



Introduction
Irvingella Ulrich and Resser in Walcott, Reference Walcott1924 is a biostratigraphically significant Furongian (upper Cambrian) trilobite that was recorded from paleocontinents as diverse as Laurentia, Siberia, Kazakhstan, Baltica, Gondwana, Avalonia, North China, and South China (Geyer and Shergold, Reference Geyer and Shergold2000; Hong et al., Reference Hong, Lee and Choi2003; Peng et al., Reference Peng, Babcock, Zuo, Zhu, Lin, Yang and Qi2012; Westrop and Adrain, Reference Westrop and Adrain2016). In Laurentia, it occurs in both the upper Steoptoean Elvinia Zone (in association with Elvinia Walcott, Reference Walcott1924) and the basal Sunwaptan I. major Zone. The Irvingella-Elvinia assemblage has been reported from numerous locations throughout North America, including northwest Canada, British Columbia, Alberta, Montana, Wyoming, Nevada, Utah, Oklahoma, Texas, Missouri, Arkansas, Wisconsin, and Pennsylvania, in shelf to outer shelf facies (e.g., Chatterton and Gibb, Reference Chatterton and Gibb2016 and references therein; Westrop and Adrain, Reference Westrop and Adrain2016).
As is generally known, the Cambrian trilobites from the Precordillera of western Argentina have a clear Laurentian aspect (e.g., Rusconi, Reference Rusconi1954a, Reference Rusconi1962; Borrello, Reference Borrello and Holland1971; Bordonaro and Banchig, Reference Bordonaro and Banchig1995; Bordonaro, Reference Bordonaro2003a, Reference Bordonaro2016; Tortello, Reference Tortello2011, Reference Tortello2014). In the 1950s, Carlos Rusconi (Museum of Natural History of Mendoza) made enormous contributions to the paleontology of the Precordillera of Mendoza, including the discovery of Elvinia and Irvingella in an exotic limestone block of the San Isidro area. Rusconi (Reference Rusconi1953, Reference Rusconi1954a) pointed out both the biostratigraphic and the paleobiogeographic implications of this finding. He also studied the collection at the species level and proposed Elvinia obliquoensis Rusconi, Reference Rusconi1953, Irvingella jorusconii Rusconi, Reference Rusconi1953, I. platycephala Rusconi, Reference Rusconi1953, and I. obliquoensis Rusconi, Reference Rusconi1953.
In the absence of new findings, the Elvinia Zone in Argentina is known only from the collection mentioned above. Although the Rusconi species were not illustrated with sufficient detail, Palmer (Reference Palmer1965) studied replicas of the type specimens and identified Elvinia roemeri (Shumard, Reference Shumard1861) and Irvingella major, and since then, these assignments have been cited frequently in the literature (Westrop, Reference Westrop1986; Pratt, Reference Pratt1992; Bordonaro and Banchig, Reference Bordonaro and Banchig1995, Reference Bordonaro and Banchig1996; Bordonaro, Reference Bordonaro and Caminos1999, Reference Bordonaro2003a, Reference Bordonaro2003b; Hong et al., Reference Hong, Lee and Choi2003; Peng et al., Reference Peng, Babcock, Zuo, Zhu, Lin, Yang and Qi2012; Chatterton and Gibb, Reference Chatterton and Gibb2016).
Recently, Westrop and Adrain (Reference Westrop and Adrain2016) provided a comprehensive systematic revision of Irvingella and discussed helpful criteria for defining its species. Westrop and Adrain (Reference Westrop and Adrain2016) proposed new diagnoses for several taxa of North America and Australia, and demonstrated that the type species I. major Ulrich and Resser in Walcott, Reference Walcott1924, which has been reported throughout Laurentia and other Cambrian continents (Pegel, Reference Pegel2000; Hong et al., Reference Hong, Lee and Choi2003; Lazarenko et al., Reference Lazarenko, Gogin, Pegel, Sukhov, Abaimova, Egorova, Fedorov, Raevskaya, Ushatinskaya, Rozanov and Varlamov2008; Rushton and Weidner, Reference Rushton and Weidner2010; Peng et al., Reference Peng, Babcock, Zuo, Zhu, Lin, Yang and Qi2012), should actually be restricted to Walcott's poorly preserved type cranidia from Wisconsin, U.S.A.
In order to provide updated systematic information on the trilobites from the Elvinia Zone of Argentina, specimens of the Rusconi collection at the Museo de Ciencias Naturales y Antropológicas J.C. Moyano (Mendoza, Argentina) are fully revised herein. They represent three valid forms of Rusconi's Irvingella and, as noted by Palmer (Reference Palmer1965), the Laurentian species Elvinia roemeri. The best-preserved specimens allow comparison with material from North America and other parts of the world. The Irvingella species appear to be endemic to the southern Precordillera, but are related to species from central Texas and northwest Canada.
Geologic setting
The Cambrian of the Precordillera of Mendoza is characterized by open-marine limestone allochthonous blocks (olistoliths) of different sizes, ranging from centimeters to hundreds of meters in thickness, occurring chaotically within the Middle and Upper Ordovician shales of the Estancia San Isidro and Empozada formations (Bordonaro et al., Reference Bordonaro, Beresi and Keller1993; Keller, Reference Keller1999; Heredia and Beresi, Reference Heredia and Beresi2004). One of the most outstanding fossil-collecting areas is San Isidro, which comprises a number of hills and creeks ~15 km west of Mendoza City (Rusconi, Reference Rusconi1952, fig. 1; Fig. 1). There, the carbonate-dominated San Martín, San Isidro, and La Cruz olistoliths (= La Cruz Limestones sensu Keller, Reference Keller1999), as well as other minor exotic blocks, contain trilobites of middle and late Cambrian age. The oldest olistoliths yield trilobites of the Glossopleura Zone (early middle Cambrian), whereas the youngest blocks are assignable to the late Furongian Saukia Zone (e.g., Rusconi, Reference Rusconi1956a and references therein; Borrello, Reference Borrello and Holland1971; Shergold et al., Reference Shergold, Bordonaro and Liñán1995; Bordonaro and Banchig, Reference Bordonaro and Banchig1996; Keller, Reference Keller1999; Bordonaro, Reference Bordonaro2003a, Reference Bordonaro2014; Bordonaro and Fojo, Reference Bordonaro and Fojo2011; Tortello, Reference Tortello2014, Reference Tortello2017, Reference Tortello2018).
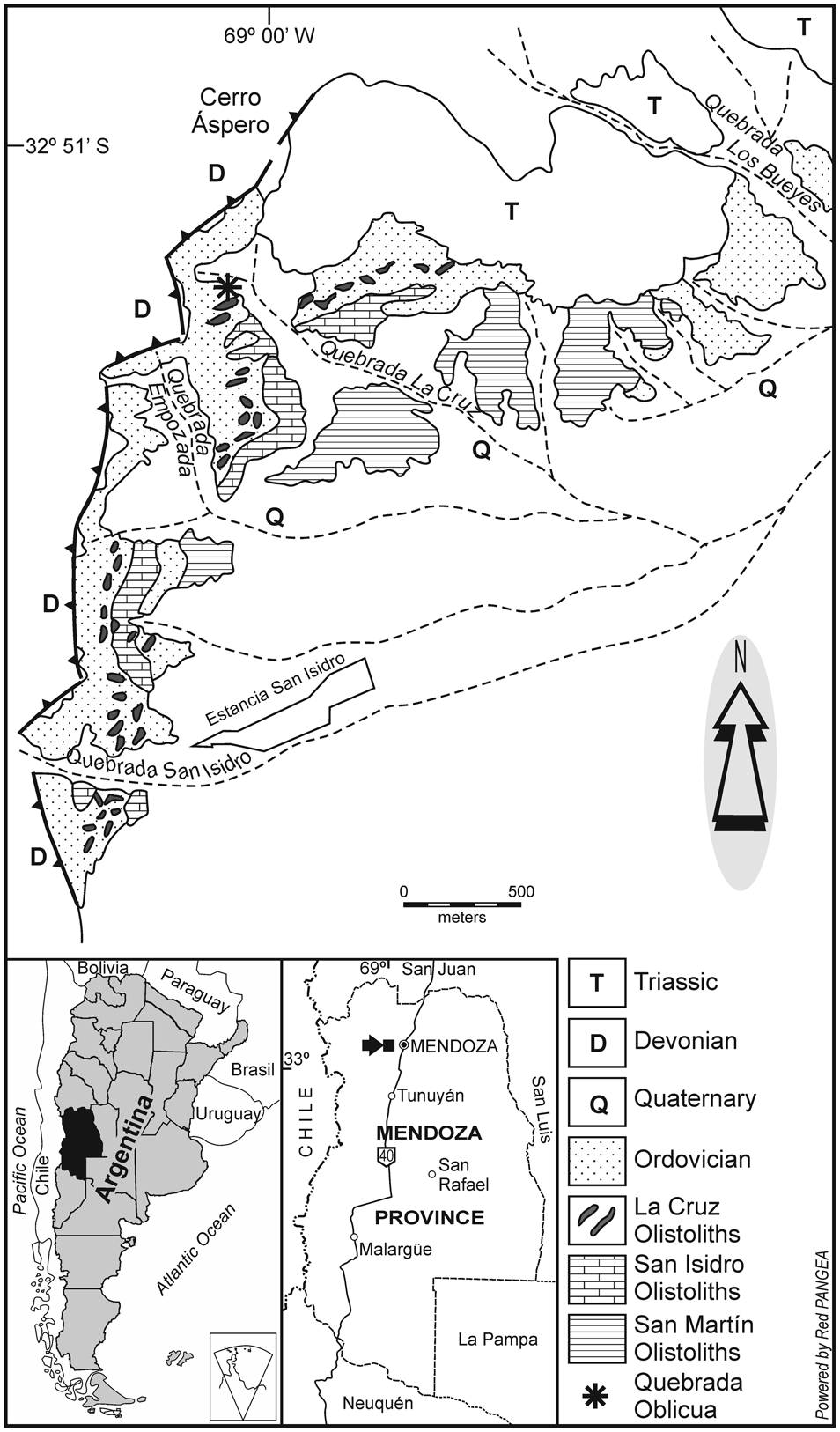
Figure 1. Map of the San Isidro area, Mendoza Province, Argentina (after Bordonaro et al., Reference Bordonaro, Beresi and Keller1993 and Tortello, Reference Tortello2014). The Quebrada Oblicua is indicated by an asterisk.
The material studied herein was found by Carlos Rusconi and Manuel Tellechea (Museo de Historia Natural de Mendoza) in the Quebrada Oblicua (=“Quebradita Oblicua” of Rusconi, Reference Rusconi1953, Reference Rusconi1954a) in the early 1950s. This fossil locality is located on the right side of the Quebrada La Cruz, which runs along the southern flank of the Cerro Áspero (Fig. 1). In a field sketch of the succession, Rusconi (Reference Rusconi1954a) stated the occurrence of light- and dark-colored limestone bearing trilobites of different ages. Specimens of Elvinia and Irvingella, the focus of the present revision, are relatively abundant (Rusconi, Reference Rusconi1954a, p. 3) and come from a Furongian dark block, which stratigraphically belongs to the La Cruz olistolith (Bordonaro, Reference Bordonaro, Gutiérrez-Marco, Saavedra and Rábano1992; Bordonaro and Banchig, Reference Bordonaro and Banchig1996).
Biostratigraphy
Biostratigraphic significance of Elvinia and Irvingella
Elvinia roemeri (Shumard) is a long-ranging middle Furongian taxon that is widely distributed over North America. Together with species of Camaraspis Ulrich and Resser in Ulrich, Reference Ulrich1924, Cliffia Wilson, Reference Wilson1951, Pterocephalia Roemer, Reference Roemer1849, Housia Walcott, Reference Walcott1916, Dellea Wilson, Reference Wilson1949, and Irvingella among others, it defines the widely recognized late Steptoean Elvinia Zone (e.g., Frederickson, Reference Frederickson1949; Wilson, Reference Wilson1949; Lochman, Reference Lochman1950, Reference Lochman1964; Nelson, Reference Nelson1951; Bell et al., Reference Bell, Feniak and Kurtz1952; Lochman and Hu, Reference Lochman and Hu1960; Grant, Reference Grant1965; Palmer, Reference Palmer1965, Reference Palmer1982; Kurtz, Reference Kurtz1975; Taylor, Reference Taylor1976; Stitt, Reference Stitt1977; Westrop, Reference Westrop1986; Hohensee and Stitt, Reference Hohensee and Stitt1989; Pratt, Reference Pratt1992; Stitt et al., Reference Stitt, Rucker, Boyer and Hart1994; Loch and Taylor, Reference Loch, Taylor, Mann and Lane1995, Reference Loch and Taylor2004; Taylor et al., Reference Taylor, Loch and Perfetta1999; Westrop and Adrain, Reference Westrop and Adrain2007; Chatterton and Gibb, Reference Chatterton and Gibb2016). A basal Sunwaptan faunule at the top of the unit, the “Irvingella major Subzone” of Palmer (Reference Palmer1979) (= I. major coquina of Wilson and Frederickson, Reference Wilson and Frederickson1950), has been formalized as a separate zone by Chatterton and Ludvigsen (Reference Chatterton and Ludvigsen1998). The latter is largely dominated by Irvingella “major” and is typically associated with Comanchia Frederickson in Wilson and Frederickson, Reference Wilson and Frederickson1950. As is the case with the Elvinia Zone, the I. major Zone is a widespread unit in North America (see a synthesis in Hohensee and Stitt, Reference Hohensee and Stitt1989, p. 860–861), although is only a few feet thick (Lochman and Wilson, Reference Lochman and Wilson1958; Palmer, Reference Palmer1965; Loch and Taylor, Reference Loch, Taylor, Mann and Lane1995; Westrop and Adrain, Reference Westrop and Adrain2007).
While Elvinia Walcott, Reference Walcott1924 is confined to Laurentia, Irvingella has been recorded from diverse paleocontinents at different latitudes (Żylińska et al., Reference Żylińska, Weidner, Ahlgren and Ahlberg2015). Such a cosmopolitan distribution may have been facilitated by the development of pelagic habits at some point of the life cycle (Rushton, Reference Rushton1967; Fortey, Reference Fortey1985; Żylińska et al., Reference Żylińska, Weidner, Ahlgren and Ahlberg2015; Chatterton and Gibb, Reference Chatterton and Gibb2016). Most of the Irvingella occurrences lie at or near the base of the Jiangshanian global stage; in Laurentia it ranges from the upper Steptoean to the basal Sunwaptan; in Australia it defines the base of the Iverian stage; in China it comes from the middle and late Changshanian; and in Kazakhstan and Siberia, from the late Sakian (Geyer and Shergold, Reference Geyer and Shergold2000; Peng et al., Reference Peng, Babcock, Zuo, Zhu, Lin, Yang and Qi2012). In the absence of other cosmopolitan taxa, Irvingella proved to be crucial for biostratigraphic correlation of Avalonia, Baltica, and the Sino-Korean block with other parts of the world (Rushton, Reference Rushton1967, Reference Rushton1983; Hong et al., Reference Hong, Lee and Choi2003; Choi et al., Reference Choi, Kim and Lee2008; Rushton and Weidner, Reference Rushton and Weidner2010). A helpful global correlation chart indicating Irvingella records was provided by Peng et al. (Reference Peng, Babcock, Zuo, Zhu, Lin, Yang and Qi2012, fig. 2).
As shown by Palmer (Reference Palmer1965) and Hong et al. (Reference Hong, Lee and Choi2003), there appears to be a succession of Irvingella species through the lower Jiangshanian. Species with a preglabellar field and a distinct anterior border on mature cranidia, such as I. typa (Kobayashi, Reference Kobayashi1935), I. megalops (Kobayashi, Reference Kobayashi1962), I. tropica Öpik, Reference Öpik1963, and I. angustilimbata Kobayashi, Reference Kobayashi1938, predate those species like I. flohri Resser, Reference Resser1942, I. nuneatonensis (Sharman, Reference Sharman1886), and I. major whose large holaspides lack a preglabellar field and have only a trace of a frontal border. The primitive forms also show distinct eye-ridges, which are barely visible or imperceptible in the derived species.
The Elvinia Zone in Argentina
The first report of Elvinia and Irvingella from the Quebrada Oblicua of Mendoza was well received by the international scientific community. In a congratulatory letter addressed to Carlos Rusconi in 1954, Dr. B.F. Howell (Princeton University) welcomed the discovery in Argentina of these “North American” age-diagnostic genera (Rusconi, Reference Rusconi1954a, p. 3). Rusconi (Reference Rusconi1953, Reference Rusconi1954a, Reference Rusconi1954b) used the name “Irvingella jorusconii Zone” for this fossil locality and, based on the occurrence of Elvinia obliquoensis, Irvingella jorusconii, I. platycephala, I. obliquoensis and “Triarthropsis” pampanus Rusconi, Reference Rusconi1953, assigned it to the upper Cambrian; however, no further details were provided on the zone, and it was not mentioned in subsequent papers. A few years later, Wilson (Reference Wilson1957, fig. 2) correlated the Quebrada Oblicua section with the lower Franconian Elvinia Zone of the standard North America trilobite zonation (see also Borrello, Reference Borrello and Holland1971).
As stated above, Palmer (Reference Palmer1965) revised replicas of the type specimens of the Rusconi collection and recognized Elvinia roemeri and Irvingella major, confirming an assignment to the Elvinia Zone. The occurrence of E. roemeri in the Argentinian Precordillera is supported by the present study, while the associated Irvingella forms, as here revised include Irvingella jorusconii Rusconi, I. platycephala Rusconi, and Irvingella sp. Species diversity in Mendoza is relatively high; other records of three co-occurring species of Irvingella have so far only been reported from the Elvinia Zone (Wujiajiania sutherlandi Subzone) of southeastern British Columbia, Canada (Clay Creek and “Site 7” of Chatterton and Gibb, Reference Chatterton and Gibb2016, p. 4, 6) and, though subject to revision, the Morgan Creek Member of the Wilberns Formation of central Texas (Cold Creek Canyon, San Saba County; Resser, Reference Resser1942).
Adult specimens of Irvingella from Mendoza are characterized by their imperceptible ocular ridges and their reduced frontal area, which lacks a preglabellar field and exhibits barely a trace of an anterior border. Thus, I. jorusconii, I. platycephala, and Irvingella sp. represent “advanced” forms in terms of morphological development in the Irvingella lineage, and these are typical of the uppermost part of the Elvinia Zone.
Although Irvingella species from Mendoza seem to be endemic to the southern Precordillera, they display a strong resemblance with poorly documented material from the Wilberns Formation of central Texas, originally assigned by Resser (Reference Resser1942) to I. oblonga and I. abrupta (= I. “major” sensu Palmer, Reference Palmer1965), as well as with I. “major” from the Rabbitkettle Formation of the southern Mackenzie Mountains (Pratt, Reference Pratt1992) (see below, Systematic paleontology). Such similarities further suggest an uppermost Steptoean age (upper Elvinia Zone), and strengthen the Laurentian aspect of the fauna studied. It is worth noting that the exotic Argentine Precordillera was possibly part of or close to southeastern Laurentia during the late Cambrian, around the Ouachita embayment (Astini et al., Reference Astini, Benedetto and Vaccari1995; Ramos, Reference Ramos1995; Keller et al., Reference Keller, Buggisch, Lehnert, Pankhurst and Rapela1998; Keller, Reference Keller1999 and references therein).
Materials and methods
Materials and preservation
The collection studied consists of ~50 disarticulated specimens that are mostly preserved as internal molds or partly exfoliated sclerites. Before photography, the material was coated with vapors of magnesium oxide. The best-preserved specimens are illustrated in dorsal, frontal, lateral, and posterior views.
Repository and institutional abbreviation
Types, figured, and other specimens examined in this study are deposited in the Museo de Ciencias Naturales y Antropológicas “Juan Cornelio Moyano” (Mendoza, Argentina) with the prefix MCNAM.
Systematic paleontology
Terminology
The morphological terms used below have been mostly defined by Whittington and Kelly (Reference Whittington, Kelly and Kaesler1997). Abbreviations: sag, sagittally; exs, exsagittally; long, longitudinally; tr, transversally.
Order Ptychopariida Swinnerton, Reference Swinnerton1915
Suborder Ptychopariina Swinnerton, Reference Swinnerton1915
Family Elviniidae Kobayashi, Reference Kobayashi1935
Subfamily Elviniinae Kobayashi, Reference Kobayashi1935
Genus Elvinia Walcott, Reference Walcott1924
Type species
Dikelocephalus roemeri Shumard, Reference Shumard1861 from the Furongian of North America, by original designation.
Remarks
Palmer (Reference Palmer1960, Reference Palmer1965) revised the scope of this genus and provided diagnoses of its two valid species, E. roemeri and E. granulata Resser, Reference Resser1942.
Elvinia roemeri (Shumard, Reference Shumard1861)
Figure 2.1–2.9, ?2.10
- Reference Shumard1861
Dikelocephalus roemeri Shumard, p. 220.
- Reference Rusconi1953
Elvinia obliquoensis Rusconi, p. 1, text-fig. 1.
- Reference Rusconi1954a
Elvinia obliquoensis; Rusconi, p. 23, text-fig. 14, pl. 2, fig. 1.
- Reference Rusconi1954a
Irvingella obliquoensis Rusconi; Rusconi, pl. 2, fig. 9 (only).
- Reference Rusconi1956b
Elvinia obliquoensis; Rusconi, pl. 4, fig. 1.
- Reference Rusconi1956b
Irvingella obliquoensis; Rusconi, pl. 4, fig. 9 (only).
- Reference Castellaro, Amos, Camacho, Castellaro and Menéndez1963
Elvinia obliquoensis; Castellaro, p. 36.
- Reference Palmer1965
Elvinia roemeri (Shumard); Palmer, p. 44, pl. 3, figs. 9, 11, 14, 16 (see for further synonymy).
- Reference Chatterton and Gibb2016
Elvinia roemeri; Chatterton and Gibb, p. 55, pl. 33, figs. 1–11, pl. 34, figs. 1–8, pl. 35, figs. 1–3, pl. 80, fig. 8 (see for further synonymy).

Figure 2. (1–9) Elvinia roemeri (Shumard, Reference Shumard1861) from the Furongian Elvinia Zone of the Quebrada Oblicua, San Isidro area, Precordillera of Mendoza. (1, 2, 4, 5, 8) Cranidium in dorsal, lateral-dorsal, anterior, and posterior views, MCNAM 17012, Elvinia obliquoensis Rusconi, Reference Rusconi1953 holotype (illustrated previously by Rusconi, Reference Rusconi1953, text-fig. 1; Rusconi, Reference Rusconi1954a, text-fig. 14, pl. 2, fig. 1; Rusconi, Reference Rusconi1956b, pl. 4, fig. 1; Castellaro, Reference Castellaro, Amos, Camacho, Castellaro and Menéndez1963, p. 36); (3) cranidium, MCNAM 17014; (6) cranidium, MCNAM 17015; (7) cranidium, MCNAM 17013, Elvinia obliquoensis Rusconi, Reference Rusconi1953 paratype; (9) cranidium, MCNAM 17047b. (10) Elvinia roemeri? (Shumard, Reference Shumard1861) from the Furongian Elvinia Zone of the Quebrada Oblicua, San Isidro area, Precordillera of Mendoza, librigena, MCNAM 17028. Scale bars = 3 mm.
Neotype
Cranidium (U.S. Nat. Mus. 70259) from the Wilberns Formation of Texas (Walcott, Reference Walcott1925, pl. 17, figs. 9, 10; Bridge and Girty, Reference Bridge and Girty1937, pl. 69, fig. 9).
Materials
Seven cranidia and one librigena (MCNAM 17012–17017, 17047b, 17028) from the Quebrada Oblicua, San Isidro area, Mendoza, Argentina. La Cruz Olistolith, Furongian, Elvinia Zone.
Remarks
Elvinia roemeri is widely distributed in the Furongian Elvinia Zone of North America (e.g., Palmer, Reference Palmer1965; Westrop, Reference Westrop1986; Pratt, Reference Pratt1992; Chatterton and Ludvigsen, Reference Chatterton and Ludvigsen1998; Chatterton and Gibb, Reference Chatterton and Gibb2016). The cranidium of this species is characterized by a truncato-conical glabella bearing a distinct, transglabellar lateral furrow S1, a moderately developed frontal area, a convex and laterally narrowing (exsag.) anterior border, a slightly divergent anterior facial suture, faint ocular ridges, broad fixigenae, and a smooth external surface of exoskeleton (e.g., Kobayashi, Reference Kobayashi1938; Frederickson, Reference Frederickson1949; Lochman, Reference Lochman1950; Palmer, Reference Palmer1965, Reference Palmer1982; Kurtz, Reference Kurtz1975). It is distinguished from E. granulata, from the Elvinia Zone of Nevada, Utah, Indiana, and New York (Palmer, Reference Palmer1965, pl. 3, fig. 12; Palmer, Reference Palmer1982, pl. 1, figs. 2, 3; Ludvigsen and Westrop, Reference Ludvigsen and Westrop1983, pl. 3, figs. 1–11, pl. 4, figs. 17, 18, pl. 5, figs. 10–13) mainly because the latter shows fine granules on the external surface of the test.
Palmer (Reference Palmer1965) examined replicas of the types of Elvinia obliquoensis from the Precordillera of Mendoza and concluded that they are conspecific with E. roemeri. The E. obliquoensis holotype, which is reillustrated herein (Fig. 2.1, 2.2, 2.4, 2.5, 2.8), compares most closely with a similarly sized cranidium from Missouri (Kurtz, Reference Kurtz1975, pl. 1, fig. 12). Additional cranidia from Mendoza (Fig. 2.3, 2.6, 2.7, 2.9) display preglabellar fields of variable length; a variation that is also seen in collections from southeastern British Columbia, Montana, Wyoming, Texas, and Pennsylvania (Bridge and Girty, Reference Bridge and Girty1937; Wilson, Reference Wilson1951; Lochman and Hu, Reference Lochman and Hu1960; Lochman, Reference Lochman1964; Chatterton and Ludvigsen, Reference Chatterton and Ludvigsen1998).
A fragmentary librigena from the Rusconi collection (Fig. 2.10) is moderately wide (tr.) and has a genal spine that is bent outward from curvature of lateral margin. Although this sclerite was originally identified as Irvingella jorusconii, it may belong to Elvinia roemeri. Similarly, a pygidium regarded by Rusconi (Reference Rusconi1954a, pl. 2, fig. 9; see also Rusconi, Reference Rusconi1956b, pl. 4, fig. 9) as Irvingella obliquoensis is characterized by having a transversely semielliptical outline; a raised, broad and subparallel-sided axis reaching the posterior border furrow; noticeable anterior pleural furrows; and a narrow (sag.) but distinct border. Such features accord with Elvinia roemeri (e.g., Westrop, Reference Westrop1986; Chatterton and Gibb, Reference Chatterton and Gibb2016), even though unfortunately the specimen was not found in the collection for further analysis.
Genus Irvingella Ulrich and Resser in Walcott, Reference Walcott1924
Type species
Irvingella major Ulrich and Resser in Walcott, Reference Walcott1924 from the Lone Rock Formation of Wisconsin, U.S.A., by original designation.
Remarks
The concept of Irvingella has been comprehensively reviewed by Palmer (Reference Palmer1960, Reference Palmer1965), Ivshin (Reference Ivshin1962), and Öpik (Reference Öpik1963). Zhang (Reference Zhang2000) and Hong et al. (Reference Hong, Lee and Choi2003) demonstrated that Komaspis Kobayashi, Reference Kobayashi1935 (type species K. typa Kobayashi, Reference Kobayashi1935 from the Furongian of Korea) is a synonym of Irvingella, and Palmer (Reference Palmer1954, Reference Palmer1965) discontinued recognition of I. (Irvingellina) Kobayashi, Reference Kobayashi1938—type species, I. (I.) protuberans Kobayashi, Reference Kobayashi1938—and I. (Parairvingella) Kobayashi, Reference Kobayashi1938—type species, I. (P.) angustilimbatus Kobayashi, Reference Kobayashi1938—from the Furongian of British Columbia, Canada, as valid subgenera.
Resser (Reference Resser1942) proposed a large number of species of Irvingella from North America, which were later synonymized with the type species Irvingella major (Frederickson, Reference Frederickson1949; Gaines, Reference Gaines1951; Palmer, Reference Palmer1965). Recently, Westrop and Adrain (Reference Westrop and Adrain2016) revised in detail most of the Laurentian and Australian species of the genus, counting, among others, a pair of valid forms (I. media; I. deckeri Resser, Reference Resser1942) originally proposed by Resser (Reference Resser1942). That review showed that several supposedly widespread species have, in fact, narrower geographic distributions (e.g., I. major; I. angustilimbata; I. flohri). Irvingella major has been reported from various Cambrian paleocontinents and used widely in the literature (Walcott, Reference Walcott1925; Ivshin, Reference Ivshin1962; Ergaliev, Reference Ergaliev1980; Pratt, Reference Pratt1992; Pegel, Reference Pegel2000; Hong et al., Reference Hong, Lee and Choi2003; Lazarenko et al., Reference Lazarenko, Gogin, Pegel, Sukhov, Abaimova, Egorova, Fedorov, Raevskaya, Ushatinskaya, Rozanov and Varlamov2008; Rushton and Weidner, Reference Rushton and Weidner2010; Peng et al., Reference Peng, Babcock, Zuo, Zhu, Lin, Yang and Qi2012; Chatterton and Gibb, Reference Chatterton and Gibb2016; among many others), but Westrop and Adrain (Reference Westrop and Adrain2016) suggested that it would be better restricted to the imperfectly preserved original type lot from Wisconsin, U.S.A.; a systematic decision that is followed herein.
Clearly, the best-documented taxa are described on the basis of more than one single sclerite type, but unfortunately to date there are several species of Irvingella without pygidia available for study (e.g., I. major sensu Westrop and Adrain, Reference Westrop and Adrain2016; I. deckeri; I. tumifrons [Hall and Whitfield, Reference Hall and Whitfield1877], I. suecica Westergård, Reference Westergård1947), and when the latter are present in the collections, they are commonly few in number (e.g., see I. tropica; I. angustilimbata; I. flohri; I. media Resser, Reference Resser1942; I. orrensis Westrop and Adrain, Reference Westrop and Adrain2016). It is interesting to note that known pygidia of Irvingella show a high range of interspecific variation regarding both the general outline of the sclerite and the size of the axis (e.g., see Chatterton and Gibb, Reference Chatterton and Gibb2016, text-figs. 11, 12).
Meanwhile, cranidial characters are common and essential components of the diagnoses at specific level, and involve mainly the contour of anterior margin, sagittal length of frontal area, presence or absence of anterior border furrow, relative development of anterior border and preglabellar field, outline and convexity of glabella, degree of expression of glabellar lateral furrows, width and shape of fixigena, presence or absence of ocular ridge, and size and curvature of palpebral furrow and palpebral lobe. To illustrate adequately these features, it is highly recommended to provide different views (e.g., frontal, lateral) of the cranidia examined and not just dorsal ones (Westrop and Adrain, Reference Westrop and Adrain2016).
Articulated specimens of Irvingella are known only from a few fossil localities (Rushton, Reference Rushton1967; Chatterton and Ludvigsen, Reference Chatterton and Ludvigsen1998). Chatterton and Gibb (Reference Chatterton and Gibb2016, p. 58) showed that the thorax also changes markedly from species to species, especially in the location and number of the longest macropleural spines, the direction of projection of the marginal spines, and even in the number of segments.
Irvingella jorusconii Rusconi, Reference Rusconi1953
Figures 3, 4.1–4.10, ?4.11–4.12, 4.13, ?4.14–4.17
- Reference Rusconi1953
Irvingella jorusconii Rusconi, p. 2, text-fig. 2a, b.
- ?Reference Rusconi1953
Irvingella obliquoensis Rusconi, p. 2, text-figs. 4, 5.
- Reference Rusconi1954a
Irvingella jorusconii; Rusconi, p. 28, text-fig. 15a, b, pl. 2, fig. 6.
- ?Reference Rusconi1954a
Irvingella obliquoensis; Rusconi, p. 31, text-figs. 17, 18, pl. 2, fig. 8 (only).
- Reference Rusconi1956b
Irvingella jorusconii; Rusconi, pl. 4, fig. 6.
- ?Reference Rusconi1956b
Irvingella obliquoensis; Rusconi, pl. 4, fig. 8 (only).
- Reference Castellaro, Amos, Camacho, Castellaro and Menéndez1963
Irvingella jorusconii; Castellaro, p. 35.
- Reference Palmer1965
Irvingella major Ulrich and Resser; Palmer, p. 46.

Figure 3. Irvingella jorusconii Rusconi, Reference Rusconi1953 from the Furongian Elvinia Zone of the Quebrada Oblicua, San Isidro area, Precordillera of Mendoza. (1–5) Cranidium in dorsal, anterior, lateral, and posterior views, MCNAM 17000, holotype (illustrated previously by Rusconi, Reference Rusconi1953, text-fig. 2; Rusconi, Reference Rusconi1954a, text-fig. 15, pl. 2, fig. 6; Rusconi, Reference Rusconi1956b, pl. 4, fig. 6; Castellaro, Reference Castellaro, Amos, Camacho, Castellaro and Menéndez1963, p. 35); (6–10) cranidium in dorsal, anterior, lateral, and posterior views, MCNAM 17001a, paratype; (11) cranidium, MCNAM 17001b, paratype; (12–14) cranidium in anterior, lateral, and posterior views, MCNAM 17002; (15) fragmentary cranidium lacking collection number. Scale bars = 3 mm.

Figure 4. (1–10, 13) Irvingella jorusconii Rusconi, Reference Rusconi1953 from the Furongian Elvinia Zone of the Quebrada Oblicua, San Isidro area, Precordillera of Mendoza. (1) Fragmentary cranidium lacking collection number; (2) small cranidium lacking collection number; (3, 6, 7) cranidium lacking collection number in lateral, dorsal, and anterior views; (4) fragmentary cranidium, MCNAM 17043; (5) fragmentary cranidium, MCNAM 17010, Irvingella obliquoensis Rusconi, Reference Rusconi1953 paratype; (8–10) pygidium in dorsal, lateral, and posterior views, MCNAM 17059; (13) pygidium, latex cast of specimen illustrated in Fig. 4.8–4.10. (11, 12, 14–17) Irvingella jorusconii? Rusconi, Reference Rusconi1953 from the Furongian Elvinia Zone of the Quebrada Oblicua, San Isidro area, Precordillera of Mendoza. (11, 12, 16, 17) Fragmentary cranidium in lateral, dorsal, anterior, and posterior views, MCNAM 17009, Irvingella obliquoensis Rusconi, Reference Rusconi1953 holotype (illustrated previously by Rusconi, Reference Rusconi1953, text-fig. 4; Rusconi, Reference Rusconi1954a, text-fig. 17, pl. 2, fig. 8; Rusconi, Reference Rusconi1956b, pl. 4, fig. 8); (14, 15) pygidium in dorsal and posterior-dorsal views, MCNAM 17137 (illustrated previously by Rusconi, Reference Rusconi1953, text-fig. 5; Rusconi, Reference Rusconi1954a, text-fig. 18). Scale bars = 3 mm.
Holotype
Cranidium (MCNAM 17000) from the Quebrada Oblicua, San Isidro area, Mendoza, Argentina (Rusconi, Reference Rusconi1954a, text-fig. 15, pl. 2, fig. 6; Fig. 3.1–3.5).
Diagnosis
Glabella inflated, slightly tapered beyond S1 and gently rounded anteriorly, strongly elevated above level of fixigenae; axial furrows conspicuous, remarkably wide (tr.); frontal area very short (sag.), lacking preglabellar field; fixigenae narrow (tr.), with maximum width of palpebral area (excluding palpebral lobe) equal to less than one-quarter of maximum cranidial width, narrowing strongly anteriorly; palpebral furrow deep, strongly arcuate; pygidium semiovate in outline, with acuminate anterolateral corners and convergent lateral margins; pygidial axis occupying about three-quarters of total pygidial length (excluding articulating half ring); pleural field narrowed backward, crossed by a well-marked anterior pleura; pygidial border furrow indistinct.
Description
Cranidium convex, subtrapezoidal in outline, with straight to slightly curved anterior margin and steeply downsloping fixigenae, length ~68% of maximum width between palpebral furrows; anterior arch weak (Fig. 3.7) to absent (Fig. 3.2); axial furrows conspicuous, remarkably wide (tr.), incised on both testate specimens and internal molds (e.g., Fig. 3.5, 3.15); glabella large, inflated, much longer than wide, strongly elevated above level of fixigenae, occupying ~95–99% of total cranidial length (sag.) and 56–58% of cranidial width (tr.) between palpebral furrows at level of abaxial tip of S1; glabellar margins subparallel at L0 and L1, becoming slightly convergent beyond S1 and gently rounded anteriorly; glabellar width at S2 equal to ~85% of width at S1; lateral profile of glabella slightly convex between L0 and S2, then strongly curved downward toward preglabellar furrow; L0 and L1 of similar size, clearly delimited by deep S0 and S1; S0 and S1 transglabellar, with a transverse medial portion and forwardly oblique lateral portions that reach axial furrows; L0 and L1 occupy, together, ~41–42% of total glabellar length; S2 faint, gently curved, transverse, expressed only abaxially; some specimens show indications of a delicate S3 (Fig. 3.7); frontal area of cranidium very short (sag.), with weakly inflated anterior border, lacking preglabellar field; because the glabella overhangs the frontal region, the anterior cranidial border is either invisible (Fig. 3.1) or hardly seen (Figs. 3.6, 4.6) in dorsal view; anterior border furrow very delicate in small holaspides (Fig. 4.2) and absent in large specimens (e.g., Fig. 3.1); fixigenae narrow (tr.), with maximum width of palpebral area (excluding palpebral lobe) equal to less than one-quarter of maximum cranidial width, narrowing strongly anteriorly; ocular ridge indistinct; palpebral furrow strongly arcuate, deep, of even width (tr.); palpebral lobe very long (exsag.), extended from level of S0 to frontal glabellar lobe; posterior fixigena with deeply incised posterior border furrow, as developed as axial furrow; external surface of cuticule smooth. Small cranidia (Fig. 4.2) show a frontal area that, in contrast with that of late holaspides, is perfectly visible in dorsal view.
Pygidium semiovate in outline, wider than long, length ~60% of maximum width, with acuminate anterolateral corners and convergent lateral margins, obtusely rounded posteriorly; axis convex, strongly elevated above level of pleural fields, a little tapered backward, rounded at posterior end, composed of three rings and a terminal piece that are defined by transverse ring furrows (first ring furrow more distinct that the second one; third represented only by a faint groove); axis occupies about three-quarters of total pygidial length (excluding articulating half ring) and 35% of maximum pygidial width; axial furrows discrete, becoming very shallow behind posterior tip of axis; articulating half ring narrow (sag.), delimited by a deep and wide (sag.) articulating furrow; pleural field narrowed backward, only slightly downsloping, crossed by an anteriormost pleura which is equally divided by a firmly impressed pleural furrow, and a very delicate second pleural furrow; border furrow indistinct; pygidial border undifferentiated; posterior margin of pygidium lacks a medial arch in posterior view.
Materials
Thirteen cranidia and two pygidia (MCNAM 17000 [holotype], 17001a [paratype], 17001b [paratype], 17002, 17008, 17010 [Irvingella obliquoensis paratype], 17043, 17059, 17060, and five numberless specimens; ?MCNAM 17009 [Irvingella obliquoensis holotype], 17137) from the Quebrada Oblicua, San Isidro area, Mendoza, Argentina. La Cruz Olistolith, Furongian, Elvinia Zone.
Remarks
Although the holotype cranidium of Irvingella jorusconii was previously illustrated only with a sketch and a small, retouched photograph (Rusconi, Reference Rusconi1954a, text-fig. 15, pl. 2, fig. 6), Rusconi (Reference Rusconi1953, p. 2; 1954a, p. 28–29) clearly stated that a tumid glabella, a short (sag.) frontal area, and a proportionately narrow (tr.) fixigena, are among the distinguishing features of this species. Together with other characters listed above, an emended diagnosis is provided here.
Palmer (Reference Palmer1965) revised replicas of the types of I. jorusconii and reassigned them to the type species I. major Ulrich and Resser in Walcott, Reference Walcott1924. However, the type material of the latter (Walcott, Reference Walcott1925, pl. 15, figs. 26–29; Westrop and Adrain, Reference Westrop and Adrain2016, p. 413–418, fig. 14A–E), although poorly preserved, clearly differs from the Argentinian specimens in having a longer (sag.) frontal area, a less tumid frontal glabellar lobe, as well as wider (tr.) fixigenae. Irvingella jorusconii is thus restated here as a diagnosable, valid taxon.
The only available pygidium of the type series of I. jorusconii (Fig. 4.8–4.10, 4.13) is tentatively kept in this species; its general morphology bears some resemblance to that of Irvingella sp. from the Orr Formation of Utah (Westrop and Adrain, Reference Westrop and Adrain2016, fig. 14A, B, F–H), though the former differs by its distinctive anterior pleura and its imperceptible border furrow. The presence of a well-defined anterior pleural furrow is shared with Bartonaspis Westrop and Adrian, Reference Westrop and Adrain2007, from the Irvingella major Zone of Nevada, Utah, and Oklahoma (e.g., Westrop and Adrian, Reference Westrop and Adrain2007, figs. 4r–t, 8a–f, 11m–n), but the pygidium of Bartonaspis is distinguished in the transversely subelliptical outline and smaller number of axial segments.
In addition, three specimens catalogued as Irvingella obliquoensis in the Rusconi collection are closely comparable with the material described above. The paratype cranidium of I. obliquoensis (Fig. 4.5), despite the fact that it is incomplete, clearly shows very wide (tr.) axial furrows and conspicuous, similarly developed S0 and S1, so it is indistinguishable from I. jorusconii. Likewise, the holotype cranidium of I. obliquoensis (Rusconi, Reference Rusconi1954a, pl. 2, fig. 8; Fig. 4.11, 4.12, 4.16, 4.17) may also be conspecific, although it is exceptionally large and, unfortunately, very fragmentary for further analysis (cf., Palmer, Reference Palmer1965). The latter is associated with a large pygidium (Fig. 4.14, 4.15), which, like that illustrated in Figure 4.8–4.10, 4.13, is semiovate in outline and exhibits a proportionately long axis, a well-defined anterior pleural furrow, and an undifferentiated border.
Besides restricting the concept of I. major to the type material from Wisconsin, Westrop and Adrain (Reference Westrop and Adrain2016, p. 415, 418) made preliminary observations on several specimens attributed to that species in Laurentia and other Cambrian continents. Unfortunately, some sclerites are scarce or too poorly documented to be evaluated properly (Westrop and Adrain, Reference Westrop and Adrain2016, p. 415). Among them, there is one occurrence that strongly resembles the material studied herein. Irvingella oblonga Resser, Reference Resser1942 (= I. “major”) (see Gaines, Reference Gaines1951; Palmer, Reference Palmer1965), which was described by Resser (Reference Resser1942, pl. 3, figs. 1–3) on one incomplete cranidium from the Morgan Creek Member of the Wilberns Formation of central Texas, exhibits, like I. jorusconii, a slightly tapered, strongly elevated anterior half of glabella, wide (tr.) axial furrows, a short (sag.) frontal area, and a relatively narrow (tr.) fixigena showing a similar width throughout its posterior half; however, further specimens are needed to improve this comparison.
Irvingella “major”, from the upper Steptoean-basal Sunwaptan of the southern Canadian Rocky Mountains, Alberta (Westrop, Reference Westrop1986, pl. 30, figs. 8–13), is comparable with I. jorusconii in sharing a similarly outlined glabella, which is laterally bounded by wide (tr.) axial furrows, a small frontal area, and relatively narrow (tr.) fixigenae. Additionally, the pygidium of the former preserves indications of one pair of pleural furrows. However, the material from Canada is separated on the basis of its less-tumid frontal glabellar lobe, and its transverse rather than obtusely rounded posterior pygidial margin.
A single cranidium of Irvingella “major” from the Furongian (Parabolina brevispina Subzone, P. spinulosa Zone) of Sweden (Rushton and Weidner, Reference Rushton and Weidner2010, fig. 3A–D) differs from I. jorusconii mainly in having a strong anterior arch and more developed fixigenae. Comparable material from the Elvinia Zone of the Great Basin, U.S.A. (e.g., Palmer, Reference Palmer1965, pl. 6, fig. 10) includes cranidia showing somewhat narrower axial furrows and pygidia with a subtrapezoidal rather than a semiovate outline, two axial rings instead of three, and a distinct border (Palmer, Reference Palmer1965, pl. 6, fig. 9).
Some complete exoskeletons of I. “major” from the Elvinia Zone (Wujiajiania sutherlandi Subzone) of southeastern British Columbia, Canada (Chatterton and Gibb, Reference Chatterton and Gibb2016, pl. 38, figs. 3, 6, pl. 39, figs. 4, 5, 7) bear pygidia that have, like that of I. jorusconii, three axial rings and terminal piece, convergent lateral sides, curved posterior margins, and vestiges of an anterior, distally curved pleura; however, these specimens are distinguished from the Argentinian species mainly by their wider fixigenae. Similarly, Irvingella dawnae Chatterton and Gibb, Reference Chatterton and Gibb2016, from the Wujiajiania sutherlandi Subzone of British Columbia (Chatterton and Gibb, Reference Chatterton and Gibb2016, pl. 39, figs. 1, 2, 10, pl. 83, fig. 3, text-fig. 12A), possesses a proportionately elongated, semiovate pygidium showing a long axis, a clearly marked anterior pleural furrow, and an obscure border furrow; however, this taxon is differentiated from I. jorusconii in the presence of a subparallel sided glabella, wider fixigenae, and a narrower pygidial axis.
The material examined herein shares proportions of the palpebral area of the fixigenae with those of I. deckeri, from the Honey Creek Formation of central Oklahoma (Resser, Reference Resser1942, pl. 3, figs. 19–27; Westrop and Adrain, Reference Westrop and Adrain2016, fig. 15A–I); however, late holaspides of I. jorusconii differ in having a more globose frontal glabellar lobe, wider (tr.) axial furrows, strongly downsloping fixigenae that are very narrow (tr.) anteriorly, and a shorter (sag.) frontal area in dorsal view.
Irvingella tumifrons was based on two cranidia from the Dunderberg Formation of Pogonop Ridge, Nevada (Hall and Whitfield, Reference Hall and Whitfield1877, pl. 2, figs. 38, 39; Resser, Reference Resser1942, pl. 4, figs. 42, 43; Westrop and Adrain, Reference Westrop and Adrain2016, fig. 18A–F) that resemble I. jorusconii in having a tumid frontal glabellar lobe and wide axial furrows, and in lacking a distinct preglabellar field; however, they are easily distinguished from the Argentinian species by their much wider (tr.) fixigenae. This comparison also applies to I. “tumifrons” from the Furongian of central Kazakhstan (Ivshin, Reference Ivshin1962, pl. 3, figs. 1–4).
Three cranidia and one pygidium of Irvingella “major” from the Ogon'or Formation of the northeastern Siberian Platform (Lazarenko et al., Reference Lazarenko, Gogin, Pegel, Sukhov, Abaimova, Egorova, Fedorov, Raevskaya, Ushatinskaya, Rozanov and Varlamov2008, pl. 17, figs. 9–12, 12a) share a tumid glabella, a reduced frontal area, relatively narrow fixigenae, and arcuate palpebral furrows with I. jorusconii, but the former have narrower axial furrows and a subtrapezoidal pygidium. Partially flattened sclerites of I. “major” from the Machari Formation (Agnostotes orientalis Zone) of Korea (Hong et al., Reference Hong, Lee and Choi2003, pl. 2, figs. 21–27) are differentiated from I. jorusconii in showing vestiges of a distinctive anterior cranidial border, broader fixigenae, and a shorter pygidial axis.
Both I. jorusconii and I. orrensis, from the Orr Formation of Utah (Westrop and Adrain, Reference Westrop and Adrain2016, figs. 19–23, 24B, C, E, I–T), exhibit a very short frontal area lacking preglabellar field, but the latter species is distinguished mainly in the presence of a subrectangular glabella that is delimited by more delicate furrows, a broader palpebral area of fixigena, and a transversely elongate pygidium. Large holaspides of Irvingella suecica, from the Alum Shale Formation of Sweden (Westergård, Reference Westergård1947, pl. 3, figs. 1–3; Rushton and Weidner, Reference Rushton and Weidner2010, fig. 8A–K), also lack a preglabellar field, but differ from I. jorusconii in showing wider fixigenae and a better defined anterior cranidial border.
Irvingella flohri, from the Dunderberg Formation of Nevada and British Columbia (Resser, Reference Resser1942, pl. 4, figs. 12–14; Palmer, Reference Palmer1965, pl. 6, figs. 16, 19, 20, 24; Chatterton and Gibb, Reference Chatterton and Gibb2016, pl. 9, fig. 7, pl. 37, figs. 2, 3, pl. 40, figs. 1–9, pl. 41, figs. 1–9, pl. 42, figs. 1–4, pl. 77, fig. 4; Westrop and Adrain, Reference Westrop and Adrain2016, figs. 9A–K, 10A–C, H–J), differs from the material studied herein because the former exhibits a little longer frontal area, a nearly transverse anterior glabellar margin, and wider fixigenae. As in I. jorusconii, the pygidium of I. flohri shows a distinct anterior pleural furrow (see Palmer, Reference Palmer1965, pl. 6, fig. 24), but is separated by having only two axial rings and less-convergent lateral margins.
Specimens from the Jiangshanian of Zhejiang, Southeast China, regarded by Peng et al. (Reference Peng, Babcock, Zuo, Zhu, Lin, Yang and Qi2012, fig. 9O–R) as I. major, are clearly differentiated from the sclerites from Argentina in having more delicate axial furrows, a longer (sag.) frontal area, and much wider (tr.) fixigenae. Similarly, Irvingella angustilimbata Kobayashi, Reference Kobayashi1938 sensu Westrop and Adrain (Reference Westrop and Adrain2016), from western Utah, eastern Nevada, and northern Canada (e.g., Resser, Reference Resser1942, pl. 4, figs. 18–22; Palmer, Reference Palmer1965, pl. 6, figs. 17, 18, 21–23; Pratt, Reference Pratt1992, pl. 11, figs. 7–13, 16; Westrop and Adrain, Reference Westrop and Adrain2016, figs. 1–8 and references therein), as well as I. media, from central Texas and Oklahoma (Resser, Reference Resser1942, pl. 3, figs. 46–54; Wilson, Reference Wilson1951, pl. 10, fig. 7, pl. 11, figs. 16, 17, 19, 20; Westrop and Adrain, Reference Westrop and Adrain2016, figs. 16, 17), are distinguished from I. jorusconii in the presence of short but distinct preglabellar fields and different fixigenal proportions. Irvingella angustilimbata has, in addition, a transversely elongate pygidium with more convergent lateral margins, and I. media bears a pygidium that is subtrapezoidal in outline.
Other species of Irvingella further differ from the material studied herein in possessing extremely broad fixigenae (I. transversa Palmer, Reference Palmer1965, from the Elvinia Zone of the Great Basin [Palmer, Reference Palmer1965, pl. 6, figs. 7, 8]; I. lata Chatterton and Gibb, Reference Chatterton and Gibb2016, from the Wujiajiania lyndasmithae Subzone of Elvinia Zone of British Columbia [Chatterton and Gibb, Reference Chatterton and Gibb2016, pl. 44, figs. 1–9, pl. 45, figs. 1, 2, text-fig. 11B]) or a well-developed preglabellar field (e.g., I. tropica, from the I. tropica Zone of Queensland, Australia [Öpik, Reference Öpik1963, pl. 4, figs. 5–8, text-fig. 36; Shergold, Reference Shergold1982, pl. 7, figs. 4–7, pl. 8, fig. 10; Westrop and Adrain, Reference Westrop and Adrain2016, figs. 11, 12, 13A–D]; I. typa, from the Eugonocare longifrons Zone of Korea [Hong et al., Reference Hong, Lee and Choi2003, pl. 1, figs. 1–11]; I. megalops from Korea [Eochuangia hana Zone], Kazakhstan [Pseudagnostus “curtare” Zone], and eastern Siberia [latest Paibian] [Ergaliev, Reference Ergaliev1980, pl. 12, figs. 12, 13; Hong et al., Reference Hong, Lee and Choi2003, pl. 1, figs. 12–23; Lazarenko et al., Reference Lazarenko, Gogin, Pegel, Sukhov, Abaimova, Egorova, Fedorov, Raevskaya, Ushatinskaya, Rozanov and Varlamov2008, pl. 17, fig. 7]).
Comparisons of I. jorusconii with the co-occurring I. platycephala and Irvingella sp. are provided below.
Irvingella platycephala Rusconi, Reference Rusconi1953
Figure 5.1–5.6
- Reference Rusconi1953
Irvingella platycephala Rusconi, p. 2, text-fig. 3a,b.
- Reference Rusconi1954a
Irvingella platycephala; Rusconi, p. 29, text-fig. 16a, b, pl. 2, fig. 7.
- Reference Rusconi1956b
Irvingella platycephala; Rusconi, pl. 4, fig. 7.

Figure 5. (1–6) Irvingella platycephala Rusconi, Reference Rusconi1953 from the Furongian Elvinia Zone of the Quebrada Oblicua, San Isidro area, Precordillera of Mendoza, cranidium in dorsal, posterior-dorsal, lateral, anterior, and anterior-oblique views, MCNAM 17004, holotype (illustrated previously by Rusconi, Reference Rusconi1953, text-fig. 3a, b; Rusconi, Reference Rusconi1954a, text-fig. 16a, b, pl. 2, fig. 7; Rusconi, Reference Rusconi1956b, pl. 4, fig. 7). (7–11) Irvingella sp. from the Furongian Elvinia Zone of the Quebrada Oblicua, San Isidro area, Precordillera of Mendoza, cranidium in dorsal, lateral, anterior-oblique, anterior, and posterior views, MCNAM 17005a, Irvingella platycephala Rusconi, Reference Rusconi1953 paratype. Scale bars = 2 mm.
Holotype
Cranidium (MCNAM 17004) from the Quebrada Oblicua, San Isidro area, Mendoza, Argentina (Rusconi, Reference Rusconi1954a, text-fig. 16a, b, pl. 2, fig. 7; Fig. 5.1–5.6).
Diagnosis
A species of Irvingella having a barrel-shaped glabella with rounded anterolateral and anterior margins; maximum glabellar width at L2; S0–S3 distinct; L1, L2, and L3 of similar length (exsag.); frontal glabellar lobe overhangs cranidial frontal area; frontal area very short (sag.), lacking preglabellar field; anterior border furrow almost imperceptible; in anterior view, there is a perceptible change in slope between fixigena and glabella.
Materials
One cranidium (MCNAM 17004, holotype) from the Quebrada Oblicua, San Isidro area, Mendoza, Argentina. La Cruz Olistolith, Furongian, Elvinia Zone.
Remarks
As stated above, Palmer (Reference Palmer1965, p. 46) examined replicas of type specimens of the Rusconi collection, although he indicated that the holotype cranidium of Irvingella platycephala was not available for study. Rusconi (Reference Rusconi1954a, pl. 2, fig. 7; 1956b, pl. 4, fig. 7) provided a small photograph of the latter, in a view that seems to be posterodorsal (comparable with the view of Fig. 5.2 herein).
The holotype of I. platycephala (Fig. 5.1–5.6) clearly differs from the rest of the type series (paratypes MCNAM 17005, 17006; additional cranidia MCNAM 17007, 17047a, 17048, 17049; Figs. 5.7–5.11, 6.1–6.15) because the former exhibits a subparallel-sided, barrel-shaped glabella that is well elevated above level of fixigenae, a maximum glabellar width at L2 instead of L1, a proportionately long (exsag.) L3, and a rounded preglabellar furrow. Thus, the concept of I. platycephala is restricted herein to the holotype, and the remainder specimens of the type lot are redescribed separately below (see Irvingella sp.).

Figure 6. Irvingella sp. from the Furongian Elvinia Zone of the Quebrada Oblicua, San Isidro area, Precordillera of Mendoza. (1, 2, 4, 5) Cranidium in dorsal, lateral, anterior, and posterior views, MCNAM 17049; (3, 6, 9) cranidium in dorsal, posterior, and anterior-dorsal views, MCNAM 17048; (7, 8, 10, 11) cranidium in dorsal, anterior-dorsal, anterior-oblique, and posterior views, MCNAM 17005b; (12, 15) cranidium in dorsal and anterior-dorsal views, MCNAM 17047a; (13) small cranidium, MCNAM 17007; (14) cranidium in anterior-dorsal view, MCNAM 17006, Irvingella platycephala Rusconi, Reference Rusconi1953 paratype. Scale bars = 2 mm.
The glabellar outline of Irvingella platycephala contrasts with that of most species of Irvingella, which usually show, in varying degrees, tapered glabellae. As stated above, I. platycephala has a barrel-shaped glabella with its maximum width at level of L2; a rare condition that is also present in a specimen from the Morgan Creek Member of the Wilberns Formation of Texas assigned to I. abrupta by Resser (Reference Resser1942) (= I. “major”, see Palmer, Reference Palmer1965). Resser (Reference Resser1942) regarded a glabella delimited by “circular” dorsal furrows, as well as distinct S0–S3, as typical characters of I. abrupta; in dorsal view, the paratype cranidium of the latter (Resser, Reference Resser1942, pl. 3, fig. 45) hardly differs from I. platycephala in having slightly narrower fixigenae.
Irvingella major from the middle Furongian of Wisconsin (Walcott, Reference Walcott1925, pl. 15, figs. 26–29; Westrop and Adrain, Reference Westrop and Adrain2016, fig. 14A–E) also has a subparallel-sided, anteriorly rounded glabella, but this species is separated from I. platycephala on the basis of its straighter axial furrows, its longer frontal area, and its obscure glabellar S2 and S3. Irvingella deckeri, from the Honey Creek Formation of central Oklahoma (Resser, Reference Resser1942, pl. 3, figs. 19–27; Westrop and Adrain, Reference Westrop and Adrain2016, fig. 15 A–I), differs, in addition, by possessing much narrower fixigenae.
Irvingella jorusconii (Rusconi, Reference Rusconi1954a, pl. 2, fig. 6; Figs. 3, 4) is clearly distinguished from I. platycephala in having a glabella that is gently tapered in front of S1 and more elevated above level of genae, wider axial furrows, smaller fixigenae, and strongly arcuate palpebral furrows.
Irvingella sp.
Figures 5.7–5.11, 6
- Reference Rusconi1954a
Irvingella platycephala Rusconi; Rusconi, p. 29 (part).
- Reference Palmer1965
Irvingella platycephala; Palmer, p. 46 (mentioned as possibly distinct from I. major).
Description
Cranidium convex, subtrapezoidal in outline, with straight (Fig. 5.7) to slightly curved anterior margin (Fig. 6.7, 6.12) and downsloping fixigenae, length ~62–66% of maximum width between palpebral furrows; anterior arch weak (Fig. 5.10) to almost imperceptible (Fig. 6.14); glabella large, slightly longer than wide, little elevated above genal region, occupying ~96–98% of total cranidial length (sag.) and 48–52% of cranidial width (tr.) between palpebral furrows at level of abaxial tip of S1, tapered strongly forward beyond middle-part of L1 and truncate anteriorly, delimited by narrow and deep axial and preglabellar furrows that are better incised on internal molds; in anterior view, there is not a marked change in slope between flanks of glabella and fixigenae (Figs. 5.10, 6.4); maximum glabellar width at L1; glabellar width at S2 equal to ~85% of width at S1; LO accounts for less than one-fifth of glabellar length; most specimens show indications of an occipital node (e.g., Fig. 6.1, 6.3); S0 transglabellar, deep, evenly curved backward, reaching axial furrows; L1 longer (exsag.) than L0, occupying ~21–25% of total glabellar length; S1 distinctive, transglabellar, reaching close to axial furrows, with a transverse medial portion and slightly curved, forwardly oblique lateral portions; S2 fainter than S1, short (tr.), oblique backward, expressed only abaxially; L3 much smaller than L2, occupying ~15–18% of total glabellar length (exsag.); S3 delicate, gently curved, transverse (Fig. 6.1) to oblique backward (e.g., Figs. 5.7, 6.7); frontal area of cranidium lacking preglabellar field; anterior cranidial border minute (sag.), bounded by a weak anterior border furrow, which is more evident in exfoliated specimens; palpebral area of fixigena wide (tr.), equal to ~25% of cranidial width at L2, flexed downward in lateral view from axial furrow to palpebral furrow; ocular ridge absent; palpebral lobe very long (exsag.), narrow (tr.) and highly convex, evenly curved, extended from the level of mid-point of L1 to the frontal glabellar lobe, delimited by a firmly impressed palpebral furrow of even width; posterior fixigena with deeply incised posterior border furrow and convex, narrow (exsag.) posterior border that widens slightly distally; external surface of cuticule smooth.
Materials
Seven cranidia (MCNAM 17005a, 17005b, 17006, Irvingella platycephala paratypes; MCNAM 17007, 17047a, 17048, 17049) from the Quebrada Oblicua, San Isidro area, Mendoza, Argentina. La Cruz Olistolith, Furongian, Elvinia Zone.
Remarks
As stated above, the specimens examined comprise the original type series of I. platycephala, excepting the holotype. These cranidia represent a separate species characterized by having a glabella that tapers strongly forward beyond L1 and is truncate anteriorly, a proportionately small glabellar lobe L3, a minute cranidial frontal area, a frontal profile showing no change in slope between fixigenae and glabella, and relatively wide fixigenae. However, until the corresponding pygidium is available for analysis, the material is provisionally left in open nomenclature.
Small holaspides of Irvingella sp. exhibit a frontal area that is proportionately longer (sag.) than that of larger holaspides (compare Fig. 6.13 with Fig. 6.1, 6.3); an ontogenetic variation that was also documented in other species of Irvingella (e.g., I. angustilimbata; I. nuneatonensis; I. media; I. orrensis; I. jorusconii) (Rushton, Reference Rushton1967, p. 341; Westrop and Adrain, Reference Westrop and Adrain2016, p. 406, 415, 427). Additionally, large specimens of Irvingella sp. bear an occipital node, which, as noted by Öpik (Reference Öpik1963, p. 96), is an exceptional character in the genus. To date, this node has also been recognized in a few cranidia from Australia (Öpik, Reference Öpik1963, text-fig. 36; Shergold, Reference Shergold1982, pl. 7, fig. 6; Westrop and Adrain, Reference Westrop and Adrain2016, figs. 11A, H, 12G), Siberia (Pegel, Reference Pegel2000, fig. 14.3; Lazarenko et al., Reference Lazarenko, Gogin, Pegel, Sukhov, Abaimova, Egorova, Fedorov, Raevskaya, Ushatinskaya, Rozanov and Varlamov2008, pl. 17, fig. 9), Korea (Hong et al., Reference Hong, Lee and Choi2003, pl. 2, fig. 13), and northwest Canada (Pratt, Reference Pratt1992, pl. 11, fig. 3).
Irvingella sp. compares most closely with a single cranidium from the Rabbitkettle Formation of the southern Mackenzie Mountains assigned by Pratt (Reference Pratt1992, pl. 11, figs. 3, 4) to I. “major”. Pratt (Reference Pratt1992, p. 49) described the latter as having an extremely conical, tumid glabella and a frontal area consisting of only a short (sag.), convex anterior border. It is also similar to Irvingella sp. in the presence of comparable glabellar lateral furrows, a relatively short L3, and indications of an occipital node. Unfortunately, the fixigena of the Canadian specimen is partially preserved and other sclerites are unknown, limiting further comparisons.
Although Irvingella sp. shares a strongly tapered glabella with several other species of the genus (e.g., Irvingella kassini Ivshin, Reference Ivshin and Osnovnye idei1960, from the upper Sakian of Kazakhstan; see Ivshin, Reference Ivshin1962, pl. 4, fig. 2; I. media; I. tropica), the former is distinguished from all of them in lacking a marked change in slope between flanks of glabella and fixigenae in frontal view. In addition, large cranidia of I. media and I. tropica differ from Irvingella sp. in retaining a distinct preglabellar field. Similarly, I. nuneatonensis, from the upper Olenus Zone of England (Rushton, Reference Rushton1967, pl. 12, text-fig. 1), is differentiated by its proportionately longer fontal area.
Acknowledgments
I am indebted to G. Campos, E. Aranguez, V. Zorrilla, and M. Parral (Museo de Ciencias Naturales y Antropológicas “Juan Cornelio Moyano,” Mendoza) for making the material available for study and their kind assistance. The manuscript benefited from constructive remarks from the referees S. Westrop and J. Adrain. I also thank B. Pratt, B. Hunda, and J. Kastigar for their editorial work. S. Morrison helped in the search of bibliographic information. This research was supported by the Consejo Nacional de Investigaciones Científicas y Técnicas and the Universidad Nacional de La Plata, Argentina.






