Introduction
Metoposaurids are a clade of large-bodied, non-marine Late Triassic (Carnian–Rhaetian) temnospondyls with a wide distribution across Laurasia and sporadic occurrences in Gondwana. In North America, three taxa are recognized: Anaschisma browni Branson, Reference Branson1905 (historically called “Buettneria perfecta” Case, Reference Case1922 or “Koskinonodon perfectus” recombined by Mueller, Reference Mueller2007, but see synonymy by Gee et al., Reference Gee, Parker and Marsh2020), Apachesaurus gregorii Hunt, Reference Hunt1993, and “Metoposaurus” bakeri Case, Reference Case1931. The vast majority of specimens have been collected from the southwestern United States, primarily from the Chinle Formation (and its equivalents) and from the Dockum Group (e.g., Colbert and Imbrie, Reference Colbert and Imbrie1956; Hunt, Reference Hunt1993; Long and Murry, Reference Long and Murry1995). A smaller, but equally important collection was also made in the Popo Agie Formation of Wyoming (Branson, Reference Branson1905; Branson and Mehl, Reference Branson and Mehl1929; Gee et al., Reference Gee, Parker and Marsh2020), with renewed collecting efforts in recent years (Kufner and Lovelace, Reference Kufner and Lovelace2018). Metoposaurids have never been reported from the mid-continent (longitudinally between Texas and the eastern seaboard), where terrestrial Late Triassic deposits are essentially unknown, and records from the eastern seaboard are both sparse and highly fragmentary (e.g., Leidy, Reference Leidy1856; Cope, Reference Cope1866, Reference Cope1868; von Huene, Reference von Huene1921; Sues and Olsen, Reference Sues and Olsen2015). However, the record from eastern North America is important for understanding patterns of metoposaurid biogeography and dispersal given the relative geographic proximity to other metoposaurid-bearing horizons in the Argana Basin of Morocco compared to other North American localities.
Baird (Reference Baird1986) summarized the eastern seaboard records, and his account remains accurate, in spite of its datedness, due to a general paucity of subsequent research on temnospondyls from the region. The most complete records come from the Cumnock Formation of North Carolina, the Wolfville Formation of Nova Scotia, and the New Oxford Formation of Pennsylvania. The earliest description of metoposaurids from Pennsylvania were Cope's descriptions (Reference Cope1866, Reference Cope1868), brief and without figures, of material from the Lockatong Formation in Chester County (Fig. 1). This material was originally described as Mastodonsaurus durus Cope, Reference Cope1866, but was subsequently placed in a new metoposaurid genus, Eupelor Cope, Reference Cope1868. A holotype was not defined in the original publication, and the diagnosis by Cope refers only to attributes of the marginal teeth. Baird (Reference Baird1986) noted that AMNH FARB 3927, a partial skull, is the type based on a handwritten note by Cope designating it as such, and this specimen was designated as the lectotype by Hunt, Reference Hunt1993. However, Baird concluded that the material was not diagnostic and could neither be distinguished from, nor synonymized with, another taxon (followed by Hunt, Reference Hunt1993, and all subsequent workers). The specimen is so incomplete as to create confusion over its skeletal identity (e.g., Colbert and Imbrie, Reference Colbert and Imbrie1956 alternatively identified it as a clavicle). We are inclined to side with Baird in his identification following personal examination by BMG, but neither sutures nor teeth could be confidently identified, and the specimen is not further addressed here. Eupelor has been treated as a nomen dubium by all other recent workers and is maintained as such here.

Figure 1. Geographic and temporal position of metoposaurid-bearing localities from the Upper Triassic of Pennsylvania. (1) Map of Pennsylvania highlighting the fossil localities where metoposaurid temnospondyls have been collected. Colors and patterns from the enlarged portion of the map highlight only the Triassic and Jurassic strata and come from the official geologic map of Pennsylvania from the Bureau of Topographic and Geologic Survey (Department of Conservation and Natural Resources, Commonwealth of Pennsylvania). (2) Upper Triassic–Lower Jurassic stratigraphy in Pennsylvania showing stratigraphic placement of the metoposaurid fossil localities. Circle = Chester County locality (type locality of Eupelor durus, AMNH FARB 3927); square = Emigsville area locality (includes AMNH FARB 1850 and 1863); star = Zions View locality.
The other records of metoposaurids in Pennsylvania come from the New Oxford Formation exposures in York County. Von Huene (Reference von Huene1921) described two specimens (AMNH FARB 1850 and 1863) from Cope's collection that were originally stated to have come from the type locality of “Eupelor durus,” but Baird (Reference Baird1986) amended the provenance to a copper mine near Emigsville (the Wheatley Copper Mine; Fig. 1). Additional material from this site is catalogued at the American Museum of Natural History (AMNH), but it is very fragmentary (Colbert and Imbrie, Reference Colbert and Imbrie1956, figs. 4, 5; BMG, personal observation). Baird (Reference Baird1986) suggested this other material was not diagnostic below the family level, with which we concur, and some of it may not even be properly referable below Temnospondyli on the basis of an absence of metoposaurid apomorphies (Table 1). The other source of Pennsylvanian metoposaurids is the Zions View locality, located ~5 km northwest of Emigsville (Fig. 1). Baird (Reference Baird1986, p. 128) noted two unprepared skulls that were collected in 1970 that he considered to be “definitely Metoposaurus,” which at the time included what is now recognized as Metoposaurus von Meyer, Reference von Meyer1842 sensu Brusatte et al., Reference Brusatte, Butler, Mateus and Steyer2015 (i.e., only European taxa) and the North American Anaschisma Branson, Reference Branson1905 (at the time, “Buettneria”). The material has been indirectly alluded to by previous workers. Doyle and Sues (Reference Doyle and Sues1995) reported Buettneria perfecta (=Anaschisma browni) in their description of the phytosaur Rutiodon Emmons, Reference Emmons1856 from the locality, which was followed by Sullivan et al. (Reference Sullivan, Lucas and Randall1995). Lucas and Sullivan (Reference Lucas and Sullivan1996) included photographs of a skull, mandible fragment, and interclavicle, but gave no description or interpretive line drawing. Similarly, the specimen is photographed in dorsal view by Schoch and Milner (Reference Schoch and Milner2000, pl. 7b), but without any details or comments. In both instances, the photographs are of little value for discerning the finer details of the cranial anatomy in a modern context and do not permit an assessment of the proposed taxonomic affinities. Most of the work on the locality has been in the context of the more substantial phytosaur material or broader biostratigraphic correlations.
Table 1. Specimen listing of metoposaurid material previously collected from Pennsylvania. All specimens were previously identified as Eupelor durus (although some were corrected to Metoposaurus sp. with a handwritten note). The Wheatley Copper Mine is located in York County; the Phoenixville Tunnel is located in Chester County.

Detailed study and informed taxonomy of the Zions View metoposaurids are important for several reasons. Firstly, much of the temnospondyl material from the eastern seaboard is fragmentary, having often been collected from active mining or tunneling operations in the late 19th to early 20th centuries, and is non-referable below the family or superfamily level. In contrast, the Zions View locality preserves at least one complete skull and other diagnostic material. Secondly, if previous identifications of the material are correct, Zions View would represent the only definitive record of Anaschisma browni from eastern North America and render the taxon as the most widely distributed metoposaurid. The eastern seaboard record represents an important geographic intermediary between the well-known southwest of North America and all other metoposaurid-bearing basins. The region would have been situated closer to the Argana Basin of Morocco than to the Chinle or Dockum basins of North America during the amalgamation of Pangea. This geographic range provides both additional data to inform on the biogeography and dispersal of metoposaurids, which is typically hindered by the restricted knowledge of many taxa to the sample from a single locality (e.g., Dutuitosaurus ouazzoui Dutuit, Reference Dutuit1976), and an avenue for examining whether intraspecific variation occurs across a geographic range. Lastly, substantial advancements have been made in metoposaurid paleobiology since the turn of the century. Detailed re-examination of historic specimens to assess cursory or tentative identifications by earlier workers are thus warranted to re-evaluate previous interpretations of alpha taxonomy and spatiotemporal trends within the clade (e.g., Buffa et al., Reference Buffa, Jalil and Steyer2019; Chakravorti and Sengupta, Reference Chakravorti and Sengupta2019; Gee et al., Reference Gee, Parker and Marsh2020).
Materials
The material (re-)described here consists of one complete skull (SMP VP-44), one partial hemimandible (SMP VP-37), one partial (SMP VP-514) and one complete (SMP VP-515) interclavicle, and two partial clavicles (SMP VP-360, SMP VP-516). All material described here comes from exposures of the middle-upper New Oxford Formation at Little Conewago Creek near Zions View, York County, Pennsylvania, USA (SMP locality 159; Fig. 1). Precise locality coordinates are available to qualified researchers upon request to the State Museum of Pennsylvania. The locality was described as the “phytosaur hole” by Baird (Reference Baird1986, p. 128), but is more typically referred to as Zions View, which is the type locality for the Conewagian Land Vertebrate Faunachron (LVF) (Huber et al., Reference Huber, Lucas and Hunt1993). It was discovered in 1909 and excavated for several years, abandoned until the 1970s, and then re-excavated for a short time (history summarized by Kochanov and Sullivan, Reference Kochanov and Sullivan1994); the more recent excavation produced the metoposaurid material described here.
Doyle and Sues (Reference Doyle and Sues1995) described substantial amounts of phytosaur material from this site, most of which was collected in the more recent excavation, and ascribed it to Rutiodon Emmons, Reference Emmons1856. Olsen et al. (Reference Olsen, McCune and Thomson1982) reported isolated skull bones and scales from the seminotid fish Synorichthys Schaeffer, Reference Schaeffer1967 and Turseodus Leidy, Reference Leidy1857. Wanner (Reference Wanner1926) described the conchostracan “Estheria” (=Wannerestheria Kozur and Weems, Reference Kozur and Weems2010) pennsylvanica from the site, and Lucas and Sullivan (Reference Lucas and Sullivan1996) noted unionid bivalves and abundant coprolites.
Repositories and institutional abbreviations
AMNH FARB, American Museum of Natural History (Fossil Amphibians, Reptiles and Birds collections), New York, New York, USA; FMNH UC, Field Museum of Natural History (University of Chicago collection), Chicago, Illinois, USA; ISI, Institute of India (Geology Museum), Kolkata, India; MNHN, Muséum national d'Histoire naturelle, Paris, France; SMP VP, State Museum of Pennsylvania (Vertebrate Paleontology collection), Harrisburg, Pennsylvania, USA; UCMP, University of California Museum of Paleontology, Berkeley, California, USA; UMMP VP, University of Michigan Museum of Paleontology (Vertebrate Paleontology collection), Ann Arbor, Michigan, USA; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Systematic paleontology
Order Temnospondyli von Zittel, Reference von Zittel1888
Suborder Stereospondyli von Zittel, Reference von Zittel1888
Superfamily Metoposauroidea Watson, Reference Watson1919
Family Metoposauridae Watson, Reference Watson1919
Anaschisma Branson, Reference Branson1905
Type species
Anaschisma browni Branson, Reference Branson1905 from the Popo Agie Formation of Wyoming, by original designation.
Anaschisma browni Branson, Reference Branson1905
Figures 2–8
- Reference Branson1905
Anaschisma browni Branson, p. 585, figs. 7, 8, 10.2.
- Reference Branson and Mehl1929
Anaschisma browni; Branson and Mehl, p. 199, fig. 6.
- Reference Colbert and Imbrie1956
Eupelor browni; Colbert and Imbrie, p. 422, pl. 26.3, 26.4.
- Reference Chowdhury1965
Metoposaurus browni; Chowdhury, p. 42.
- Reference Gregory1980
Anaschisma browni; Gregory, p. 129.
- Reference Hunt1993
Metoposauridae indet. Hunt, p. 86.
- Reference Schoch and Milner2000
Metoposauridae indet. Schoch and Milner, p. 125.
- Reference Gee, Parker and Marsh2020
Anaschisma browni; Gee et al., p. 236, figs. 2–6.

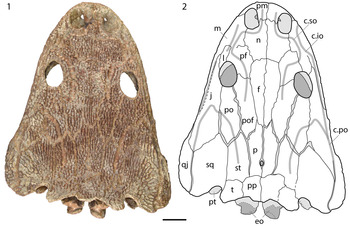
Figure 2. Dorsal view of the referred skull of Anaschisma browni, SMP VP-44. (1) photograph; (2) line drawing of the same. Abbreviations: c.io, infraorbital canal; c.po, postorbital canal; c.so, supraorbital canal; eo, exoccipital; f, frontal; j, jugal; l, lacrimal; m, maxilla; n, nasal; p, parietal; pf, prefrontal; pm, premaxilla; po, postorbital; pof, postfrontal; pp, postparietal; pt, pterygoid; qj, quadratojugal; sq, squamosal; st, supratemporal; t, tabular. Scale bar = 5 cm.

Figure 3. Comparison of the circumorbital elements of the referred skull of Anaschisma browni, SMP VP-44, with other metoposaurid specimens. (1–3) Left circumorbital region of SMP VP-44 in different angled lighting; (4) line drawing of the same; (5) line drawing of the holotype of Metoposaurus krasiejowensis (ZPAL AbIII/358; modified from Sulej, Reference Sulej2007, fig. 13); (6) line drawing of the holotype of Anaschisma browni (FMNH UC 447; modified from Gee et al., Reference Gee, Parker and Marsh2020, fig. S1; right side reflected here); (7) line drawing of the holotype of “Buettneria perfecta” (UMMP VP 7475; modified from Case, Reference Case1922, fig. 1A); (8) line drawing of the holotype of “Metoposaurus” bakeri (UMMP VP 13055; modified from Case, Reference Case1932, fig. 2); (9) line drawing of the holotype of Apachesaurus gregorii (UCMP 63845; modified from Spielmann and Lucas, Reference Spielmann and Lucas2012, fig. 10A; right side reflected here); (10) line drawing of the holotype of Arganasaurus azerouali (MNHN.F.ARG 5; modified from Buffa et al., Reference Buffa, Jalil and Steyer2019, fig. 1B); (11) line drawing of the holotype of Panthasaurus maleriensis (ISI A 56; modified from Chakravorti and Sengupta, Reference Chakravorti and Sengupta2019, fig. 3b). Abbreviations: j, jugal; l, lacrimal; m, maxilla; n, nasal; pf, prefrontal. Scale bar in (4) = 1 cm; line drawings (5–11) are not scaled relative to each other or to 3.4.

Figure 4. Occipital view of the referred skull of Anaschisma browni, SMP VP-44. (1) Photograph in posterior profile; (2) line drawing of the same. Abbreviations: eo, exoccipital; f.m, foramen magnum; f.p, paraquadrate foramen; pp, postparietal; psp, parasphenoid; pt, pterygoid; pt.oc, oblique crest of the pterygoid; q, quadrate; qj, quadratojugal; sq, squamosal; t, tabular. Scale bar = 3 cm.

Figure 5. Ventral (occlusal) view of the referred skull of Anaschisma browni, SMP VP-44. (1) Photograph; (2) line drawing of the same. Palatal dentition other than the enlarged “fangs” is not figured. Abbreviations: cp, cultriform process; ect, ectopterygoid; eo, exoccipital; ipv, interpterygoid vacuity; j, jugal; m, maxilla; pal, palatine; pm, premaxilla; psp, parasphenoid; pt, pterygoid; q, quadrate; qj, quadratojugal; stf, subtemporal fenestra; t.p, enlarged palatine teeth; t.v, enlarged vomerine teeth; v, vomer; v.r, vomerine ridge. Scale bar = 5 cm.

Figure 6. Close-up photograph of the ornamentation on the parasphenoid of the referred skull of Anaschisma browni, SMP VP-44 in ventral (occlusal) view. Scale bar = 5 cm.

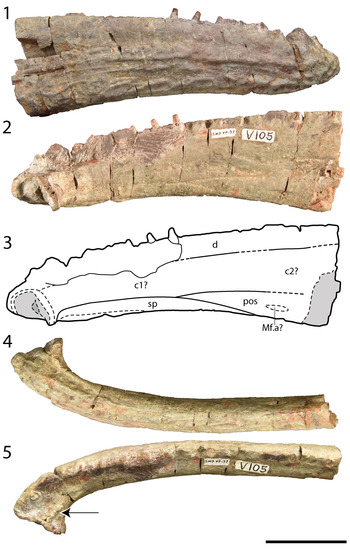
Figure 7. Photographs of the referred hemimandible of Anaschisma browni, SMP VP-37. (1) Photograph in labial profile; (2) photograph in lingual profile; (3) line drawing of the lingual profile; (4) photograph in ventral profile; (5) photograph in dorsal profile; arrow indicates the row of adsymphyseal teeth. Abbreviations: c1?, ?anterior (or ?first) coronoid; c2?, ?second coronoid; d, dentary, Mf.a?, anterior Meckelian fenestra?; pos, postsplenial; sp, splenial. Scale bar = 5 cm.

Figure 8. Photographs of the referred postcrania. (1, 2) SMP VP-515, complete interclavicle in (1) ventral profile and (2) dorsal profile; (3, 4) SMP VP-514, partial interclavicle in (3) ventral profile and (4) dorsal profile; (5) SMP VP-516, left clavicle in ventral profile; (6) SMP VP-360, partial right clavicle in ventral profile. All specimens are scaled relative to each other; scale bar = 5 cm.
Holotype
Complete skull (FMNH UC 447) from the Popo Agie Formation of Wyoming (Branson, Reference Branson1905).
Occurrence
The age of Zions View is not precisely constrained due to a lack of radioisotopic dating, challenges of correlating the Gettysburg Basin with other Late Triassic basins on the eastern seaboard, and the vagueness of what exactly constitutes the “middle” New Oxford Formation. The upper Gettysburg Basin had traditionally been identified as late Carnian (e.g., Olsen et al., Reference Olsen, McCune and Thomson1982; Doyle and Sues, Reference Doyle and Sues1995) prior to Muttoni et al.'s (Reference Muttoni, Kent, Olsen, Stefano, Lowrie, Bernasconi and Hernández2004) revision of the Carnian-Norian boundary that now situates it within the early Norian (see Fig. 1).
The New Oxford Formation has long been correlated with the lower part of the Lockatong Formation of the Newark Basin on the basis of palynological data (e.g., Cornet, Reference Cornet1977), and these collectively form the New Oxford-Lockatong palynofloral zone. This zone's lowest boundary is in the uppermost portions of the Stockton Formation (e.g., Olsen et al., Reference Olsen, Kent, Cornet, Witte and Schlische1996). Comparison of the New Oxford-Lockatong palynofloral zone with the Chinle Formation exposures from Petrified Forest National Park indicate a correlation with palynological zone II of the Chinle Formation (e.g., Lindström et al., Reference Lindström, Irmis, Whiteside, Smith, Nesbitt and Turner2016; Baranyi et al., Reference Baranyi, Reichgelt, Olsen, Parker and Kürschner2018). This zone extends from the Blue Mesa Member into the lower part of the Sonsela Member (e.g., Reichgelt et al., Reference Reichgelt, Parker, Martz, Conran, van Konijnenburg-van Cittert and Kürschner2013). This correlation would place Zions View as part of the Adamanian (Conewagian) Estimated Holochronozone (Martz and Parker, Reference Martz, Parker, Ziegler and Parker2017). This portion of the Chinle Formation and Dockum Group has long produced extensive remains of Anaschisma browni in Arizona, Texas, and New Mexico (e.g., Hunt and Lucas, Reference Hunt and Lucas1993; Long and Murry, Reference Long and Murry1995; Parker and Irmis, Reference Parker and Irmis2005; Parker and Martz, Reference Parker and Martz2010). Conversely, Stocker et al. (Reference Stocker, Sues and Jasinski2017), after reviewing the record of Rutiodon from the locality and determining that it represented Parasuchus Lydekker, Reference Lydekker1885, suggested that this documentation implied either a Carnian age of the tetrapod-bearing horizons of the New Oxford Formation or a greater temporal extent of Parasuchus. Both Parasuchus and A. browni are also known from Carnian-age sediments in Wyoming (Branson, Reference Branson1905; Branson and Mehl, Reference Branson and Mehl1929; Lucas et al., Reference Lucas, Heckert and Rinehart2007; Gee et al., Reference Gee, Parker and Marsh2020). Lastly, the conchostracan Wannerestheria pennsylvanica from Zions View is found in the upper units of the Lockatong Formation (Kozur and Weems, Reference Kozur and Weems2010). Kozur and Weems (Reference Kozur and Weems2010, p. 343–344) summarized the history of stratigraphic positioning of the locality (described as strata along Little Conewago Creek in that study) and favored a placement of the site in the lowest part of the Gettysburg Formation (Norian), which overlies the New Oxford Formation. There is an obvious need for further work to resolve both the stratigraphic position of the site and its relation to the broader regional framework. Here we consider the site to be Norian (in agreement with Kent et al., Reference Kent, Olsen and Muttoni2017), although it is not of great importance in the context of this study because A. browni is already known from both Carnian and Norian deposits, so the confirmation of its occurrence in this study does not contribute to this conundrum.
Description
Only one skull, SMP VP-44, is present in collections. Numerous unprepared specimens are reposited at the SMP, and the second skull noted by Baird (Reference Baird1986) and Schoch and Milner (Reference Schoch and Milner2000), without specified catalogue number, may be among them (Baird, Reference Baird1986 specifically noted in his report that both skulls were unprepared). SMP VP-44 is complete and only slightly transversely distorted (Fig. 2), such that the right side of the skull is slightly posteriorly shifted relative to the equivalent left side. The skull measures 42 cm in midline length from the premaxilla to the postparietals. The orbits are anteroposteriorly long ovals, and the nares are only slightly smaller and more circular. Towards the posterior skull table, some of the elements have slightly separated such that the definition of the elements is exaggerated.
The premaxilla is a subtriangular element framing the external naris (Fig. 2). The suture with the nasal is a sinuous, transversely oriented contact, except for adjacent to the medial margin where a short alary process extends posteriorly. The nasal is a broad quadrangular element that expands laterally to frame the naris from behind before tapering along the contact with the prefrontal to meet the frontal in a mostly horizontal transverse suture (Fig. 2). The maxilla is a long, slender element with most of the marginal dentition (Fig. 2). Its dorsal exposure is narrow except for posterior to the naris where it expands to meet the nasal, widely separating the lacrimal from the naris.
The circumorbital elements warrant more detailed description because the position of the lacrimal remains useful for differentiating the North American taxa (e.g., Hunt, Reference Hunt1993; Spielmann and Lucas, Reference Spielmann and Lucas2012; Gee et al., Reference Gee, Parker and Marsh2020). The lacrimal enters the orbit in Anaschisma browni and is excluded from the orbit in Apachesaurus gregorii and “Metoposaurus” bakeri (Case, Reference Case1922, Reference Case1931, Reference Case1932; Spielmann and Lucas, Reference Spielmann and Lucas2012; Lucas et al., Reference Lucas, Rinehart, Heckert, Hunt and Spielmann2016; Fig. 3). Although the sutures are not as clear in SMVP VP-44 as in some other North American specimens, there is clearly a sutural contact on the left lateral orbital margin just anterior to the mid-length (Fig. 3.1–3.3). From the orbital margin, the suture extends more or less straight anteriorly along a long crack. This mirrors the condition of An. browni (Fig. 3.6, 3.7); when the lacrimal is excluded, a short jugal-prefrontal suture extends anterolaterally from the orbit and then bifurcates, laterally for the jugal-lacrimal suture and medially for the lacrimal-prefrontal suture. There is no evidence for this bifurcation, and we are thus confident that the crack occurred along the jugal-lacrimal suture. Another line of evidence is in the point of origination of the presumed lacrimal-jugal suture at the lateral orbital margin. In most specimens of either Ap. gregorii or “M.” bakeri, the prefrontal does not expand that far laterally (Case, Reference Case1932; Spielmann and Lucas, Reference Spielmann and Lucas2012; Fig. 3.8, 3.9); there is also a slight medial expansion of the jugal to complete the exclusion of the lacrimal. The lacrimal is not as well defined on the right side of the skull, but the jugal and prefrontal are definitively not in contact. On both sides, the jugal extends anterior to the orbit and partially overlaps the lacrimal medially. Posteriorly, it expands in the temporal region to wedge between the quadratojugal and the squamosal.
The prefrontal is a square element that sutures to the postfrontal to omit the frontal from the orbital margin (Fig. 2). The suture is angled anterolaterally and contacts the orbit slightly anterior to the mid-length of the orbit on the right side and at the mid-length on the left side. The postfrontal is an elongate rectangle that tapers posteriorly to wedge between the parietal and supratemporal (Fig. 2). The frontal is a slender sub-triangular element that tapers gradually posteriorly (Fig. 2). The postorbital is an elongate pentagonal element that expands slightly posteriorly to wedge between the supratemporal and squamosal (Fig. 2).
The parietal is a slender pentagonal element that expands slightly anteriorly (Fig. 2). A narrow anterior process partially wedges between the postfrontal and the frontal. The pineal foramen is situated in the posterior third of the parietals. The postparietal is a rectangular element, slightly longer than it is wide (Fig. 2). The tabular is a slightly smaller element of more equant proportions (Fig. 2). On both sides, the occipital margin is damaged (normally it formed a smooth posteriorly concave surface). The tabular horns are also slightly weathered, although they are clearly defined and contribute to the medial margin of the otic notch. The supratemporal is a pentagonal element of equal width throughout, with an anterior process wedging between the postfrontal and postorbital (Fig. 2). The squamosal is a much broader pentagonal element, incised posteriorly for the well-developed otic notch (Fig. 2). Its anterior process is medially offset from the element's midline and wedges between the postorbital and jugal. The quadratojugal is a curved element with a subtriangular dorsal profile (Fig. 2). A slender anterior process extends along the jugal's posterolateral margin. The quadratojugal also curves ventrolaterally.
Ornamentation on the skull is typical of most large-bodied metoposaurids, consisting primarily of circular pitting on the snout and interorbital region (Fig. 2). Longer radiating grooves are common on the posterior and temporal elements. The lateral line canals are well defined throughout. The supraorbital canal originates anteriorly on the premaxilla, curving around the medial narial margin and toward the lacrimal, then curving back toward the midline in front of the orbit and extending posteriorly onto the postfrontal. The postorbital canal is generally “U”-shaped, with a small kink as it crosses the jugal-postorbital suture. One posterior termination is at the posterior skull margin at the suture between the squamosal and the quadratojugal. It extends anteriorly onto the jugal and then curves across onto the postorbital and back onto the supratemporal, on which it terminates. The infraorbital canal originates on the medial expansion of the maxilla and extends posteriorly along the lateral margin of the skull to meet the postorbital canal on the right side. The canal is not completely preserved anteriorly on the left side and curves onto the lateral surface of the skull posteriorly such that it does not connect to the postorbital canal.
The occiput is complete but distorted, with some dorsoventral compression having dislodged the exoccipitals posteriorly such that they project farther than they would have when the animal was alive (Figs. 2, 4). Consequently, the foramen magnum is not defined dorsally and dorsolaterally and appears generally more triangular than “keyhole-shaped” as in most temnospondyls without ossification of the synotic tectum. There are well-defined lamellose processes that project medially from the column of the exoccipital; these normally divide the foramen magnum proper from the unossified synotic tectum to produce the keyhole-shaped opening. The artificial condition produced by compression here is not equivalent to the superficially similar biological condition found in Apachesaurus gregorii (Spielmann and Lucas, Reference Spielmann and Lucas2012). This compression has also distorted the paraquadrate foramina in the temporal region, and, as in many metoposaurid specimens, it is not possible to discern whether any accessory paraquadrate foramina were present at the posterolateral end of the larger foramen. The sutures of the roofing elements are essentially indiscernible in occipital view, although it may be inferred that the point at which the occiput fractured and was partially extended posteriorly was in the region where the postparietal and the tabular would meet the exoccipital to jointly frame the posttemporal foramen (not defined in this specimen). The oblique crest of the pterygoid (sometimes termed the ascending lamina) is also well exposed and projects dorsally and slightly posteriorly beneath the otic notch (see Fig. 2).
The palate is complete, but sutures are generally difficult to identify confidently, and the total number of tooth positions cannot be discerned (Fig. 5). The midline sutures appear to have been weathered, while other sutures are more tentatively identified on the basis of prominent fractures in the approximate region in which sutures would be expected (e.g., the parasphenoid-pterygoid, the vomer-palatine, and the ectopterygoid-palatine contacts) or where only portions of the sutures are identifiable. The small palatal exposure of the jugal can be predicted, but the suture with the maxilla and the ectopterygoid is not discernible. There are practically no features of the palate that differ among metoposaurids, and those that do relate to qualitative aspects, such as the degree of parasphenoidal ornamentation (Fig. 6), rather than to differences in the morphology and sutures. The cultriform process is broad and flat, like all metoposaurids other than Apachesaurus gregorii. The typical palatal tooth arrangement is identified, with paired “fangs” or “tusks” on the vomer, anterior to the choana, and on the palatine, posterior to the choana. Teeth of the same size as those of the premaxilla and maxilla occur from the palatine “fangs” to the end of the tooth row. Small teeth occur as a transvomerine row on a slightly elevated (ventrally offset) ridge and extend posterolaterally along the medial margin of the choana to the palatine “fangs” (Fig. 5). These are more clearly defined on the left vomer and palatine.
SMP VP-37 is a partial right hemimandible, preserved anteriorly from the symphyseal region and containing less than half of the tooth row (Fig. 7). Over its preserved length, it measures 20.4 cm. Based on the partially identified sutures, the fragment is probably ~40–45% complete, and not more than 50% of the total length, and thus would belong to a similarly sized individual to SMP VP-44. No collections records indicate a clear association with SMP VP-44, and metoposaurid-bearing assemblages are typically characterized by many individuals of similar sizes, so size alone is not a reliable means for determining association. The dentary, splenial, postsplenial, and anterior (first) coronoid are preserved (Fig. 7.2, 7.3). A second coronoid can be inferred based on the conserved mandibular configuration of metoposaurids (e.g., Sulej, Reference Sulej2007; Lucas et al., Reference Lucas, Rinehart, Heckert, Hunt and Spielmann2016), but the suture with the first coronoid is not identified. The symphysis is marked by an unossified, medially facing concave surface. The splenial makes a very small contribution to the ventromedial surface of the symphysis. A small depression toward the posterior portion of the preserved postsplenial is probably the anterior Meckelian window. Ornamentation on the labial surface comprises a mixture of oval pits and elongate grooves bounded by low ridges. The pits are more abundant toward the ventral margin and symphysis, whereas the grooves and ridges are more dominant dorsally. A shallow longitudinal groove (the superior mandibular groove or the oral groove) divides the ornamented ventrolabial surface from the smooth dorsolabial surface. Teeth are mostly absent, but the tooth sockets are sufficiently defined to estimate a tooth count along the preserved region of 27–29 positions in addition to the paired symphyseal “fangs.” The tooth count agrees with estimation of the hemimandible's complete size based on the general dentary tooth count of metoposaurids, which is in the range of 70–75 teeth (e.g., Sulej, Reference Sulej2007). There is also a row of poorly defined, small teeth, marked by an elevated ridge that is posterior (lingually situated) to the symphyseal “fangs”; the count is unclear given their quality of preservation. As preserved, the teeth are circular in cross-section and tapered to form conical structures; carinae are not apparent.
SMP VP-515 is a large, essentially complete interclavicle (Fig. 8.1, 8.2). The element is broadly diamond-shaped, with an anteriorly directed process. The ornamentation on the ventral surface comprises the typical central region with circular pits that radiate outwards in all directions to form elongate grooves. The region of circular pitting is proportionately large, as in the large North American metoposaurids (e.g., Case, Reference Case1922, Reference Case1931) and in contrast to the European metoposaurids (Sulej, Reference Sulej2002, Reference Sulej2007). Broad unornamented surfaces for the articulation of the clavicles are found on each side and meet along the midline anterior to the anterior extent of the ornamentation.
SMP VP-514 is a partial interclavicle with the right lateral margin and some of the central plate preserved (Fig. 8.3, 8.4). The posterior end of the unornamented shelf for the clavicular articulation is preserved. Neither the anterior nor posterior extent of the entire element is defined, nor is the center of ossification. The ornamented region is partially preserved and consists almost entirely of elongate grooves. Over the overlapping region with SMP VP-515, there are no observable differences.
SMP VP-516 is a partial left clavicle still partially embedded in a large block of matrix and with only the ventral surface exposed (Fig. 8.5). The dorsal process appears to have been completely lost. Some damage has occurred at the posterolateral corner of the element, but it preserves a large area of circular pitting that radiates medially and anteriorly in the form of more elongate grooves. The lateral margin is nearly straight.
SMP VP-360 is a partial right clavicle lacking the medial and anterior portions (Fig. 8.6). The specimen is currently mounted at the Pennsylvania State Capitol building and can only be presented here in ventral profile, but the dorsal region is not preserved in any event. In its present condition, it is much longer than wide and is relatively flat; the margins are mostly damaged such that the contour is only vaguely discernible. Ornamentation consists of circular pitting toward the posterolateral edge that radiates into elongate grooves anteriorly. In contrast to the nearly straight margin in SMP VP-516, the margin is slightly convex in SMP VP-360, although this is not an appreciable difference.
Remarks
The revised osteology and diagnosis of Anaschisma browni have been recently presented by Gee et al. (Reference Gee, Parker and Marsh2020) and a forthcoming publication by Kufner and Gee; therefore, no new information on the diagnosis is presented here.
Discussion
Taxonomic identity
Various apomorphies or differential features identified by previous authors, and most recently by Gee et al. (Reference Gee, Parker and Marsh2020), for Anaschisma browni (e.g., alary process of the premaxilla, anteroposteriorly short lacrimal with broad contribution to the orbit) permit confident referral of SMP VP-44 to this taxon. The latter feature is particularly important in separating this material from the only other diagnostic metoposaurid material on the eastern seaboard—the small, referred specimen of “Metoposaurus” bakeri from Nova Scotia (first referred by Baird, Reference Baird1986, and accepted by all other authors; see Sues and Olsen, Reference Sues and Olsen2015, fig. 12, for photograph), which lacks this contact, like its conspecifics from Texas. While identification of the Zions View material is in agreement with previous authors’ interpretations dating back to at least the 1990s, it bears noting that the specimen has never been properly described, with justification for taxonomic referral, either implicitly by way of interpretive figures or explicitly by documentation of apomorphic features or unique combinations of characters. Such justification is essential for ensuring rigorous propagation of data that can be independently assessed by other workers in some fashion. Large temnospondyl material from the east coast of North America should be scrutinized more carefully for several reasons: (1) these localities were spatially closer to other metoposaurid-bearing regions in Europe and Morocco at that time (not coeval at present, but possible spatiotemporal range extensions); (2) these localities are less well-constrained stratigraphically and temporally; and (3) the east coast is the only region in North America where a large-bodied, non-metoposaurid taxon (Calamops paludosus) occurs in the Late Triassic (Sues and Schoch, Reference Sues and Schoch2013).
The remainder of the material can only be referred based on circumstantial evidence and by the absence of any features that clearly differ from either Metoposauridae (for which most diagnostic features are cranial; e.g., Buffa et al., Reference Buffa, Jalil and Steyer2019) or Anaschisma browni specifically. For example, there are no diagnostic features of the clavicle per the diagnosis of Buffa et al. (Reference Buffa, Jalil and Steyer2019); stereospondyls generally have similar interclavicular morphology and ornamentation (e.g., Warren and Snell, Reference Warren and Snell1991). SMP VP-515 is nearly complete, so the subtler nuances in contour allow for differentiation from even the closely related Middle Triassic metoposauroid Callistomordax kugleri Schoch, Reference Schoch2008, which has a narrower, more rhomboidal interclavicle with prominent equant anterior and posterior styluses. SMP VP-514 is much more fragmentary, but it at least differs in no overlapping regions from SMP VP-515. Beyond this, metoposaurid interclavicles are essentially differentiated only by the size of the circular pitted region on the ventral surface by a qualitative binary metric: small or large. Anaschisma browni and “Metoposaurus” bakeri typically have been characterized by relatively large circular pitted regions (e.g., Case, Reference Case1922, Reference Case1931, Reference Case1932; Sulej, Reference Sulej2002, Reference Sulej2007; though see Lucas et al., Reference Lucas, Rinehart, Heckert, Hunt and Spielmann2016, for a contrary example from Rotten Hill), like in SMP VP-515, although the intraspecific variability remains to be better assessed (e.g., Spielmann and Lucas, Reference Spielmann and Lucas2012). From a differential sense, the interclavicles can thus only be differentiated from some taxa and compared favorably with others. Stereospondyl clavicles are arguably even more highly conserved than interclavicles, and the only diagnostic features of Metoposauridae (following Buffa et al., Reference Buffa, Jalil and Steyer2019) relate to the clavicular blade and the articulation with the interclavicle. The latter cannot be assessed here, and the former refers to an intraspecifically variable indentation in the medial edge (Sulej, Reference Sulej2007) that also occurs in brachyopoids (Warren and Marsicano, Reference Warren and Marsicano2000), which is not completely preserved in either SMP VP-360 or SMP VP-516. Even if a metoposaurid identity is accepted, isolated clavicles usually cannot be referred to a particular taxon. Size of circular ornamentation has sometimes been used to differentiate taxa (e.g., Hunt, Reference Hunt1993; Sulej, Reference Sulej2002). One feature noted by Gee et al. (Reference Gee, Parker and Marsh2020) is the presence or absence of a sensory groove on the clavicle (absent in A. browni, but present in Panthasaurus maleriensis Chowdhury, Reference Chowdhury1965). The absence of this feature on SMP VP-360 and SMP VP-516, therefore, at least partially excludes some taxa from consideration.
The hemimandible (SMP VP-37) warrants particular attention because this is the only region of the skeleton that is known from the trematosauroid Calamops paludosus from the lower Stockton Formation in Pennsylvania (Sues and Schoch, Reference Sues and Schoch2013). Of the four diagnostic features listed by Sues and Schoch (Reference Sues and Schoch2013), the only one that can be assessed in SMP VP-37 is the presence of rows of deep pits on the symphysis. Although there are pits on the symphysis of SMP VP-37, this is a common feature of metoposaurids as a continuation of the ventrolabial ornamentation pattern, and these pits are neither deep nor arranged in rows. Other differential features of C. paludosus include the absence of adsymphyseal teeth (found in most metoposaurids; Konietzko-Meier and Wawro, Reference Konietzko-Meier and Wawro2007; Sulej, Reference Sulej2007) and the absence of a well-ossified surface medial to the symphyseal “fang.” SMP VP-37 differs in no regards from other metoposaurids, which have a highly conserved mandibular morphology (see Brusatte et al., Reference Brusatte, Butler, Mateus and Steyer2015, for some possible differentiating features).
Based on the conventional resemblance-based method of identification, there is little reason to suspect that all temnospondyl material from Zions View belongs to anything other than a metoposaurid. More specifically, based on the taxonomic affinities of the skull, probably all of the material belongs to Anaschisma browni. No attributes of any given specimen are inconsistent with this interpretation. Our referrals are thus based primarily on four lines of circumstantial evidence: (1) the near total absence of large-bodied non-metoposaurids in the Late Triassic of North America, (2) the tendency for metoposaurid-bearing deposits to be monotaxic with respect to temnospondyls, (3) the apomorphy-based identification of SMP VP-44 (skull), and (4) our ability to differentiate SMP VP-37 from Calamops paludosus. These lines of evidence have historically been regarded as relatively robust, or at least to be the most viable given the nature of the fossil record.
Lastly, we note that while our detailed study agrees with previously published interpretations regarding the taxonomy of the Zions View metoposaurid material, this study is the first to provide any justification or corresponding evidence for this interpretation. There are inherent limitations to studies that report occurrences without providing the relevant data to support the conclusions. Furthermore, the recognition of C. paludosus as a non-metoposaurid trematosauroid (Sues and Schoch, Reference Sues and Schoch2013) indicates the presence of at least one other large-bodied stereospondyl at a local geographic scale and underscores the need for explicit rationale for taxonomic assignment of material that does not rely largely on absolute size alone. The relatively poor record of temnospondyls on the eastern seaboard only further underscores the point that metoposaurid affinities cannot be assumed for all large specimens. Detailed re-study and re-description of historic specimens is important both for the sake of reproducibility (because taxonomic identifications are hypotheses rather than permanent, static facts) and for facilitating future studies by other workers who may be unable to personally examine the material but who may be interested in incorporating either qualitative (e.g., comparative anatomy) or quantitative (e.g., morphometrics) data from these specimens.
Ontogenetic maturity
Ontogenetic assessment of metoposaurid specimens remains of interest given the debate over Apachesaurus gregorii, which is represented only by relatively small individuals. Based strictly on size alone, SMP VP-44 was at least a sub-adult individual (midline length of 42 cm); metoposaurid skulls rarely exceed 55 cm in length (e.g., see measurements provided by Colbert and Imbrie, Reference Colbert and Imbrie1956; Dutuit, Reference Dutuit1976; Sulej, Reference Sulej2007; Lucas et al., Reference Lucas, Rinehart, Heckert, Hunt and Spielmann2016). Most patterns of metoposaurid ontogeny hinge on the interpretation of A. gregorii because it comprises most of the very small (<20 cm skull length) metoposaurid specimens (Gee and Parker, Reference Gee and Parker2020, table 1). Furthermore, because metoposaurids were paedomorphic, with underdeveloped neurocrania and postcrania, qualitative attributes used for non-paedomorphic temnospondyls (e.g., degree of ossification) are not useful here. These issues have been previously addressed and are not repeated here. The observation that the sutures of the posterior skull have separated slightly does suggest some immaturity of SMP VP-44. Beyond this, no other feature confers robust information one way or the other.
Intraspecific variation within Anaschisma browni.—Metoposaurids are among the most abundantly known Mesozoic temnospondyls, with dozens of known complete skulls among thousands of other fragmentary specimens. Such a sample size is relatively rare for extinct tetrapods and makes metoposaurids an excellent case study for studying intraspecific variation. There are several reasons why the documentation of intraspecific variation is critical for paleontological research at large. Firstly, increased sample size and hypothesized intraspecific variation permits continuous assessment of qualitative taxonomic diagnoses and differentiation—there are numerous examples across the tree of life of taxa named for specimens whose morphological differentiators are now recognized as the result of intraspecific variation (whether due to ontogeny or to other factors). This consideration is particularly salient for metoposaurids because the standard of morphological differentiation that warranted taxonomic distinction within the clade was historically quite low (e.g., Branson and Mehl, Reference Branson and Mehl1929). As such, these narrow typological standards led to an extensive number of taxa erected on minor or single differences that were often questionably valid or that ignored non-taxonomic explanators (e.g., taphonomic distortion, ontogenetic transformation). For example, features such as “eyes not as far forward” (Branson, Reference Branson1905, p. 588–589) for “Anaschisma brachygnatha” (=Anaschisma browni), “clavicle of striking characteristics” (Branson and Mehl, Reference Branson and Mehl1929, p. 233) for “Buettneria major” (=A. browni), and “small size” (Schoch and Milner, Reference Schoch and Milner2000, p. 121–122) for both Apachesaurus gregorii and Arganasaurus lyazidi Dutuit, Reference Dutuit1976 have been previously utilized to diagnose taxa. In the same vein, modern taxonomy, informed by phylogenetics, must also account for variation in scoring of phylogenetic matrices. In the context of the Metoposauridae, this consideration is important given the markedly different topological results of three recent studies (Buffa et al., Reference Buffa, Jalil and Steyer2019; Chakravorti and Sengupta, Reference Chakravorti and Sengupta2019; Gee et al., Reference Gee, Parker and Marsh2020). Some of the disparity results from differing characters (especially the use of predominantly quantitative characters by Chakravorti and Sengupta, Reference Chakravorti and Sengupta2019), but differing scoring (i.e., whether or not polymorphisms were scored when noted in the literature) should also be considered. Not scoring polymorphisms when they are known may contribute to artificially well-resolved topologies because characters appear more discretely informative than they truly are.
With respect to intraspecific variation, the most studied metoposaurid is Metoposaurus krasiejowensis Sulej, Reference Sulej2007. Sulej (Reference Sulej2007) identified several examples of intraspecifically variable cranial sutures, such as the relationship of the lacrimal to the orbit (a longstanding phylogenetic character), contact between the postorbital and the parietal, and contact between the prefrontal and the maxilla. In all three cases, the condition is documented to vary at the level of the individual. Furthermore, Teschner et al. (Reference Teschner, Sander and Konietzko-Meier2018) examined a broad range of humeri and identified distinct histotypes, while Antczak and Bodzioch (Reference Antczak and Bodzioch2018) identified variation in ornamentation patterns on the skull and pectoral girdle. Such variation would be further expanded if Sulej's (Reference Sulej2002) original subspecies designation were restored (i.e., Metoposaurus diagnosticus krasiejowensis vs. M. d. diagnosticus von Meyer, Reference von Meyer1842), or if his species-level framework was utilized without his subspecies addition (i.e., Metoposaurus krasiejowensis = M. diagnosticus).
In contrast, the published osteology of most metoposaurids is based exclusively or primarily from single localities, which in some instances might represent a single event that records a single population. Spatial restriction is true for Metoposaurus krasiejowensis and M. algarvensis Brusatte et al., Reference Brusatte, Butler, Mateus and Steyer2015 in Europe (Sulej, Reference Sulej2007; Brusatte et al., Reference Brusatte, Butler, Mateus and Steyer2015), all three of the Moroccan taxa (Arganasaurus azerouali Dutuit, Reference Dutuit1976, A. lyazidi, Dutuitosaurus ouazzoui; Khaldoune et al., Reference Khaldoune, Jalil, Germain and Steyer2016; Buffa et al., Reference Buffa, Jalil and Steyer2019), and the Indian taxon (Panthasaurus maleriensis; Chakravorti and Sengupta, Reference Chakravorti and Sengupta2019). The North American Apachesaurus gregorii has been described from many localities (Long and Murry, Reference Long and Murry1995), but most of this material can only be said to belong to small-bodied individuals and is not referable under an apomorphy-based system, notwithstanding doubts about the validity of the original interpretation of the taxon as a metoposaurid with a distinctly smaller adult body size (Gee and Parker, Reference Gee and Parker2020). Elimination of the single diagnostic postcranial feature, elongate intercentra (following Gee et al., Reference Gee, Parker and Marsh2017; Gee and Parker, Reference Gee and Parker2020), substantially reduces the number of properly referred specimens of Ap. gregorii. Thus, only Metoposaurus diagnosticus, “Metoposaurus” bakeri, and Anaschisma browni are confidently known from several localities that span an appreciable geographic range. It is worth noting that indeterminate material from localities in Algeria, France, Italy, and Madagascar (in addition to already known metoposaurid-bearing localities or regions) could hypothetically belong to known taxa and thus extend geographic ranges if restudied (Koken, Reference Koken1913; Corroy, Reference Corroy1928; Leonardi, Reference Leonardi1952; Lehman, Reference Lehman1971; Dutuit, Reference Dutuit1978; Cuny and Ramboer, Reference Cuny and Ramboer1991; Fortuny et al., Reference Fortuny, Arbez, Mujal and Steyer2019). Metoposaurus diagnosticus is confined to Germany (Schoch and Milner, Reference Schoch and Milner2000; Sulej, Reference Sulej2002, Reference Sulej2007; Milner and Schoch, Reference Milner and Schoch2004), while “M.” bakeri is known almost exclusively from Texas, with only the additional single specimen in Nova Scotia (Baird, Reference Baird1986; Sues and Olsen, Reference Sues and Olsen2015). Anaschisma browni is definitively recorded from at least five states (Arizona, New Mexico, Pennsylvania, Texas, Wyoming) and numerous localities within each state that collectively form a sizeable range within North America. Documented intraspecific variation in this taxon is thus more likely to capture a fuller range of variation given that even coeval localities in close geographic proximity probably represent distinct populations, let alone those from geographically distant localities.
SMP VP-44 exhibits essentially no deviations from the typical morphology of Anaschisma browni (Case, Reference Case1922; Lucas et al., Reference Lucas, Rinehart, Heckert, Hunt and Spielmann2016). The skull is relatively symmetrical, although slight transverse distortion of the skull has resulted in a slight offset along the longitudinal axis of major features (e.g., anterior orbital margin). The course of the lateral line grooves is also consistent with previous descriptions, although it may be noted that the post-orbital groove is angled more sharply posteromedially as it passes from the jugal onto the postorbital and then onto the supratemporal; in some other individuals, the groove passes medially onto the postorbital and then posteriorly onto the supratemporal. This variation falls within the range previously reported for A. browni and is not considered taxonomically meaningful in this instance. It may be noted (only in a qualitative sense) that specimens of A. browni from the Popo Agie Formation have proportionately larger nares that are nearly the same size as the orbits (e.g., Branson and Mehl, Reference Branson and Mehl1929; Gee et al., Reference Gee, Parker and Marsh2020) when compared to conspecifics from more near-equatorial latitudes in which the nares are distinctly smaller than the orbits (e.g., Case, Reference Case1922; Lucas et al., Reference Lucas, Rinehart, Heckert, Hunt and Spielmann2016). The importance of this, if it is biologically important, is unclear. The relative size of the nares and the orbits in SMP VP-44 is somewhat intermediate to the end members of A. browni, but this should not be interpreted as a suggestion of gradational evolution associated with biogeographic trajectories. The absence of any substantial morphological variation across a broad geographic range in A. browni is not necessarily surprising given the morphological conservatism across metoposaurids in general, but this description provides clear, positive evidence for a hypothesis of intraspecific conservatism.
Temnospondyl turnover in North America
The Late Triassic temnospondyl record of North America is notable for two attributes. Firstly, North America preserves the most complete and diverse record of small-bodied temnospondyls from the Late Triassic (Wilson, Reference Wilson1948; Bolt and Chatterjee, Reference Bolt and Chatterjee2000), including putative stem caecilians (Pardo et al., Reference Pardo, Small and Huttenlocker2017; So et al., Reference So, Kufner, Huttenlocker, Pardo and Lovelace2018) and unequivocal crown batrachians (Stocker et al., Reference Stocker, Nesbitt, Kligman, Paluh, Marsh, Blackburn and Parker2019). Small-bodied stereospondyls are rare in any setting in the Mesozoic, and their ecology and phylogenetic relatedness to Lissamphibia remain to be more comprehensively explored. Secondly, the record of large-bodied taxa is conspicuously dominated by metoposaurids, a pattern that otherwise occurs only in regions with a substantially smaller history and body of collection of both metoposaurids and of terrestrial tetrapods at large (e.g., Morocco, Madagascar). As noted above, the only record of large-bodied, non-metoposaurid temnospondyls from the Late Triassic of North America is Calamops paludosus from the lower Stockton Formation (Solesbury Member; Carnian). Assuming that the correlation of the middle-upper New Oxford Formation with the Lockatong Formation is correct, C. paludosus likely pre-dates all metoposaurid occurrences in eastern North America. However, we note that the specimens collected near Emigsville (including AMNH FARB 1850 and 1863) are potentially from near the base of the New Oxford Formation, which may make them close to temporally equivalent with C. paludosus.
The longstanding perception of metoposaurid exclusivity remains true in the Late Triassic of western North America, which is also the source of the diminutive temnospondyl taxa. In contrast, the Middle Triassic record of temnospondyls in this region is substantially more diverse, similar to other geographic regions, and preserves brachyopoids, non-metoposaurid trematosauroids, and capitosauroids (e.g., Welles and Cosgriff, Reference Welles and Cosgriff1965; Schoch, Reference Schoch2000; Rinehart et al., Reference Rinehart, Lucas and Schoch2015). All three of these clades persist into the Late Triassic in other geographic regions where they sometimes co-occur with metoposaurids, as in western Europe (e.g., Milner and Schoch, Reference Milner and Schoch2004; Sulej and Majer, Reference Sulej and Majer2005), or succeed metoposaurids, as in India (e.g., Sengupta, Reference Sengupta2003). The extensive sampling in the southwestern region of the United States for more than a century suggests that this stark faunal disparity is not the result of poor sampling, but rather that metoposaurids were genuinely the only large-bodied temnospondyls in these basins by the Late Triassic. However, the fossil record of this transition is not captured in this part of North America, with the regional Tr-3 unconformity representing a several-million-year gap between deposition of the Holbrook Member of the Moenkopi Formation (Anisian) and the next occurrences of temnospondyls in the late Carnian of Wyoming and Texas. There are no definitive records of temnospondyls from the Ladinian of North America, although some of the other poorly constrained horizons on the east coast cannot be totally excluded from being Ladinian (e.g., the Economy Member of the Wolfville Formation; Sues and Olsen, Reference Sues and Olsen2015). Consequently, the mechanisms that contributed to the development of a metoposaurid zone of exclusivity by the latest Carnian (Otischalkian LVF) in North America remain unclear. Climatic variation along paleolatitudinal gradients (e.g., Sues and Olsen, Reference Sues and Olsen2015) and climatic instability (e.g., Whiteside et al., Reference Whiteside, Lindström, Irmis, Glasspool, Schaller, Dunlavey, Nesbitt, Smith and Turner2015) are popular hypotheses invoked for explaining the distribution of amniotes (e.g., Dunne et al., Reference Dunne, Farnsworth, Greene, Lunt and Butler2020), such as the absence of the herbivorous sauropodomorph dinosaurs, but hypotheses specifically regarding the distribution of large-bodied stereospondyls have not been formulated. A large body of recent work has emerged examining the role of the Carnian Pluvial Episode (CPE) in shaping terrestrial tetrapod evolution, but few have explicitly addressed temnospondyls. Buffa et al. (Reference Buffa, Jalil and Steyer2019) and Fortuny et al. (Reference Fortuny, Arbez, Mujal and Steyer2019) suggested a correlation between metoposaurid diversification and the CPE, which has been increasingly linked to major events of Triassic tetrapod evolution (e.g., Simms and Ruffell, Reference Simms and Ruffell1989, Reference Simms and Ruffell1990; Ruffell et al., Reference Ruffell, Simms and Wignall2016; Dal Corso et al., Reference Dal Corso, Bernardi, Sun, Song, Seyfullah, Preto and Gianolla2020), particularly to the radiation of dinosaurs (e.g., Benton et al., Reference Benton, Bernardi and Kinsella2018; Bernardi et al., Reference Bernardi, Gianolla, Petti, Mietto and Benton2018; Mancuso et al., Reference Mancuso, Benavente, Irmis and Mundil2020). However, as those workers also noted, it remains to be tested how this climate event would have favored some clades of large-bodied stereospondyls over others; most studies remain focused on amniotes. The occurrence of the non-metoposaurid metoposauroid Callistomordax kugleri in the upper Ladinian (Schoch, Reference Schoch2008) confers another line of evidence that the timing of metoposaurid origination and diversification may be tied to global events in the middle Carnian.
A likely possibility that would reconcile this uncertainty is that presently un- or underrecognized physiological disparity between taxa is a major explanator of Mesozoic temnospondyl distribution (e.g., Muhkerjee et al., Reference Mukherjee, Ray and Sengupta2010). It has long been known that the majority of Mesozoic taxa are medium- to large-bodied aquatic taxa, but it has not been shown that they were all ecologically similar; the frequent colloquial parallel to extant crocodilians may obfuscate the true ecological diversity of large stereospondyls. Indeed, finite element analysis (FEA) of various stereospondyls to infer feeding behavior has identified the potential for ecological differentiation among the distinct morphotypes (e.g., Fortuny et al., Reference Fortuny, Marcé-Nogué, de Esteban-Trivigno, Gil and Galobart2011, Reference Fortuny, Marcé-Nogué, Gil and Galobart2012, Reference Fortuny, Marcé-Nogué, Steyer, de Esteban-Trivigno, Mujal and Gil2016, Reference Fortuny, Marcé-Nogué and Konietzko-Meier2017; Lautenschlager et al., Reference Lautenschlager, Witzmann and Werneburg2016; Konietzko-Meier et al., Reference Konietzko-Meier, Gruntmejer, Marcé-Nogué, Bodzioch and Fortuny2018). Additionally, an increasing body of histological literature has identified disparate physiological capabilities and growth patterns among closely related taxa (e.g., Konietzko-Meier and Klein, Reference Konietzko-Meier and Klein2013; Muhkerjee et al., Reference Mukherjee, Sengupta and Rakshit2020), indicating differing responses to local environmental conditions. Metoposaurids are actually the best sampled clade in this regard. Most of the comparative work has focused on disparities between the Moroccan Dutuitosaurus ouazzoui (Steyer et al., Reference Steyer, Laurin, Castanet and de Ricqlès2004) and the Polish Metoposaurus krasiejowensis (Konietzko-Meier and Klein, Reference Konietzko-Meier and Klein2013; Konietzko-Meier and Sander, Reference Konietzko-Meier and Sander2013). The former appears to have been under more pronounced environmental stress that forced it to aestivate for a prolonged time (evidenced by discrete lines of arrested growth) compared to much shorter, sporadic cessations in growth in the latter. Teschner et al. (Reference Teschner, Sander and Konietzko-Meier2018) further documented two distinct histotypes within M. krasiejowensis that implied distinct environmental conditions for perhaps two distinct local populations. Most recently, histological data from the relatively high paleolatitude Indian taxon, Panthasaurus maleriensis, indicates a similarly temperate climate to that of Poland, with a similar environmental signal to that found in M. krasiejowensis (Teschner et al., Reference Teschner, Chakravorti, Sengupta and Konietzko-Meier2020).
Other clades of large-bodied stereospondyls are generally not as well-sampled histologically, thereby precluding robust comparisons with Metoposauridae. However, a number of studies do hint at cryptic ecological and physiological differentiation that could in turn explain the different geographic ranges of various clades. Muhkerjee et al. (Reference Mukherjee, Ray and Sengupta2010, Reference Mukherjee, Sengupta and Rakshit2020) suggested that some paracylotosaurid capitosaurs were more terrestrially capable than other stereospondyls, perhaps indicating a more shallow-water spatial occupation when in water. Some differences could also relate to the tolerance of various taxa. For example, the ability to tolerate differing environments seems to have been particularly high for the long-ranging plagiosaurid Gerrothorax pulcherrimus Nilsson, Reference Nilsson1934, and it has been suggested that this taxon was a “pioneer species” that inhabited more unstable areas in which other taxa could not survive (Sanchez and Schoch, Reference Sanchez and Schoch2013). Similarly, greater tolerance for cold water temperatures has been suggested to explain the persistence of post-Triassic brachyopoids at high paleolatitudes by means of exclusion of crocodilians (Warren et al., Reference Warren, Rich and Vickers-Rich1997). Most other clades remain very poorly sampled—there is essentially no modern histology on non-metoposaurid trematosaurs, for example—but the available data already demonstrate some apparent differences between and within clades. Integrating these data with the observable external anatomy would likely elucidate a clearer narrative regarding stereospondyl distribution.
In lieu of histological data for most metoposaurids, the paleolatitudinal range of these clades can provide alternative insight. All metoposaurids ranged from ~35°N to 35°S (see Brusatte et al., Reference Brusatte, Butler, Mateus and Steyer2015, fig. 13), as demonstrated by the central European and Indian populations, which bracket this range. Other stereospondyls are known from higher paleolatitudes in both hemispheres, particularly brachyopoids, although Late Triassic temnospondyls as a whole were more abundant at the lower latitudes where presumably seasonality was less pronounced (Dunne et al., Reference Dunne, Farnsworth, Greene, Lunt and Butler2020), much like lissamphibians today (Pyron and Wiens, Reference Pyron and Wiens2013). The North American metoposaurid-bearing basins, specifically those in the southwestern United States, would have been the closest to the equator; the Argana Basin in Morocco lay intermediate between these basins and those of central Europe. In both North America and Morocco (the western part of metoposaurids’ range), there is no evidence for other large-bodied stereospondyl taxa co-existing with or succeeding metoposaurids. The general paleoclimate of the southwestern United States has been well studied from a variety of perspectives, especially in the Chinle Formation (e.g., Dubiel et al., Reference Dubiel, Parrish, Parrish and Good1991, Reference Dubiel, Hasiotis, Davidson, Leleu and North2011; Atchley et al., Reference Atchley, Nordt, Dworkin, Ramezani, Parker, Ash and Bowring2013; Baranyi et al., Reference Baranyi, Reichgelt, Olsen, Parker and Kürschner2018; Jin et al., Reference Jin, Dworkin, Atchley and Nordt2018; Rasmussen et al., Reference Rasmussen, Mundil, Irmis, Geisler, Gehrels, Olsen and Kent2020) and indicates a humid climate during deposition of the lower units, with a progressively arid and seasonally pronounced shift throughout the Norian. Similar seasonality has been inferred from histological data of the Moroccan metoposaurids (Steyer et al., Reference Steyer, Laurin, Castanet and de Ricqlès2004), while the Polish and Indian taxa seem to have been subjected to a comparatively mild climate that did not lead to growth cessations (Konietzko-Meier and Klein, Reference Konietzko-Meier and Klein2013; Teschner et al., Reference Teschner, Chakravorti, Sengupta and Konietzko-Meier2020). This physiological variation is most likely tracking environmental variation since metoposaurid anatomy is so highly conserved, suggesting that the paleolatitudinal restriction of metoposaurids at large also owes to environmental barriers, whether by availability of corridors for dispersal or through various aspects of the climate (e.g., seasonality, ephemerality of freshwater bodies).
Considering the sampling in Late Triassic terrestrial ecosystems, including those that produce other temnospondyl remains, the apparent latitudinal restriction of metoposaurids seems fairly robust as a crude approximation of the true range. In this vein, the recent report of metoposaurid “osteoderms” from Zimbabwe (~40°S) by Barrett et al. (Reference Barrett, Sciscio, Viglietti, Broderick, Suarez, Sharman and Jones2020) warrants some discussion here. This is an intriguing novel occurrence, not yet addressed by temnospondyl workers, that has already been propagated by several studies (Dunne et al., Reference Dunne, Farnsworth, Greene, Lunt and Butler2020; Sciscio et al., Reference Sciscio, Viglietti, Barrett, Broderick, Munyikwa, Chapelle, Dollman, Edwards, Zondo and Choiniere2020) and has been utilized as a line of evidence for broader claims, such as biome analogy with geographically disparate deposits (Barrett et al., Reference Barrett, Sciscio, Viglietti, Broderick, Suarez, Sharman and Jones2020). However, there are several reasons to exercise caution with this purported occurrence. Firstly, metoposaurids do not have osteoderms (Sciscio et al., Reference Sciscio, Viglietti, Barrett, Broderick, Munyikwa, Chapelle, Dollman, Edwards, Zondo and Choiniere2020 more generically termed these “dermal bones”), and the photographs indicate only flat elements with circular ornamentation. These appear to be pectoral in nature, which, as previously noted here, are very hard to properly refer in isolation and when fragmentary. The authors neither described the material (it is only figured in the supplementary data) nor justified their taxonomic identification and, most crucially, the specimens were not collected. Reported occurrences without voucher specimens are problematic for the sake of reproducibility (Bell et al., Reference Bell, Gauthier and Bever2010). Circumstantial evidence (e.g., regional monotaxicity, temporal occurrence) does not lend particular credence to a specific temnospondyl clade in the same way that it does for North America (e.g., chigutisaurids occur in both the lower and upper horizons of the Elliot Formation of South Africa; Warren and Damiani, Reference Warren and Damiani1999). Finally, phytosaurs occur at high paleolatitudes of both hemispheres without metoposaurids (e.g., Bonaparte, Reference Bonaparte1966; Brusatte et al., Reference Brusatte, Butler, Niedźwiedzki, Sulej, Bronowicz and Satkūnas2013), while other stereospondyl clades have been reported to occur alongside phytosaurs in the absence of metoposaurids. Examples include chigutisaurids in India (Sengupta, Reference Sengupta1988), plagiosaurids in Thailand (Suteethorn et al., Reference Suteethorn, Janvier and Morales1988), and plagiosaurids and capitosauroids in Luxembourg (Milner et al., Reference Milner, Duffin and Delsate1996). In Europe, metoposaurids co-occur with phytosaurs and at least one other large-bodied temnospondyl clade (Milner and Schoch, Reference Milner and Schoch2004; Sulej and Majer, Reference Sulej and Majer2005). Lastly, metoposaurids were replaced by other clades in Europe (by capitosauroids; Sulej and Majer, Reference Sulej and Majer2005) and India (by chigutisaurids; Sengupta, Reference Sengupta1988, Reference Sengupta2003) during the late Norian and Rhaetian. Radioisotopic dating of the phytosaur-bearing site in Zimbabwe constrained a maximum age of 209.2 ± 4.5 Ma, near the Norian-Rhaetian boundary (Barrett et al., Reference Barrett, Sciscio, Viglietti, Broderick, Suarez, Sharman and Jones2020). Most metoposaurid-bearing localities are considered to occur in the Carnian, including those from India and Morocco (Buffa et al., Reference Buffa, Jalil and Steyer2019). The Malagasy metoposaurids, the geographically closest to the Zimbabwe occurrence, are of uncertain stratigraphic placement (Fortuny et al., Reference Fortuny, Arbez, Mujal and Steyer2019), but the upper Isalo II interval (Norian) does not contain any metoposaurid material (Burmeister et al., Reference Burmeister, Flynn, Parrish and Wyss2006), and the material is probably from the lower Isalo II and thus Carnian in age. The only metoposaurid records from the Rhaetian (208.5–201.3 Ma) are from North America (Hunt, Reference Hunt1993; Spielmann et al., Reference Spielmann, Lucas and Heckert2007; Spielmann and Lucas, Reference Spielmann and Lucas2012), and there are practically no other post-Norian occurrences of temnospondyls other than brachyopoids (see Konietzko-Meier et al., Reference Konietzko-Meier, Werner, Wintrich and Sander2019, for summary, and Maisch et al., Reference Maisch, Matzke and Sun2004, for a possible Jurassic trematosauroid). Therefore, the occurrence of a metoposaurid from Zimbabwe should be considered unreliable until more diagnostic material is both collected and described. We emphasize that this discussion should not be interpreted to mean that we are confident that the material is not metoposaurid, but rather that the only data that support metoposaurid affinities are weakly supported circumstantial lines of evidence (i.e., large-bodied stereospondyl co-occurring with a phytosaur). This discussion also underscores one of the key motivations of our own study: to provide a rigorous, detailed justification for taxonomic identification of material that may be considered to be a geographic outlier.
Conclusions
We have provided a thorough description of the most complete and confidently taxonomically referable material of metoposaurid temnospondyls from the Late Triassic of eastern North America. Our study confirms previous authors’ cursory identification of the Zions View temnospondyl material to what is now referred to as Anaschisma browni (Fig. 9). We further did not identify any intraspecific variation, a somewhat surprising conclusion given the wide geographic separation from the next closest population of A. browni in west Texas. The Zions View material represents the most complete metoposaurid occurrence from the eastern seaboard and highlights the potential for localities outside of the classic collecting areas. With the confident verification of A. browni in Pennsylvania, this taxon is by far the most widespread metoposaurid, occurring across practically all of the continental United States and hinting at the potential for a vast undersampling/biased sampling of the clade.

Figure 9. Life reconstruction of the Zions View Anaschisma browni by Sergey Krasovskiy.
The Late Triassic deposits on the eastern seaboard are less well explored than the southwest and south-central regions, and, accordingly, have produced substantially less material of virtually all documented tetrapod clades from this time, but the importance of these records for understanding large-scale trends and distributions should not be understated. Within North America, the Gettysburg Basin is significant because it may be the only geographic region to capture the fine temporal scale turnover among large-bodied stereospondyls, which almost certainly occurred between the Ladinian–early Carnian when there is no definitive record of temnospondyls from North America. Other Carnian metoposaurid-bearing deposits include the Popo Agie Formation in Wyoming (e.g., Branson and Mehl, Reference Branson and Mehl1929), the Camp Springs Conglomerate (sometimes classified as a member of the Santa Rosa Formation or a formation of the Dockum Group) in Texas (e.g., Case, Reference Case1931, Reference Case1932), the Cumnock Formation in North Carolina, and the Wolfville Formation in Nova Scotia (e.g., Sues and Olsen, Reference Sues and Olsen2015). Non-metoposaurid temnospondyls of any age have never been reported from North Carolina, and the only ones reported from Wyoming are small-bodied fossorial specimens from the Popo Agie Formation that resemble Chinlestegophis jenkensi Pardo et al., Reference Pardo, Small and Huttenlocker2017 from the Chinle Formation and Rileymillerus cosgriffi Bolt and Chatterjee, Reference Bolt and Chatterjee2000 from the Dockum Group (So et al., Reference So, Kufner, Huttenlocker, Pardo and Lovelace2018). In eastern New Mexico, the Dockum Group unconformably overlies the Anton Chico Member of the Moenkopi Formation, which has also produced a high diversity of non-metoposaurid temnospondyls in Arizona. The Wolfville Formation is thus the only other unit that may directly capture the North American transition from a diverse temnospondyl assemblage to a metoposaurid-exclusive (among large-bodied clades) assemblage at a local geographic scale (Sues and Olsen, Reference Sues and Olsen2015 were uncertain of the Anisian–Ladinian age of the Economy Member of the Wolfville Formation). In this respect, localities such as Zions View have far greater potential than the classic collecting areas to inform on numerous paleobiological aspects of temnospondyls and tetrapods more broadly, such as paleobiogeographic distribution and dispersal and morphological variation associated with broad ranges. Either renewed or additional work outside of the historically sampled depositional basins (e.g., the Popo Agie Formation in Wyoming and Chinle Formation exposures outside of Arizona) will be important for further refining the nature and setting of the evolutionary history of not only temnospondyls but of all Late Triassic tetrapods.
Acknowledgments
Thanks to the reviewers, W. Parker and V. Buffa, and the editor, H.-D. Sues, for constructive feedback that greatly improved this manuscript. BMG acknowledges previous graduate studies funding support from the University of Toronto and current postdoctoral funding from NSF grant #ANT-1947094 (to C. Sidor). Thanks to M. Norell and C. Mehling for permission to visit the collections at the American Museum. We thank S. Krasovskiy for the wonderful reconstruction of the Zions View Anaschisma browni.