



Introduction
Although the Afro-Arabian landmass has played a pivotal role in the evolutionary history of early placental mammals (Werdelin and Sanders, Reference Werdelin and Sanders2010), the sub-Saharan Paleogene fossil record remains very scarce, particularly in West Africa, where relatively little paleontological work has been carried out. However, the potential to discover Eocene placentals was demonstrated over half a century ago when a fragmentary proboscidean molar was described from the middle Eocene (Lutetian) of M'Bodione Dadere, Senegal, by Gorodiski and Lavocat (Reference Gorodiski and Lavocat1953). These authors compared this molar with those of Moeritherium, a genus from the late Eocene–early Oligocene period. Interestingly, they correctly noticed that the M'Bodione Dadere proboscidean was significantly smaller than all known Moeritherium species. Despite this substantial morphological difference, Gorodiski and Lavocat (Reference Gorodiski and Lavocat1953) were reluctant to establish a new taxon based on this fragmentary molar, judging that Paleogene proboscideans were too poorly known to substantiate the definition of a new taxon. At the time, early proboscidean evolution was documented by just four genera, known only from the late Eocene (Moeritherium and Barytherium) and early Oligocene (Moeritherium, Phiomia, and Palaeomastodon) of the Fayum, Egypt (e.g., Andrews, Reference Andrews1906; Osborn, Reference Osborn1936). After more than 50 years of field research, the fossil record of stem proboscideans is now much better known (Gheerbrant and Tassy, Reference Gheerbrant and Tassy2009; Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa, Delmer, Werdelin and Sanders2010). Considerable progress has been made in several other parts of Africa (Morocco, Algeria, Libya, Kenya, and Ethiopia) and the Arabian Peninsula (Oman and Saudi Arabia), notably at the late Paleocene–early Eocene transition (Gheerbrant et al., Reference Gheerbrant, Sudre, Cappetta and Bignot1998a, Reference Gheerbrant, Sudre, Tassy, Amaghzaz, Bouya and Iarochene2005, Reference Gheerbrant, Bouya and Amaghzaz2012; Gheerbrant, Reference Gheerbrant2009) and, to a lesser degree, at the late Eocene–early Oligocene transition (Delmer et al., Reference Delmer, Mahboubi, Tabuce and Tassy2006; Delmer, Reference Delmer2009; Adnet et al., Reference Adnet, Cappetta and Tabuce2010; Jaeger et al., Reference Jaeger, Salem, Elbakai and Abuturruma2012; Seiffert et al., Reference Seiffert, Nasir, Al-Harthy, Groenke, Kraatz, Stevens and Al-Sayigh2012; Pickford, Reference Pickford2015; Al-Kindi et al., Reference Al-Kindi, Pickford, Al-Sinani, Al-Ismaili, Hartman and Heward2017).
Despite these fundamental advances, some unresolved problems persist. Among them, the origin and phylogenetic position of the genus Moeritherium remain uncertain. Two phylogenetic scenarios have been recently proposed for the relationships of Moeritherium within Proboscidea. First, according to Seiffert et al. (Reference Seiffert, Nasir, Al-Harthy, Groenke, Kraatz, Stevens and Al-Sayigh2012), Moeritherium could have a basal position in the phylogeny, near Daouitherium and Numidotherium, two stem proboscideans from the Ypresian of Morocco and Algeria, respectively. Second, Moeritherium has a more elevated position in the phylogeny, being considered as the sister group of the clade Deinotheriidae-Elephantiformes (Seiffert, Reference Seiffert2007; Delmer, Reference Delmer2009; Gheerbrant and Tassy, Reference Gheerbrant and Tassy2009; Ferretti and Debruyne, Reference Ferretti and Debruyne2011; Cooper et al., Reference Cooper, Seiffert, Clementz, Madar, Bajpai, Hussain and Thewissen2014). In any case, these contradictory scenarios illuminate longstanding uncertainties regarding the transition from early proboscideans to elephantiforms (palaeomastodonts and elephantoids). Particularly, the homology between the lower incisors of early proboscideans and the ever-growing lower tusks of elephantiforms is still debated (Delmer, Reference Delmer2009; Jaeger et al., Reference Jaeger, Salem, Elbakai and Abuturruma2012; Al-Kindi et al., Reference Al-Kindi, Pickford, Al-Sinani, Al-Ismaili, Hartman and Heward2017).
Current data suggest that the transition from early proboscideans to elephantiforms occurred during the end of the Eocene (Priabonian; Fig. 1a), but a simple examination of the Paleogene proboscidean fossil record reveals geographical and temporal sampling biases in our understanding of the evolutionary history of proboscideans. Earliest proboscideans are documented only from Moroccan deposits, while other Paleogene species are known from a few African (e.g., Algeria, Egypt, Libya, Ethiopia, and Kenya) and Arabian (Saudi Arabia and Oman) sites. Most important, this history is largely truncated by a hiatus encompassing the end of the Ypresian to the early Priabonian, so that the proboscidean fossil record is virtually unknown during most of the Eocene (Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa, Delmer, Werdelin and Sanders2010; Fig. 1a). It is noteworthy that the group is absent from the rich faunas from Gour Lazib (late early or early middle Eocene, Algeria), Chambi (late early or early middle Eocene, Tunisia), and Namibia (?Lutetian–Bartonian) (e.g., Pickford et al., Reference Pickford, Senut, Morales, Mein and Sanchez2008; Adaci et al., Reference Adaci, Bensalah, Tabuce, Mebrouk, Marivaux, Otero, Zaoui, Benyoucef and Mahboubi2016). Proboscideans are also absent from the only two unambiguous middle Eocene continental mammalian sites in Africa, the faunas of Aznag in Morocco (Tabuce et al., Reference Tabuce, Adnet, Cappetta, Noubhani and Quillevéré2005) and Mahenge in Tanzania (Kaiser et al., Reference Kaiser, Ansorge, Arratia, Bullwinkel, Gunnell, Herendeen, Jacobs, Mingram, Msuya and Musolff2006). By contrast, the only two ?mid-Eocene localities that yielded proboscidean remains were from West Africa. Arambourg et al. (Reference Arambourg, Kikoine and Lavocat1951) described Moeritherium sp. from two isolated molars discovered from marine sediments near In Tafidet, Mali (middle or late Eocene; see Gheerbrant et al., Reference Gheerbrant, Sudre, Cappetta and Bignot1998a, fig.6c; Delmer et al., Reference Delmer, Mahboubi, Tabuce and Tassy2006; O'Leary et al., Reference O'Leary, Roberts, Bouare, Sissoko and Tapanila2006). Gorodiski and Lavocat (Reference Gorodiski and Lavocat1953) described ?Moeritherium sp., also from middle Eocene sediments, on the basis of the Senegalese molar here studied. Since its initial description, this molar has never been figured; Domning et al. (Reference Domning, Ray and McKenna1986), Gheerbrant et al. (Reference Gheerbrant, Sudre, Cappetta and Bignot1998a), and Delmer et al. (Reference Delmer, Mahboubi, Tabuce and Tassy2006) briefly discussed its morphology and concluded that it was too small to belong to a Moeritherium species.
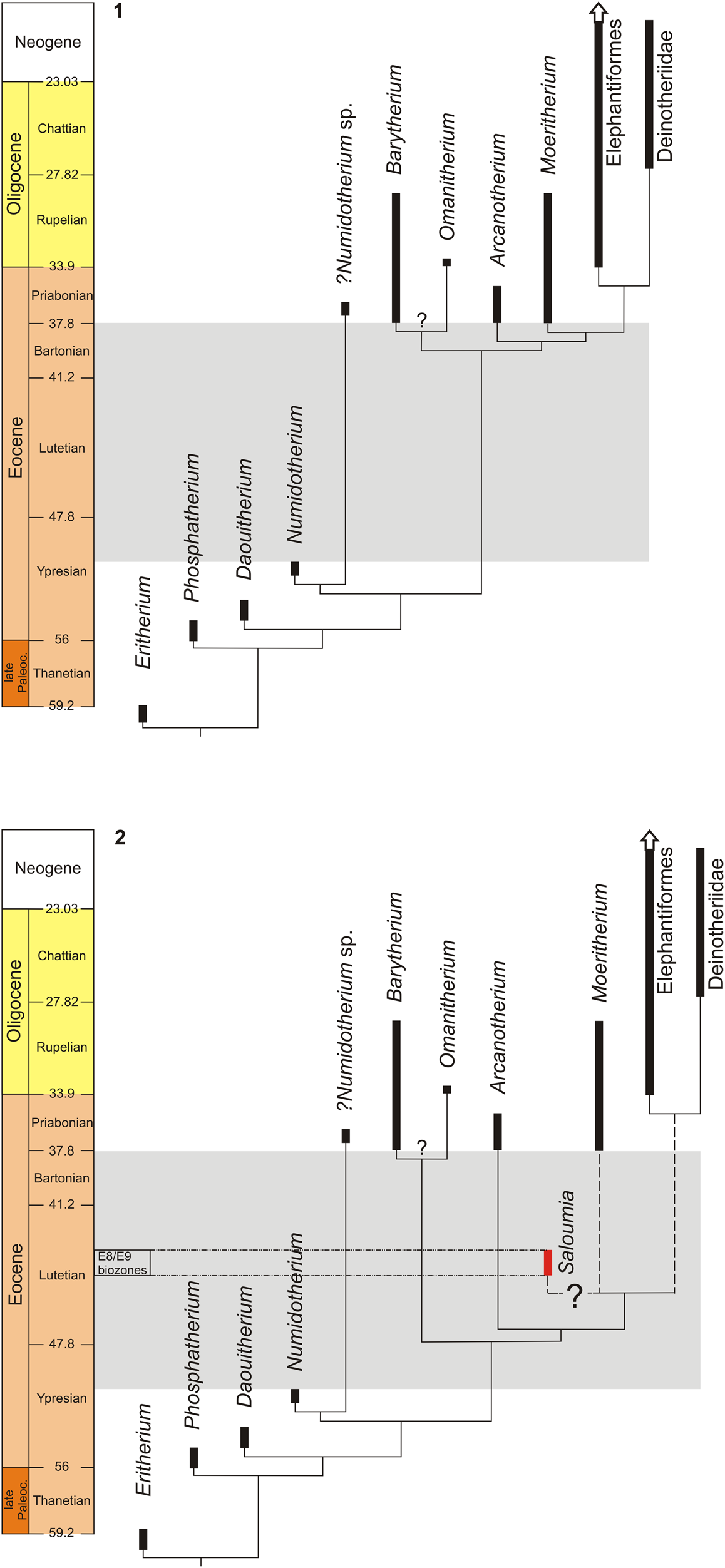
Figure 1. Phylogeny of Paleogene proboscideans based on the cladistics analysis proposed by Delmer (Reference Delmer2009), Gheerbrant and Tassy (Reference Gheerbrant and Tassy2009), Gheerbrant et al. (Reference Gheerbrant, Bouya and Amaghzaz2012), and Seiffert et al. (Reference Seiffert, Nasir, Al-Harthy, Groenke, Kraatz, Stevens and Al-Sayigh2012); the gray zone shows the important gap in their fossil record. (1) The question mark indicates that Omanitherium could be closer to Arcanotherium and Numidotherium than to Barytherium as proposed by Pickford (Reference Pickford2015) and Al-Kindi et al. (Reference Al-Kindi, Pickford, Al-Sinani, Al-Ismaili, Hartman and Heward2017); (2) possible phylogenetic relationships of Saloumia gorodiskii n. gen. n. sp. from the mid-Lutetian of Senegal. Following this scenario, Saloumia could represent a suprageneric taxon close to the clade including Moeritherium and Elephantiformes.
To better document this peculiar Eocene proboscidean, we organized systematic prospecting for fossil vertebrates at and around M'Bodione Dadere in 2010 and 2011. Although we discovered a rich marine fauna represented by numerous sharks and rays, no additional mammalian specimens have been discovered except an indeterminable minute mammalian tooth fragment. Considering the difficulties in discovering new specimens of proboscideans in the rare Eocene Senegalese outcrops, we reassess the Moeritherium-like molar from M'Bodione Dadere. Both its provenance (West Africa) and age (Lutetian) make this tooth a pivotal asset to better understand the early history of proboscideans as it fills an important gap in their paleogeographic and chronological fossil record.
Geological setting
M'Bodione Dadere village is located north of the town of Kaolack in the Sine-Saloum region of the central-western part of Senegal (Fig. 2). In this flat area, outcrops are absent due to an important coverage of Quaternary to Recent sands (10–20 m), which unconformably overlie Paleogene to Pliocene levels. Access to a detailed stratigraphy and to Paleogene deposits is only possible thanks to drill cores and water-well cuttings. Fortuitously, sediments excavated from levels attributed to two Lutetian–Bartonian shallow marine formations yielded vertebrate fossils.

Figure 2. Geographic position, stratigraphic position, and regional correlations of M'Bodione Dadere site that yielded the holotype of Saloumia gorodiskii n. gen. n. sp.
From the water well of Tiavandou, South to Kaolack (Fig. 2), nummulitic limestone attributed to the Taïba Formation (late Lutetian to early Bartonian; Roger et al., Reference Roger, Banton, Barusseau, Castaigne, Comte, Duvail, Nehlig, Noël, Serrano and Travi2009a; Sarr, Reference Sarr2013) yielded several teeth, 18 vertebrae, and rib fragments of an archeocete identified as ‘Zeuglodon’ cf. Z. osiris by C. Arambourg (Elouard, Reference Elouard1966). More recently, in a revision of the African fossil record of archeocetes, Gingerich (Reference Gingerich, Werdelin and Sanders2010) suggested that these remains were too large to be attributed to Saghacetus osiris (Dames, Reference Dames1894) (the new name for Zeuglodon osiris), the size being more similar to Dorudon atrox (Andrews, Reference Andrews1906), another archeocete well known from the early Priabonian of the Fayum. Unfortunately, after unfruitful research in the collections of the University of Dakar, the specimens seem to have been lost.
In the northern part of the Sine-Saloum region, other water-well cuttings attributed to the middle Eocene Lam Lam Formation yielded vertebrate remains. The most recent example is the discovery of an actinopterygian maxilla attributed to an amiid (O'Leary et al., Reference O'Leary, Sarr, Malou, Sow, Lepre and Hill2012) in Tewrou Poram, North of the town of Gossas (Fig. 2). The earliest discovery of vertebrates in the Lam Lam Formation corresponds to that of the M'Bodione Dadere proboscidean. On 24 August 1952, Alexandre Gorodiski was in charge of geological mapping in the area of Thiès (on behalf of the Federal Direction of Mines and Geology of French West Africa) when he discovered the proboscidean molar from cuttings of the 20 m deep well of M'Bodione Dadere; the molar was described by R. Lavocat (Gorodiski and Lavocat, Reference Gorodiski and Lavocat1953; see also Lavocat, Reference Lavocat1955). By chance, A. Gorodiski also collected samples of the coquina limestone from the well cuttings. This revealed two new fragments of mammal teeth showing ‘condylarth’ affinities (Gevin et al., Reference Gevin, Mongereau and Truc1975; Sudre, Reference Sudre1979); these indeterminable specimens, housed in the collections of the University of Montpellier, also evoke herodotiine macroscelideans by size and gross morphology (R. Tabuce, personal observation).
To reconstruct the geological context of the M'Bodione Dadere proboscidean, we compared the stratigraphic sequence established by Gorodiski and Lavocat (Reference Gorodiski and Lavocat1953) with nearby logs from drill cores (Gossas, Ngolothie, and Kaolack; Fig. 2) and with our direct observations of recent water-well cuttings located at Sanghaye Bele village (11 km southeast of M'Bodione Dadere; Fig. 2). At M'Bodione Dadere and Sanghaye Bele, the fossiliferous deposits attributed to the Lam Lam Formation correspond to those of a coquina limestone and a yellow to white clayey limestone alternating with marls; at M'Bodione Dadere these deposits are capped by phosphate nodules. The invertebrate fauna is composed of echinoids and mollusks, among them the lamellibranch Ostrea meunieri, Douvillé, Reference Douvillé1920, which is known from the ‘Lutetian’ of Senegal (Tessier, Reference Tessier1952; see Roger et al., Reference Roger, Barusseau, Duvail, Nehlig, Noël and Serrano2009b, p. 16). Well samples from M'Bodione Dadere and Sanghaye Bele did not provide any characteristic microfauna, but equivalent levels from the Ngolothie borehole near Kaolack (Fig. 2) yielded the planktonic foraminifera Globigerinatheka gr. index Finlay, Reference Finlay1939, Cassigerinelloita amekiensis Stolk, Reference Stolk1965, and Pseudohastigerina micra (Cole, Reference Cole1927), which characterize E8 to E11 biozones (middle Lutetian to early Bartonian; Sarr, Reference Sarr2013). North of the Senegalese basin, the base of the Lam Lam Formation (Pallo limestone and phosphate clays) is E8 in age (middle Lutetian; presence of Hantkenina aragonensis Nuttall, Reference Nuttall1930; Flicoteaux, Reference Flicoteaux1975; Roger et al., Reference Roger, Banton, Barusseau, Castaigne, Comte, Duvail, Nehlig, Noël, Serrano and Travi2009a), whereas the top of the formation (Lam Lam marls) yielded a rich planktonic foraminifera association (Truncorotaloides rohri Brönnimann and Bermudéz [Reference Brönnimann and Bermúdez1953], Acarinina topilensis [Cushman, Reference Cushman1925], Globigerapsis sp., G. index, Acarinina collactea [Finlay, Reference Finlay1939], Pseudoglobigerinella bolivariana [Petters, Reference Petters1954], Acarinina pentacamerata [Subbotina, Reference Subbotina1947], and Hantkenina spp.), which indicates E9 biozone (middle Lutetian; Flicoteaux, Reference Flicoteaux1974; Roger et al., Reference Roger, Barusseau, Duvail, Nehlig, Noël and Serrano2009b). These regional correlations indicate a secure middle Lutetian age (E8–E9 biozones, ca. 44 Ma, Vandenberghe et al., Reference Vandenberghe, Hilgen, Speijer, Gradstein, Ogg, Schmitz and Ogg2012) for the M'Bodione Dadere proboscidean.
Material and methods
Repositories and institutional abbreviations
The holotype MNHN.F.MCA 1 and unique specimen of Saloumia gorodiskii is housed in the collections of the Museum National d'Histoire Naturelle, Paris (MNHN). A computed tomography (CT) scan of MNHN.F.MCA 1 was acquired at the Montpellier Rio Imaging (MRI) micro-CT imaging station Skyscan 1076 (Montpellier, France). Image segmentation and three-dimensional (3D) rendering were performed using manual segmentation with Avizo 7.1 (VSG). A 3D reconstruction of MNHN.F.MCA 1 was deposited in MorphoMuseum (https://doi.org/10.18563/journal.m3.104). Other institutional abbreviations: AMNH (American Museum of Natural History, New York, USA) and NHMUK (Natural History Museum, Department of Palaeontology, London, UK).
Systematic paleontology
Class Mammalia Linnaeus, Reference Linnaeus1758
Order Proboscidea Illiger, Reference Illiger1811
Family incertae sedis
Genus Saloumia new genus
Type species
Saloumia gorodiskii n. gen. n. sp., only species.
Diagnosis
As for the type species, by monotypy.
Etymology
From the coastal river Saloum, which flows south of the village of M'Bodione Dadere.
Remarks
The specimen described here is identified as a proboscidean on the basis of its relative large size, quadrituberculy, wrinkled enamel, small parastyle, well-developed lingual cingulum, blunt paraconule mesiolabially located to the protocone, and the lack of a centrocrista. The particular combination of these dental characters is unknown is other African ‘ungulates’ from the Paleogene (hyraxes, embrithopods, sirenians, and ‘condylarths’).
Saloumia gorodiskii new species
Figs. 3, 4

Figure 3. Saloumia gorodiskii n. gen. n. sp., MNHN.F.MCA 1, right upper molar, in (1) occlusal (stereo pair), (2) oblique mesial, and (3) oblique lingual views.

Figure 4. Saloumia gorodiskii n. gen. n. sp., MNHN.F.MCA 1, right upper molar, in occlusal view (3D rendering from X-ray microtomography).
Holotype
MNHN.F.MCA 1, right upper molar, which rests on indurated coquina limestone.
Diagnosis
Saloumia gorodiskii differs from Eritherium and Phosphatherium by larger size, wrinkled enamel, strong lingual cingulum, and no centrocrista; its morphology also differs from Phosphatherium by a pronounced bunodonty. Saloumia gorodiskii differs from Moeritherium ssp., Phiomia, and Palaeomastodon by smaller size, more bunodont and not bunolophodont dental pattern, relatively lower crown, and more labial positions of the protocone and hypocone.
Occurrence
M'Bodione Dadere (14°20.706′N, 16°11.151′W), Lam Lam Formation, middle Lutetian, planktonic foraminiferal biozones E8–E9, ca. 44 Ma.
Description
MNHN.F.MCA 1 is a low-crowned and incompletely preserved upper molar, missing its distal part (length > 10.7 mm; width = 12.3 mm; Figs. 3, 4). The apexes of the four principal cusps (paracone, metacone, protocone, and hypocone) were broken away (‘scalped’), possibly before fossilization; there is no significant wear on these cusps. Only the mesial flanks of the metacone and hypocone are preserved. The hypocone is relatively centrally positioned and as large as the protocone and certainly wider than the metacone. The paracone and protocone are fully bunodont and connected at their base but without a high loph connecting them. However, this loph, if it was incipiently developed, may have been erased as the apexes of protocone and paracone were broken away. A large paraconule, whose apex is also broken away, is underlined by dental wear facets; it is closely appressed to the mesiolabial flank of the protocone; the paraconule also connects to the mesial cingulum. There is no trace of a metaconule. The shallow valley between the paracone and metacone is deeper and larger than that between the protocone and hypocone. The transverse valley is thus poorly defined. The cingulum is continuous from the labial border of the paracone to the lingual face of the teeth; it is particularly well developed, forming an entostyle located lingual to the transverse valley. The cingulum is also inflated labially to the transverse valley, forming an ectostyle. The enamel is wrinkled, especially on the paracone and lingually to the protocone.
Etymology
The species is named after the geologist Alexandre Gorodiski, who discovered the fossiliferous locality and the holotype.
Remarks
The thick enamel, the lack of a well-defined ectoflexus, and the parastyle, which is not enlarged and not mesially projected but strongly appressed to the paracone, precludes that MNHN.F.MCA 1 is a deciduous premolar. Likewise, the probability that MNHN.F.MCA 1 corresponds to a fully molariform premolar is also excluded because the molarization of premolars in proboscideans is documented only during the mid–early Oligocene (in the elephantiforms Phiomia and Palaeomastodon).
Systematic attribution and discussion
Gorodiski and Lavocat (Reference Gorodiski and Lavocat1953) attributed MNHN.F.MCA 1 to ?Moeritherium sp. in view of its quadritubercular dental pattern and the morphology of its cingulum. However, they insisted that ‘it would not be prudent to exclude the possibility that this specimen may belong to another [new] primitive bunodont genus, especially because main cusps appear to have been more obviously bunodont than those of known Moeritherium species’ (Gorodiski and Lavocat, Reference Gorodiski and Lavocat1953, p. 316). During the 1950s, Paleogene proboscideans were documented only by Moeritherium, Barytherium, Phiomia, and Palaeomastodon. The genus Moeritherium was then considered as ancestral among proboscideans. Today, the phylogenetic position of Moeritherium remains debated. Such an uncertainty is surprising considering that dental, cranial, and postcranial anatomy of Moeritherium has been well known for more than a century. The genus is particularly well documented in the Fayum, where it is known by two species. The consensus view consists in attributing all specimens from the Gebel el Qatrani Formation (early Oligocene) to Moeritherium trigodon Andrews, Reference Andrews1904, and those from the Qasr el Sagha Formation (late Eocene) to M. lyonsi (Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa, Delmer, Werdelin and Sanders2010; but see Delmer et al., Reference Delmer, Mahboubi, Tabuce and Tassy2006).
These two Fayumian moeritheres are characterized by a bunolophodont dental pattern. The oldest attested Moeritherium species, M. chehbeurameuri Delmer et al., Reference Delmer, Mahboubi, Tabuce and Tassy2006, corresponds to a smaller and more lophodont taxon; it was described from the Priabonian of Bir El Ater, Algeria (Delmer et al., Reference Delmer, Mahboubi, Tabuce and Tassy2006). This species was also listed from the ?late Eocene locality of Dur At-Talah, Libya (Jaeger et al., Reference Jaeger2010, Reference Jaeger, Salem, Elbakai and Abuturruma2012) and from the late Eocene locality of Birket Qarun-2, Fayum (W. Sanders, personal communication, 2019; see also Seiffert et al., Reference Seiffert, Bown, Clyde, Simons, Fleagle and Gilbert2008; Seiffert, Reference Seiffert, Werdelin and Sanders2010).
Saloumia evokes all these Moeritherium species by three derived characters: wrinkled enamel, the lack of centrocrista, and the well-developed lingual cingulum that was considered as a diagnostic feature of the genus (Figs. 3, 4). As a result, these three dental characters could support a sister-group relationship between Saloumia and Moeritherium. However, the association of a convolute and a distocrista (see Delmer et al., Reference Delmer, Mahboubi, Tabuce and Tassy2006) distally to the hypocone, which is also a diagnostic character of the upper molars of Moeritherium, cannot be described in Saloumia due to the poor preservation of the holotype. Saloumia also evokes the Fayumian Moeritherium species by quadrituberculy, small parastyle, blunt paraconule mesiolabially located to the protocone, and blunt crest joining the protocone to the parastyle via the paraconule. However, these traits are primitive among proboscideans and thus do not support a close relationship between Saloumia and Fayumian Moeritherium species. Moreover, Saloumia is half as large as Moeritherium trigodon and M. lyonsi and differs from these two species by having a relatively lower crown and a more labial position of both the protocone and hypocone and by the occurrence of a labial cingulum. We note, however, that M. ‘gracile’ Andrews, Reference Andrews1902, which is a synonym of M. lyonsi Andrews, Reference Andrews1901 (see Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa, Delmer, Werdelin and Sanders2010; W. Sanders, personal communication, 2019), has a labial cingulum, according to Delmer et al. (Reference Delmer, Mahboubi, Tabuce and Tassy2006). Finally, as mentioned by Gorodiski and Lavocat (Reference Gorodiski and Lavocat1953), the bunodonty of Saloumia differs markedly from the bunolophodonty of all the Fayumian moeritheres.
The bunolophodonty that characterizes the Fayumian Moeritherium species was long viewed as primitive for Proboscidea. As such, Moeritherium has long been recognized as close to the origin of the order (see also Seiffert et al., Reference Seiffert, Nasir, Al-Harthy, Groenke, Kraatz, Stevens and Al-Sayigh2012). By contrast, Moeritherium is often interpreted as a highly specialized genus (Court, Reference Court1994), more closely related to the deinotheriid–elephantiform clade than to the older genera Phosphatherium, Daouitherium, and Numidotherium (e.g., Delmer, Reference Delmer2009; Gheerbrant and Tassy, Reference Gheerbrant and Tassy2009; Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa, Delmer, Werdelin and Sanders2010; Ferretti and Debruyne, Reference Ferretti and Debruyne2011; Cooper et al., Reference Cooper, Seiffert, Clementz, Madar, Bajpai, Hussain and Thewissen2014). The true lophodonty of earliest Eocene genera was then considered as the primitive dental pattern of proboscideans (Gheerbrant et al., Reference Gheerbrant, Sudre, Tassy, Amaghzaz, Bouya and Iarochene2005). Following this hypothesis, the bunolophodonty of the Fayumian Moeritherium would derive from a true lophodonty. This result was strengthened by the description of the Priabonian lophodont M. chehbeurameuri.
Nevertheless, in our opinion, the question of the primitive dental pattern of moeritheriids is not resolved yet (see also Seiffert et al., Reference Seiffert, Nasir, Al-Harthy, Groenke, Kraatz, Stevens and Al-Sayigh2012). Indeed, Moeritherium sp. from the middle/late Eocene of In Tafidet, Mali (Arambourg et al., Reference Arambourg, Kikoine and Lavocat1951) is clearly more bunodont than its penecontemporaneous M. chehbeurameuri (Supplemental Data). As a result, we cannot exclude the possibility that Moeritherium derives from a bunodont incipiently bilophodont lineage morphologically close to Saloumia. Interestingly, the Malian Moeritherium also shares with Saloumia a minute ectostyle on the labial cingulum. This trait is primitive when compared to M. chehbeurameuri. The species from In Tafidet appears also primitive by comparison with M. chehbeurameuri and Saloumia by the occurrence of both a postparacrista and premetacrista (Supplemental Data); these crests constitute the centrocrista when associated with the ectostyle (this style is then named mesostyle, e.g., Gheerbrant et al., Reference Gheerbrant, Bouya and Amaghzaz2012). Accordingly, the primitive dental morphology of Moeritherium is yet uncertain. Interestingly, a bunodont incipiently bilophodont origin for this genus recently received significant support from the discovery of Eritherium, a late Paleocene proboscidean characterized by bunodont incipiently bilophodont molars (Gheerbrant, Reference Gheerbrant2009). Furthermore, Khamsaconus, another early Ypresian proboscidean from Morocco, displays the same bunodont incipiently bilophodont pattern. This dental pattern is today viewed as primitive for proboscideans (Gheerbrant et al., Reference Gheerbrant, Bouya and Amaghzaz2012).
Direct comparisons with the holotype of Khamsaconus, a very small DP4 and unique specimen, is however impossible. Note that the fragmentary bunodont–bunolophodont lower molar attributed to Khamsaconus by Gheerbrant et al. (Reference Gheerbrant, Sudre, Sen, Abrial, Marandat, Sigé and Vianey-liaud1998b) is now attributed to a basal hyracoid (see Gheerbrant et al., Reference Gheerbrant, Sudre, Tassy, Amaghzaz, Bouya and Iarochene2005, p. 286). Khamsaconus was never included in a phylogenetic analysis with both Eritherium and Phosphatherium, but Gheerbrant et al. (Reference Gheerbrant, Bouya and Amaghzaz2012) favored a close phylogenetic relationship among these three genera. Saloumia shares with Eritherium only primitive features: quadrituberculy, low crown, reduced parastyle (compared with Phosphatherium), blunt crest joining the protocone to the parastyle, and labial position and larger size of the protocone and hypocone relative to the paracone and metacone, respectively. Most important, Saloumia differs from Eritherium and Phosphatherium by four derived characters: larger size, wrinkled enamel, lack of a continuous centrocrista, and strong lingual cingulum. Saloumia further differs from Eritherium by a more pronounced bunodonty.
Phosphatherium and other Eocene genera (Daouitherium, Barytherium, and Arcanotherium) clearly depart from Saloumia by their true lophodonty. Among them, the undetermined species Daouitherium sp., known by a unique lower molar (Gheerbrant et al., Reference Gheerbrant, Sudre, Tassy, Amaghzaz, Bouya and Iarochene2005, p. 285), is of similar size. Although larger and fully lophodont, Arcanotherium evokes Saloumia by its moderately wrinkled enamel and the development of its lingual cingulum. Such derived characters are also present in the bunolophodont Phiomia and Palaeomastodon. These basal elephantiforms also share with Saloumia the lack of centrocrista, a strong lingual cingulum, and a variable occurrence of an ectostyle. The latter character is for example present in AMNH 13449 (Palaeomastodon beadnelli Andrews, Reference Andrews1901; see Osborn, Reference Osborn1936, p. 144) and NHMUK M8851 (Phiomia serridens Andrews and Beadnell, Reference Andrews and Beadnell1902; see Tassy Reference Tassy1982, p. 234). Palaeomastodon and Phiomia differ from Saloumia by a larger size, the lack of connection between the paraconule and the parastyle, a more lingual position of both the protocone and hypocone, and four main cusps of similar size. Finally, the tritoloph (the third loph, distal to the metaloph), which is diagnostic of Phiomia and Palaeomastodon, cannot be checked in Saloumia due to the poor preservation of the specimen.
To conclude, although poorly known, Saloumia appears sufficiently distinct from other early proboscideans to be considered a valid genus. Saloumia shares with stem early Paleogene proboscideans (Eritherium and Phosphatherium) only primitive features. Most important, three derived characters observed in Saloumia (wrinkled enamel, lack of centrocrista, and strong lingual cingulum) could indicate a possible affinity with either Moeritherium or basal elephantiforms. If correct, this hypothesis would push back the origin of the (Moeritherium–(Elephantiformes–Deinotheriidae)) clade well into the middle Eocene and generate considerable ghost lineages within proboscidean phylogeny (Fig. 1.2). However, this hypothesis remains highly speculative given our current level of knowledge on Saloumia. Its very pronounced bunodonty is well distinct from the bunolophodonty of Moeritherium and basal elephantiforms, so that Saloumia could also document an early experiment in dental diversity among Paleocene–Eocene proboscideans, with no direct relationships with later Paleogene proboscideans.
Acknowledgments
3D data acquisition was performed using the μCT facilities of the MRI platform member of the national infrastructure France-BioImaging supported by the French National Research Agency (ANR-10-INBS-04, ‘Investments for the future’) and of the Labex CEMEB (ANR-10-LABX-0004) and NUMEV (ANR-10-LABX-0020). This research is part of the PaleoSen project (www.paleosen.com) and was supported by the French ANR-PALASIAFRICA Program (ANR-08-JCJC-0017), PICS-CNRS, the International Exchange Scheme of the Royal Society, and the National Geographic Society's Global Exploration Fund (Northern Europe).
Accessibility of supplemental data
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.51c59zw4c.




