



Introduction
Dissorophidae is a Pennsylvanian–Permian clade of temnospondyl stem amphibians that are found in fluvial and floodplain deposits of the southwestern United States, Central Europe, European Russia, and north-central China (Eichwald, Reference Eichwald1848; Cope, Reference Cope1895; Carroll, Reference Carroll1964a; Gubin, Reference Gubin1980; Witzmann, Reference Witzmann2005; Schoch and Milner, Reference Schoch and Milner2014; Liu, Reference Liu2018). They belong to the Dissorophoidea, which most authors consider the clade from which some or all extant lissamphibians originated (Bolt, Reference Bolt1969; Anderson et al., Reference Anderson, Reisz, Scott, Fröbisch and Sumida2008a; Pardo et al., Reference Pardo, Small and Huttenlocker2017). Recently, new dissorophid taxa have been reported and long-known taxa revised, paving the way for phylogenetic analyses of the group (Reisz et al., Reference Reisz, Schoch and Anderson2009; Schoch, Reference Schoch2012; Holmes et al., Reference Holmes, Berman and Anderson2013; Maddin et al., Reference Maddin, Fröbisch, Evans and Milner2013; Schoch and Sues, Reference Schoch and Sues2013; Gee, Reference Gee2018; Gee and Reisz, Reference Gee and Reisz2018).
Here we focus on a long-neglected dissorophid, Parioxys ferricolus Cope, Reference Cope1878, whose affinities have remained unclear. In a series of publications, Moustafa (Reference Moustafa1952a, Reference Moustafa1955a, Reference Moustafab) worked out many anatomical details of this species and considered P. ferricolus an eryopiform (in modern parlance). Basing his work largely on Eryops megacephalus, he traced in ink what he considered sutures on the roughly prepared adult skull (MCZ 1162; Fig. 1.8, 1.9). Only after much further work on dissorophoids by Carroll (Reference Carroll1964a) and later authors did the skeletal features become sufficiently understood to permit referral of P. ferricolus to Dissorophoidea, which was first explicitly done by Schoch and Milner (Reference Schoch and Milner2014), who listed it as a eucacopine dissorophid because of features shared particularly with Cacops spp.

Figure 1. Parioxys ferricolus Cope, Reference Cope1878, skulls. (1, 2) AMNH 2445: (1) photograph in dorsal view; (2) drawing of same specimen. (3, 4) AMNH 4309 (holotype): (3) photograph; (4) drawing. (5) AMNH 2445, lateral view. (6, 7) USNM 544065: (6) photograph; (7) drawing. (8, 9) MCZ 1162: (8) photograph; (9) drawing. d = dentary; f = frontal; ju = jugal; la = lacrimal; lep = laterally exposed palatine; m = maxilla; n = nasal; nar = naris; p = parietal; pf = postfrontal; pm = premaxilla; pp = postparietal; prf = prefrontal; q = quadrate; qj = quadratojugal; sm = septomaxilla; st = supratemporal; ta = tabular.
The objective of the present study is: (1) to review the existing material of P. ferricolus, part of which has been further prepared, with emphasis on previously unreported cranial features and (2) to assess the phylogenetic position of this taxon.
Materials and methods
During this study, material from three institutions was examined.
Material examined
Four skulls of different sizes (AMNH 2445, 4309; MCZ 1162; USNM 544065), a range of skull fragments (MCZ 1162a, 1162b), and various postcranial bones (MCZ 1162; USNM 544065) form the basis of the present study. Specimens were examined with a WILD M3Z binocular microscope. Illustrations were based on photographs taken with a Panasonic Lumix DMC-TZ22.
The material is moderately well preserved, but preparation is difficult because the bones are covered by a tightly adhering carbonatic crust that cannot be removed entirely without damaging the underlying bone surface. Therefore, sutures are not visible in AMNH 2445 and 4309 (Fig. 1) and only partially exposed in USNM 544065 (Figs. 1, 2). In MCZ 1162, where the dermal ornament and sutures are better exposed, sutures were drawn on the skull by Y.S. Moustafa during his dissertation research; however, some of them could not be confirmed by the present study.
Repositories and institutional abbreviations
AMNH, American Museum of Natural History, New York; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts; USNM, National Museum of Natural History, Smithsonian Institution, Washington, D.C.
In preparing the description, we used the following studies for comparative data: Actiobates peabodyi Eaton, Reference Eaton1973 (Gee and Reisz, Reference Gee and Reisz2020), Anakamacops petrolicus Li and Cheng, Reference Li and Cheng1999 (Liu, Reference Liu2018), Broiliellus texensis Williston, Reference Williston1914, Cacops spp. (Williston, Reference Williston1910; Reisz et al., Reference Reisz, Schoch and Anderson2009; Gee and Reisz, Reference Gee and Reisz2018), Conjunctio multidens Carroll, Reference Carroll1964a (Schoch and Sues, Reference Schoch and Sues2013), Dissorophus multicinctus Cope, Reference Cope1895 (DeMar, Reference DeMar1968; Schoch, Reference Schoch2012), Kamacops acervalis Gubin, Reference Gubin1980 (Schoch, Reference Schoch1999), Mattauschia laticeps Milner, Reference Milner2019, Palodromeus bairdi Schoch, Henrici, and Hook, Reference Schoch, Henrici and Hook2020, Scapanops neglecta Schoch and Sues, Reference Schoch and Sues2013, and Tersomius texensis Case, Reference Case1910 (Carroll, Reference Carroll1964a; Maddin et al., Reference Maddin, Fröbisch, Evans and Milner2013).
Systematic paleontology
Temnospondyli Zittel, Reference Zittel1888
Rhachitomi Watson, Reference Watson1919 sensu Schoch, Reference Schoch2013
Dissorophoidea Bolt, Reference Bolt1969
Olsoniformes Anderson et al., Reference Anderson, Henrici, Sumida, Martens and Berman2008b
Dissorophidae Boulenger, Reference Boulenger, Wallace, Hadley and Chisolm1902
Eucacopinae Schoch and Sues, Reference Schoch and Sues2013
Parioxys Cope, Reference Cope1878
Type species
Parioxys ferricolus Cope, Reference Cope1878.
Diagnosis
(1) Intercentra closed dorsally to form complete rings, (2) adductor ridge of femur not bifurcated at proximal end, (3) length of tibia 60% of femur length (based on MCZ 2262; Moustafa, Reference Moustafa1955a; Carroll, Reference Carroll1964a; Schoch and Milner, Reference Schoch and Milner2014).
Occurrence
Early Permian (Cisuralian); Texas, United States of America.
Remarks
Moustafa (Reference Moustafa1955b) named a second species of Parioxys, P. romeri, on the basis of the distal end of a humerus (MCZ 1965) from the Petrolia Formation near Dundee, Archer County, Texas. The validity of this taxon is questionable. Carroll (Reference Carroll1964b) proposed a third species of Parioxys, P. bolli, on the basis of a partial postcranial skeleton (AMNH 7118) from the Archer City Formation near Archer City, Archer County, Texas. He noted that only a few features distinguished Parioxys bolli from P. ferricolus.
Parioxys ferricolus Cope, Reference Cope1878
Figures 1–4

Figure 2. Detailed drawings of Parioxys ferricolus Cope, Reference Cope1878. (1) USNM 544065 in dorsal view. (2) MCZ 1162 in dorsal view.

Figure 3. Parioxys ferricolus Cope, Reference Cope1878. (1, 2) Palate in ventral view. MCZ 1162: (1) photograph; (2) drawing. (3, 4) USNM 544065: (3) photograph; (4) drawing. (5) MCZ 1162a, drawing of parasphenoid in ventral view. (6) MCZ 1162b, drawing of marginal palate in ventral view. a = angular; bp = basipterygoid joint; ch = choana; cp = cultriform process; d = dentary; e = ectopterygoid; ef = ectopterygoid fang; eo = exoccipital; m = maxilla; mt = maxilla tooth; oc = occipital condyle; ot = otic region; pl = palatine; plf = palatine fang; pm = premaxilla; pt = pterygoid; se = sphenethmoid; ub = unidentified bone; vo = vomer; vod = vomerine depression; vof = vomerine fang; vot = vomerine tooth row.
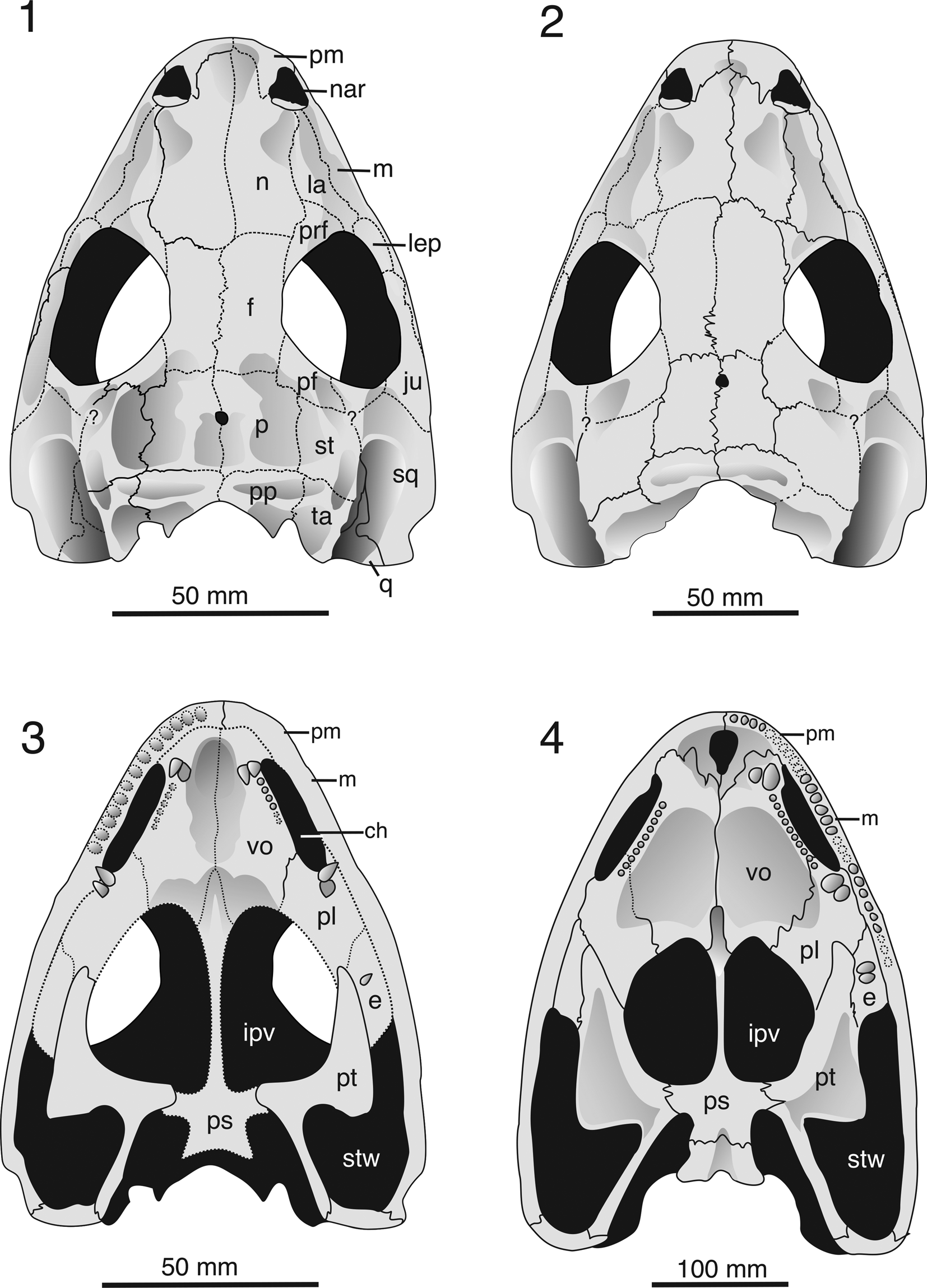
Figure 4. (1–3) Parioxys ferricolus Cope, Reference Cope1878. Reconstructions of skulls: (1) large juvenile skull based on USNM 544065, dorsal view; (2) adult skull based on MCZ 1162, dorsal view; (3) Juvenile skull based largely on USNM 544065, supplemented by MCZ 1162, ventral view. (4) Kamacops acervalis, ventral view (modified from Schoch, Reference Schoch1999). ch = choana; e = ectopterygoid; f = frontal; ipv = interpterygoid vacuity; ju = jugal; la = lacrimal; lep = laterally exposed palatine; m = maxilla; n = nasal; nar = naris; p = parietal; pf = postfrontal; pl = palatine; pm = premaxilla; pp = postparietal; prf = prefrontal; ps = parasphenoid; pt = pterygoid; q = quadrate; sq = squamosal; st = supratemporal; stw = subtemporal window; ta = tabular; vo = vomer.
Holotype
AMNH 4309, poorly preserved, slightly distorted skull (120 mm length), damaged by crude preparation (Moustafa Reference Moustafa1952a, Reference Moustafa1955a, Reference Moustafab; Schoch and Milner, Reference Schoch and Milner2014). Moustafa (Reference Moustafa1952a) identified this specimen as the lectotype from two specimens that Case (Reference Case1911) considered syntypes of Parioxys ferricolus and referred the other skull (AMNH 4310) to Eryops.
Diagnosis
Autapomorphies: (1) length of preorbital region 1.5 times that of posterior skull table in juveniles and twice that in adults; (2) parietal only two-thirds the length of the frontal; (3) interpterygoid vacuities shorter than half skull length; (4) choana extended posteriorly; (5) vomer with enlarged, V-shaped median depression; (6) palatine ramus of pterygoid anteriorly broadened to wedge between palatine and ectopterygoid.
Synapomorphies with Cacops spp. and Kamacops acervalis: lacrimal laterally constricted by wide maxillary shelf, narrower than alary process; LEP large, oval feature with concave dorsal surface; and tabular extended posteriorly.
Plesiomorphies (contrasting the condition in Cacops spp., Kamacops acervalis, and Anakamacops petrolicus): interorbital distance of adults wide (0.29); tabular not contacting or fused to quadrate.
Occurrence
Near Mt. Barry, Wichita County, Texas, USA; Petrolia Formation, Wichita Group, early Permian (Cisuralian). The referred specimens MCZ 1162 and USNM 544065 were collected from the Petrolia Formation at Dundee in Baylor County, Texas. The tetrapod assemblage from the Petrolia Formation is part of the Seymouran Land Vertebrate Faunachron sensu Lucas (Reference Lucas, Lucas, Cassinis and Schneider2006).
Description
Parioxys ferricolus has a moderately wide skull (width/length ratio = 0.92), which is narrower than in dissorophines, with relatively large orbits (orbit length/skull length ratio = 0.32) and a tapering preorbital region. The proportions of the skull roof (Figs. 1, 2) are intermediate between those of Conjunctio multidens and Cacops morrisi Reisz, Schoch, and Anderson, Reference Reisz, Schoch and Anderson2009. In the holotype, which is a small juvenile, the preorbital region measures 1.35 times the length of the postorbital skull table. This ratio increases in larger skulls (USNM 544065: 1.5; MCZ 1162: 2.0).
The preorbital region tapers toward the relatively narrow tip, which at the posterior level of the nares measures only 0.55–0.58 times the width of the skull level with the anterior orbital margin. This feature is isometric throughout the preserved size range. The external naris is like that of K. acervalis and Cacops spp. in being confined to the tip of the snout, contrasting with the much larger opening in Conjunctio multidens. There is a shallow depression between the nares in some specimens, but no fontanelle (Figs. 1.7, 4.1). A wedge-shaped septomaxilla is preserved and well exposed on the posterior margin of the naris of MCZ 1162 (Fig. 1.9); it remains unclear whether the bone also included an internal narial portion as in some other temnospondyls.
The premaxilla bears five large teeth. The most completely preserved maxilla has 23 teeth (USNM 544065), with space for 10 additional ones, judging from their relatively equidistant implantation. The nasal, lacrimal, and prefrontal have proportions similar to those in C. morrisi, and the concave lateral suture of the lacrimal is particularly similar in these two taxa. A faint central depression is developed on the dorsal surface of the nasals and anterior portion of the frontals, but crushing may have somewhat altered the surface relief.
The pineal foramen is much smaller than in Cacops morrisi and Conjunctio multidens, and it is emplaced on a gentle eminence at the anterior margin of a medial depression (Fig. 1). The postparietal is short, its dorsal portion reaching only one-third the length of the parietal. In USNM 544065 and AMNH 2445, it bears a transverse ridge along the occipital margin that continues onto the tabular where it ends at the posterior tip of the tabular horn. In AMNH 4309 and MCZ 1162, the ridge is much lower and not continuous (Figs. 1, 2). In USNM 544065, the transverse ridge bifurcates on the tabular, with an anterior branch extending along the supratemporal and postfrontal, similar to the condition in Cacops morrisi.
The postorbital region of the skull has a slightly rounded, convex lateral margin. It is shorter and wider than in other eucacopines, with the length/width ratio of the postorbital skull region ranging from 0.43 in juveniles to 0.34 in adults. A contact between the postorbital and squamosal is probably present, considering the proportions of the bones, but not exposed. The supratemporal is longer and the parietal is anteriorly wider than in C. morrisi, both having rectangular outlines. The suture between the supratemporal and parietal is markedly curved in double S-shaped fashion in USNM 544065 (Fig. 1).
The squamosal and quadratojugal are not preserved in the larger specimens, but this region is known in AMNH 2445 (Fig. 1). In lateral view, the skull is dorsoventrally deepest at the level of the posterior margin of postparietals and continuously decreases in height toward the tip of the snout. The quadrate has a rudimentary dorsal process, much less developed than in juveniles of C. morrisi; thus, a late-ontogenetic closure of the squamosal embayment was absent in P. ferricolus, which differs strongly from the condition in Cacops spp., Kamacops acervalis, Anakamacops petrolicus, and Zygosaurus lucius Eichwald, Reference Eichwald1848.
Palate
The palate is morphologically intermediate between those of Cacops spp. and Kamacops acervalis in many features. Its entire preserved ventral surface is covered by closely set, uniformly-sized denticles (Fig. 2). The vomer, palatine, and ectopterygoid are almost completely preserved, but the pterygoid is known only by fragments of its palatine and basipterygoid rami in USNM 544065 and the outline of the quadrate ramus in MCZ 1162. The basal plate of the parasphenoid is well preserved in MCZ 1162a (Fig. 3.5), whereas most of the cultriform process remains unknown. Sutures between any palatal elements cannot be traced in USNM 544065, but there is a fragment of the marginal palate (MCZ 1162b) that preserves sutures well (Fig. 3.6).
The vomer is proportionately longer than in K. acervalis and more expanded posteriorly than in any other known dissorophoid except for Macrerpeton huxleyi Moodie, Reference Moodie1909 (Schoch and Milner, Reference Schoch and Milner2021). In USNM 544065, the complete vomerine dentition is exposed, covering the entire surface of the element. The medial region is strongly concave, spanning the entire length of the vomers. Although probably homologous with the anterior depression in C. morrisi, this depression is more comparable in size to the intervomerine depression of zatracheids than to that of dissorophoids (Witzmann and Schoch, Reference Witzmann and Schoch2006). The dentigerous lateral parts of the vomers are gently convex on the ventral side.
The long and narrow choanae are exposed in both MCZ 1162 and USNM 544065 (Fig. 3). This proportionately matches the length of the choana in K. acervalis and is much longer than in other eucacopines (Fig. 4).
The interpterygoid vacuity is smaller than in Cacops spp. but substantially larger than that in K. acervalis. It shares the straight sagittal lateral margin with both. The basipterygoid ramus, which is entirely covered with denticles, is shorter than in C. morrisi but not as abbreviated and plate-like as in K. acervalis. The dorsal side bears a large, raised socket for the epipterygoid. The palatine ramus of the pterygoid forms a broad, medially expanded plate with the palatine. This region is best preserved in the fragmentary specimen MCZ 1162b (Fig. 3.6), where the sutures are visible. The palatine is an abbreviated element, completely and widely separated from the interpterygoid vacuity by a broadened palatine ramus of the pterygoid (Figs. 3.6, 4.3). This ancestral condition is unparalleled in dissorophids, but a contact between pterygoid and vomer is common in trematopids (Polley and Reisz, Reference Polley and Reisz2011; Milner, Reference Milner2019). However, the marked broadening of the palatine ramus and its wedging between the palatine and ectopterygoid form apomorphic character states. The vomer, palatine, and ectopterygoid all bear large, recurved tusks, with those of the vomer slightly taller than the others (Fig. 4).
The parasphenoid is well preserved in MCZ 1162a, where the base of the very slender cultriform process is present. Consistent with Kamacops, the basal plate is very long with pronounced, offset posterolateral wings. Unlike the latter, the suture with the exoccipital is hardly visible, and the ventral surface of the plate bears a large inverted-V-shaped depression. Otherwise, this region is smooth without tooth patches, and foramina for the carotid artery are absent. The basipterygoid process is also clearly offset and appears to have been less firmly sutured with the pterygoid than in other eucacopines.
Neurocranium
The sphenethmoid and otic region are exposed in MCZ 1162 (Fig. 3.2). The sphenethmoid is relatively wide but ends well anterior to the otic region. The otic was fully ossified, without separation into pro- and opisthotic portions.
Visceral skeleton
The stapes and elements of the hyobranchial apparatus remain unknown.
Mandible
Moustafa (Reference Moustafa1955a) figured and described the mandible in detail. The lower jaw agrees in most features with that in C. morrisi (Gee and Reisz, Reference Gee and Reisz2018). The posterior half of the jaw is lower dorsoventrally than in C. morrisi, and its ventral margin is almost straight. The angular is not as expanded dorsally, but the coronoid process is taller and more rounded than in C. morrisi, with a continuously sloping anterior margin rather than forming an abrupt edge. The dentary bears at least 26 large conical and recurved teeth. The size of the teeth decreases somewhat posteriorly, where they have broader bases. The adductor fenestra has a low lingual margin formed by the prearticular. The size of the coronoid and splenial was correctly illustrated by Moustafa (Reference Moustafa1955a).
Postcranial skeleton
MCZ 1162 comprises a set of at least eight partial skeletons (Moustafa, Reference Moustafa1952b). As Moustafa (Reference Moustafa1955a) already illustrated many postcranial elements in detail, we do not provide figures here. He emphasized the rhachitomous structure of the anterior vertebral column, with semilunar intercentra; in the posterior portion, they are higher and nearly form closed discs, consistent with the condition in other dissorophids (Moustafa, Reference Moustafa1955b, pl. 4). The previously undescribed USNM 544065 includes fragments of limb bones and a complete right half of the pelvis. The girdles and limbs include some interesting features that differ from the corresponding ones in Cacops morrisi. The scapula is dorsoventrally lower in P. ferricolus, dorsally expanded, and co-ossified with the coracoid. The humerus is more robust than in C. morrisi, but its articular ends are not as wide as in C. aspidephorus Williston, Reference Williston1910. The supinator process is more clearly defined than in the two species of Cacops. In the fully ossified pelvis, the ilium has an anterodorsally expanded portion and the dorsal margin slopes posteroventrally, forming a posterior process. The ischium is elongate and bears a posterolateral process, and the pubis has a slight anterodorsal extension.
Remarks
None.
Phylogenetic relationships
To assess the phylogenetic position of Parioxys ferricolus, we performed a phylogenetic analysis using the character–taxon matrix for dissorophoids recently published by Schoch et al. (Reference Schoch, Henrici and Hook2020; see supplementary Appendix).
Phylogenetic analysis
The analysis was performed using TNT 1.5 (available at https://cladistics.org/tnt/), under the New Technology Search option (Fig. 5). It recovered five equally most parsimonious trees (tree length [TL] = 291 steps; consistency index [CI] = 0.432; retention index [RI] = 0.76). The character descriptions and character–taxon matrix are provided at https://doi.org/10.5061/dryad.tb2rbp02d. All multistate characters were ordered. Dendrysekos helogenes Steen, Reference Steen1934 (=Dendrerpeton acadianum Owen, Reference Owen1853 of many former authors; see discussion in Schoch and Milner, Reference Schoch and Milner2014) was used as the outgroup. Scores for the following additional taxa were added: Anakamacops petrolicus (Gee and Reisz, Reference Gee and Reisz2020), Kamacops acervalis (Gubin, Reference Gubin1980; Schoch, Reference Schoch1999), Scapanops neglecta (Schoch and Sues, Reference Schoch and Sues2013), and Parioxys ferricolus (based on the present study).

Figure 5. Phylogeny of dissorophoid temnospondyls, with particular emphasis on the Dissorophidae and the position of Parioxys ferricolus Cope, Reference Cope1878.
In a variant analysis, the dissorophoid Reiszerpeton renascentis (Maddin et al., Reference Maddin, Fröbisch, Evans and Milner2013) was added, which has recently been found as a potential relative of eucacopines (Gee et al., Reference Gee, Berman, Henrici, Pardo and Huttenlocker2021). This analysis obtained four most parsimonious trees with poorer resolution of eucacopines (TL = 298 steps; CI = 0.42; RI = 0.75).
Phylogenetic position of Parioxys ferricolus
The topology of the trees is essentially like that found by Schoch (Reference Schoch2019) and Schoch et al. (Reference Schoch, Henrici and Hook2020). Perryella olsoni Carlson, Reference Carlson1987 nests at the base of Dissorophoidea, Micromelerpeton credneri Bulman and Whittard, Reference Bulman and Whittard1926 (representing Micromelerpetidae, which is not further discussed here) is recovered as the basalmost dissorophoid, followed by a clade Amphibamiformes + Olsoniformes. Amphibamiformes comprises four successive clades, Micropholidae (Eoscopus, Tersomius, Pasawioops, Micropholis), Amphibamidae (Platyrhinops, Amphibamus, Doleserpeton), Branchiosauridae (Apateon, Leptorophus, Schoenfelderpeton), and Lissamphibia plus its respective stem groups (Georgenthalia, Gerobatrachus, Eocaecilia, Karaurus, Triadobatrachus). The internal structure of Olsoniformes is not fully resolved, with Actiobates, Mattauschia, and Palodromeus forming an unresolved polytomy with the rest, followed by a second polytomy with Ecolsonia + Fedexia and Tambachia, Phonerpeton, and Acheloma, and finally Dissorophidae. The latter comprises two subclades, Dissorophinae with Broiliellus and Dissorophus and Eucacopinae, with the following branching sequence: (1) Scapanops neglecta, (2) Conjunctio multidens, (3) Anakamacops petrolicus, (4) Cacops morrisi, (5) Parioxys ferricolus, and (6) Kamacops acervalis.
The resolution of eucacopines is lowered when Reiszerpeton renascentis is added to the analysis, which results in a polytomy within that clade. In such a variant analysis, R. renascentis is not found to nest with dissorophids, but with Ecolsonia cutlerensis Vaughn, Reference Vaughn1969 within a poorly resolved basal part of the olsoniform tree, which probably highlights our poor knowledge of that taxon.
Discussion
Parioxys ferricolus shares many features with eucacopines and belongs to a more inclusive group of that clade comprising Cacops spp. and Kamacops acervalis. However, understanding the phylogeny of eucacopines is hampered by the fact that most taxa are still incompletely known: the postcranium is well preserved in only two taxa, Cacops morrisi and C. aspidephorus; the palate remains unknown in Scapanops neglecta; and the sutures are only partially documented for both Parioxys ferricolus and Anakamacops petrolicus and remain unknown in Kamacops acervalis.
The lost holotype of Zygosaurus lucius, a steinkern that lacked most of the bone from the middle Permian (Guadalupian) of Bashkortostan (Russia), is rather different from other eucacopines, but its otic notch closely resembles that in Cacops spp. (Eichwald, Reference Eichwald1848). Unlike all other eucacopines, Z. lucius and A. petrolicus share an enlarged otic notch that constricts the postorbital skull region laterally, but the orbits differ strongly in size.
Phylogenetic analysis (Fig. 6) indicates that Scapanops neglecta may be the basalmost eucacopine, but this remains tentative due to the large amount of missing data. In addition to the many unknown features, Scapanops neglecta has a highly autapomorphic skull roof, but it shares with eucacopines the abbreviated postparietal and internarial fenestra. Like Conjunctio multidens, it has numerous tiny maxillary teeth, indicating that eucacopines may initially have subsisted on small invertebrate prey, just as the basal olsoniform Palodromeus bairdi and amphibamiforms in general. Autapomorphic features of Scapanops neglecta are the transverse ridge on the posterior portion of the nasals and the dorsally placed, elongate oval external nares.

Figure 6. Character evolution in dissorophids, with particular emphasis on the eucacopines and Parioxys ferricolus.
A rather well-supported clade encompasses Conjunctio multidens, Cacops spp., Parioxys ferricolus, and Kamacops acervalis. C. multidens retained a relatively long postorbital skull table, like dissorophines, trematopids, and Palodromeus bairdi, but the general proportions of the orbits and snout are like those of Cacops spp. and Parioxys ferricolus.
Eucacopines more derived than Conjunctio multidens have fewer and larger marginal teeth and greatly enlarged, recurved palatal tusks, indicating a change to seizing and swallowing larger prey. Kamacops acervalis, the poorly known Iratusaurus vorax Gubin, Reference Gubin1980, and Anakamacops petrolicus probably had evolved to become major predators in their respective ecosystems.
Eucacopines more derived than Conjunctio have more robust skulls with a more pronounced surface relief. The eminences on the skull roof include a prominent ridge connecting the supratemporal with the postfrontal, which is also present in Dissorophus multicinctus, Cacops spp., and Parioxys ferricolus, whereas it extends further laterally across the postorbital in Conjunctio multidens. The tabular and postparietal flanges along the occipital margin increase in size from Conjunctio multidens, Cacops spp., and P. ferricolus to K. acervalis, A. petrolicus, and Z. lucius. In all these taxa, except for P. ferricolus, the tabular contacts the quadrate; this closure of the otic notch occurred only in late ontogeny in C. morrisi (Reisz et al., Reference Reisz, Schoch and Anderson2009).
Cacops spp. and P. ferricolus share additional apomorphic character states, such as the shorter prefrontal, the smaller external naris, and the enlarged LEP with its concave dorsal surface. These features are possibly shared with K. acervalis and others, which remains unclear due to missing data concerning sutures. However, P. ferricolus and K. acervalis appear to be advanced beyond the condition in Cacops spp. in the following character states: (1) smaller interpterygoid vacuities, resulting from the medial expansion of palatine and pterygoid (condition more extreme in K. acervalis), (2) the enlarged vomer with the posteriorly elongated, slit-like choana, and (3) the absence of an internarial fenestra. All eucacopines more derived than Conjunctio multidens share the tendency to form pronounced dermal sculpture on the skull roof, which usually obscures the sutures in adults.
The large eucacopines probably evolved into niches occupied by trematopids in other ecosystems. It is noteworthy that the former apparently did not co-exist with the latter, and, in at least some assemblages with both trematopids and eucacopines, the latter were substantially smaller, such as the Sakmarian-age fissure fillings at Fort Sill, Oklahoma (Polley and Reisz, Reference Polley and Reisz2011; Gee and Reisz, Reference Gee and Reisz2018).
Acknowledgments
We thank C. Mehling (AMNH) and the late F. Jenkins (MCZ) for access to specimens in their respective collections and M. Kamenz and I. Rosin (SMNS) for preparation of USNM 544065 and casting additional material. The software TNT is made freely available by the Willi Hennig Society. We thank N. Fröbisch and F. Witzmann for their constructive reviews.
Data availability statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.tb2rbp02d






