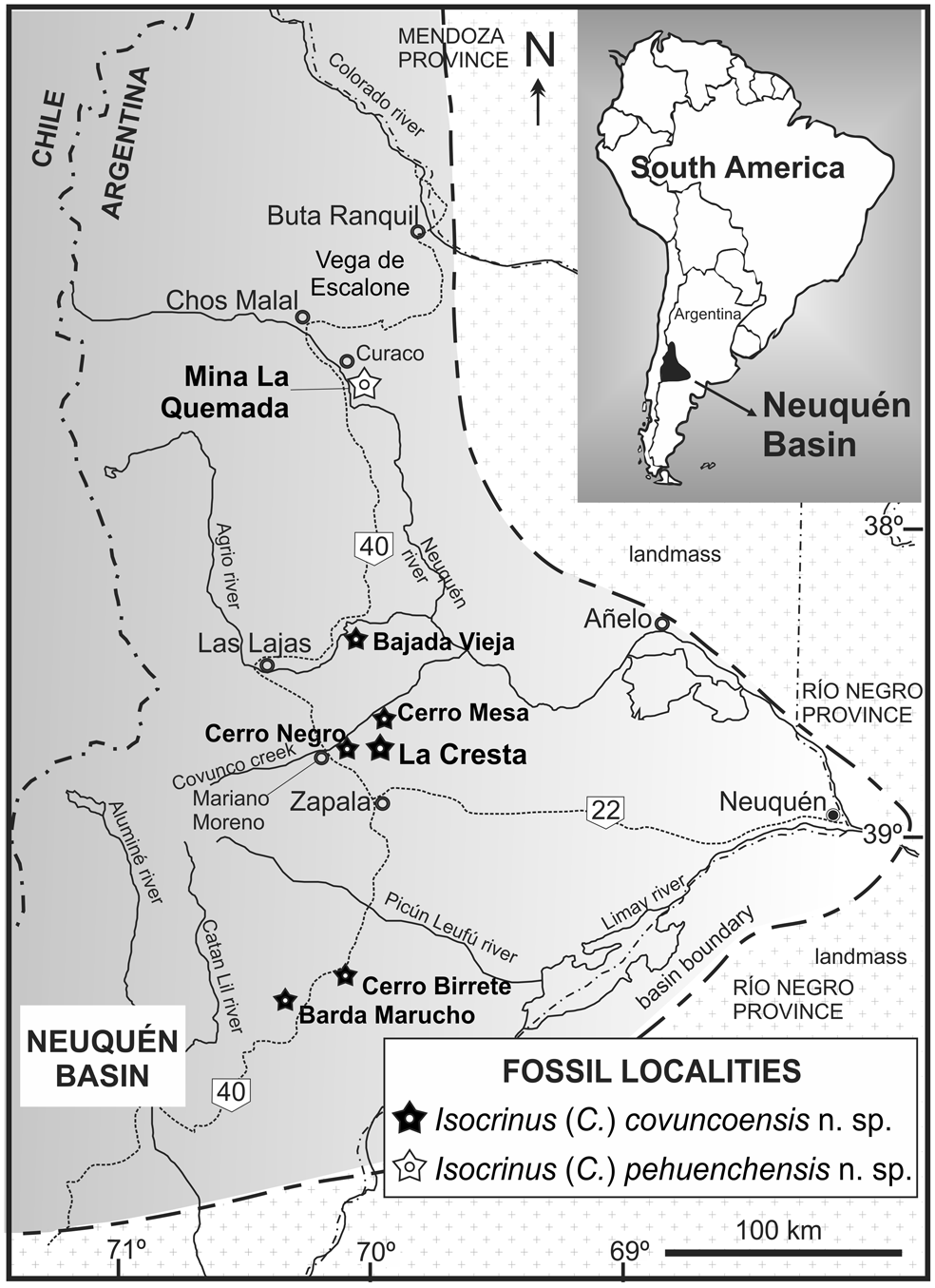
Figure 1. Location map of the Neuquén Basin in west-central Argentina showing fossil localities from where two new species of stalked crinoids are described. Type material of Isocrinus (Chladocrinus) covuncoensis n. sp. (late Valanginian) proceeds from La Cresta locality. Additional material of the species was recorded from Bajada Vieja, Cerro Birrete, Cerro Mesa, Cerro Negro and Barda Marucho. Type material of Isocrinus (Chladocrinus) pehuenchensis n. sp. (late Hauterivian) proceeds from Mina La Quemada locality.
During the Jurassic and Early Cretaceous, the basin accumulated a thick succession of marine and continental clastic, evaporitic, and carbonate deposits, which are divided into several sedimentary cycles (Vergani et al., Reference Vergani, Tankard, Belotti and Welsink1995; Howell et al., Reference Howell, Schwarz, Spalletti, Veiga, Veiga, Spalletti, Howell and Schwarz2005). One of these, the Mendoza Group, corresponds mostly to marine sedimentary rocks that reach a thickness of thousands of meters, deposited from Kimmeridgian to latest Hauterivian times (Leanza et al., Reference Leanza, Hugo, Repol, Gonzalez and Danieli2005; Aguirre-Urreta et al., Reference Aguirre-Urreta, Naipauer, Lescano, López-Martínez, Pujana, Vennari, De Lena, Concheyro and Ramos2019). The Agrio Formation (Fig. 2), defined by Weaver (Reference Weaver1931) in the upper section of the Mendoza Group, has outcrops from Mendoza Province in the north to Piedra del Águila, located in the south of Neuquén Province. It was deposited from early Valanginian to latest Hauterivian times, and comprises three members: the lower Pilmatué Member, the middle Avilé Member, and the upper Agua de la Mula Member (Weaver, Reference Weaver1931; Leanza et al., Reference Leanza, Hugo, Repol, Gonzalez and Danieli2005; Aguirre-Urreta et al., Reference Aguirre-Urreta, Naipauer, Lescano, López-Martínez, Pujana, Vennari, De Lena, Concheyro and Ramos2019). Both the Pilmatué and Agua de la Mula members are mainly composed of black to dark-gray shales and calcareous shales with intercalations of thin sandstone bodies and shell beds that were deposited in a shallow, mixed clastic-carbonate ramp under an overall storm influence (Spalletti et al., Reference Spalletti, Veiga, Schwarz, Leanza, Arregui, Carbone, Danieli and Vallés2011). Both the lower and upper members are characterized by an abundant and diverse marine macrofossil content composed of bivalves, gastropods, cephalopods (nautiloids and ammonoids), corals, sponges, serpulids, and echinoids forming shell beds. A comprehensive monograph on the taxonomy and stratigraphy of these faunas was compiled in Weaver (Reference Weaver1931), but during the last 20 years there have been new detailed studies on these important faunas (e.g., Lazo et al., Reference Lazo, Cichowolski, Rodríguez, Aguirre-Urreta, Veiga, Spalletti, Howell and Schwarz2005; Aguirre-Urreta et al., Reference Aguirre-Urreta, Lazo, Griffin, Vennari, Parras, Cataldo, Garberoglio, Luci, Leanza, Arregui, Carbone, Danieli and Vallés2011; Lazo and Luci, Reference Lazo and Luci2013). On the other hand, the Avilé Member is characterized by aeolian and fluvial sandstones deposited during a short but significant regressive episode of the basin (Veiga et al., Reference Veiga, Spalletti and Flint2002).
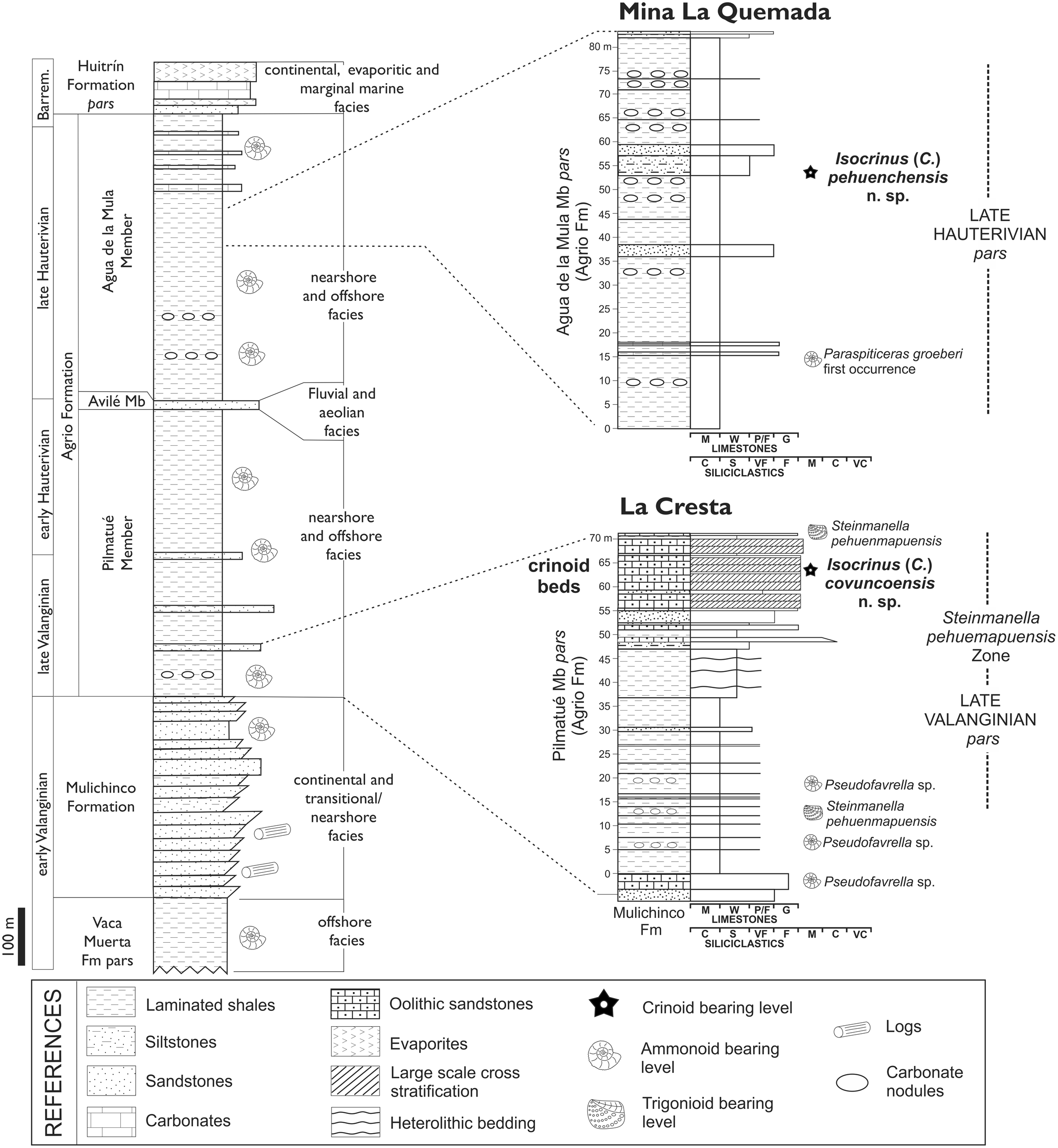
Figure 2. General stratigraphy and ages of the Lower Cretaceous Mendoza Group including the Agrio Formation at the top plus detailed sedimentary logs performed at Mina La Quemada, the type locality of Isocrinus (Chladocrinus) pehuenchensis n. sp., and La Cresta, the type locality of Isocrinus (Chladocrinus) covuncoensis n. sp. Lithostratigraphy based on Leanza et al. (Reference Leanza, Hugo, Repol, Gonzalez and Danieli2005); biostratigraphy and ages from Aguirre-Urreta and Rawson (Reference Aguirre-Urreta and Rawson2010) and Lazo and Luci (Reference Lazo and Luci2013).
Fossil localities and ages
The studied crinoid material came from two localities (La Cresta and Mina La Quemada) and two different stratigraphic positions within the Agrio Formation, each locality corresponding to one of the positions (Fig. 2). Detailed sedimentologic logs of both sections were measured considering the associated ammonoid fauna and following the local ammonoid zonation (see Aguirre-Urreta et al., Reference Aguirre-Urreta, Mourgues, Rawson, Bulot and Jaillard2007; Aguirre-Urreta and Rawson, Reference Aguirre-Urreta and Rawson2010, Reference Aguirre-Urreta and Rawson2012) and the local trigonioid zonation (Lazo and Luci, Reference Lazo and Luci2013). There is additional crinoid material that corresponds to disarticulated ossicles (mostly columnals) and isolated noditaxis coming from other localities, all of them from the Pilmatué Member of the Agrio Formation (see below).
La Cresta and laterally equivalent localities
Isocrinus (Chladocrinus) covuncoensis n. sp. is based mainly on materials collected at La Cresta fossil locality (38°45'S, 69°55'W). The best-preserved specimens in terms of completeness were recorded there, including types. La Cresta (The Crest) is here designated a new fossil locality and its name refers to a hill looking similar to a crest because it is longer than wide and sharp, spanning ~4.5 km along strike. It is placed 1 km south of Cerro Mesa of Covunco (38°44'S, 69°54'W) on the south margin of Covunco Creek.
Cerro Mesa (Table Hill) is in close proximity to La Cresta, and both are ~20 km northeast of Zapala city and 10 km east of Mariano Moreno village (Fig. 1). They are part of the same anticline and easily correlated between each other by means of lithology and fossil content. The sedimentary succession at La Cresta is similar to that outcropping in Cerro Mesa, but beds are dipping SE and outcrops are elongated in NE-SW direction for ~5 km along strike. The Cerro Mesa area is a classical Lower Cretaceous area of the basin where several regional geological and biostratigraphic works have been done in the last century (see further details in Aguirre-Urreta and Rawson, Reference Aguirre-Urreta and Rawson2010 and Veiga and Schwarz, Reference Veiga and Schwarz2017). Description of the Valanginian–Hauterivian succession at Cerro Mesa and logs are presented in Keidel (Reference Keidel1925, fig. 2), Gentili (Reference Gentili1950), and Leanza (Reference Leanza1981, fig. 2). Cerro Mesa was also visited by Weaver (Reference Weaver1931) who collected trigonioid specimens from gray shales at the base of the Agrio Formation. The uppermost beds of the Mulichinco Formation and basal third of the Pilmatué Member of the Agrio Formation are exposed on the flank of Cerro Mesa and La Cresta encompassing the Pseudofavrella angulatiformis and Holcoptychites neuquensis zones of late Valanginian to early Hauterivian age (see Aguirre-Urreta and Rawson, Reference Aguirre-Urreta and Rawson2010). The studied crinoid beds have been accurately dated as late Valanginian by the presence of trigonioid bivalves belonging to the Steinmanella pehuenmapuensis Zone that partially overlaps the Pseudofavrella angulatiformis ammonoid Zone (Fig. 2; see Lazo and Luci, Reference Lazo and Luci2013).
At La Cresta, several thin lenticular crinoid beds were found 60 m above the base of the Pilmatué Member of the Agrio Formation (Fig. 2). The crinoid beds are included in a thick interval that reaches 30 m in thickness that consists of cross-bedded sandstones and carbonate-clastic mixed sediments with variable proportions of bioclasts, ooids, and terrigenous sands. In this succession, the crinoid beds develop throughout a 40 cm thick lens-shaped interval associated with large-scale cross-bedded sets and several reactivation surfaces separating cross-bedded sets (up to 3.5 m thick each set; see Fig. 3.1, 3.2). The scale and architecture of these cross-bedded strata suggest that they represent the migration of large subaqueous dunes (or sand waves) in tide-influenced subtidal settings (see further details in Veiga and Schwarz, Reference Veiga and Schwarz2017).

Figure 3. Field pictures of the crinoid-bearing facies of the Agrio Formation, Valanginian–Hauterivian, Neuquén Basin, west-central Argentina. (1) Cross section view of crinoid beds alternating with sandstones deposited over a reactivation surface that separates two large-scale cross-bedded sets (up to 5 m thick each set). The crinoid beds are included in a thick interval of cross-bedded sandstones, suggesting migration of large subaqueous dunes or sand waves; (2) plan view of the base of the key crinoid bed containing exceptional preservation of several specimens of Isocrinus (Chladocrinus) covuncoensis n. sp.; the bed is considered as a local Konservat Lagerstätte; (3, 4) detailed plan view of the lower surface (base) of the key crinoid bed showing general orientation of specimens (white arrows) and morphological details (column, cirri, radials, and brachitaxes up to Ax6) of Isocrinus (C.) covuncoensis n. sp.; (5) panoramic view of the upper third of the Agua de la Mula Member, Agrio Formation, at Mina La Quemada, showing the stratigraphic position of the concretion level bearing type material of Isocrinus (Chladocrinus) pehuenchensis n. sp., late Hauterivian; (6) cross-section view of the concretion level bearing type material of Isocrinus (C.) pehuenchensis n. sp.; the level is placed in the boundary between laminated shales below and siltstones above. (1–4) From La Cresta locality, Pilmatué Member, Agrio Formation; (5, 6) from Mina La Quemada, Agua de la Mula Member, Agrio Formation.
The best-preserved specimens (MOZ-PI5837) were collected from the lower surface (base) of a key crinoid bed that was easily accessible for study in this outcrop (Fig. 3.3, 3.4). Some of these specimens are almost completely articulated including column with cirri, aboral cup, arms with pinnules, and arm tips. Other specimens are represented by articulated arms with attached pinnules or by isolated pluricolumnals with articulated cirri. The exceptional preservation of articulated crinoids indicates that these specimens were suddenly buried, probably during life, and no subsequent reworking or bioturbation took place, and thus may well be considered a local Konservat Lagerstätte due to sporadic obrution events (see Veiga and Schwarz, Reference Veiga and Schwarz2017). Apart from the key crinoid surface, the rest of the crinoid beds at La Cresta are composed of moderately to highly disarticulated remains with dense ossicle packing. The preservational features of the latter beds suggest significant exposure time prior to final burial in the dune trough.
A laterally equivalent interval of calcareous sandstones and crinoid beds is recorded at the top hill succession at Cerro Mesa, but due to the outcrop condition these beds are only observable in cross-section, not in plan view, and thus observation and collection of crinoids were rather difficult.
Several occurrences of isolated crinoid ossicles occur in beds that are lateral equivalents to the crinoid beds of La Cresta locality (see Fig. 1). These records come from sandstone beds at Cerro Negro, Cerro Birrete and Barda Marucho (Gentili, Reference Gentili1950, p. 13; Leanza, Reference Leanza1981, p. 2–3; Leanza and Hugo, Reference Leanza and Hugo1997, p. 49; Lescano and Concheyro, Reference Lescano and Concheyro2009, p. 77; Daroca, Reference Daroca2011, p. 16) and one thin shell bed at Bajada Vieja (Lazo, Reference Lazo2004).
The sections outcropping at Cerro Mesa and Cerro Negro are very similar to each other, including the upper part of the Mulichinco Formation and the basal half of the Agrio Formation. Cerro Negro section is within a military base and only isolated columnals were observed in the basal third of the Pilmatué Member of the Agrio Formation within the Pseudofavrella angulatiformis Zone (D.G.L., personal observation, 2011).
Isolated noditaxis and columnals were observed in sandstone beds that lie in the transition between the Bajada Colorada and Agrio formations in Cerro Birrete and Barda Marucho localities (D.G.L., personal observation, 2008). Just above these beds there is a shale interval of the Pilmatué Member that includes the first occurrence of ammonoids of the Decliveites crassicostatus Subzone of late Valanginian age, thus they are probably laterally equivalent of those crinoid beds outcropping at La Cresta and Cerro Mesa localities. For more information on the last two localities see Lescano and Concheyro (Reference Lescano and Concheyro2009) and Cataldo and Lazo (Reference Cataldo and Lazo2012).
Mina La Quemada
The second locality in importance (after La Cresta) in terms of crinoid preservation is Mina La Quemada (Fig. 1), which corresponds to an abandoned coal mine (37°35'S, 70°00'W) east of state road 9, ~7 km south from Curaco settlement, Pehuenches County. The studied material of Isocrinus (Chladocrinus) pehuenchensis n. sp. was found in an isolated, 12 cm-long, subspherical, silty calcareous concretion in the upper third of the Agua de la Mula Member of the Agrio Formation (Fig. 2), ~40 m above the first occurrence of the ammonoid Paraspiticeras groeberi Aguirre-Urreta and Rawson, Reference Aguirre-Urreta and Rawson1993, late Hauterivian (Aguirre-Urreta et al., Reference Aguirre-Urreta, Naipauer, Lescano, López-Martínez, Pujana, Vennari, De Lena, Concheyro and Ramos2019). One articulated specimen (the holotype), including aboral cup, proximal arms (IBr, IIBr, and proximal IIIBr), and first pinnules, was recorded in the concretion along with fragments of column and cirri. The concretion was sampled from a level of concretions situated along a sharp boundary between a tabular shale interval below and a tabular siltstone to very fine-grained sandstone interval above (Fig. 3.5, 3.6). Several other inspected concretions from this level were unfortunately barren. The presence of a concretion level along this boundary may indicate a minor sedimentary pause that enhanced carbonate cementation. The crinoid concretion sample, although found isolated, is quite significant in terms of associated facies, because it was found in finer-grained facies than the previously mentioned crinoid beds, and thus represents a different setting from that envisaged for I. (Chladocrinus) covuncoensis n. sp. in the Pilmatué Member.
Materials and methods
Repositories and institutional abbreviations
The great majority of the studied specimens are deposited in the repository of the Museo Juan Olsacher (MOZ-Pi), Zapala city, Neuquén Province (Argentina), except one specimen that is housed in the Museo Patagónico de Ciencias Naturales (MPCN-Pi), General Roca city, Río Negro Province (Argentina) that was collected by J.C. Salgado, its former director. Suffix numbers indicate the specimen number of a given catalog number.
Systematic paleontology
The general terminology utilized to describe the crinoid endoskeleton follows Ubaghs (Reference Ubaghs, Moore and Teichert1978), Simms (Reference Simms1989), and Hess and Messing (Reference Hess, Messing and Selden2011); while the classification down to the genus level follows the proposal of Hess and Messing (Reference Hess, Messing and Selden2011). Description follows, when possible, the standard approach proposed by Fearnhead (Reference Fearnhead2008) and some measurements, ratios, and categories were based on those proposed by Simms (Reference Simms1989).
Measurements were taken on type material, unless stated otherwise and are as follows: H: cup/ossicle height; W: cup/ossicles width; D: ossicle diameter, measured in the same direction as ossicle width but used instead when ossicle is rounded; x/y: ratio measured on columnals (see Simms, Reference Simms1989, text-fig. 3); % width cirrus socket: width of cirrus socket expressed as percentage of nodal width; % width axial canal: width of axial canal expressed as percentage of columnal width; Wproxis/distis: width of proxistele/dististele. On the basis of the number of columnals per noditaxis, these can be classified into three groups: short (<10 columnals per noditaxis), moderate (10–25 columnals per noditaxis), and long (>25 columnals per noditaxis). Columnal outline is defined by the ratio of two measurements (x, y), which are dependent upon the angle between adjacent interradii (see Simms, Reference Simms1989, p. 6, text-fig. 3b).
Subclass Articulata Zittel, Reference Zittel1879
Order Isocrinida Sieverts-Doreck, Reference Sieverts-Doreck, Moore, Lalicker and Fischer1952
Suborder Isocrinina Sieverts-Doreck, Reference Sieverts-Doreck and Priveteau1953
Family Isocrinidae Gislén, Reference Gislén1924
Subfamily Isocrininae Gislén, Reference Gislén1924
Diagnosis
Two primibrachials (IBr1-2); synarthry between secundibrachials 1 and 2 (IIBr1-2, Hess and Messing, Reference Hess, Messing and Selden2011).
Occurrence
Middle Triassic (?Anisian)–Recent (Hess and Messing, Reference Hess, Messing and Selden2011).
Remarks
Specimens of the family Isocrinidae are characterized by synarthry between primibrachials 1 and 2 (IBr1-2). Synarthry is a type of very flexible ligamentary articulation conformed by two opposing bundles of ligaments that are separated by a fulcral ridge. It is easily identified laterally by the presence of the middle fulcral ridge crossing the articulation face. Specimens of the subfamily Isocrininae are characterized by the presence of only two primibrachials and synarthry between secundibrachials 1 and 2 (IIBr1-2). These characteristics are recognized in both studied species (see below).
Genus Isocrinus von Meyer in Agassiz, Reference Agassiz1836
Type species
Isocrinites pendulus von Meyer in Agassiz, Reference Agassiz1836, p. 195, holotype specimen considered lost (see Hess and Messing, Reference Hess, Messing and Selden2011).
Other species
The best known species of Isocrinus worldwide is I. (I.) nicoleti (Desor) from the Middle Jurassic of Europe, but also four species from the Lower Cretaceous are well-known: I. aptiensis Taylor, Reference Taylor1966; I. australis (Moore, Reference Moore1870); I. (Chladocrinus) hanaii Oji, Reference Oji1985; and Isocrinus parvus Howchin, Reference Howchin1921, all of which are used for morphological comparisons with the studied material. There are also well-known Lower Jurassic species, such as Isocrinus (Chladocrinus) basaltiformis (Miller, Reference Miller1821) and Isocrinus (Chladocrinus) tuberculatus (Miller, Reference Miller1821) that, due to their completeness, deserve morphological comparisons with the studied material.
Diagnosis
Basals small, visible from outside, not forming contiguous circlet. Columnals low, except in distalmost part of column, cirrus sockets large, as high as nodals (Hess and Messing, Reference Hess, Messing and Selden2011).
Occurrence
Upper Triassic (?Carnian), Lower Jurassic–Lower Cretaceous, Europe, Russia, Crimea, Caucasus, Japan, Australia, Antarctica, and Argentina (Hess and Messing, Reference Hess, Messing and Selden2011; this paper).
Remarks
The studied material of both species match the diagnostic characteristics of the genus Isocrinus.
Subgenus Chladocrinus Agassiz, 1836
Type species
Pentacrinites basaltiformis Miller, Reference Miller1821
Diagnosis
Straight cryptosyzygy between secundibrachials 3 and 4 (IIBr3-4). Internodes long, with as many as 20 columnals (Hess and Messing, Reference Hess, Messing and Selden2011).
Occurrence
Upper Triassic (?Carnian), Lower Jurassic (Hettangian)–Lower Cretaceous (Aptian), Europe, Russia, Japan and Argentina (Hess and Messing, Reference Hess, Messing and Selden2011; this paper).
Remarks
The studied material of both species match the diagnostic characteristics of the subgenus Chladocrinus, except for the absence of long noditaxis showing instead short noditaxis with up to 6–7 columnals.
Isocrinus (Chladocrinus) covuncoensis new species
Figures 4–9
Types
Three specimens preserved on the base of one block of crinoidal limestone (Fig. 4). Holotype MOZ-Pi5837-1 (Fig. 5) corresponds to a single individual that includes aboral cup, proximal arms (up to IVBr), some pinnules, proximal half of column, and cirri; paratype MOZ-Pi5837-2 (Fig. 6.1, 6.2) is a single individual that includes an almost complete crown (up to VIIBr), proximal column, and cirri; paratype MOZ-Pi5837-3 (Fig. 6.3, 6.4) is a single individual that includes only part of the crown (up to VIBr). There are two other relatively complete individuals surrounding the mentioned specimens, but they are highly recrystallized and are difficult to study or figure. There are also loose pluricolumnals with cirri and isolated ossicles around the types (see Fig. 4).

Figure 4. Isocrinus (Chladocrinus) covuncoensis n. sp., general aspect of limestone block MOZ-Pi 5837 in plan view (lower surface) showing the three types (MOZ-Pi 5837-1, -2, -3) placed at the base of the key crinoid bed. Arrow shows slightly curved distal portion of column.
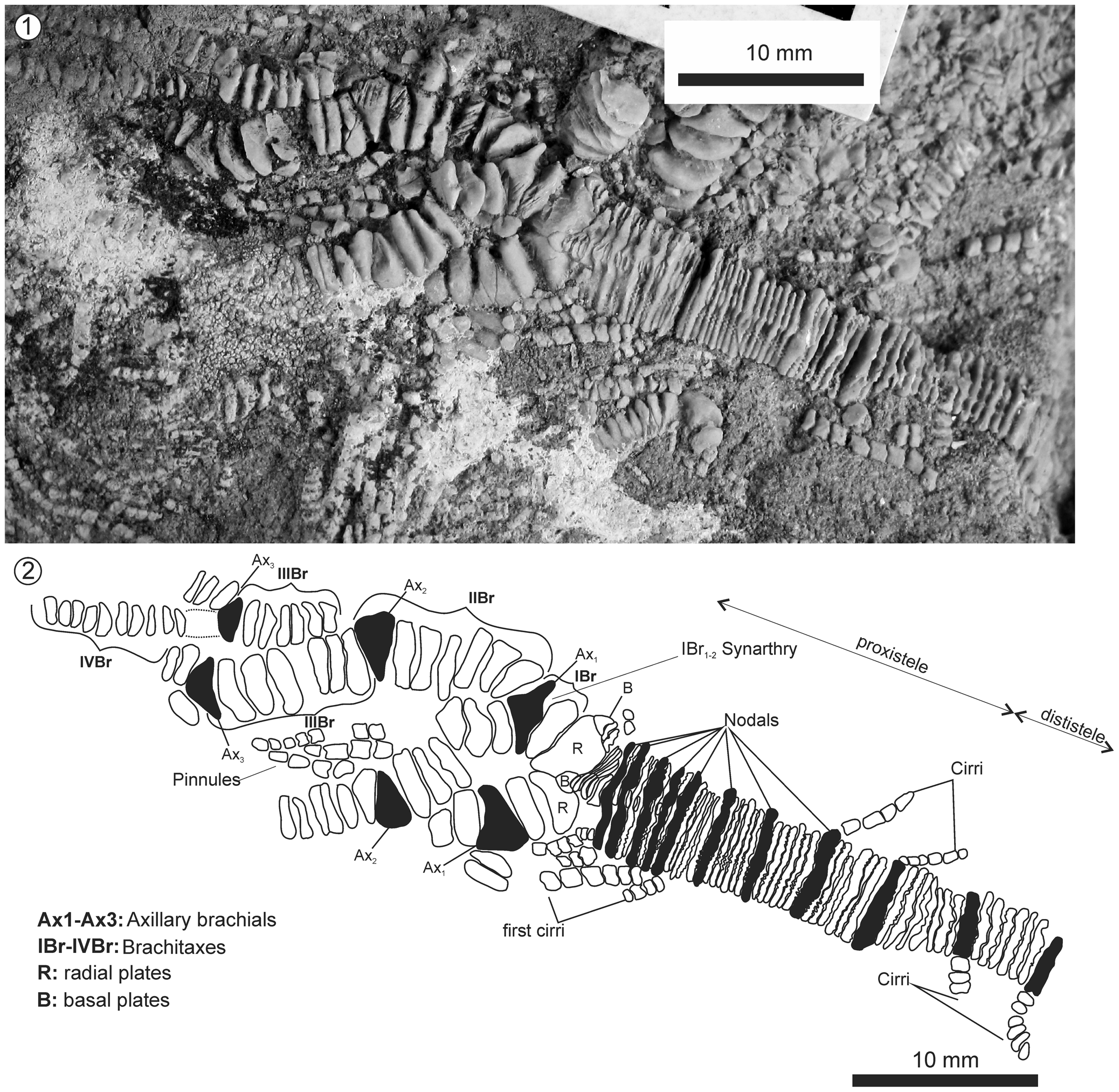
Figure 5. Isocrinus (Chladocrinus) covuncoensis n. sp., La Cresta locality, Pilmatué Member, Agrio Formation, late Valanginian. (1) Detailed picture of holotype MOZ-Pi 5837-1; (2) schematic drawing of the same specimen showing morphological details.
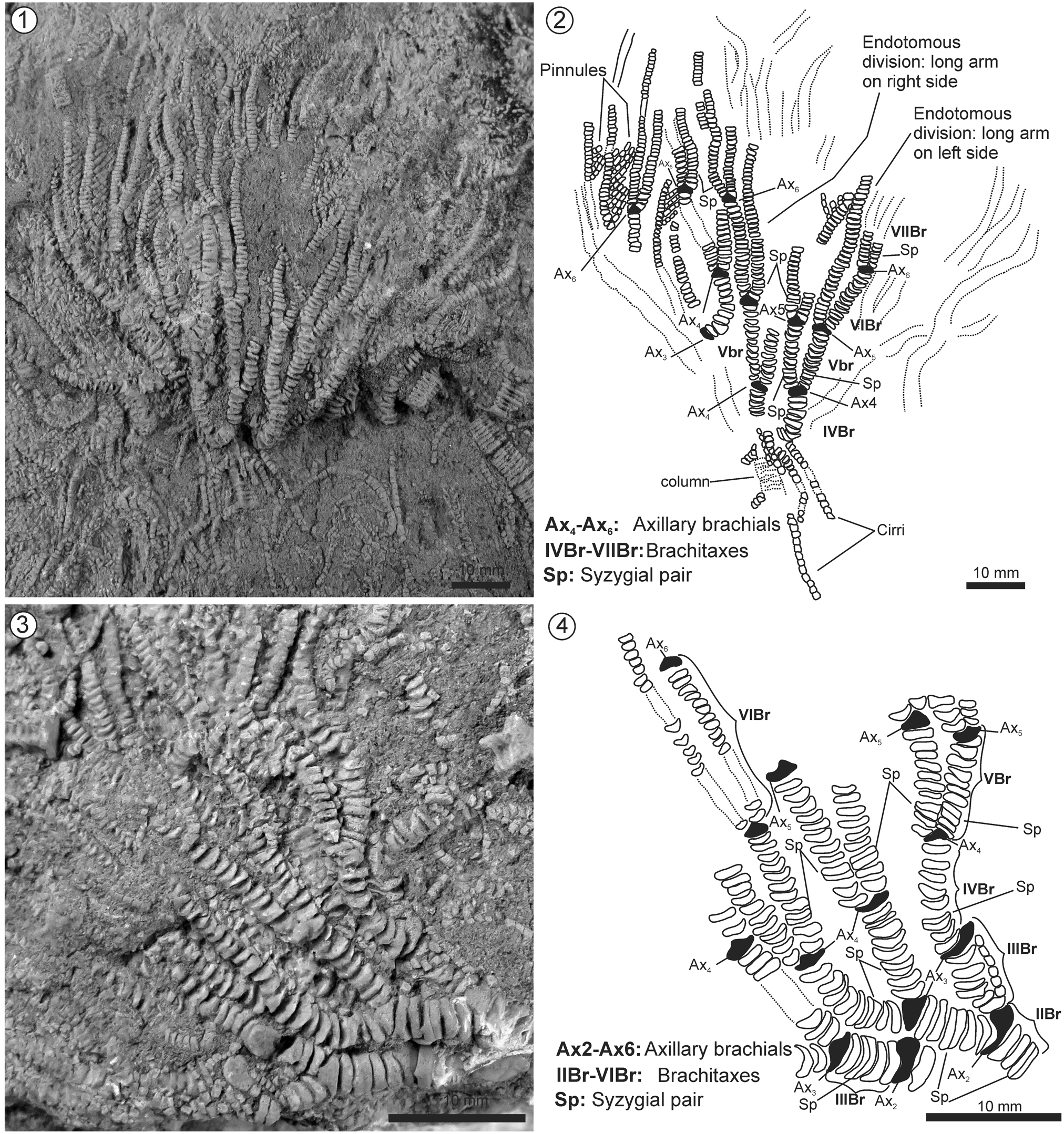
Figure 6. Isocrinus (Chladocrinus) covuncoensis n. sp., La Cresta locality, Pilmatué Member, Agrio Formation, late Valanginian. (1) Detailed picture of paratype MOZ-Pi 5837-2; (2) schematic drawing of the same specimen showing morphological details; (3) detailed picture of paratype MOZ-Pi 5837-3; (4) schematic drawing of the same specimen showing morphological details.

Figure 7. Isocrinus (Chladocrinus) covuncoensis n. sp., La Cresta locality, Pilmatué Member, Agrio Formation, late Valanginian. (1) Detailed picture of specimen MOZ-Pi 5836-1a; (2) schematic drawing of the same specimen showing morphological details.

Figure 8. Schematic representation of branching style of Isocrinus (Chladocrinus) covuncoensis n. sp., length and width of arms are not to scale. For abbreviations see Figures 5–7.
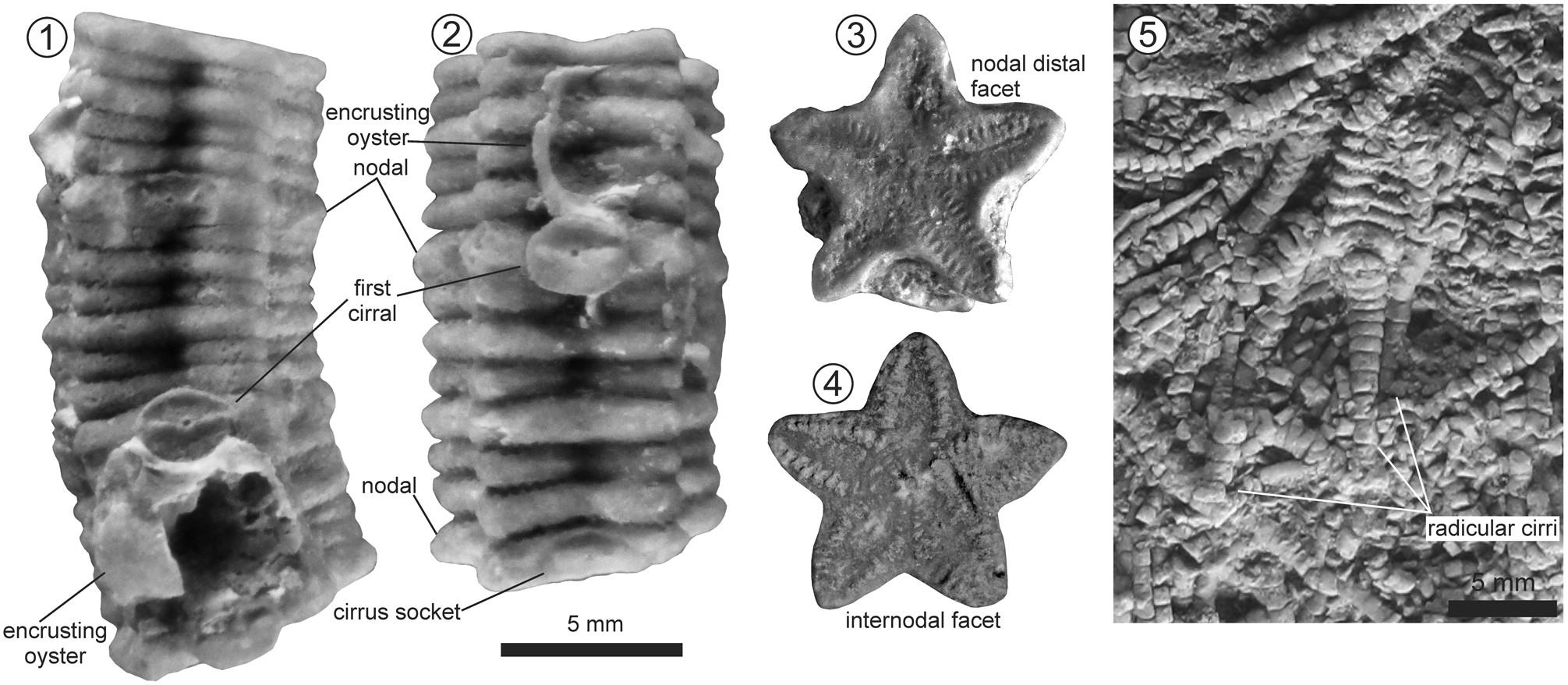
Figure 9. Column of Isocrinus (Chladocrinus) covuncoensis n. sp., La Cresta locality, Pilmatué Member, Agrio Formation, late Valanginian. (1) Pluricolumnal MOZ-Pi 5838-1; (2) pluricolumnal MOZ-Pi 5838-2; (3) detail of slightly concave nodal distal facet showing cryptosymplectial articulation, MOZ-Pi 5838-1; (4) internodal facet showing symplectial articulation, MOZ-Pi 5838-8; (5) detailed picture of specimen MOZ-Pi 5836-1b showing radicular cirri.
Diagnosis
Small aboral cup with depressed base; slender, multiramose crown with six arm divisions, isotomous branching except endotomous in axillary 5 (Ax5); seven (or rarely eight) secundibrachials (IIBr); 240 arm tips; uniform pinnules; crown slightly higher than column; column smooth and stout; differentiated proxistele and dististele; pentalobate columnals; interradius acuminated; short noditaxis (6–7 columnals); petaloid crenullae, 22–24 in number per interradius, axial canal small.
Occurrence
Types were collected from crinoid beds 60 m above the base of the Agrio Formation at La Cresta locality (Fig. 1). These beds were accurately dated as late Valanginian by the presence of trigonioid bivalves belonging to the Steinmanella pehuenmapuensis Zone that partially overlaps the Pseudofavrella angulatiformis ammonoid Zone (see Lazo and Luci, Reference Lazo and Luci2013). The latter partially overlaps the Criosarasinella furcillata Zone of the standard ammonoid zonation of the Mediterranean Region (see Reboulet et al., Reference Reboulet, Rawson, Moreno-Bedmard, Aguirre-Urreta, Barragán, Bogomolov, Company, González-Arreola, Stoyanova, Lukeneder, Matrion, Mitta, Randrianaly, Vašiček, Baraboshkin, Berto, Bersac, Bogdanova, Bulot, Latil, Mikhailova, Ropolo and Szives2011). Isocrinus (C.) covuncoensis n. sp. is endemic to the Neuquén Basin, west-central Argentina. Besides the type locality, it has been recorded as isolated noditaxis and columnals from late Valanginian sandstone beds of the Pilmatué Member of the Agrio Formation at the following localities: Cerro Mesa, Cerro Negro, Cerro Birrete, Barda Marucho, and Bajada Vieja (Fig. 1).
Description
Crown fan-shaped, (H = 83 mm, W = 61 mm), slender, with delicate arm branching especially towards arm tips, multiramose.
Dorsal (aboral) cup small, flat (W = 6.9 mm; H = 2.5 mm; H/W = 0.36), a little wider than the column (column width ~4.9 mm), depressed base, cone-shaped, radial symmetry, externally smooth. Infrabasals not visible in adult cup; basals small, separated on surface; radials poorly projected laterally, shorter than wide (H = 2.4 mm, W = 3.9 mm, H/W = 0.61), separated in its lower half from adjacent radials by small subconical basals (H = 0.9 mm, W = 1.12 mm, H/W = 0.80). Radial-radial articulation is flat, probably zygosynostosial. Tegmen unknown.
The arms reach ~80 mm in height in the largest specimen (Fig. 8), with up to six axillaries, brachitaxes range from IBr to VIIBr. All axillaries show isotomous divisions, except axillary 5 (Ax5), which shows endotomous division giving way to one long arm tip and a short arm; the short one has another axillary (Ax6) that in turn shows isotomous division; eventually giving way to two equally long arm tips. Arms slenderer toward distal ends, ranging in diameter from 4.4 mm (IBr) proximally to 1.0 mm (VIIBr) distally; almost uniform in diameter within each brachitaxis, except hexabrachials and heptabrachials narrowing distally. Each arm branches in 48 arm tips (see Fig. 8) reaching a total of 240 arm tips. Cryptosyzygial ligament articulation (=syzygial pair) developed between third and fourth brachial in each brachitaxis from IIBr to VIIBr (Figs. 6, 7).
Primibrachials two; articulation IBr1-2 synarthrial (Fig. 5), IBr2 axillary 1. Secundibrachials seven; articulation IIBr1-2 synarthrial (Fig. 7), articulation IIBr3-4 cryptosyzygial conforming a syzygial pair (Figs. 6, 7). Tertibrachials (IIIBr) seven, quartibrachials (IVBr) usually 10, but up to 13, pentabrachials (VBr) usually 12–13, but occasionally 15, hexabrachials (VIBr) 14 (short arm up to Ax6) to more than 30 (i.e., long arm), probably finials; heptabrachials (VIIBr) more than 25, but exact number unknown due to poor arm tip preservation, probably represent finials due to distal tapering.
First pinnule at IIBr2 (Fig. 7); subsequent pinnules arise from alternative sides of successive brachials, except for hypozygals that lack pinnule; uniserial; close-packed arrangement; tapering gradually to distal end. No apparent differentiation of pinnules throughout arms. Pinnulars subrectangular (=cylindrical) to subquadrate; ~10 in number in longest pinnules; 10 mm maximum length, tapering in size gradually to distal end; lateral smooth, unornamented.
Column stout (Wcolumnal/Wcup = 1.01); latera smooth; maximum measured height 65 mm (specimen not sampled, measured in the field); shorter than crown; straight; heteromorphic; divided into two portions: proxistele and dististele. Proxistele ~20 mm in height; maximum width ~6.5 mm; columnals reduced in height (H < 1 mm), almost flat, columnal outline pentalobate (x/y = 0.2, see categories in Simms, Reference Simms1989, p. 6); nodals very close to each other, higher than internodals; number of internodals gradually increasing from one to six.
Dististele (H ≤ 45 mm) higher than proxistele; columnals pentalobate; interradius accuminated; usually 5 or 6 internodals (i.e., 6–7 ossicles per noditaxis; short noditaxis); Hnodal = 1.25 mm; Hinternodal = 0.85 mm; maximum width ~6.5–7 mm; nodals slightly higher than internodals; although some internodals are almost as high as the nodals; alternating slightly in diameters. Dististele generally straight, except distal portion slightly curved (Figs. 4, 9). Articulation between nodal and infranodal cryptosymplexy; nodal distal facet slightly concave; proximal articulation of nodals and articulation between internodals symplexy. Central circular axial canal represents 4.29% of columnal width; with surrounding areola. Petaloid crenularium with 22–24 crenullae per interradius.
Cirrus sockets five in number per nodal; as high as the nodal, moderate sized, 34–38% of nodal width; distinct transverse ridge that divides laterally into two short oblique ridges (synarthry); central circular axial canal with raised lumen. Cirri slender and long, present along the entire length of column (Fig. 5); undivided; generally perpendicular to column, at least initially, bending upwards or downwards (proximally) or downwards (distally) becoming somewhat parallel to column; proximal cirri covering aboral cup. Cirrals narrow (proximally) to barrel-shaped (distally), smooth; distal articulation by traverse ridge and central circular axial canal; widest cirrus with 28 cirrals (W = 21 mm; Fig. 4); tapering distally. Cirri at the distal column pointing downwards, acting as radicular or anchoring cirri; undivided (Fig. 9).
Etymology
After Covunco Creek, which is the main stream next to La Cresta fossil locality, from where types were collected.
Materials
MOZ-Pi5836 is a batch of two blocks of crinoidal limestone containing three specimens. MOZ-Pi5836-1a is a single individual that includes articulated arms up to VIIBr, but without cup and column (Fig. 7); MOZ-Pi5836-2 is a single individual that includes a partially disarticulated crown; MOZ-Pi5836-1b corresponds to one distal pluricolumnal with cirri (Fig. 9); MOZ-Pi5838 is a batch of 15 loose pluricolumnals (Fig. 9) plus one isolated arm with pinnules. All from the same crinoid beds of La Cresta locality. MPCN-Pi 000693 corresponds to one block of crinoidal limestone, partially recrystallized, containing one articulated specimen plus several isolated columns with cirri from the Lower Cretaceous of Cerro Mesa of Covunco that is currently exhibited in the Museo Patagónico de Ciencias Naturales, General Roca city.
Measurements
See Table 1.
Table 1. Measurements used for description of Isocrinus (Chladocrinus) covuncoensis n. sp. and I. (C.) pehuenchensis n. sp. based on type specimens MOZ-Pi 5837 and MOZ-Pi 5823. For explanations on ratios and percentages see the Systematic Paleontology section. Some measurements, ratios, and categories are based on Simms (Reference Simms1989).

Remarks
Isocrinus (Chladocrinus) covuncoensis n. sp. is described on the basis of beautifully preserved materials, being one of the most complete examples among Cretaceous isocrinids in the world. It could be described in detail based on several articulated specimens from the same key crinoid bed from La Cresta locality. Therefore, it is easy to compare with other Cretaceous species of Isocrinus. Those complete specimens (e.g., MOZ-Pi5837) of the studied material that are relatively of a uniform size may represent the adult stage of I. (C.) covuncoensis n. sp. Morphological comparison with I. (C.) pehuenchensis n. sp. is presented below the formal description of the latter.
There are many Cretaceous species of isocrinids that are tentatively assigned to Isocrinus only on the basis of isolated columnals or pluricolumnals (e.g., Rasmussen, Reference Rasmussen1961; Salamon and Gorzelak, Reference Salamon and Gorzelak2010; Lach, Reference Lach2016; Lach and Salamon, Reference Lach and Salamon2016; Salamon, Reference Salamon2018; Salamon et al., Reference Salamon, Ferré and Gorzelak2019), which have mostly been assigned to “Isocrinus?” and therefore is useless to compare them one by one with I. (C.) covuncoensis n. sp. because columnal variability may be high even within a single species and thus it does not warrant reliable taxonomic discussion. The present species will be compared only to those well-known Jurassic and Cretaceous species of Isocrinus, including those from the Lower Cretaceous of Antarctica, Australia, and Japan.
Isocrinus (Chladocrinus) basaltiformis (Miller, Reference Miller1821, p. 62–63, pl. II), originally described from the Lower Jurassic (Pliensbachian) of England, differs by having pentagonal columnals, ornamented latera, and larger number (up to 20) of internodals (see Klikushin, Reference Klikushin1987, fig. 1, A–C; Hess and Messing, Reference Hess, Messing and Selden2011, fig. 26, 1a–c).
Isocrinus (Chladocrinus) tuberculatus (Miller, Reference Miller1821, p. 64–65), originally described from the Lower Jurassic of England, differs by having larger number of secundibrachials, sub-pentalobate columnal outline, and higher columnals (see Hess and Messing, Reference Hess, Messing and Selden2011, fig. 26, 1d).
Isocrinus nicoleti (Desor, Reference Desor1845), originally described from the Middle Jurassic (Bathonian) of France, is similar to I. (C.) covuncoensis n. sp. in terms of cup morphology, number of primibrachials, number of internodals, and columnal morphology, but it differs by having more secundibrachials (7–10), more tertibrachials (14–17), fewer arm tips (32), and higher column (up to 89 mm) (see de Loriol, Reference de Loriol1887, p. 165, pl. 154–161; Hess, Reference Hess2012, fig. 4).
Isocrinus aptiensis Taylor (Reference Taylor1966, fig. 6a, b, d, f) from the Aptian of the east coast of Alexander Island, Antarctica, was described on the basis of several specimens with almost complete crowns and proximal columns articulated. It is easily differentiated from I. (C.) covuncoensis n. sp. by its smaller size, higher column than crown, greater number of IIBr (13–15) and IIIBr (≥15), higher than wide brachials, and less ramified crown (up to Ax2).
Isocrinus australis (Moore, Reference Moore1870, p. 243, pl. 17, fig. 3, pl. 18, fig. 1) is a well-known species from the Lower Cretaceous of Australia, especially from the late Aptian Doncaster Member of the Wallumbilla Formation (see Cook, Reference Cook2012). It was originally described from Queensland with subsequent further material described from the same state (Etheridge, Reference Etheridge1901, pl. 1, fig. 4, pl. 3, figs. 1–3). Additional opalized material was described from coeval deposits in White Cliffs, New South Wales (Etheridge, Reference Etheridge1904, pl. 28, as subspecies I. australis albascopularis) and from the Stuart Range, Southern Australia (Howchin, Reference Howchin1921, pl. 1, figs. 1, 2). Isocrinus australis has a similar cup configuration, equal number of IBr (2), and isotomous branching on proximal arms, but it has greater number of IIBr (13), IIIBr (15), and VBr (≥54) than I. (C.) covuncoensis n. sp. If VBr are taken as finials (see Etheridge, Reference Etheridge1901, p. 9), then I. australis has fewer axillaries and arm tips. Height of crown is also larger in the Australian species (>200 mm).
Isocrinus parvus Howchin (Reference Howchin1921, pl. 1, fig. 3) from the Aptian–Albian of Southern Australia is based on a single specimen that corresponds to an upper fragment of a crown including brachials and pinnules (H = 50.8 mm) without aboral cup. There are 28–35 finials above a basal isotomous division. It is difficult to compare this species because it is highly incomplete, and thus should be taken with uncertainty. It should be checked whether it is a different species or synonymous with I. australis. Taking into account the size of the finial arms, this Australian species is larger than I. (C.) covuncoensis n. sp.
Isocrinus (Chladocrinus) hanaii Oji (Reference Oji1985, pl. 77, pl. 78, figs. 1–5, 7, 8, pl. 79, figs. 1–7, text-fig. 4A) from the upper Aptian of the Hiraiga Formation, Miyako Group, Northeast Japan is based on well-preserved specimens, including crowns and columns. It has some similarities with I. (C.) covuncoensis n. sp. including cup configuration and presence of cryptosyzygial pairs between third and fourth brachials in nearly every brachitaxis, but differs in having: (1) stouter and less dense crown; (2) different arm branching pattern corresponding to three proximal isotomous divisions followed by a fourth endotomous division and a last fifth isotomous division, with a total of 120 arm tips; (3) greater number of IIBr (12–13), IIIBr (17, 19, 21), and IVBr (21, 25, 37); (4) different shape of columnals that are pentagonal to subpentagonal; (5) greater number of internodals (7–8, rarely 9); and (6) larger cirri.
Isocrinus (Chladocrinus) pehuenchensis new species
Figure 10
Holotype
One subspherical calcareous concretion (MOZ-Pi 5823), 11 cm in diameter, containing one articulated aboral cup, proximal arms, first pinnules, and first four columnals (H ~16 mm). In addition, one fragment of column (H ~11.8 mm) with cirri, preserved in the same calcareous concretion. However, not directly articulated with the mentioned cup, it is most probably part of the same specimen because they are closely spaced in the concretion.

Figure 10. Isocrinus (Chladocrinus) pehuenchensis n. sp., holotype MOZ-Pi 5823, Mina La Quemada, Agua de la Mula Member, Agrio Formation, late Hauterivian. (1, 2) Detailed views of crown; (3, 5) detailed views of column and cirri; (4) detailed view of distal facet of proximal columnal. For abbreviations see Figures 5–7.
Diagnosis
Isocrinin with small aboral cup with flat base; isotomous branching; eight or nine secundibrachials (IIBr); column slender; differentiated proxistele and dististele; columnals pentastellate and ornamented with medial ridge with fine tubercles; interradius acuminated with fine tubercles on its tip; short noditaxis (6 columnals); triangular crenullae, 14 in number per interradius, axial canal large.
Occurrence
Isocrinus (Chladocrinus) pehuenchensis n. sp. is recorded in the upper third of the Agua de la Mula Member of the Agrio Formation, Mina La Quemada locality, 40 m above the first occurrence of Paraspiticeras groeberi Aguirre-Urreta and Rawson, Reference Aguirre-Urreta and Rawson1993, belonging to the homonym ammonoid zone, upper Hauterivian. The latter partially overlaps the Pseudothurmannia ohmi Zone of the standard ammonoid zonation of the Mediterranean Region (see Reboulet et al., Reference Reboulet, Rawson, Moreno-Bedmard, Aguirre-Urreta, Barragán, Bogomolov, Company, González-Arreola, Stoyanova, Lukeneder, Matrion, Mitta, Randrianaly, Vašiček, Baraboshkin, Berto, Bersac, Bogdanova, Bulot, Latil, Mikhailova, Ropolo and Szives2011).
Description
Crown of unknown shape and size, only proximal arm branching is preserved.
Dorsal (aboral) cup small, very flat (W = 5.9 mm; H = 1.4 mm; H/W = 0.23), much wider than the column (Wcolumn ~3.45 mm), slightly depressed base, radial symmetry, externally smooth. Infrabasals not visible in adult cup; basals small, separated on surface; distal surface of basals shows symplectial articulation (Fig. 10.4) with first columnal; radials are subrectangular and projected laterally; shorter than width (H = 1.7 mm, W = 4.6 mm, H/W = 0.36), separated in lower half from adjacent radials by small subconical basals (H = 1.2 mm, W = 1.3 mm, H/W = 0.92). Upper surface of radials (just in contact with IBr1) forms an obtuse angle with its lower part, probably a muscular articulation. Radial-radial articulation is flat, probably synostosial. Tegmen unknown.
Only proximal brachitaxes preserved, from IBr to basal IIIBr. Primibrachials (W = 4.5 mm) two; articulation IBr1-2 synarthrial (Fig. 10.1, 10.2), IBr2 axillary 1. Secundibrachials eight or nine; articulation IIBr1-2 synarthrial (Fig. 10.1, 10.2), articulation IIBr3-4 cryptosyzygial conforming a syzygial pair (Fig. 10.1, 10.2). Isotomous branching up to Ax2.
IIBr2 with first pinnule (Fig. 10.1, 10.2); subsequent pinnules arise from alternative sides of successive brachials, except for hipozygals that lack pinnules; uniserial. Pinnulars subrectangular (=cylindrical) to subquadrate; lateral smooth unornamented.
Column slender (Wcolumnal/Wcup = 0.36); latera ornamented by middle ridge with fine tubercles (Figs. 10.3, 10.5); straight; heteromorphic; divided at least into two portions: proxistele and dististele.
Proxistele narrowing distally (maximum width ~3.7 mm); columnals reduced in height (H << 1 mm), almost flat, columnal outline pentastellate (x/y = 0.45; see categories in Simms, Reference Simms1989, p. 6; Fig. 10.1, 10.2); central circular axial canal large (7% of columnal width); triangular crenularium with 14 crenullae per interradius.
Dististele (H ≥ 11.6 mm); columnals pentastellate; interradius acuminated, three fine tubercles on its tip (Fig. 10.3); five internodals (i.e., six ossicles per noditaxis; short noditaxis); nodals slightly higher than internodals (Hnodal = 0.7 mm; Hinternodal = 0.5 mm); column articulation symplexy.
Cirrus sockets five in number per nodal; as high as the nodal, moderate sized, 32.3% of nodal width; distinct transverse ridge that divides laterally into two short oblique ridges (synarthry); central circular axial canal with raised lumen, represents 5.8% of columnal width. Cirri undivided; curved downwards. Cirrals rectangular, smooth; sinuous articular facet; widest cirrus with nine cirrals (W = 4 mm; Fig. 10.3, 10.5); tapering distally; perpendicular to column, at least initially, and then amply curved downwards.
Etymology
After Pehuenches County, where Mina La Quemada fossil locality is placed and holotype was collected.
Measurements
See Table 1.
Remarks
Isocrinus (Chladocrinus) pehuenchensis n. sp. is only known from one specimen, the holotype, and although this is not an ideal situation, the quality of its preservation and the presence of the aboral cup, proximal arm branching, and column makes this example one of the most complete specimens among Cretaceous isocrinids. It is superficially similar to Isocrinus (C.) covuncoensis n. sp., both are isocrinids, but I. (C.) pehuenchensis n. sp. has many important morphological differences with the former (see Table 1). The most important of which include: (1) different H/W cup proportion, being larger in I. (C.) covuncoensis n. sp.; (2) different Wdististele/Wcup being larger in I. (C.) covuncoensis n. sp.; (3) smaller Wcolumn in I. (C.) pehuenchensis n. sp.; (4) number of secundibrachials, being 8–9 in I. (C.) pehuenchensis n. sp. and 7 in I. (C.) covuncoensis n. sp.; (5) columnal outline, being pentastellate in I. (C.) pehuenchensis n. sp. and pentalobate in I. (C.) covuncoensis n. sp.; (6) column ornamentation, being smooth in I. (C.) covuncoensis n. sp. and decorated in I. (C.) pehuenchensis n. sp.; and (7) column articulation symplexy in contrast to symplexy and cryptosymplexy in I. (C.) covuncoensis n. sp. The combination of quantitative and qualitative characters may warrant the true separation of both species. In addition, they have a different stratigraphic occurrence, with I. (C.) covuncoensis n. sp. older from upper Valanginian beds and I. (C.) pehuenchensis n. sp. younger from upper Hauterivian beds, but also were recorded from different facies and thus different paleoenvironmental settings are envisaged (see discussion).
Isocrinus (Chladocrinus) basaltiformis (Miller, Reference Miller1821, p. 62–63, pl. II), originally described from the Lower Jurassic (Pliensbachian) of England, differs by having pentagonal columnals, latera heavily ornamented, and larger number (up to 20) of internodals (see Klikushin, Reference Klikushin1987, fig. 1A–C; Hess and Messing, Reference Hess, Messing and Selden2011, fig. 26, 1a–c).
Isocrinus nicoleti (Desor, Reference Desor1845), originally described from the Middle Jurassic (Bathonian) of France, is similar to I. (C.) pehuenchensis n. sp. in terms of cup morphology, number of primibrachials, number of internodals, and columnal morphology; but it differs by having a wider column (maximum 6 mm) and smooth latera of columnals (see de Loriol, Reference de Loriol1887, p. 165, pl. 154–161; Hess, Reference Hess2012, fig. 4).
Isocrinus aptiensis Taylor, Reference Taylor1966, from the Aptian of the east coast of Alexander Island, Antarctica is differentiated from I. (C.) pehuenchensis n. sp. by having a larger number of IIBr (13–15), higher than wide brachials and unornamented column (Taylor, Reference Taylor1966, p. 4, fig. 6a, b, d, f).
Isocrinus australis Moore, Reference Moore1870 from the late Aptian of Australia is differentiated essentially by having a larger number of IIBr (13). Isocrinus parvus (Howchin, Reference Howchin1921, pl. 1, fig. 3) from the Aptian–Albian of Southern Australia is difficult to be compared because it corresponds to finials, which are absent in I. (C.) pehuenchensis n. sp. However, as stated above, this Australian species should be taken with uncertainty for its incompleteness.
Isocrinus (Chladocrinus) hanaii Oji, Reference Oji1985, from the upper Aptian of the Hiraiga Formation, Miyako Group, Northeast Japan is differentiated by having a larger number of IIBr (12–13), pentagonal to subpentagonal columnals, and larger number of internodals (7–8, rarely 9).
Discussion and conclusions
The presence of stalked crinoids in the Jurassic–Cretaceous of South America has been long recognized, but this is the first contribution that describes two species on the basis of complete specimens, and thus for the first time a discussion on taphonomy, facies relationships, and paleoecology can be done. The newly described species belong to different sedimentary settings and ages, even though they come from the same formation. Both are recorded in offshore facies, but I. (Chladocrinus) covuncoensis n. sp. belongs to a high-energy sandy setting while I. (Chladocrinus) pehuenchensis n. sp. belongs to a low-energy muddy setting.
Isocrinus (C.) covuncoensis n. sp. was recorded mainly in crinoid beds at La Cresta, associated with a particular sedimentary setting dominated by large subaqueous dunes deposited by strong unidirectional currents that evolved in an offshore shelf, below the fair-weather wave base. The excellent preservation of the key crinoid bed can be considered as a local Konservat Lagerstätte. Crinoids that dwelled (at least) at the dune toes and interdunes were buried suddenly by the acceleration of the lee face migration (see Veiga and Schwarz, Reference Veiga and Schwarz2017). It was a rather small species with a maximum height of 148 mm in the adult The highly mobile sandy substrate affected by strong currents suggests that the slightly curved distal part of the column along with terminal cirri laid horizontally on the bottom, instead of being erected vertically, and based on modern relatives, it can be interpreted that the species was capable of crawling and local relocation if needed. Regenerated arms or predation marks were not detected, indicating that predation pressure was not an important issue in the populations of the species.
Subaqueous dunes were common in the past, but also have been recorded at present times (Testa and Bosence, Reference Testa and Bosence1999). Cariocrinus andreae beds from the lower Hauptrogenstein, Bajocian of Switzerland were interpreted as deposited in subaqueous dune-dominated settings (see Hess, Reference Hess, Hess, Ausich, Brett and Simms1999, fig. 211, 212). Complete specimens have been found in a larger area than that of the present study, at about 30 sites, and crinoid density is much higher, reaching 400 specimens per square meter. One striking similarity is that the best-preserved specimens occur in the lowest bedding plane of the recorded set of crinoid beds, which is very similar to the position of the key crinoid bed described here (Fig. 3.2–3.4). This would indicate that exceptional preservation is recorded just after a prolonged pause in the lee face migration of dunes and that its reactivation, including avalanching processes, may have buried suddenly the living crinoids that settled on the dune trough.
Notably, I. (C.) covuncoensis n. sp. conformed a monospecific crinoid association, not associated with any other megafossils, except for some trace fossils (e.g., Ophiomorpha isp.) restricted to the sandstones, which are otherwise common and abundant in the Agrio Formation. Only very small cemented oysters were detected attached to some columns. It seems that the subaqueous dune setting was not hospitable for settlement of benthic invertebrate larvae other than crinoids, small oysters, and possibly decapods. Perhaps the strong current activity and highly mobile substrate might have acted as limiting factors. Recent subaqueous dune fields of north-east Brazil, 13–20 m in depth, are colonized by dense seaweed, including red and coralline algae in the dune, but they were not detected either in the present study (Testa and Bosence, Reference Testa and Bosence1999).
Isocrinus (C.) pehuenchensis n. sp. is based on a single specimen recorded in a calcareous silty concretion at Mina La Quemada. This locality is distally placed in the Agrio Formation shelf and predominantly has records of basinal to distal offshore facies (see Legarreta and Uliana, Reference Legarreta and Uliana1991), supporting a relatively deeper setting than that envisaged for I. (C.) covuncoensis n. sp. The concretion level is associated with offshore fine-grained deposits settled from suspension on a low-energy sea floor. This succession is generally barren of benthic elements, but frequent in ammonoid levels. The excellent state of preservation of the crinoid in this case was due to an early cementation process by carbonate that enhanced the lithification of the concretion formed during a sedimentation pause just after the specimen was entombed, and not by sudden burial, as happened for I. (C.) covuncoensis n. sp. The crinoid could have acted as the nucleus for carbonate precipitation. Even though this is an isolated record, it shows that isocrinids established in the deeper offshore of the Agrio Formation on a muddy sea floor under low-energy conditions. The abundance of crinoid specimens at this level seems to be much less than that of the former case, suggesting that further colonization by benthic elements (including crinoids) of low-energy muddy substrates was hampered by some limiting environmental factor/s, such as hardened substrate consistency and/or low benthic oxygen level.
The studied Valanginian–Hauterivian stalked crinoids from the Neuquén Basin are recorded from offshore facies (=outer shelf, no storm influence) that represent deep marine settings, while shallower facies (nearshore to inner shelf) are devoid of crinoids. In our studied case, the deep water distribution of crinoids cannot be explained by predation pressure only, because there is no predatory evidence, not only on crinoids (absence of arm regeneration and predation marks) but on the overall mollusk and echinoid faunas reported from the Agrio Formation (see Aguirre-Urreta et al., Reference Aguirre-Urreta, Lazo, Griffin, Vennari, Parras, Cataldo, Garberoglio, Luci, Leanza, Arregui, Carbone, Danieli and Vallés2011). Therefore, the absence of crinoids in nearshore facies of the Agrio Formation may be better explained as a combination of taphonomic biases and/or environmental limiting factors, but not due only to predation pressure. This is in line with the proposal of Whittle et al. (Reference Whittle, Hunter, Cantrill and McNamara2018) that isocrinid migration from shallow to deep water did not occur at the same time all over the world and with the fact that the youngest crinoids in nearshore facies persisted from the Cretaceous to the Eocene/Oligocene boundary.
Acknowledgments
We are grateful to P. Chafrat and C. Brito for their help while visiting the collection of the Museo Patagónico de Ciencias Naturales (General Roca, Río Negro, Argentina). We are indebted with M.B. Aguirre-Urreta (IDEAN, UBA-CONICET, Buenos Aires, Argentina), C.S. Cataldo (IDEAN, UBA-CONICET, Buenos Aires, Argentina), and P.F. Rawson (Hull University, UK) for sharing important field observations from Mina La Quemada locality and useful references. T.K. Baumiller (University of Michigan, Ann Arbor, USA), E. Olivero (CADIC, Ushuaia, Argentina), A. Pisera (Polish Academy of Sciences, Warsaw, Poland), and J. Stilwell (Monash University, Clayton, Australia) are thanked for their help regarding queries and bibliography. Comments and suggestions by A. Hunter (University of Cambridge) and T. Oji (University Museum, Nagoya University) are especially appreciated. This work was supported in part by UBACyT grant 2013-2016 “Grupo en Formación” and ANPCyT PICT 2015-1381 grant. This is the contribution R-293 of the Instituto de Estudios Andinos “Don Pablo Groeber” (IDEAN).











