



Introduction
The paleocontinent of Baltica is well known for its rich Silurian crinoid fauna, primarily from Gotland, Sweden (Angelin, Reference Angelin1878; Bather, Reference Bather1893; Franzén, Reference Franzén1982, Reference Franzén1983). However, the oldest Gotland strata are latest Telychian (Llandovery) in age, with the Gotland strata primarily Wenlock and Ludlow. The record of Silurian crinoids from Baltica has been expanded recently with description of faunas from the western Estonian Islands and central Estonia (Ausich et al., Reference Ausich, Wilson and Vinn2012, Reference Ausich, Wilson and Vinn2015b, Reference Ausich, Wilson and Tinn2019; Ausich and Wilson, Reference Ausich and Wilson2016). Significantly, these recent papers on Estonian crinoids described both Llandovery and Pridoli faunas. This included a small Llandovery (Rhuddanian) fauna from Hiiumaa Island (western Estonia). Disarticulated crinoidal remains suggested a much larger fauna than indicated by the following named taxa: Hiiumaacrinus vinni Ausich and Wilson, Reference Ausich and Wilson2016; Protaxocrinus estoniensis Ausich and Wilson, Reference Ausich and Wilson2016; Eomyelodactylus sp.; and calceocrinids. Paerticrinus arvosus Wright and Toom, Reference Wright and Toom2017, the second oldest flexible crinoid from Baltica, has also been described recently (lower Rhuddanian, Llandovery). Ausich et al. (Reference Ausich, Wilson and Tinn2019) described two additional Aeronian (Llandovery) crinoids from the Kalana Lagerstätte in central Estonia: Kalanacrinus mastikae Ausich et al., Reference Ausich, Wilson and Tinn2019 (diplobathrid camerate) and Tartucrinus kalanaensis Ausich et al., Reference Ausich, Wilson and Tinn2019 (disparid). New taxa described herein further enhance our understanding of the early Silurian crinoid radiation on Baltica.
Geographic and stratigraphic occurrences
The crinoids reported here are from Rhuddanian and Aeronian (Llandovery, Silurian) fine-grained carbonates in quarries in central and northwestern Estonia (Figs. 1, 2) (Nestor et al., Reference Nestor, Einasto, Männik and Nestor2003; Männik, Reference Männik, Bauert, Hints, Meidla and Männik2014). The oldest of these new crinoids are from the Varbola Formation (Juuru Regional Stage, Rhuddanian) in the Reinu Quarry, Rapla County, northwestern Estonia (59.087299°N, 24.73768°E). The other crinoids are from three additional quarries in the Raikküla Regional Stage (Aeronian, Llandovery), including the Kalana Quarry, Ellakvere Quarry, and the Rõstla Quarry. The Kalana Quarry exposes the Jõgeva Beds of the Nurmekund Formation (Aeronian) (Männik et al., Reference Männik, Tinn, Loydell and Ainsaar.2016) and is near the village of Kalana, in central Estonia (58.722118°N, 26.038226°E) (Fig. 1). The new crinoids reported here from the Kalana Quarry are at a stratigraphically higher level than those reported from the Kalana Lagerstätte (Ausich et al., Reference Ausich, Wilson and Tinn2019). The Ellakvere Quarry was near Jõgeva in Jõgeva County, central Estonia (58.742244°N, 26.418208°E) and exposed Jõgeva Beds of the Nurmekund Formation (Aeronian). The Rõstla Quarry is also in Jõgeva County, central Estonia (58.646504°N, 26.806853°E) and strata exposed in this quarry are the Imavere and Mõhkküla beds of the Nurmekund Formation (Aeronian) (Figs. 1, 2).

Figure 1. Locality map of the described crinoids, with the Nurmekund Formation outcrop area shaded (modified after Männik et al., Reference Männik, Tinn, Loydell and Ainsaar.2016).

Figure 2. Stratigraphic section of the Llandovery of central and western Estonia and the western Estonian Islands; crinoid occurrences are noted. Based on the published scheme of Männik (Reference Männik, Bauert, Hints, Meidla and Männik2014). B. = Beds; Fm. = Formation; Mb. = Member; O = Ordovician; O3 = Late Ordovician; Reg. = Regional.
Specimens described here are from museum collections. The precise stratigraphic beds from which these specimens were collected were destroyed through quarrying activities. However, crinoids from the Ellakvere, Kalana, and Reinu quarries are preserved in micrite, and those from the Rõstla Quarry are preserved in dolomicrite, indicating that the Rõstla Quarry specimens are from a different stratigraphic unit compared to other crinoids from the Nurmekund Formation.
Llandovery crinoids from Estonia
Llandovery (early Silurian) crinoid faunas are of particular significance because they record recovery from the Late Ordovician mass extinctions (Sepkoski, Reference Sepkoski and Walliser1996; Eckert, Reference Eckert1988; Donovan, Reference Donovan1989, Reference Donovan1994; Alroy, Reference Alroy2008, Reference Alroy2010a, Reference Alroyb; Peters and Ausich, Reference Peters and Ausich2008; Harper et al., Reference Harper, Hammarlund and Rasmussen2014; and many others). Further, this recovery represents the initial radiation of the Middle Paleozoic Crinoid Evolutionary Fauna (Ausich et al., Reference Ausich, Kammer and Baumiller1994; Baumiller, Reference Baumiller, David, Guille, Féral and Roux1994), which was a Silurian to middle Mississippian crinoid macroevolutionary fauna comprised dominantly of monobathrid camerate, flexible, and eucladid crinoids (Ausich and Deline, Reference Ausich and Deline2012). Previously described Llandovery crinoids are principally from the Laurentia and Avalonia paleocontinents (e.g., Ausich, Reference Ausich1984a, Reference Ausichb, Reference Ausich1985, Reference Ausich1986a, Reference Ausichb, Reference Ausichc, Reference Ausich1987; Eckert, Reference Eckert1984, Reference Eckert1990; Donovan, Reference Donovan1993; Fearnhead and Donovan, Reference Fearnhead and Donovan2007; Ausich and Copper, Reference Ausich and Copper2010; Ausich et al., Reference Ausich, Peter and Ettensohn2015a; Ausich and Wilson, Reference Ausich and Wilson2016; Ausich et al., Reference Ausich, Wilson and Tinn2019). The new faunas reported here are a further expansion of our developing knowledge of Baltica Llandovery crinoid faunas, including that of Donovan and Franzén (Reference Donovan and Franzén1988) and recent work by Ausich and Wilson (Reference Ausich and Wilson2016), Wright and Toom (Reference Wright and Toom2017), and Ausich et al. (Reference Ausich, Wilson and Tinn2019). Estonian Llandovery crinoids reported here include two eucladids—Euspirocrinus hintsae n. sp. (Rhuddanian) and Oepikicrinus perensae n. gen. n. sp. (Aeronian)—and a diplobathrid camerate, Rozhnovicrinus isakarae n. gen. n. sp. (Aeronian). These new taxa bring the total of Llandovery crinoids known from Estonia to eight named species and three taxa left in open nomenclature (Table 1).
Table 1. Rhuddanian and Aeronian crinoids from Estonia. * = Kalana Lagerstätte occurrence.

Euspirocrinus hintsae n. sp. and Paerticrinus Wright and Toom, Reference Wright and Toom2017 (Varbola Formation, early Rhuddanian) are the oldest Silurian crinoids recognized in Baltica. The fauna from Hiiumaa (Hilliste Formation, late Rhuddanian) includes Hiiumaacrinus Ausich and Wilson, Reference Ausich and Wilson2016, Protaxocrinus Springer, Reference Springer1906, Eomyelodactylus Foreste, Reference Foerste1919, and one or two calceocrinid taxa left in open nomenclature. This includes all major clades except the monobathrid eucamerates. Hiiumaacrinus (diplobathrid eucamerate), Eomyelodactylus (disparid), and the calceocrinids (disparid) belong to major clades that were dominant during the Early Paleozoic Crinoid Evolutionary Fauna (Ordovician); however, the three families to which these taxa belong (Dimerocrinitidae, Myelodactylidae, and Calceocrinidae, respectively) are exceptions to the general rule and did diversify during the Silurian. Euspirocrinus Angelin, Reference Angelin1878 (eucladid), Paerticrinus (flexible), and Protaxocrinus (flexible) belong to major clades that would diversify and eventually dominate the Middle Paleozoic Evolutionary Fauna. Thus, the initial Silurian (Rhuddanian) crinoid diversification on Baltica largely paralleled the diversifications on Laurentia and Avalonia.
Aeronian crinoids from Estonia are Kalanacrinus Ausich, Wilson, and Tinn, Reference Ausich, Wilson and Tinn2019, Oepikicrinus n. gen. (dimerocrinitid diplobathrid eucamerates), Tartucrinus Ausich, Wilson, and Tinn, Reference Ausich, Wilson and Tinn2019 (iocrinid disparid), and Rozhnovicrinus n. gen. (eucladid). Kalanacrinus and Rozhnovicrinus n. gen. represent further diversification of the dimerocrinitid clade; Oepikicrinus n. gen. represents the continued radiation of eucladids; and Tartucrinus is the youngest genus known of the Iocrinidae and the only Silurian iocrinid outside of Laurentia. During the Aeronian, based on known material, Baltica crinoid evolution largely expanded the Rhuddanian clades already present on Baltica. In contrast, many other clades radiated, and new clades originated on Laurentia and Avalonia. By the Wenlock, a largely cosmopolitan fauna existed on Laurentia, Avalonia, and Baltica.
Materials and methods
The superfamilial classification used here follows Cole (Reference Cole2017), Wright (Reference Wright2017), and Wright et al. (Reference Wright, Ausich, Cole, Peter and Rhenberg2017); family-level classifications typically follow Moore and Teichert (Reference Moore and Teichert1978). Morphological terminology follows Ubaghs (Reference Ubaghs, Moore and Teichert1978a) and Ausich et al. (Reference Ausich, Brett, Hess, Simms, Hess, Ausich, Brett and Simms1999) with modifications as noted by Ausich et al. (Reference Ausich, Wilson and Vinn2015b) and Ausich (Reference Ausich2018). The plating of interrays is given by the number of plates in each range from the proximalmost plate to the last range before the tegmen. In the posterior interray, the primanal is indicated by ‘P,’ and the first interradial plate in regular interrays is indicated by ‘1.’ A question mark indicates that more distal plating is unknown. Abbreviations used in designating measurements include: ACH = aboral cup height; ACW = aboral cup width; BH = basal plate height; BW = basal plate width; CaH = calyx height; CaW = maximum calyx width; CrH = crown height; IH = infrabasal plate height; IW = infrabasal plate width; RH = radial plate height; RW = radial plate maximum width; RFW = radial facet width; and * = measurement incomplete or of a compressed specimen. All measurements are in millimeters.
Repositories and institutional abbreviations
Specimens were studied from: TUG = University of Tartu, Tartu, Estonia; and GIT = Department of Geology, Tallinn University of Technology, Tallinn, Estonia. Specimens illustrated in Figure 3 were uncoated when photographed, whereas those in Figure 5 were photographed after whitening with ammonium chloride sublimate.

Figure 3. Oepikicrinus perensae n. gen. n. sp., new eucamerate from the Llandovery of Estonia: (1) lateral view of two paratypes, TUG 999-1-1 and 999-1-2; (2) lateral view of partially disarticulated paratype GIT 405-254-3; (3) lateral view of holotype GIT 405-254-1, with complete arms, also note long pinnules; (4) lateral view of compacted paratype GIT 405-254-2, with proximal arms. Scale bars = 1.0 mm (2); 2.5 mm (1, 3, 4).
Systematic paleontology
Class Crinoidea Miller, Reference Miller1821
Subclass Camerata Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Infraclass Eucamerata Cole, Reference Cole2017
Order Diplobathrida Moore and Laudon, Reference Moore and Laudon1943
Family Dimerocrinitidae von Zittel, Reference von Zittel1879
Remarks
As defined in Ubaghs (Reference Ubaghs, Moore and Teichert1978b), the Dimerocrinitidae is one of three Ordovician diplobathrid families that survived through the end-Ordovician extinctions and diversified during the Llandovery (Ausich and Deline, Reference Ausich and Deline2012). The oldest members of the Dimerocrinitidae are Eodimerocrinites Donovan and Gilmour, Reference Donovan and Gilmour2003, Euptychocrinus Brower, Reference Brower1994, and Ptychocrinus Wachsmuth and Springer, Reference Wachsmuth and Springer1885 from Laurentia (Katian of Scotland and the United States). During the Katian, this family persisted on Laurentia and migrated to northern Gondwana (Rosfacrinus Le Menn and Spjeldnaes, Reference Le Menn and Spjeldnaes1996; Morocco). The family persisted in Laurentia into the Hirnantian.
During the recovery from end-Ordovician extinctions, the family rapidly diversified during the Llandovery in both Laurentia and Baltica. Three Llandovery genera are known from Laurentia (Ausich and Copper, Reference Ausich and Copper2010) and now, three additional genera are recorded from Baltica. Baltic taxa are Hiiumaacrinus (Rhuddanian), Kalanacrinus (Aeronian) and Oepikicrinus n. gen. (Aeronian) (Ausich and Wilson, Reference Ausich and Wilson2016; Ausich et al., Reference Ausich, Wilson and Tinn2019). Both Kalanacrinus and Oepikicrinus n. gen. occur in strata in the Kalana Quarry of central Estonia, with Kalanacrinus from the Kalana Konservat Lagerstätte and Oepikicrinus n. gen. from younger Aeronian strata. After the Llandovery, dimerocrinitids continued to diversify (Ausich and Deline, Reference Ausich and Deline2012). Dimerocrinitids reappeared in northern Gondwana during the Wenlock (Dimerocrinites Phillips, Reference Phillips and Murchison1839; Le Menn et al., Reference Le Menn, Gourvennec, Piçarra and Robardet2003). The youngest dimerocrinitids are Devonian.
Genus Oepikicrinus new genus
Type species
Oepikicrinus perensae n. gen. n. sp., by monotypy.
Diagnosis
Dimerocrinitid with medium-sized, cone-shaped calyx, smooth plate sculpturing, infrabasal plates visible in lateral view (15% of CaH), anitaxial ridge absent with anitaxis plates broadly convex, interray regions slightly depressed, first primibrachial tetragonal, primaxil pentagonal, no axillary secundibrachial, fixed pinnules absent, 10 free arms, each free arm with one isotomous division, rectilinear uniserial pinnulate brachials, and circular columnals.
Occurrence
Silurian (Llandovery, Aeronian), Estonia (Baltica paleocontinent).
Etymology
The name for this new genus recognizes Armin Öpik (1898–1983) for his significant contributions to understanding fossil echinoderms from Estonia.
Remarks
Oepikicrinus n. gen. is compared to all other Silurian Dimerocrinitidae in Supplemental Table 1. It is distinguished from other Estonian taxa as follows. Hiiumaacrinus has a medium-sized, cone-shaped calyx, smooth calyx plate sculpturing, infrabasal plates not visible in lateral view, anitaxial ridge absent, depressed interradial regions on calyx, first primibrachial tetragonal to hexagonal, primaxil heptagonal, second secundibrachial axillary, fixed pinnules absent, 20 free arms (free arm branching and free arm brachials unknown), and a circular column. Kalanacrinus has a high cone- to bowl-shaped calyx, nodose calyx plate sculpturing, distal tips of the infrabasal plates visible in lateral view (presence of anitaxial ridge unknown), interradial regions not depressed, first primibrachial hexagonal, primaxil heptagonal, third secundibrachial axillary, fixed pinnules present, 20 free arms, free arms atomous, brachials uniserial, and a circular column. In contrast, Oepikicrinus n. gen. has a medium-sized, cone-shaped calyx, smooth plate sculpturing, infrabasal plates visible in lateral view (15% of CaH), anitaxial ridge absent with anitaxis plates broadly convex, interray regions slightly depressed, first primibrachial tetragonal, primaxil pentagonal, no axillary secundibrachials, fixed pinnules absent, 10 free arms, each free arm with one isotomous division, rectilinear uniserial pinnulate brachials, and circular columnals.
Oepikicrinus perensae new species
Figures 3, 4.1

Figure 4. Plate diagrams of Oepikicrinus n. gen. and Rozhnovicrinus n. gen.: (1) exploded plate diagram that is an interpretation of plating for Oepikicrinus n. gen.; (2, 3) Rozhnovicrinus isakarae n. gen. n. sp.: (2) plate diagram of GIT 405-252-1, also note unusual posterior interray plating and peneplenary radial facets; (3) drawing of holotype GIT 405-252-2. Black filling = radial plates; cross-ruled pattern = radianal plate; dotted pattern = additional anal plates; P = primanal. Scale bars = 1.0 mm.
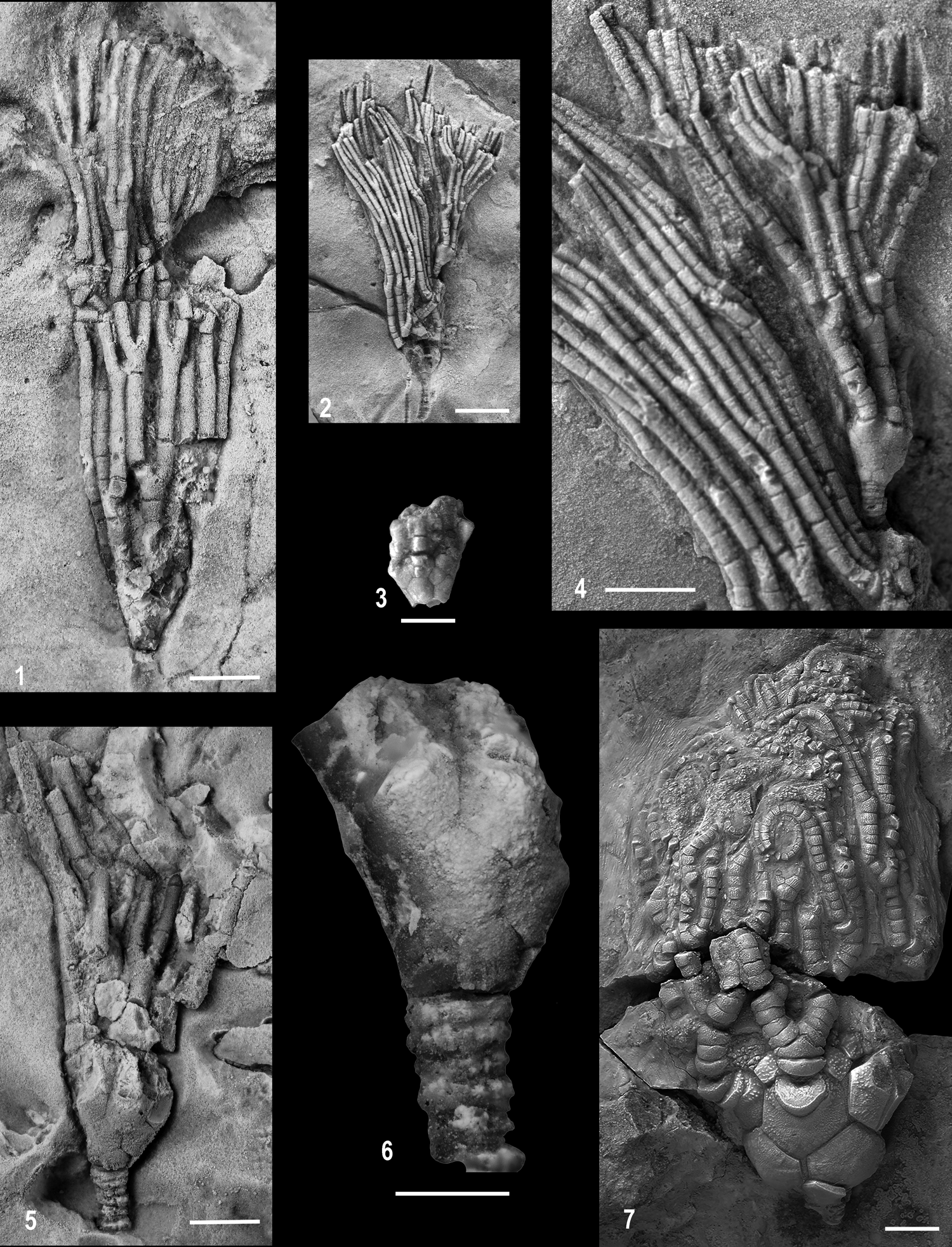
Figure 5. Lateral views of new eucladid crinoids from the Llandovery of Estonia: (1–6) Rozhnovicrinus isakarae n. gen. n. sp.: (1) crown with damaged aboral cup, paratype TUG 1329-14-1; (2) two specimens, the larger with only an impression of the aboral cup is paratype GIT 405-252-1, and the smaller complete specimen is holotype GIT 405-252-2; (3) D-ray lateral view of aboral cup of paratype GIT 405-252-1; (4) enlargement of holotype GIT 405-252-2 and arms of paratype GIT 405-252-1 (see Fig. 5.2); (5, 6) paratype TUG 1329-14-4: (5) aboral cup and proximal arms; (6) enlargement of aboral cup; (7) Euspirocrinus hintsae n. sp., holotype GIT 405-256, note distal coiling of arms. Scale bars = 2.5 mm (1, 3–6); 5.0 mm (2, 7).
Holotype
GIT 405-254-1. Type locality: Nurmekund Formation; Silurian (Llandovery, Aeronian); unknown quarry in the vicinity of Jõgeva, Estonia.
Diagnosis
See generic diagnosis above.
Occurrence
Oepikicrinus perensae n. gen. n. sp. is known with certainty from the Nurmekund Formation in the Kalana (TUG 1736, 1372) and Rõstla (TUG 999) Quarries. It was also collected from the Nurmekund Formation in an unknown quarry in the vicinity of Jõgeva (personal communication, H. Perens, 2019) (GIT 405-254).
Description
Crown and aboral cup small to medium-sized; calyx medium cone-shaped, as high as wide; arms not grouped; calyx plate sculpturing smooth. Infrabasal circlet ~15% of calyx height (Fig. 3.1); presumably five infrabasal plates. Basal circlet ~30% of calyx height (Fig. 3.2); presumably five basal plates hexagonal except CD basal heptagonal. Radial circlet ~30% of calyx height; radial plates presumably five, pentagonal, approximately as wide as high; approximately same size as basal plates. Normal interrays very poorly preserved but interpreted to have only the first interradial plate between the first primibrachials. Primanal separating C and D radial plates (Fig. 3.4), hexagonal, CD interray wider than normal interrays; plating P-1-1 (Fig. 4.1), anitaxis broadly convex. First primibrachial fixed, tetragonal wider than high, smaller than radial plates; second primibrachial beginning of free arms, axillary, pentagonal, larger than first primibrachial. Tegmen unknown. Free arms ten, one isotomous branching (Fig. 3.3), second primibrachial (first free brachial) axillary. Brachial plates rectilinear uniserial, first secundibrachial higher than wide, subsequent secundibrachials progressively lower so by at least mid-arm brachials wider than high. Arms with long pinnules. Column circular, holomeric, and heteromorphic. Proxistele with one internodal alternating with nodals.
Etymology
This species recognizes Helle Perens, whose work on Silurian geology of Estonia was instrumental to this study.
Materials
Holotype GIT 405-254-1 and paratypes GIT 405-254-2, 405-254-3, TUG 999-1-1, and 999-1-2; specimens questionably assigned to this taxon include TUG 1736-19-1, 1736-19-2, and 1372-2.
Measurements
Holotype GIT 405-254-1: CrH 13.6, CaH 3.0, CaW 3.0; paratype TUG 999-1-1: CrH 10.0, CaH 3.9, CaW 3.8.
Remarks
As noted above, Oepikicrinus perensae n. gen. n. sp. is a relatively small Dimerocrinitidae and further expands diversification of this family in Baltica during the Llandovery.
Parvclass Cladida Moore and Laudon, Reference Moore and Laudon1943
Magnorder Eucladida Wright, Reference Wright2017
Superorder Cyathoformes Wright et al., Reference Wright, Ausich, Cole, Peter and Rhenberg2017
Family Euspirocrinidae Bather, Reference Bather1890
Genus Euspirocrinus Angelin, Reference Angelin1878
Type species
Euspirocrinus spiralis Angelin, Reference Angelin1878, by monotypy.
Other species
Euspirocrinus cirratus Strimple, Reference Strimple1963; Euspirocrinus gagnoni Ausich and Copper, Reference Ausich and Copper2010; Euspirocrinus girvanensis Donovan, Reference Donovan1992; Euspirocrinus heliktos Ausich, Reference Ausich1986b; Euspirocrinus wolcottense Eckert and Brett, Reference Eckert and Brett2001; and Euspirocrinus hintsae n. sp.
Occurrence
Ordovician (Hirnantian) to Silurian (Ludlow), Laurentia and Baltica.
Remarks
Euspirocrinus hintsae n. sp. is the first report of Euspirocrinus outside of the Laurentian paleocontinent.
Euspirocrinus hintsae new species
Figure 5.7
Holotype
GIT 405-256. Type locality: Varbola Formation, Silurian (Llandovery, Rhuddanian), Reinu Quarry, Rapla County, Estonia.
Diagnosis
Euspirocrinus with low bowl-shaped aboral cup, fine pustulose sculpturing on aboral cup plates and arms, size of anal sac plates and number of plates in posterior interray unknown, two or three primibrachials, arms branching with poor isotomy proximally and endotomously distally, keels or nodes on brachials absent; column pentameric, heteromorphic; column length unknown.
Occurrence
Varbola Formation, Juuru Regional Stage (Rhuddanian, Llandovery, Silurian), Reinu Quarry, Rapla County, Estonia.
Description
Crown medium-sized (Fig. 5.7). Aboral cup low bowl-shaped, width to height ratio ~1.7, plates gently convex, fine pustulose surface sculpturing. Infrabasals probably five. Basals probably five, hexagonal, approximately the same size as radial plates. Radial plates probably five, pentagonal, slightly higher than wide. Radial facets angustary, horseshoe-shaped, declivate, occupying ~70% of distal radial plate width, deep V-shaped adoral groove; radial facet topography not known. Posterior interray and oral region not preserved. Arms branching four times with poor isotomy proximally and endotomous branching distally (Fig. 5.7). First primibrachial much wider than high, primibrachial two or three axillary, primaxil 2.0 times wider than high. First secundibrachials in lateral contact medially, secundibrachial three or four axillary; first tertibrachials in lateral contact medially, where branched tertibrachial four to seven axillary. All brachials rectangular uniserial, aborally convex, with same sculpturing as calyx plates, nonaxillary brachial dimensions variable but commonly ~2.0 times higher than wide. Distal arms planispirally coiled inward. Column circular, heteromorphic, and pentameric; nodals convex laterally; other details of column unknown.
Etymology
This new species name recognizes Linda Hints, who has made significant contributions to Estonian paleontology and who found the holotype of this new taxon.
Measurements
Holotype GIT 405-256: CrH 47.4*; ACH 11.5*; ACW 16.2*; BH 6.4; BW 7.0; RH 9.1; RW maximum 9.1; RFW 3.9.
Remarks
During the Ordovician, Euspirocrinus was present only on Laurentia. Euspirocrinus hintsae n. sp. is the earliest known occurrence of this genus outside of Laurentia, and this genus persisted into the Wenlock of Baltica (Sweden). The characteristic angustary facets and planispirally coiled distal arms are common among all species of this genus.
The aboral cup with a low bowl shape, finely pustulose aboral cup plates, arm branching poorly isotomous proximally and endotomous distally, and pentameric column are features that distinguish Euspirocrinus hintsae n. sp. from other species of this genus (Ausich and Copper, Reference Ausich and Copper2010, table 30). It is most similar to Euspirocrinus wolcottense from the Walcott Formation (late Llandovery, New York), however, the latter has an aboral cup with a medium bowl shape, ridges and pustulose plate sculpturing, weakly endotomous arm branching, and pentameric only in the distal column.
Family Dendrocrinidae Wachsmuth and Springer, Reference Wachsmuth and Springer1886
Genus Rozhnovicrinus new genus
Type species
Rozhnovicrinus isakarae n. sp., by monotypy.
Diagnosis
Dendrocrinidae with a medium cone-shaped aboral cup, smooth plate sculpturing, angustary radial facets, the radianal below and to the left of the CD basal plate, fixed plates in regular interrays absent, five primibrachials, arm branching isotomous, column heteromorphic, column circular, column construction holomeric, and unknown lumen shape.
Occurrence
Silurian (Llandovery, Aeronian), Estonia (Baltica paleocontinent).
Etymology
The name for this crinoid genus recognizes Sergei V. Rohznov for his significant contributions to the understanding of fossil echinoderms from Estonia.
Remarks
Rozhnovicrinus n. gen. has a unique anal plate arrangement for an Ordovician-Silurian dendrocrinid. The radianal is beneath the left side of the C radial plate and above and to the right of the CD basal plate. However, the radianal plate is one of three plates in sutural contact with the CD basal plate, which also has a large plate above and to the left and a small plate immediately above the CD basal plate. This small plate has a very short suture with the CD basal plate, and it separates the other two plates in sutural contact with the CD basal plate.
Rozhnovicrinus n. gen. is compared to all Ordovician and Silurian Dendrocrinidae in Supplemental Table 2, and comparisons to other Llandovery Dendrocrinidae are given below. Dendrocrinus Hall, Reference Hall1852 has a medium cone-shaped aboral cup, smooth plate sculpturing, angustary radial facets, the radianal in contact with the CD basal plate, radianal directly beneath the C radial plate, fixed plates in regular interrays absent, three to seven primibrachials, arm branching isotomous, column xenomorphic to homeomorphic, column circular to pentagonal, column construction pentameric or holomeric, and lumen circular. Aetocrinus Ausich and Copper, Reference Ausich and Copper2010 has a low cone-shaped aboral cup, smooth plate sculpturing, angustary radial facets, the radianal in contact with the infrabasals, radianal beneath and to the left of the C radial plate, fixed plates in regular interrays absent, five to nine primibrachials, arm branching isotomous, column heteromorphic, column circular, column construction holomeric, and (lumen shape unknown). Laurucrinus Ausich and Copper, Reference Ausich and Copper2010 has a medium cone-shaped aboral cup, smooth plate sculpturing, angustary radial facets, the radianal is below and to the left of the CD basal plate, fixed plates in regular interrays absent, three or four primibrachials, arm branching isotomous, column heteromorphic, column pentagonal, column construction holomeric, and (lumen shape unknown). Rozhnovicrinus n. gen. has a medium cone-shaped aboral cup, smooth plate sculpturing, angustary radial facets, the radianal is below and to the left of the CD basal plate, fixed plates in regular interrays absent, five primibrachials, arm branching isotomous, column heteromorphic, column circular, column construction holomeric, and unknown lumen shape.
Rozhnovicrinus isakarae new species
Figures 4.2, 4.3, 5.1–5.6
Type material
Holotype GIT 405-252-2. Nurmekund Formation, Silurian (Llandovery, Aeronian), Ellakvere Quarry, central Estonia. Paratypes: GIT 405-252-1, TUG 1329-14-1, and 1329-14-4.
Diagnosis
See generic diagnosis above.
Occurrence
This new crinoid is from the Aeronian Nurmekund Formation (Llandovery, Aeronian); in the Kalana (TUG 1329) and Ellakvere (GIT 405-252) Quarries in central Estonia.
Description
Crown small, conical to subcylindrical (Fig. 5.1). Aboral cup small, medium cone-shaped (Fig. 5.6), width to height ratio ~1.0, plates gently convex, smooth. Infrabasal plates five, equal in size; infrabasal circlet ~20% of aboral cup height, base truncate (Fig. 4.3). Basal plates five, hexagonal, CD basal larger than others, somewhat higher than wide, larger than radials; basal circlet ~45% of aboral cup height. Radials five, hexagonal except D radial that is ~1.5 times wider than as high; radial circlet ~35% of aboral cup height. Radial facets peneplenary (Fig. 5.3), horseshoe shape, declivate, shallow V-shaped adoral groove; radial facet topography not known. Five anal plates in or partially in the aboral cup. Three anal plates above the CD basal plate (Fig. 4.2). Radianal plate on upper right shoulder of CD basal and below toward left of C radial plate; slightly wider than high, diamond shape. Anal X on left upper shoulder of CD basal plate and below to left of D radial plate but not in sutural contact with the radianal plate, diamond-shaped, higher than wide. Right tube plate between radianal and anal X with short suture below to CD basal plate, heptagonal, approximately as high as wide. Radianal and right tube plate support a series of plates leading to the anal sac. Tegmen not known. Anal sac shorter than arms, comprised of numerous small, slightly concave, hexagonal plates (as known). Arms branching three times with poor isotomy (Fig. 5.2, 5.4, 5.5). First and second primibrachial as high as wide, more distal brachials higher than wide; primibrachial five axillary; secundibrachial five axillary, secundibrachial five or six axillary; tertibrachials eight to 12 axillary. All brachials rectangular uniserial, aborally convex. Where preserved, 2.5 ambulacral cover plates per brachial. Column circular, holomeric, and heteromorphic; other details of column unknown.
Etymology
The species name recognizes Mare Isakar for her significant and continuing contributions to the paleontology of Estonia.
Materials
The holotype GIT 405-252-2 is a small specimen with some arms and a short length of poorly preserved column. Paratypes plus one additional specimen (GIT 405-252-3).
Measurements
Holotyoe GIT 405-252-2: CrH 28.4, ACH 2.8, ACW 2.7, BH 1.4, BW 1.2, RH 1.0, RW 1.2, RFW 0.8. Paratype GIT 405-252-1: CrH 13.2, ACH 1.9, ACW 1.9, IH 0.5, IW 0.4, BH 0.8, BW 0.8, RH 0.8, RW 0.8, RFW, 0.5; paratype TUG 1329-14-1: CrH 22.0; paratype TUG 1329-14-4: IH 1.8, IW 1.6, BH 2.5, BW 2.5, RH 2.5, RW 3.3.
Remarks
With three plates in sutural contact with the CD basal plate, Rozhnovicrinus n. gen. is an unusual Silurian cladid that can be interpreted in more than one way. The preferred interpretation in the description above accounts for all of the plates. In other cladids, the anal X is in sutural contact with the radianal plate, the right tube plate is not in sutural contact with the CD basal, and the plate above the right tube plate does not share a suture with both the right tube plate and the radianal. This is a unique CD interray plating and is the basis by which the new genus is defined. The combination of a radianal plate below to the left of the C radial plate, having the anal X to the left rather than to the left and above the radianal, and more than three anal plates fixed in the aboral cup ally this specimen to the Euspirocrinidae, as do the relatively large, horseshoe-shaped, declivate radial facets.
Accessibility of supplemental data
Diagnostic tables for new genera are available from Dryad Digital Repository: https://doi.org/10.5061/dryad.33622fn.
Acknowledgments
We thank the following groups for their support that made this work possible: National Geographic Society (NGS 60031112; to WIA and MAW); Ohio State University Emeritus Academy (to WIA); and the Luce Fund at The College of Wooster (to MAW). M. Isakar, O. Tinn, and O. Vinn were instrumental in developing our knowledge of Estonian geology and paleontology as was G. Baranov for the photos of echinoderms. The authors also thank S.R. Cole and S.K. Donovan for their thoughtful reviews.






