



Introduction
Basilosaurus cetoides (Owen, Reference Owen1839) is an extinct cetacean common to the southeastern coastal plain of the United States. The most complete B. cetoides skeleton from Georgia, USA, to date was discovered by employees of the Georgia Department of Natural Resources during a routine fish survey of the Flint River in downtown Albany (Dougherty County) in the fall of 2010. Five post-thoracic vertebrae, portions of at least two additional post-thoracic vertebrae, and possible rib fragments were observed at the site. Excavation of this specimen began in August 2012, but by October 2012 the site was looted, and most of the skeleton was lost to science. All that remains of this Basilosaurus are portions of a single vertebra, additional bone fragments, photographs, field notes, and sketches. Despite the loss of the majority of this specimen, its discovery increases the number of reported Basilosaurus localities in Georgia to five and provides potential for further understanding the late Eocene landscape of the southeastern United States. Here we explore the temporal, environmental, and geographic distribution of Basilosaurus in the Atlantic and Gulf coastal plains to re-evaluate the position of the late Eocene shoreline and investigate possible limits to the distribution of this taxon.
Background
Basilosauridae are fully aquatic archaeocete cetaceans that occupied marine environments around the world during the Bartonian and Priabonian (late middle Eocene to late Eocene). They are known from deposits in Antarctica (e.g., Buono et al., Reference Buono, Fernández, Reguero, Marenssi, Santillana and Mörs2016), Austria (Uhen and Tichy, Reference Uhen and Tichy2000), Egypt (see Gingerich, Reference Gingerich1992, and the references therein), Germany (Uhen and Berndt, Reference Uhen and Berndt2008), Italy (Pilleri and Cigala Fulgosi, Reference Pilleri, Cigala Fulgosi and Pilleri1989), Jordan (Zalmout et al., Reference Zalmout, Mustafa and Gingerich2000), Libya (Wight, Reference Wight, Salem and Busrewil1980), Morocco (e.g., Gingerich and Zouhri, Reference Gingerich and Zouhri2015), New Zealand (Köhler and Fordyce, Reference Köhler and Fordyce1997), Pakistan (Gingerich et al., Reference Gingerich, Arif, Bhatti, Anwar and Sanders1997), Peru (Martínez-Cáceres and de Muizon, Reference Martínez-Cáceres and de Muizon2011), Russia (Kalmykov, Reference Kalmykov2012), Senegal (Elouard, Reference Elouard1966), Tunisia (Batik and Fejfar, Reference Batik and Fejfar1990), Ukraine (Gol'din and Zvonok, Reference Gol'din and Zvonok2013), the United Kingdom (Halstead and Middleton, Reference Halstead and Middleton1972), and the United States (see Uhen, Reference Uhen2013, and the references therein, as well as Uhen and Taylor, Reference Uhen and Taylor2020). Basilosauridae includes 19–24 species, depending on the equivocal interpretations of certain specimens (e.g., Uhen, Reference Uhen2013, Reference Uhen, Würsig, Thewissen and Kovacs2018; Gingerich and Zouhri, Reference Gingerich and Zouhri2015; van Vliet et al., Reference van Vliet, Bosselaers, Vahldiek, Paymans and Verheijen2020). They are characterized by: a lack of upper third molars; the presence of multiple, well-developed accessory denticles on cheek teeth; modification of forelimbs into flippers; greatly reduced hind limbs with no articulation between the pelvic girdle and sacrum; and an elevated number of posterior thoracic and/or lumbar vertebrae (Uhen, Reference Uhen2013). Basilosauridae has often been divided into the subfamilies Dorudontinae and Basilosaurinae, with the latter exhibiting a larger body size and elongated posterior thoracic, lumbar, and anterior caudal vertebrae (e.g., Uhen, Reference Uhen and Thewissen1998). However, this distinction has not often been followed in recent years due to the uncertainty of phylogenetic relationships among basilosaurids and the possibility that one of these subfamilies may be paraphyletic (Uhen, Reference Uhen2013).
The first known archaeocete specimens were an elongated vertebral centrum and partial dentary discovered in Louisiana in the early nineteenth century. Harlan (Reference Harlan1834) coined the genus Basilosaurus (meaning “king lizard”) for this specimen based on the interpretation that these elements belonged to a marine reptile, but he failed to give the taxon a specific epithet. Owen (Reference Owen1839) recognized the mammalian nature of the material and coined the name Zeuglodon cetoides, with the specific epithet signifying the taxon's cetacean affinities. Gibbes (Reference Gibbes1847) recognized the priority of Harlan's genus and proposed the new combination Basilosaurus cetoides for this taxon. The genus has come to include a well-known Egyptian species, B. isis (Andrews, Reference Andrews1904), and B. drazindai (Gingerich et al., Reference Gingerich, Arif, Bhatti, Anwar and Sanders1997), a proposed species from Pakistan that was based on a single, well-preserved lumbar vertebra.
Basilosaurus is so abundant along the Gulf Coast of the United States that it has been adopted as an official state fossil of both Mississippi and Alabama. Reel (Reference Reel1972) speculated that a large number of fossil whales were preserved in this region because its warm climate, ample food sources, and openness to the Atlantic Ocean made it an ideal mating area during the late Eocene. Regardless of the reason for their abundance, documenting the distribution of Basilosaurus through space and time has implications for understanding the paleogeography of the southeastern coastal plain of the United States during the late Eocene.
Materials and methods
The primary specimen in this study is GSM 1465, which is the most complete Basilosaurus discovered in Georgia to date. The fossil was embedded in the late Priabonian Ocala Limestone Formation along the banks of the Flint River in Albany, Georgia, USA (Paleobiology Database collection 138667). It was oriented in an approximately east-west position, perpendicular to the direction of river flow, with the anterior to the east. Fossils found in association with the Basilosaurus were an isolated carcharhiniform shark vertebra, the bivalve Amusium ocalanum Dall, and an unidentified, microscopic gastropod, all of which were collected prior to looting. The Basilosaurus fossil is described based on measurements, field notes, and photographs (Fig. 1). Measurements of length, width, and height were collected from each vertebra, when preservation allowed, and compared to data from Buchholtz (Reference Buchholtz2001) to determine the position of each along the vertebral column.

Figure 1. (1) View of GSM 1465 in situ, upon discovery; 12-inch ruler for scale. (2) Illustration of vertebrae visible in (1). (3) Close up of the three most complete vertebrae. (4) Illustration of vertebrae visible in (3). Photography by Rob Weller (1) and Arthur Berger (3).
Locations of Basilosaurus fossils in North America were sourced from the Paleobiology Database (PBDB) (paleobiodb.org; as downloaded on June 30, 2020) and the literature. GSM 1465 was already included in the PBDB based on an abstract (Smith et al., Reference Smith, Hastings and Bebej.2013). The following data, when available, were recorded for each locality (Supplemental Data 1): (1) county and state, (2) geologic age, (3) site stratigraphy, (4) environment of deposition, (5) latitude and longitude, (6) literature reference, and (7) PBDB collection number. All data from the PBDB were crosschecked against the original references. The geologic age of a Basilosaurus locality lacking explicit age data was interpreted from site stratigraphy.
All Basilosaurus localities (N = 61) were mapped using ArcMap 10.6. Basilosaurus localities with stratigraphic data (N = 53) were added to a schematic of late Eocene stratigraphy to illustrate the temporal distribution of the genus. Basilosaurus localities with more specific stratigraphic information (N = 36) were sorted into four time bins: early Bartonian, Bartonian/Priabonian boundary, early to middle Priabonian (in which the upper boundary of the Pachuta Marl Member of the Yazoo Clay Formation marks the end of the “middle Priabonian” for this study), and late Priabonian. These 36 sites were coded by time bin and plotted on a second map. The time bins were chosen with the intention of including as many specimens as possible, given the limited stratigraphic data for some specimens. Different time bins were considered, but these differences would not have affected the overall picture of Basilosaurus dispersal during the late Eocene. For example, substituting the late Priabonian category with a middle to late Priabonian category would change the designation for two Georgia specimens, including GSM 1465, but the map would still show that dispersal was generally later in the eastern states than in the western states.
Previously published paleogeographic features of the late Eocene in the southeastern United States were added to the map of Basilosaurus localities produced in this study. Schuchert (Reference Schuchert1929, fig. 6) posited the location of a late Eocene shoreline based on Jackson (late Eocene) and Claiborne (middle Eocene) deposits, broadly defining a shoreline for, presumably, the period stretching from the end of the Bartonian to the late Priabonian. Rainwater (Reference Rainwater1964, fig. 7) posited a shoreline based on marine deposits at the base of the upper Eocene Moody's Branch Formation (late Bartonian). Reel (Reference Reel1972, fig. 5) based a late Eocene shoreline on the presence of Jackson Group sediments, as interpreted from Rainwater (Reference Rainwater1964), with the caveat that the true shoreline lies farther landward due to the erosion of original coastal sediments. Huddlestun (Reference Huddlestun1993, figs. 51, 54) postulated an approximate location of the shoreline in Georgia during the latest Priabonian based on lithology, a tentative position for the terminal Eocene low sea stand in the state, and an approximate location and the direction of the Suwannee Current, which flowed through the relatively deep Gulf Trough during the middle to late Eocene. Tew and Mancini (Reference Tew and Mancini1995, fig. 9D) presented a latest Eocene shoreline in Mississippi and Alabama based on the northernmost extent of deposition for the Shubuta Member of the Yazoo Clay. Scott (Reference Scott, Buster and Holmes2011, fig. 2.3) illustrated the axis of the Ocala Platform, a topographically high feature in Florida that arose in the post-early Cenozoic.
A new late Eocene shoreline was developed as a composite of the maximum landward portions of each previously interpreted shoreline and the maximum landward portions of a new shoreline developed using the location of Basilosaurus fossils. Because shorelines are constantly shifting features, each shoreline used in this study most likely accurately depicts the location of the shoreline for some time slice, however large or small, for the late Eocene. Our goal is to combine paleogeographic evidence from multiple sources to produce one shoreline that best represents the maximum transgression of the epicontinental sea across the Atlantic and Gulf coastal plains from the early Bartonian to the late Priabonian, which is the time when Basilosaurus inhabited North America.
Abbreviations
Ca, caudal vertebra; L, lumbar vertebra; PBDB, Paleobiology Database; S, sacral vertebra; V, vertebra.
Repositories and institutional abbreviations
Specimens examined in this study are deposited in the Georgia Southern University Museum Paleontology Collections (GSM), Statesboro, GA, USA. Additional discussed specimens are deposited in the National Museum of Natural History (USNM), Washington, DC, USA; the Academy of Natural Sciences (ANSP), Philadelphia, PA, USA; and the Florida Museum of Natural History, University of Florida (UF), Gainesville, FL, USA.
Systematic paleontology
Cetacea Brisson, Reference Brisson1762
Pelagiceti Uhen, Reference Uhen2008
Basilosauridae Cope, Reference Cope1868
Basilosaurus Harlan, Reference Harlan1834
Type species
Basilosaurus cetoides (=Zeuglodon cetoides Owen, Reference Owen1839).
Other species
Basilosaurus isis (=Zeuglodon isis Andrews, Reference Andrews1904).
Diagnosis
Basilosaurus is unique among basilosaurids for its long body (length >15 m) and post-thoracic vertebrae that are approximately twice as long as they are wide or tall (Uhen, Reference Uhen2013). Basilosaurus cetoides has been differentiated from B. isis based primarily on the former's larger size (Gingerich et al., Reference Gingerich, Smith and Simons1990), but additional work is needed to assess the validity of this distinction and whether there are any diagnostic morphological characters (Uhen, Reference Uhen2013).
Basilosaurus cetoides Owen, Reference Owen1839
Figures 1, 2
- Reference Owen1839
Zeuglodon cetoides Owen, p. 69.
- Reference DeKay1842
Zeuglodon harlani DeKay, p. 123.
- Reference Koch1845a
Hydrargos sillimanii Koch, p. 1.
- Reference Koch1845b
Hydrarchos sillimanii Koch, p. 1.
- Reference Koch1845a
Hydrarchos harlani; Koch, p. 1.
- Reference Wyman1845
Zeuglodon ceti Wyman, p. 65.
- Reference Carus1847
Basilosaurus cetoides Geinitz in Carus, p. 1.
- Reference Carus1847
Basilosaurus cetoides Reichenbach in Carus, p. 13.
- Reference Hammerschmidt1848
Basilosaurus harlani Hammerschmidt, p. 323.
- Reference Müller1849
Zeuglodon macrospondylus Müller, p. 3.
- Reference Abel1906
Alabamornis gigantea Abel, p. 450.
- Reference Kellogg1936
Basilosaurus cetoides; Kellogg, p. 15.

Figure 2. Recovered portions of GSM 1465. All were found anterior to V1, but none was in articulation with V1. Partial vertebral epiphysis (referred to as V0 in the text) in (1) exterior and (2) interior view. Fragment of vertebral epiphysis in (3) exterior and (4) interior view. Isolated bone fragment in (5) exterior and (6) interior view. Vertebral centrum fragment in (7) anterior or posterior, (8) lateral, and (9) interior view.
Holotype
Elongated vertebral centrum (ANSP 12944A) from the undifferentiated Jackson Group, Caldwell Parish, Louisiana, USA (Harlan, Reference Harlan1834, plate XX, figs. 1, 2).
Diagnosis
Basilosaurus cetoides is the only species in the genus known from the United States to date. Because GSM 1465 possesses the characteristic elongate vertebrae of a Basilosaurus and was found in the United States, the specimen is referred to B. cetoides.
Occurrence
GSM 1465 was discovered on the eastern bank of the Flint River in downtown Albany, Dougherty County, GA, USA (31.58°N, 84.15°W). GSM 1465 was encased in the late Eocene Ocala Limestone Formation of the Jackson Group. In Dougherty County, Georgia, this formation is exposed as a “white to very light pink pure to sandy aphanitic oolitic and crystalline limestone [that contains] bones of zeuglodonts … abundant foraminifera, macroshells, and bryozoan” (Wait, Reference Wait1963, p. 18). In Florida, where the formation was originally identified (Dall and Harris, Reference Dall and Harris1892), it is exposed as fine-grained, white- to cream-colored fossiliferous limestone (both packstone and grainstone) that is sometimes dolomitized and rarely contains the macroscopically visible foraminifera Lepidocyclina sp. in the lower member (Scott, Reference Scott2001). Lepidocyclina is not observed at the Flint River site, but because the Flint River tends to expose only the upper section of the Ocala (Wait, Reference Wait1963) and Lepidocyclina has been identified at outcrops in Dougherty County, the site is likely an exposed portion of the upper Ocala (middle to late Priabonian).
Fossils found at the same stratum as GSM 1465 are a single vertebra of a small, carcharhiniform shark (Dana Ehret, personal communication, 2019; Fig. 3.1), multiple specimens of the pectinid bivalve Amusium ocalanum (Dall) (David Campbell, personal communication, 2014; Fig. 3.2), as well as unidentified gastropod and bivalve shell fragments. The sedimentary provenance and associated fauna indicate a shallow marine environment of deposition.

Figure 3. Fossils found in association with GSM 1465. (1) A single vertebral centrum from an unknown carcharhiniform shark (GSM 1466) in anterior (left) and posterior (right) view. (2) Disarticulated valves of the pectinid bivalve Amusium ocalanum (GSM 4724).
Description
GSM 1465 originally consisted of seven partial to nearly complete post-thoracic vertebrae (heretofore referred to as V0–V6) and additional bone fragments. Five post-thoracic vertebrae were partially articulated (V1–V5 in Fig. 1): V1 is represented only by an epiphysis; V2 lacks a neural arch, neural spine, and transverse processes; V3 and V4 lack neural spines and transverse processes; V5 has a broken neural arch and lacks a neural spine; V2 and V4 have unfused epiphyses (Fig. 1), indicating that this specimen may not be fully mature. The size of the vertebrae, comparable to measurements reported in the literature (e.g., Uhen, Reference Uhen and Thewissen1998; Buchholtz, Reference Buchholtz2001), suggests the specimen is an adult. No juvenile specimens of Basilosaurus have been found to date, however, so no comparison between GSM 1465 and a juvenile can be done. V0 and V6, not in articulation, are represented by epiphyses (V6 in Fig. 1; V0 in Fig. 2); V6 was oriented epiphysis up, suggesting more of the bone may be encased in the rock.
V1, V2, V3, and V4 were lost to science when the site was looted. V0 was collected prior to looting and is currently held in the Vertebrate Paleontology Collections of the Georgia Southern Museum. The original excavations were conducted in drought conditions; since 2012, V5 and V6 have been inundated by the Flint River and presumably remain in place. The full amount of the skeleton preserved remains unknown.
Remarks
Many of the vertebral features that would allow for interpretation of position along the vertebral column either were not preserved or were lost when the fossils were stolen, so the proposed position of each vertebra is tentative. The possible positions of these vertebrae were interpreted using measurements of epiphyseal width and height (as measured on the anterior), anteroposterior centrum length, and qualitative morphological characteristics, as compared to a composite skeleton of Basilosaurus cetoides (USNM 4675 and USNM 12261) from Kellogg (Reference Kellogg1936), graphically represented in Buchholtz (Reference Buchholtz2001), and modified here in Figure 4. Vertebral length was obtained for V2, V3, and V5. Centrum length decreases from V2 to V3, then increases to V5. Vertebral width was obtained for V0 and V6. Width slightly increases from V0 to V6. Height was obtained for V6. There is a substantial difference between width and height for V6. Based on these dimensions, and following the identifications of Kellogg (Reference Kellogg1936), this sequence of vertebrae falls between vertebrae 34 and 43 (inclusive), either L12 through Ca3 or S2 through Ca6. The remnants of neural arches on V3 and V4 are positioned slightly towards the anterior, and the metapophyses, although broken, are positioned in such a way that suggests they were directed more anteriorly than dorsally. Based on comparison with images in Kellogg (Reference Kellogg1936, pl. 6), this morphology is most pronounced on Ca3 through Ca5. The combination of quantitative and qualitative morphological data suggests that this vertebral sequence more likely represents S2 through Ca6.
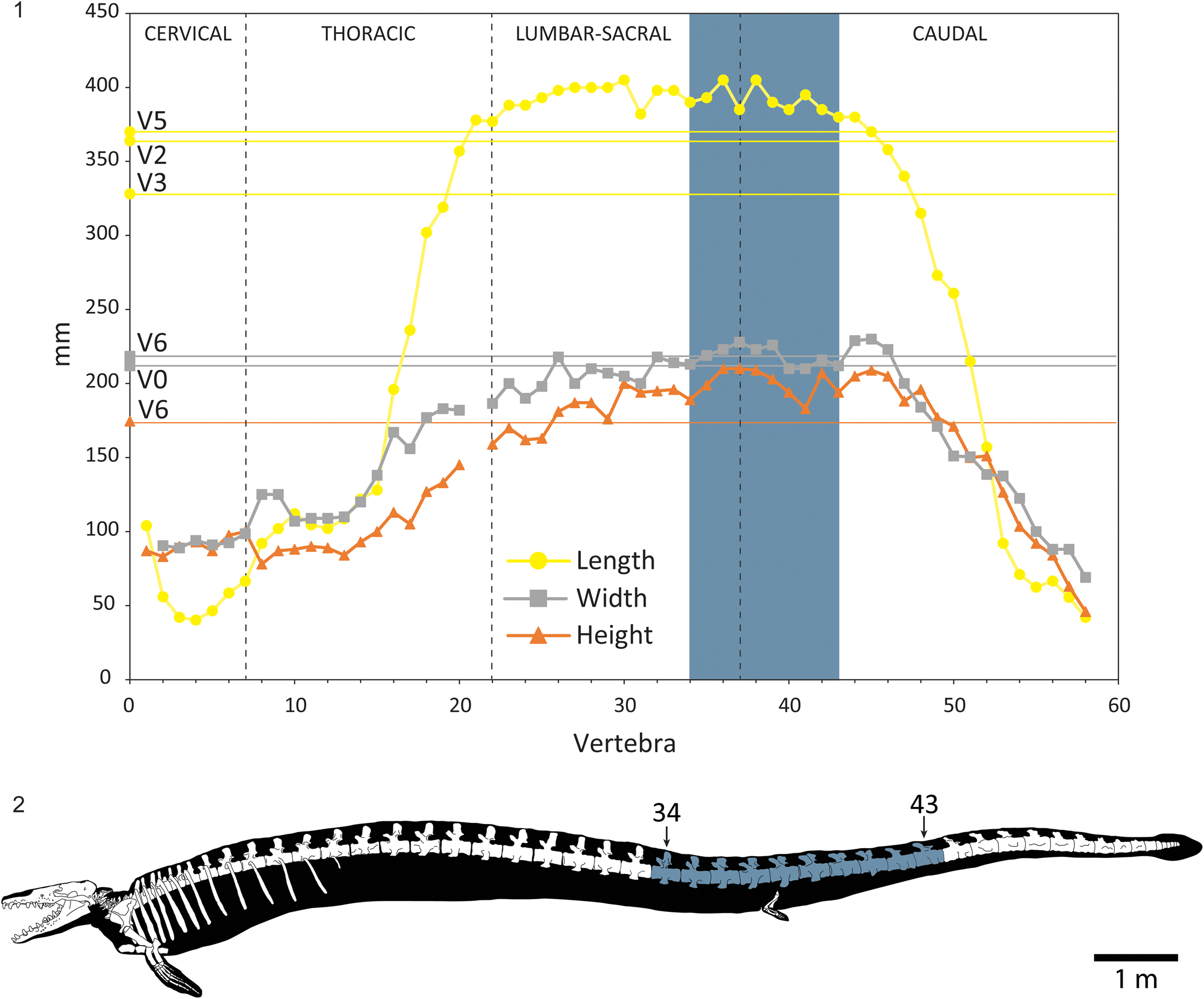
Figure 4. (1) Measurements of length, width, and height for GSM 1465 vertebrae (this study) compared to the same measurements from a composite skeleton of Basilosaurus cetoides (Buchholtz, Reference Buchholtz2001). (2) Schematic of a Basilosaurus skeleton, with possible positions of the GSM 1465 vertebrae highlighted.
However, Kellogg's (Reference Kellogg1936) vertebral designations require updating. The composite specimen of Basilosaurus cetoides was reconstructed from two non-overlapping vertebral series. The anterior end of the vertebral column (USNM 4675) included a consecutive series of seven cervical, 15 thoracic, and one lumbar vertebrae, while the posterior end of the vertebral column (USNM 12261) included a consecutive series of what Kellogg identified as 12 lumbar, two sacral, and 21 caudal vertebrae. This gives an estimated vertebral formula of C7:T15:L13:S2:Ca21. Gidley (Reference Gidley1913) justified the non-overlapping nature of these series based on anatomical differences between the lone lumbar of USNM 4675 and the anteriormost lumbar of USNM 12261, but noted that the overall lumbar count may be an underestimate. Buchholtz (Reference Buchholtz and Thewissen1998) noted that the sacral vertebrae identified by Kellogg lack intervertebral fusion and bear no articulations with the ilia, referring to them as ‘sacral lumbars’ and including them in the lumbar count (C7:T15:L15:Ca21). However, in recent years, it has been recognized that earlier archaeocetes with definitive vertebral counts possessed four fused sacral vertebrae, and vertebral counts for fully aquatic archaeocetes have typically identified the last four ‘lumbars’ anterior to the caudal region as sacral vertebrae (Bebej et al., Reference Bebej, ul-Haq, Zalmout and Gingerich2012; Uhen, Reference Uhen2014; Buchholtz and Gee, Reference Buchholtz and Gee2017). Following this convention, Kellogg's formula for B. cetoides should be revised to C7:T15:L11:S4:Ca21. Basilosaurus isis from Egypt possesses a vertebral formula of C7:T16:L19:S4:Ca20 (Bebej et al., Reference Bebej, ul-Haq, Zalmout and Gingerich2012), suggesting that the lumbar series of B. cetoides may indeed be missing multiple vertebrae. Regardless, the vertebral series described here overlaps with the terminal sacral vertebra and first six caudal vertebrae in Kellogg's composite specimen, and is tentatively identified as S4 through Ca6 (V0/S4, V1/Ca1, V2/Ca2, V3/Ca3, V4/Ca4, V5/Ca5, V6/Ca6).
Chondrichthyes Huxley, Reference Huxley1880
Carcharhiniformes Compagno, Reference Compagno, Greenwood, Miles and Patternson1973
Carcharhiniformes indet.
Figure 3.1
Occurrence
Same as GSM 1465.
Description
GSM 1466, isolated vertebral centrum (Fig. 3.1). The centrum is disk shaped with concave anterior and posterior surfaces. Two foramina for attachment of the neural and hemal arches are present dorsally and ventrally, respectively. The vertebra has an anteroposterior length of 9.1 mm, a dorsoventral height of 16.6 mm, and a width of 17.9 mm.
Remarks
Carcharhiniformes are the most abundant sharks in late Eocene sediments of Georgia (Parmley and Cicimurri, Reference Parmley and Cicimurri2003). Carcharhiniform shark vertebrae lack identifying features, so taxonomic resolution remains coarse for this specimen. However, Negaprion (lemon shark), the most abundant shark in the Priabonian Hardie Mine Local Fauna of central Georgia (Parmley and Cicimurri, Reference Parmley and Cicimurri2003), has vertebrae of similar size to GSM 1466, so it is a likely candidate for the taxon represented here.
Pterioida Newell, Reference Newell1965
Pectinidae Wilkes, Reference Wilkes1810
Amusium Röding, Reference Röding1798
Type species
Amusium pleuronectes (=Ostrea pleuronectes Linnaeus, Reference Linnaeus1758).
Amusium ocalanum Dall, Reference Dall1898
Figure 3.2
Holotype
From Levy County, Florida (Dall, Reference Dall1898, pl. 29).
Occurrence
Same as GSM 1465. Amusium ocalanum is restricted to the southeastern United States; it is common in the Ocala Limestone of Georgia and Florida and less common in similarly aged clastic units (Glawe, Reference Glawe1974).
Description
GSM 4724, internal mold of one valve and multiple disarticulated valves (Fig. 3.2). Valves are convex and not quite bilaterally symmetrical, apical angle is acute, and internal ribbing is present. Specimens recovered at this location range in size from ~10 mm to >30 mm at their greatest width.
Remarks
Valves may exhibit well-defined, obvious ribbing, or may be nearly smooth (Dall, Reference Dall1898).
Results
Stratigraphic and biogeographic distribution of Basilosaurus in North America.—Fossil occurrences of Basilosaurus in North America are restricted to the Bartonian and Priabonian of the Atlantic and Gulf coastal plains (Figs. 5, 6). The oldest and most southern Basilosaurus fossil is from the early Bartonian portion of the Avon Park Formation in Florida. No other Basilosaurus fossils of early or middle Bartonian age have been reported on the continent to date. Basilosaurus next occurs at the Bartonian/Priabonian boundary in several locations, including western Tennessee, extending the geographic range of the taxon to the west and north. Basilosaurus exhibits its largest geographic range and greatest abundance in the early to middle Priabonian, with localities as far west as Arkansas, as far east as South Carolina, and at every state in between (except Florida). By the late Priabonian, the abundance of the taxon declined, and its distribution was limited to southeastern Mississippi, southwestern Alabama, and southwestern Georgia.
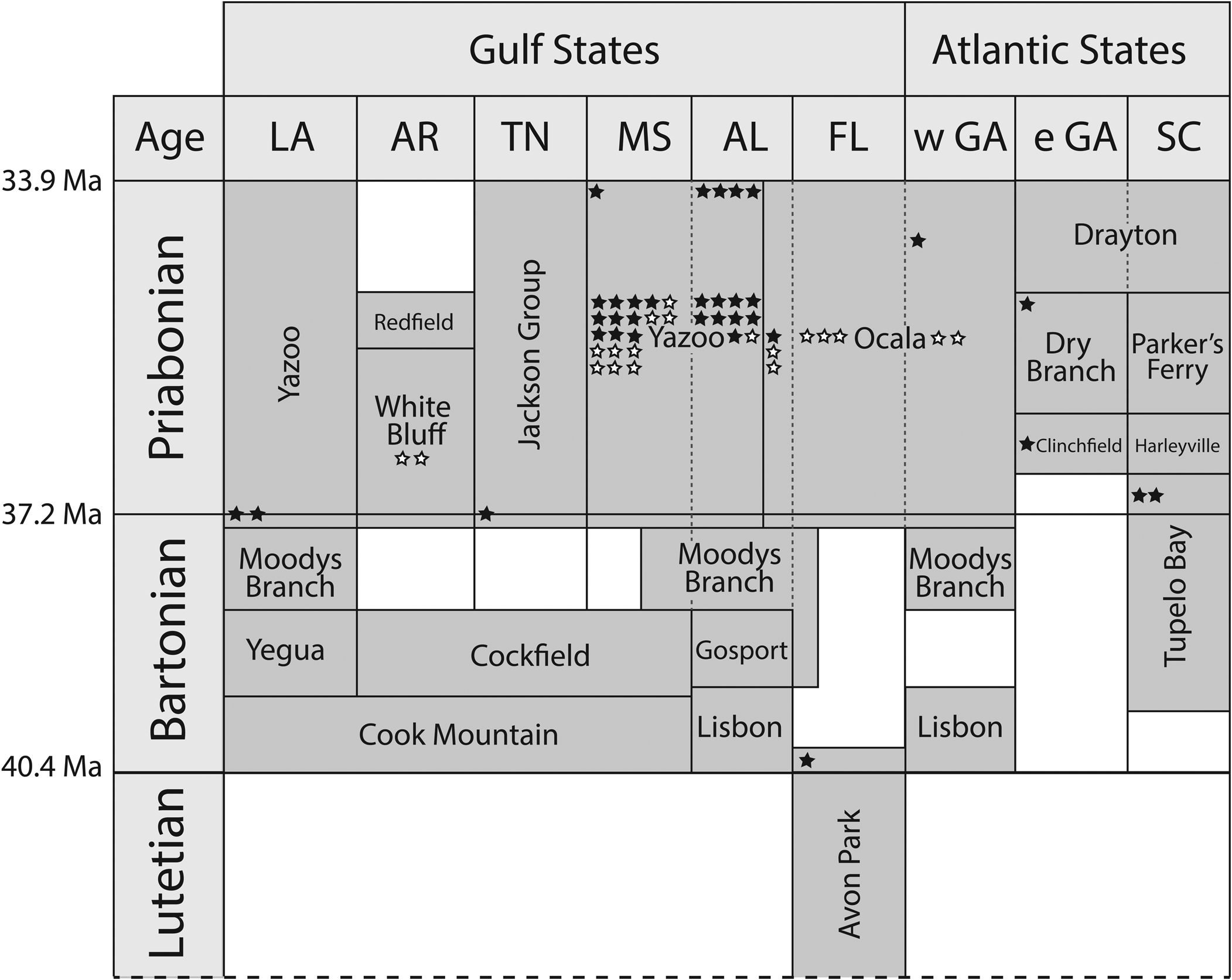
Figure 5. Stratigraphy of the Gulf and Atlantic coastal plains from the middle Lutetian to the end-Priabonian, modified from Uhen (Reference Uhen2013, fig. 7). Data from the PBDB and additional sources (Supplemental Data 1). Closed stars indicate that there is some information regarding where in the stratigraphic column the specimen was found; open stars indicate that the position within the stratigraphic column is unknown.
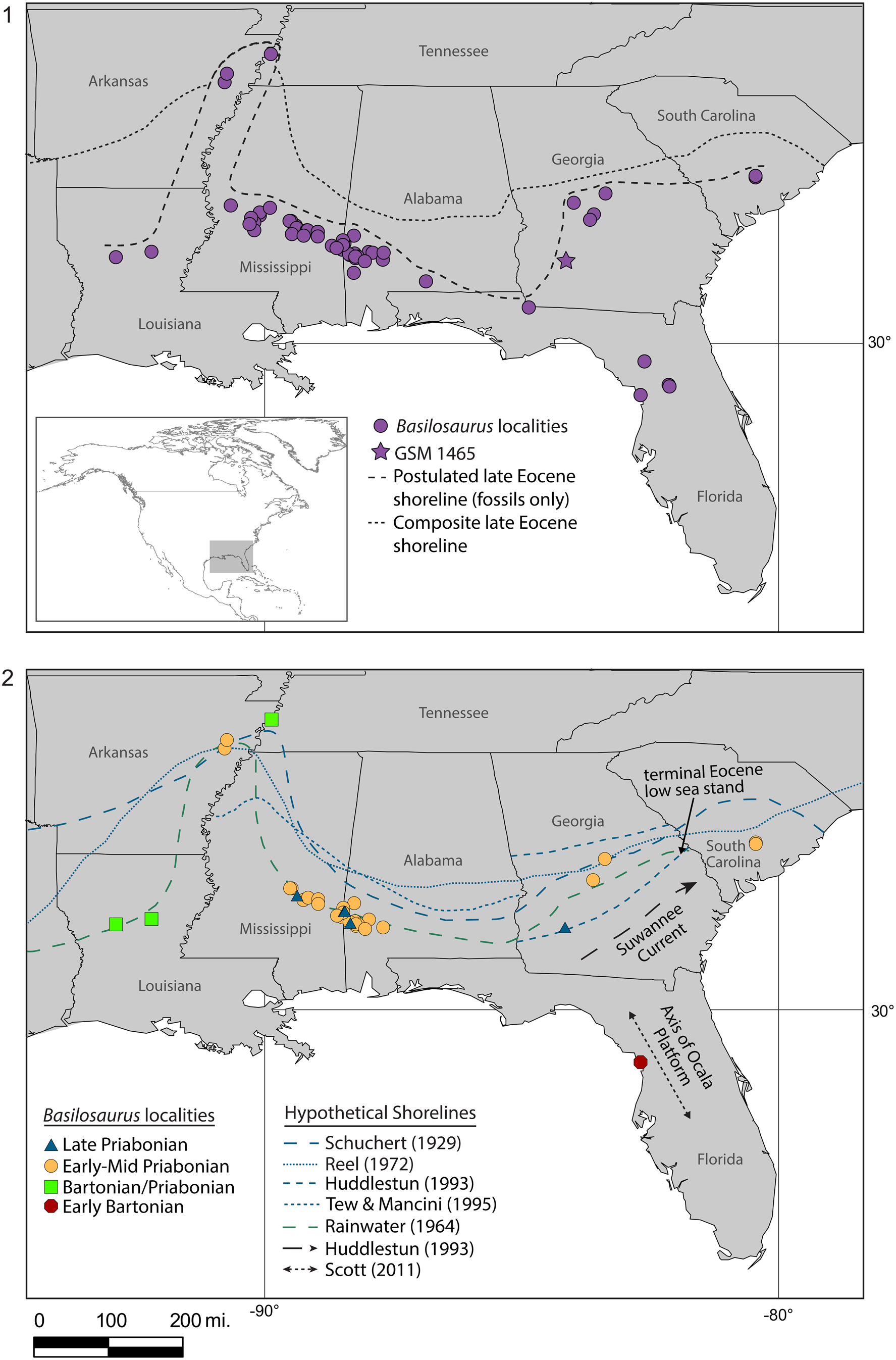
Figure 6. (1) Location of all Basilosaurus fossils listed in Supplemental Data 1, a postulated shoreline based only on these fossil occurrences, and a composite of the shorelines from Schuchert (Reference Schuchert1929), Rainwater (Reference Rainwater1964), Reel (Reference Reel1972), Huddlestun (Reference Huddlestun1993), Tew and Mancini (Reference Tew and Mancini1995), and this study. (2) Location of Basilosaurus fossils that could be sorted into one of four time bins and shorelines from the literature used to draw the composite in (1). Shoreline colors roughly correspond with the color key for stratigraphic age. The axis the of Ocala Platform is redrawn from Scott (Reference Scott, Buster and Holmes2011) and the track of the Suwannee Current through the Gulf Trough is redrawn from Huddlestun (Reference Huddlestun1993).
Paleogeography of the late Eocene shoreline in North America
A hypothetical position of the late Eocene shoreline, based on 61 Basilosaurus localities, is illustrated in Figure 6.1. Because this shoreline is defined by specimens found in a range of shallow marine environments, from nearshore to outer neritic, the true maximum extent of the late Eocene marine transgression was likely north of this hypothetical shoreline. Late Eocene shorelines from the literature are shown in Figure 6.2.
Basilosaurus localities can be roughly associated with paleogeographic features reported in the literature. The oldest reported Basilosaurus in North America (early Bartonian) lies just west of the axis of the Ocala Platform in Florida. Basilosaurus fossils of the Bartonian/Priabonian boundary broadly track the late Bartonian Rainwater (Reference Rainwater1964) shoreline. The early-middle Priabonian Basilosaurus fossils of Arkansas, Mississippi, and Alabama fall on or around the Rainwater (Reference Rainwater1964) shoreline; Arkansas specimens also align with the overlapping middle to late Eocene Schuchert (Reference Schuchert1929) and late Eocene Reel (Reference Reel1972) shorelines. In Georgia, one early–middle Priabonian locality falls on the intersection of the Schuchert (Reference Schuchert1929) and Reel (Reference Reel1972) shorelines, and another is just north of the Rainwater (Reference Rainwater1964) shoreline; early–middle Priabonian specimens of South Carolina are just south of the Reel (Reference Reel1972) shoreline. No Basilosaurus specimens from the late Priabonian are located north of any shorelines from the literature. In the west, these specimens fall on or south of the Rainwater (Reference Rainwater1964) shoreline that extends the farthest seaward. The solitary late Priabonian locality in the east is located on the terminal Eocene low sea stand (Tew and Mancini, Reference Tew and Mancini1995). Only two Basilosaurus localities are farther north than any of the shorelines in the literature: the early–middle Priabonian site in St. Francis County, Arkansas (Palmer, Reference Palmer1939), and the Bartonian/Priabonian boundary site in Lauderdale County, Tennessee (Moore and Brown, Reference Moore and Brown1969). These localities extend the composite late Eocene shoreline to the middle of Tennessee's western boundary, farther inland than previously noted (Fig. 6.1).
Environmental distribution of Basilosaurus in North America
The majority of Basilosaurus depositional environments in this study (N = 39) are described as “coastal indeterminate,” “carbonate indeterminate,” or “marine indeterminate,” and the remainder are associated with some subset of the shallow marine environment (Supplemental Data 1). Four sites are associated with deeper zones of the epeiric sea. The Ouachita and Grant County, Louisiana, sites (Harlan, Reference Harlan1834; Lancaster, Reference Lancaster, Schiebout and van den Bold1986), both in the Bartonian/Priabonian boundary time bin, are both described as offshore shelf, likely middle neritic environments. Two sites in Jasper County, Mississippi (Reel, Reference Reel1972; early–middle Priabonian), are described as offshore, likely outer neritic environments. All four localities are positioned well south of the composite shoreline of this study. Multiple sites are in closer proximity to the paleocoastline, and are described as foreshore (N = 6), estuary (N = 2), and nearshore marine (N = 1). Specimens specifically associated with a coastal environment are found in Arkansas, Georgia, and Mississippi, and, with the exception of the Arkansas locality, are not notably closer to the composite shoreline than are deeper-water specimens.
Discussion
In the late Eocene, most of the southeastern coastal plain of the United States was covered by a relatively deep epicontinental sea, which resulted from a marine transgression that began in the late Bartonian (Frederiksen, Reference Frederiksen1980). This sea was the environment in which Basilosaurus expanded its range across the region. Sea level in Mississippi reached a high in the eastern part of the state by the middle to late Priabonian (Huff, Reference Huff1970), corresponding in time and geography with the majority of Basilosaurus occurrences in the region.
Basilosaurus was present across a spectrum of shallow marine environments, from nearshore to outer neritic, indicating that it, like Basilosaurus isis at Wadi Al-Hitan in Egypt (Peters et al., Reference Peters, Antar, Zalmout and Gingerich2009), was a generalist of the epicontinental seas. Because fossils of the genus were most abundant when and where the sea level was the highest, Basilosaurus in North America may have had a slight preference for a deeper, middle- to outer neritic environments, also like B. isis at Wadi Al-Hitan (Peters et al., Reference Peters, Antar, Zalmout and Gingerich2009). Of all specimens with specific information on the environment of deposition, those associated with the deepest waters are present in Louisiana at the Bartonian/Priabonian boundary, when there was rapid, but slight, deepening of the epicontinental sea to a midsublittoral environment (Huff, Reference Huff1970), and the middle Priabonian of Mississippi, when the environment ranged from midsublittoral to shallow, outer sublittoral (Huff, Reference Huff1970; Supplemental Data 1). Nearly all Basilosaurus occurrences are well south of the late Eocene composite shoreline (Fig. 6.1), again suggesting an environmental preference for deeper zones of the shallow sea. In this way, the relative paucity of the taxon in the Atlantic Coastal Plain as compared to the Gulf Coastal Plain may be related to environmental differences, in which Basilosaurus preferred the presumably deeper waters over the Gulf.
The proximity of two Bartonian/Priabonian boundary Basilosaurus localities to the Rainwater (Reference Rainwater1964) shoreline of approximately the same age at first appears to counter the hypothesis that Basilosaurus preferred middle to outer neritic environments. These two specimens, however, are associated with offshore shelf and middle neritic environments, suggesting that the shoreline was actually farther inland at this time than shown by Rainwater (Reference Rainwater1964). Another possible counterargument involves a Basilosaurus (UF 115000) found in a nearshore, shallow subtidal environment, adjacent to the long axis of the Ocala Platform. This topographically high point in the Cenozoic of Florida (Scott, Reference Scott, Buster and Holmes2011) would have created a shallow zone around the possibly inundated platform, making it a potential death trap for the Basilosaurus. The three occurrences located at the landward edge of the Mississippi embayment, also a shallow area, may have suffered a similar fate.
Basilosaurus has not been found near the Suwannee Current in Georgia, which is somewhat surprising because the current flowed through the relatively deep Gulf Trough of the late Eocene (Huddlestun, Reference Huddlestun1993), making it a seemingly favorable environment for the taxon. The area surrounding the Gulf Trough, however, may have been topographically high, creating a shallow-water barrier that prevented Basilosaurus dispersal to the deeper waters over the trough itself. The lack of Basilosaurus occurrences in the Priabonian of Florida likewise could be related to water depth, in which the topographically high Ocala Platform of central Florida (Fig. 6.2; Scott, Reference Scott, Buster and Holmes2011) created a zone of water too shallow for Basilosaurus to inhabit throughout much of the late Eocene.
Changes to the temporal distribution of Basilosaurus fossils in the Gulf Coastal Plain can be associated with regional marine transgressions and regressions. The first appearance of Basilosaurus in the North American fossil record is equivalent in time to the early Bartonian Cook Mountain transgression, which is associated with intertidal to inner neritic environments (Rainwater, Reference Rainwater1964). The middle Bartonian Yegua regression, which followed the Cook Mountain transgression, is dominated by continental and intertidal environments (Rainwater, Reference Rainwater1964) and coincides with an absence of Basilosaurus occurrences across the entire southeastern United States. Basilosaurus reappears in the fossil record at the Bartonian/Priabonian boundary, in conjunction with a slight deepening of the epicontinental sea (Huff, Reference Huff1970). The northernmost Basilosaurus lived at this time, when an inner to middle neritic environment spread across much of the Gulf Coastal Plain (Rainwater, Reference Rainwater1964; Huff, Reference Huff1970). Basilosaurus was most abundant during the middle Priabonian, in the middle to outer neritic zones of the deepening Jackson Sea (Huff, Reference Huff1970). The last appearance of Basilosaurus occurs near the end of the Jackson depositional episode/Jackson transgression (Rainwater, Reference Rainwater1964; Galloway, Reference Galloway2008), when portions of the Gulf Coastal Plain were well within the outer neritic zone (Huff, Reference Huff1970).
The limited number of Basilosaurus localities in the fossil record of the Atlantic Coastal Plain prevents a detailed assessment of any connection between Basilosaurus abundance and sea level in this region. In general, all Basilosaurus localities of the Atlantic Coastal Plain can be chronologically associated with the Jackson transgression of the Gulf Coastal Plain. The maximum landward extent of the Jackson sea was north of the Fall Line (Huddlestun, Reference Huddlestun1993), and all Basilosaurus occurrences are located south of this Jackson shoreline. GSM 1465 lies on the terminal Eocene shoreline, a low sea stand, but because this specimen can only be broadly dated as middle to late Priabonian it is equally possible that it lived in the deeper sea of the middle Priabonian.
Basilosaurus fossils provide evidence that a Mississippi embayment extended as far north as central Tennessee at the Bartonian/Priabonian Boundary (PBDB Collection 32902; Moore and Brown, Reference Moore and Brown1969). The presence of this embayment has been noted in the early Eocene, corresponding to the Upper Wilcox depositional episode (Galloway, Reference Galloway2008), in a middle (Bartonian/Lutetian boundary) and late (Priabonian) Eocene worldwide paleocoastline map (Smith et al., Reference Smith, Smith and Funnell1994), in the broadly defined “Jackson Time” of the late Eocene (Schuchert, Reference Schuchert1929), and in the Jackson Group outcrops reflecting the late Eocene seas (Reel, Reference Reel1972). To the best of our knowledge, this feature has not previously been shown extending as far north as in our interpretation. The depositional environments of the two northernmost specimens used to extend this feature are nearshore (estuary/bay and coastal indeterminate), so this embayment likely did not extend much farther north than depicted by our composite shoreline.
Summary and conclusions
The temporal range and geographic distribution of Basilosaurus in North America suggest that the maximum landward extent of the late Eocene epicontinental sea occurred between the late Bartonian and middle Priabonian. The decline in Basilosaurus localities in the late Priabonian suggests that a regional marine regression was occurring by this time. The most notable change to the late Eocene shoreline, based on Basilosaurus fossil occurrences, is the northward shift of the Mississippi embayment at the Bartonian/Priabonian boundary.
The environmental distribution of Basilosaurus across the southeastern coastal plain provides support for a previously proposed preference of the taxon for middle to outer neritic environments of the epicontinental seas. The absence of Basilosaurus fossils in the middle Bartonian, then, is likely related to a marine regression that resulted in a temporary lack of favorable environments for the taxon in the region. Likewise, the relatively few Basilosaurus occurrences in the Atlantic Coastal Plain suggests the environment there was less favorable for the taxon than it was over the Gulf Coastal Plain. The difference between regions is likely due to the presence of topographic highs, such as the Ocala Platform in Florida and, potentially, the area surrounding the Gulf Trough in Georgia, that resulted in a shallower epicontinental sea and barriers to dispersal in the east.
Acknowledgments
The authors would like to thank Rob Weller from the Fisheries Division of the Georgia Department of Natural Resources in Albany, Georgia, USA, for bringing this fossil to our attention, inviting us to excavate it, and assisting with site access and excavation. Bruce Maples, engineer with the City of Albany, Georgia, and Tommy Gregors, director of the Thronateeska Heritage Center, Albany, Georgia, were integral in providing us access to the site. Art Berger provided invaluable photography of the fossil and the site. We are grateful to David Campbell and Dana Ehret for identifying fauna found in association with the Flint River Basilosaurus. Thank you to Nicole Dzenowski and the paleontology volunteers at the Science Museum of Minnesota for preparation of the fossil material. We especially thank our excavation team of Georgia Southern University students, faculty, and friends: Zach Ansley, Mike D'Emic, Dustin Fackler, Shawna Felkel, Christina Gondry, Johanne Lewis, and Kelly Simpson. The Georgia Southern University College of Science and Mathematics provided funding to support the excavation. Excavation was conducted with the permission of the United States Army Corps of Engineers (Savannah District), project number SAS-2012-00529. Finally, we thank Stephen Godfrey and an anonymous reviewer for helpful and constructive comments on this manuscript.
Data availability statement
Data available from the Zenodo Digital Repository: https://doi.org/10.5281/zenodo.5167386






