



Introduction
The Cretaceous platform carbonates in the Geyik Dağı area represent a part of the so-called L'Axe Calcaire du Taurus (Ricou et al., Reference Ricou, Argyriadis and Marcoux1975), Anatolide-Tauride Carbonate Platform (ATCP) (Şengör and Yılmaz, Reference Şengör and Yılmaz1981), Taurus Carbonate Platform (Koçyiğit, Reference Koçyiğit1981), Menderes-Taurus Platform (Görür and Tüysüz, Reference Görür and Tüysüz2001), Menderes Carbonate Platform (Vlahović et al., Reference Vlahović, Tišljarb, Velić and Matičec2005). Most previous works on the Geyik Dağları range aim to explain its complex geological and tectonic evolution (e.g., Özgül, Reference Özgül1976, Reference Özgül1997; McPhee et al., Reference McPhee, van Hinsbergen, Maffione and Altıner2018), except for Özer and Kahrıman (Reference Özer and Kahrıman2019), in which Cenomanian canaliculate rudists were documented. In the Anamas-Akseki area, which is the continuation of the Geyik Dağları range to 65 km northwest, there are few studies on the detailed biostratigraphy and facies of the Cretaceous platform carbonate sequences (e.g., Martin, Reference Martin1969; Monod, Reference Monod1977; Altıner et al., Reference Altıner, Ömer Yılmaz, Özgül, Akçar, Bayazıtoğlu and Gaziulusoy1999; Solak et al., Reference Solak, Taslı and Koç2017, Reference Solak, Taslı, Özer and Koç2019; Solak, Reference Solak2019). This study forms part of a broader effort to contribute to the understanding of geologic evolution of the Geyik Dağı area during the Cretaceous.
The Albian Stage is little or poorly known in the ATCP due to emersion of some parts of the platform during the late Aptian (e.g., Farinacci and Köylüoğlu, Reference Farinacci and Köylüoğlu1982; Taslı and Solak, Reference Taslı and Solak2019; Solak et al., Reference Solak, Taslı and Koç2020), in addition to the lack of detailed micropaleontologic and biostratigraphic works. This study focuses on documenting and illustrating the Albian benthic foraminifera and on describing stratigraphically important ones. The first micropaleontological results allowed us to recognize a new species of benthic foraminifera from the Albian, Phenacophragma oezeri Solak and Taslı, Reference Solak and Taslı2020 from the peri-Mediterranean platforms.
Geologic setting
The Geyik Dağı area is located at the western part of the Central Taurides (Fig. 1.1) consisting of autochthonous and allochthonous rock assemblages (Özgül, Reference Özgül1976). In this area, autochthonous rocks, which were named as the Geyik Dağı Unit by Özgül (Reference Özgül1976), are overthrust by the Antalya Nappes (Lefèvre, Reference Lefèvre1967) or Antalya Complex (Robertson and Woodcock, Reference Robertson, Woodcock, Dixon and Robertson1984) to the south and by the Aladağ Unit (Özgül, Reference Özgül1976) to the north. Accretion of these nappes is thought to be the result of continental and oceanic subduction below an oceanic upper plate lithosphere (Şengör and Yılmaz, Reference Şengör and Yılmaz1981). The Geyik Dağı Unit includes carbonate and clastic sediments of Cambrian–Ordovician age, unconformably overlain by platform-type sequences of Middle Jurassic–Cenomanian age (Polat Limestone) and Maastrichtian–Paleocene age (Çataloluk Limestone), ending with Lutetian clastics (Özgül, Reference Özgül1997). The Maastrichtian to Lutetian cherty pelagic limestones (Kuşca Limestone) also occur in a separate tectonic slice of the Yıldızlı Dağ Unit (Özgül, Reference Özgül1997). On the other hand, McPhee et al. (Reference McPhee, van Hinsbergen, Maffione and Altıner2018) called the same rock assemblages the Geyikdağı nappe, which was deformed by a thin-skinned thrust fault imbricate system that affected the uppermost Mesozoic carbonates and by a deeper thrust duplex system that incorporates Ordovician and older basement rocks. During field work in the Geyik Dağı area, we have observed two carbonate successions of different paleoenvironmental settings separated by faults: one of them contains orbitolinid and rudistid bioclastic limestones of Cenomanian age (cf. upper part of the Polat Limestone of Özgül, Reference Özgül1997) and overlying hemipelagic to pelagic limestones of Santonian to Danian age (cf. Kuşca Limestone of Özgül, Reference Özgül1997), while the other consists entirely of an inner platform rudistid limestone succession of Albian to Late Cretaceous age (cf. Polat Limestone plus Çataloluk Limestone of Özgül, Reference Özgül1997), including a post-Cenomanian disconformity. The section studied belongs to the Hadim Unit (Özgül, Reference Özgül1997, p. 117, fig. 4), in which the Cretaceous is represented entirely by platform-type carbonates. It corresponds to the upper part of the Middle Jurassic–Cenomanian Polat Limestone of the Hadim Unit, which contains rudist-rich limestones. See Özgül (Reference Özgül1997, p. 118, fig. 5) for a generalized section of the Geyik Dağı Unit in the Hadim area.

Figure 1. Location and geologic maps of the study area. (1) Geographical subdivision of the Taurides (after Özgül, Reference Özgül1984); (2, 3) schematic geologic maps of the Geyik Dağları area showing the location of studied section (simplified after Özgül, Reference Özgül1984).
Materials and methods
The Geyik Dağları is a mountain range lying in the northwest-southeast direction on the Taşeli Plateau in the north of Alanya (Antalya), and accessible by car and hard-surface roads (one lane wide) only in dry weather during the summer. The highest mountain is the Geyik Dağı (2877 m). The Cretaceous platform-type carbonate succession (Fig. 1.2, 1.3) is exposed along a rocky ridge, including Göbekçal Hill (2391 m), located 10 km northwest of Geyik Dağı peak. This ridge is limited by faults and Pleistocene glacial sediments. The succession consists of beige- and cream-colored, thick- to very thick-bedded limestones that frequently contain whole and fragmented rudists, gastropods, and large bivalves. Laminae, fenestrae, and emersion/paleokarst breccia/karst infillings are common. Algal-foraminiferal packstone microfacies intercalated with ostracod-miliolid/wackestone/mudstone microfacies indicates peritidal environments in an inner platform setting. Although the succession is interrupted by step faults and karstic depressions, the section is continuous up to a major disconformable surface covering the Cenomanian strata. The section was logged from the lowermost exposed limestone layers (36°58'26.66"N, 32° 6'41.76"E). Micropaleontologic analyses of limestone samples were performed on 62 thin sections and 30 serial acetate-peels obtained from the 23 limestone samples. Determination of the stratigraphic position of the investigated succession that is shown in Figure 2 is based on the identification of benthic foraminifera and on using them as index fossils. The stratigraphic value of taxa identified is discussed in the “Discussion and biostratigraphic remarks” section.

Figure 2. Stratigraphic distribution of the Albian benthic foraminifera in the Göbekçal Hill section.
The higher taxonomic classification follows Pawlowski et al. (Reference Pawlowski, Holzmann and Tyszka2013). The lower taxonomic classification follows Kaminski (Reference Kaminski2014) for the agglutinating taxa and Loeblich and Tappan (Reference Loeblich and Tappan1988) for representatives of the Miliolida. The terminology used in the text is defined in Hottinger (Reference Hottinger2006). The section types passed parallel or perpendicular to the plane of biseriality in biserially arranged forms are variously termed: axial section (Hottinger, Reference Hottinger2006, fig. 6B), vertical section (BouDagher-Fadel, Reference BouDagher-Fadel2018, pl. 5.10), longitudinal section (Cherchi et al., Reference Cherchi, Schroeder and Ruberti2009, pl. 1; Cvetko Tešović et al., Reference Cvetko Tešović, Glumac and Bucković2011). We preferred to use the terms “longitudinal section perpendicular/parallel to the plane of biseriality” and “transverse section” for sections of biserial forms.
Repository and institutional abbreviation
The studied Albian thin-sections, which are labeled as A8–A30, are deposited in the collection of Paleontology at the General Geology Laboratory, Department of Geological Engineering, Mersin University, Turkey.
Systematic paleontology
Phylum Foraminifera d'Orbigny, Reference d'Orbigny1826
Class Globothalamea Pawlowski et al., Reference Pawlowski, Holzmann and Tyszka2013
Order Lituolida Lankester, Reference Lankester1885
Suborder Nezzazatina Kaminski, Reference Kaminski, Bubík and Kaminski2004
Superfamily Nezzazatoidea Hamaoui and Saint-Marc, Reference Hamaoui and Saint-Marc1970
Family Nezzazatidae Hamaoui and Saint-Marc, Reference Hamaoui and Saint-Marc1970
Subfamily Nezzazatinae Hamaoui and Saint-Marc, Reference Hamaoui and Saint-Marc1970
Genus Nezzazata Omara, Reference Omara1956
Type species
Nezzazata simplex Omara, Reference Omara1956, p. 889, pl. 102, figs. 7–13, text-fig. 6, western Sinai, Egypt.
Nezzazata isabellae Arnaud-Vanneau and Sliter, Reference Arnaud-Vanneau and Sliter1995
Figure 3.1–3.4
- Reference Altıner and Decrouez1982
Nezzazata sp. C; Altıner and Decrouez, pl. 4, fig. 26.
- Reference Arnaud-Vanneau and Sliter1995
Nezzazata isabellae Arnaud-Vanneau and Sliter, p. 552, text-fig. 7 (A–D), pl. 2, figs. 11–24.
- Reference Arnaud-Vanneau and Premoli Silva1995
Nezzazata isabellae; Arnaud-Vanneau and Premoli Silva, p. 206, pl. 2, figs. 1–3.
- Reference Mancinelli and Chiocchini2006
Nezzazata isabellae; Mancinelli and Chiocchini, p. 92, pl. 4, figs. 11–21.
- Reference Chiocchini, Pampaloni and Pichezzi2012
Nezzazata isabellae; Chiocchini et al., pl. 66, figs. 2, 3, 5–7, 9–12, 14.
- Reference Velić2019
Nezzazata isabellae; Taslı and Solak., fig. 10 (8).
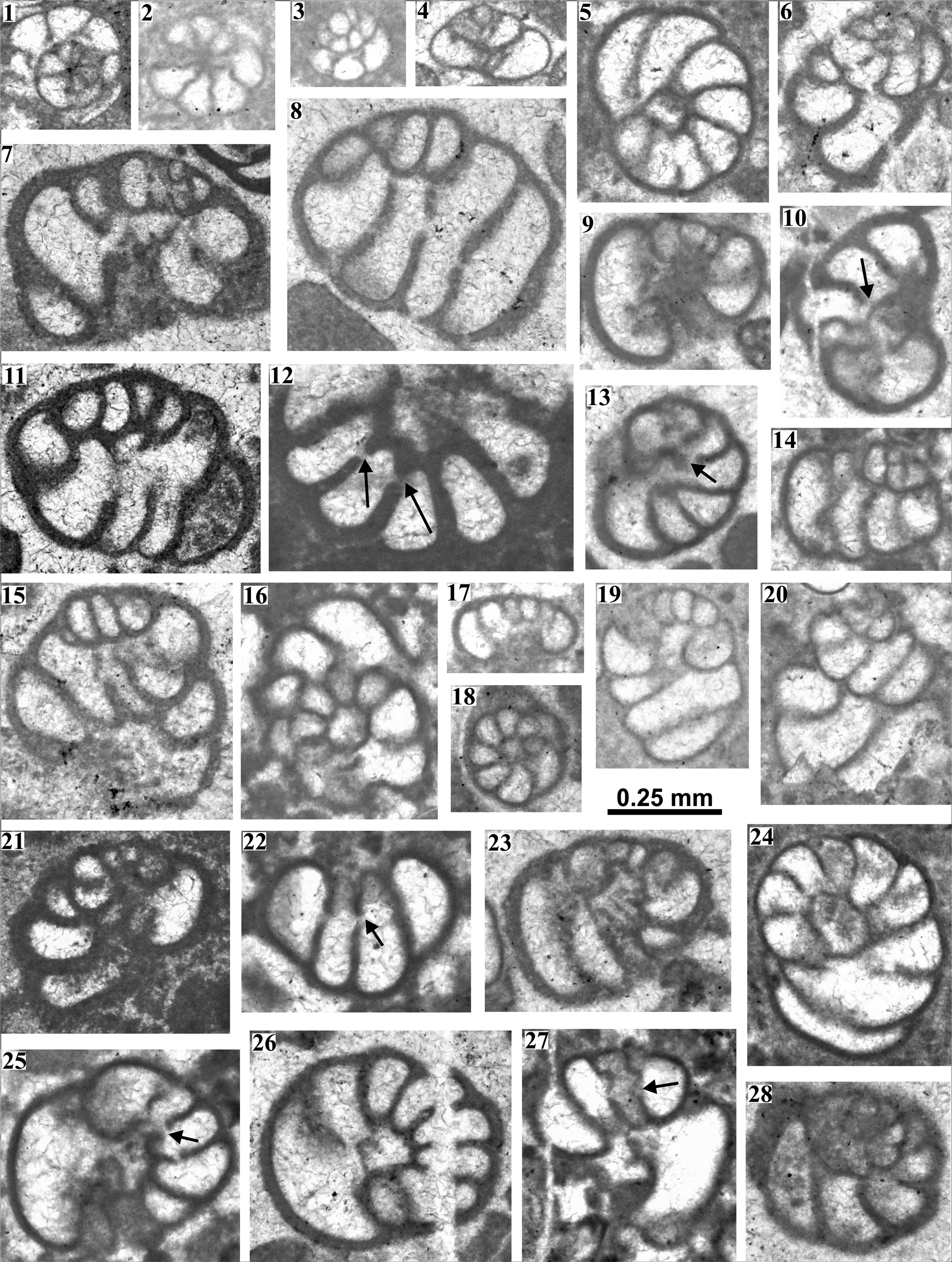
Figure 3. Nezzazata isabellae Arnaud-Vanneau and Sliter, Reference Arnaud-Vanneau and Sliter1995 (1–4) from the Albian platform limestones, Geyik Dağı area. (1, 2) Transverse sections (A12/1, A30/1); (3) oblique axial section (A14/1); (4) axial section (A26/1). Nezzazatinella sp. (5–28) from the Albian platform limestones, Geyik Dağı area. (5, 10, 12, 16, 18, 24, 26) Nearly transverse sections (A22/1, A9/1, A22/4, A12/1, A9/2, A8/1, A9/1); (7, 9, 17, 21, 23) nearly axial sections (A9/3, A9/2, A8/1, A27/9, A9/2); (6, 13, 19, 25, 28) oblique transverse sections (A12/1, A9/1, A8/1, A12/2, A9/1); (8, 11, 14, 15, 20, 22, 27) tangential sections (A9/1, A9/3, A12/1, A9/1, A8/1, A12/1, A9/1, A12/1); arrows indicate aperture.
Holotype
Axial section (USNM 483970) from the Allison Guyot in the Mid-Pacific Mountains (Arnaud-Vanneau and Sliter, Reference Arnaud-Vanneau and Sliter1995, pl. 2, fig. 11).
Remarks
This species is characterized by a small test with a rounded periphery (Arnaud-Vanneau and Sliter, Reference Arnaud-Vanneau and Sliter1995). It frequently occurs throughout the studied section and is especially abundant in the lower part. The Geyik Dağı specimens, represented by numerous specimens in various section types, have a height (h) of mostly 0.100–0.148 mm (rarely up to 0.162 mm), diameter (d) of 0.175–0.203 mm (rarely up to 0.243 mm), and mostly 2–3 (rare) whorls. Although there are a few larger individuals, they correspond to type specimens of Nezzazata isabellae described by Arnaud-Vanneau and Sliter (Reference Arnaud-Vanneau and Sliter1995) from the (late?) Aptian to early Albian.
Nezzazata isabellae represents one of the oldest species of the genus Nezzazata (Arnaud-Vanneau and Sliter, Reference Arnaud-Vanneau and Sliter1995) and differs from other species by its smaller size—minimum height of 0.2 mm, minimum diameter of 0.3 mm in 7 Nezzazata species of Smout (Reference Smout1956) and minimum diameter of 0.51 mm in Nezzazata simplex Omara (Reference Omara1956).
Genus Nezzazatinella Darmoian, Reference Darmoian1976
Type species
Nezzazatinella adhami Darmoian, Reference Darmoian1976, p. 525, pl. 1, figs. 1–7, Shat Al Arab Formation, Basrah, southeastern Iraq.
Nezzazatinella sp.
Figure 3.5–3.28
Remarks
This species differs from the other known species of Nezzazatinella in having a widely rounded periphery, fewer numbers of chambers in the last whorl, and small sizes (see Table 1). It resembles Dobrogelina? cartusiana Arnaud-Vanneau, Reference Arnaud-Vanneau1980 in its widely rounded periphery, but the aperture is a single slit (Fig. 3.10, 3.12, 3.13, 3.22, 3.25, 3.27), instead of apertural pores.
Table 1. Comparative table of Nezzazatinella sp. with the other Nezzazatinella species and Dobrogelina? cartusiana Arnaud-Vanneau, Reference Arnaud-Vanneau1980.

Order Loftusiida Kaminski and Mikhalevich in Kaminski, Reference Kaminski, Bubík and Kaminski2004
Suborder Loftusiina Kaminski and Mikhalevich in Kaminski, Reference Kaminski, Bubík and Kaminski2004
Family Spirocyclinidae Munier-Chalmas, Reference Munier-Chalmas1887
Subfamily Cyclammininae Marie, Reference Marie1941
Genus Reissella Hamaoui, Reference Hamaoui1963
Type species
Reissella ramonensis Hamaoui, Reference Hamaoui1963, p. 58, Upper Cretaceous (Cenomanian), Judea Limestone Group, Israel.
Reissella sp.
Figure 4.17–4.25
Description
The test is very small, peneropliform, planispirally enrolled, and involute, with one (Fig. 4.18) to two whorls (Fig. 4.22); later, it may tend to uncoil (Fig. 4.19, 4.21) with up to three uniserial chambers (Fig. 4.19). Proloculus spherical to subspherical, probably composed of proloculus and deuteroloculus (Fig. 4.18, 4.19). The last whorl contains 10–13 chambers. There is no significant change in chamber height, while chamber width increases gradually during ontogenesis. Chambers are subdivided by very short beams, aligned in successive chambers, producing quadrangular meshes in the marginal zone of chambers (Fig. 4.24). The wall is thin, simple, and microgranular, with no agglutinated particles. Aperture is cribrate with numerous pores scattered over the apertural face (Fig. 4.17, 4.23). Proloculus diameter 0.06–0.08 mm (rarely 0.10 mm), equatorial diameter 0.20–0.35 mm, axial thickness 0.20 mm, test height 0.30–0.40 mm.
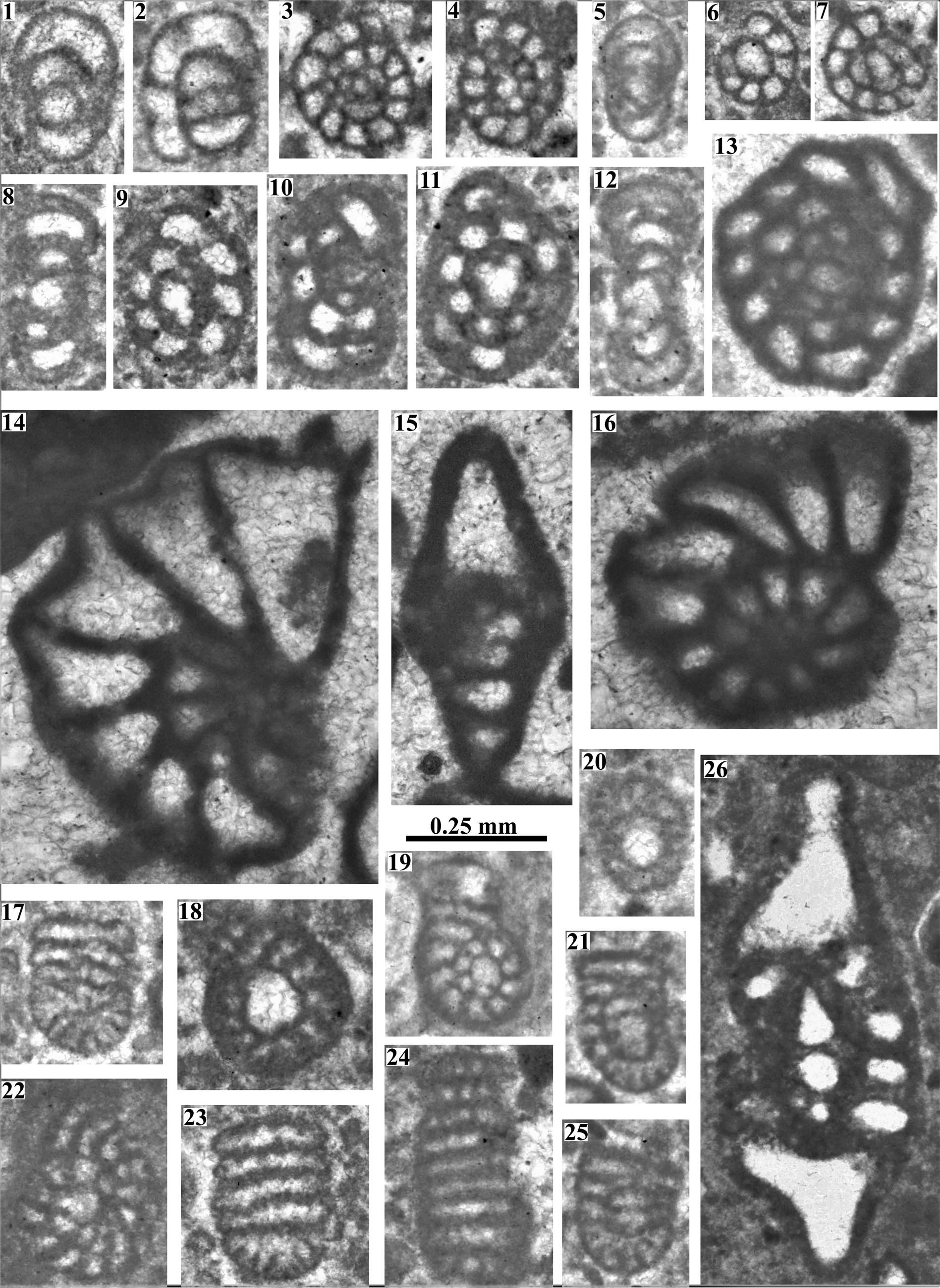
Figure 4. Benthic foraminifera from the Albian platform limestones, Geyik Dağı area. (1–7) Haplophragmoides globosus Lozo, Reference Lozo1944: (1, 5) Axial sections (A26/1, A26/1), (2) oblique subequatorial section (A26/1), (3, 4, 6, 7) equatorial sections (A12/1, A26/1, A12/1, A19/6). (8–13) Trochamminoides coronus Loeblich and Tappan, Reference Loeblich and Tappan1946: (8, 12) Axial sections (A22/1, A14/1), (9, 11, 13) equatorial sections (A26/1, A26/1, A19/6), (10) oblique subequatorial section (A14/1). (14, 15) Phenacophragma oezeri Solak and Taslı, Reference Solak and Taslı2020, subequatorial and subaxial sections (A19/1, A19/1). (16, 26) Mayncina bulgarica Laug et al., Reference Laug, Peybernès and Rey1980, equatorial and oblique axial sections (A19/1, A27/8). (17–25) Reissella sp., (17, 23–25) subaxial sections (A26/1, A26/1, A26/1, A26/1), (18–21) equatorial sections (A26/1, A26/1, A26/1, A27/6), (22) oblique equatorial section (A26/1).
Remarks
The type species, described from the Cenomanian of Israel, has a larger test and numerous short rafters in addition to elongate primary and short secondary beams. A slit-like aperture could not be observed in our specimens. This species seems to be a primitive representative of the genus and is probably new. It is very rarely found in numerous thin-sections of two samples (A26 and A27).
Suborder Ataxophragmiina Fursenko, Reference Fursenko1958
Family Cuneolinidae Saidova, Reference Saidova1981
Subfamily Cuneolininae Saidova, Reference Saidova1981
Genus Cuneolina d'Orbigny, Reference d'Orbigny1839
Type species
Cuneolina pavonia d’Orbigny, Reference d'Orbigny1846, p. 253, pl. 21, figs. 50–52, Upper Cretaceous, Charente, France.
Cuneolina parva Henson, Reference Henson1948
Figure 5.1–5.3
- Reference Henson1948
Cuneolina pavonia var. parva var. nov. Henson, p. 624, pl. 14, figs. 1–6, pl. 17, figs. 7–12, pl. 18, figs. 12–14.
- Reference Arnaud-Vanneau and Sliter1995
Cuneolina parva; Arnaud-Vanneau and Sliter, p. 554, pl. 4, figs. 6, 7, 9.
- Reference Cvetko Tešović, Glumac and Bucković2011
Cuneolina parva; Cvetko Tešović et al., fig. 12J, K.

Figure 5. Cuneolinid foraminifera from the Albian platform limestones, Geyik Dağı area. (1–3) Cuneolina parva Henson, Reference Henson1948: (1) tangential section oblique to the plane of biseriality (A9/4), (2) oblique transverse section (A9/1), (3) tangential section showing beams in embrionic apparatus (19/2). (4, 5) Cuneolina sliteri Arnaud-Vanneau and Premoli Silva, Reference Arnaud-Vanneau and Premoli Silva1995, longitudinal sections perpendicular to the plane of biseriality (A12/1, A14/1). (6, 7) Vercorsella arenata Arnaud-Vanneau, Reference Arnaud-Vanneau1980, tangential sections (A8/1, A19/1). (8–13) Vercorsella scarsellai (De Castro, Reference De Castro1963): (8) longitudinal section perpendicular to the plane of biseriality (A19/6), (9, 13) tangential sections (A19/1, A9/4), (10) longitudinal section of a specimen revealing that the plane of biseriality is twisted (A9/1), (11) transverse section (A19/2), (12) oblique transverse section (A9/1). (14–18) Akcaya minuta (Hofker, Reference Hofker1965): (14, 16) longitudinal sections perpendicular to the plane of biseriality (A27/6, A8/1), (15, 17) tangential sections (A9/3, A12/1), (18) transverse section (A9/3). (19–23) Akcaya auruncensis (Chiocchini and Di Napoli-Alliata, Reference Chiocchini and Di Napoli-Alliata1966): (19, 20) longitudinal sections perpendicular to the plane of biseriality (A12/1, A9/1), (21) sublongitudinal section perpendicular to the plane of biseriality (A19/2), (22, 23) transverse sections (A9/3, A19/3). (24) Novalesia? sp., longitudinal section perpendicular to the plane of biseriality (A9/1).
Holotype
Specimen (P.39115) from Egypt; Parker's Monument Section (Henson, Reference Henson1948, p. 624, not figured holotype).
Remarks
About 10 specimens that have random sections were examined. Some sections display proloculus covered by a subdivided deuteroloculus (e.g., Fig. 5.1, 5.3). Chambers have one or two rafters (Fig. 5.1, 5.2). The morphological characters and measured values of the Geyik Dağı specimens correspond to the type specimens of Cuneolina parva (maximum height of 1.35 mm in our specimens, versus 1.6 mm in Henson, Reference Henson1948). The biserial stage contains up to nine pairs of chambers. The apical angles of specimens in thin sections are not comparable.
Two large cuneolinids, Cuneolina pavonia and Cuneolina parva, are frequently reported species. Cuneolina pavonia differs from Cuneolina parva by its larger test and by having 2–3 rafters in the chambers. Cuneolina sliteri Arnaud-Vanneau and Premoli Silva, Reference Arnaud-Vanneau and Premoli Silva1995, which is the smallest species of Cuneolina, is distinguished by its smaller, non-flabelliform test and by lack of rafters in the first chambers.
Cuneolina sliteri Arnaud-Vanneau and Premoli Silva, Reference Arnaud-Vanneau and Premoli Silva1995
Figure 5.4–5.5
- Reference Arnaud-Vanneau and Premoli Silva1995
Cuneolina sliteri Arnaud-Vanneau and Premoli Silva, p. 207, pl. 3, figs. 1–9.
- Reference Mancinelli and Chiocchini2006
Cuneolina sliteri; Mancinelli and Chiocchini, p. 94, pl. 4, figs. 1–10.
- Reference Chiocchini, Pampaloni and Pichezzi2012
Cuneolina sliteri; Chiocchini et al., pl. 62, figs. 2–12.
Holotype
Longitudinal section in the plane of biseriality from Takuyo-Daisan Guyot, Sample 144-879A-5R-1 (Arnaud Vanneau and Premoli Silva, Reference Arnaud-Vanneau and Premoli Silva1995, pl. 3, fig. 1).
Remarks
More than 15 specimens in mostly longitudinal sections were examined. They have a height of 0.31–0.90 mm and a maximum basal diameter up to 0.23 mm. Embryonic apparatus made by a proloculus and a deuteroloculus subdivided by a few rafters and beams, followed by a biserial stage with up to mostly five or six pairs of chambers (only one specimen up to 10). The deuteroloculus displays mostly only beams in thin sections (Fig. 5.4, 5.5). These specimens are within the range of morphological characters and biometric values (maximum height of 1.15 mm, maximum diameter of 0.78 mm) of Cuneolina sliteri described by Arnaud-Vanneau and Premoli Silva (Reference Arnaud-Vanneau and Premoli Silva1995). The comparison with frequently reported other representatives of Cuneolina is made in remarks for Cuneolina parva.
Genus Vercorsella Arnaud-Vanneau, Reference Arnaud-Vanneau1980
Type species
Vercorsella arenata Arnaud-Vanneau, Reference Arnaud-Vanneau1980, p. 519, pl. 46, figs. 1, 2, pl. 71, figs., 1–7, Western Alps, France.
Remarks
In the generic diagnosis by Arnaud-Vanneau (Reference Arnaud-Vanneau1980) and Loeblich and Tappan (Reference Loeblich and Tappan1988), the aperture of Vercorsella is expressed as to be a basal slit and used as the main criterion to distinguish it from Cuneolina, which has a row of pores. Arnaud-Vanneau and Sliter (Reference Arnaud-Vanneau and Sliter1995) provided emendations of Vercorsella and Cuneolina and noted that the aperture of Vercorsella consists of a row of pores. Arnaud-Vanneau and Sliter (Reference Arnaud-Vanneau and Sliter1995) used a keriothecal test and subdivided deuteroloculus in Cuneolina to distinguish it from Vercorsella. We distinguish Cuneolina from Vercorsella by its rafters (Vercorsella has fewer well-developed rafters) in the last chambers (e.g., Loeblich and Tappan, Reference Loeblich and Tappan1988; Schlagintweit and Gawlick, Reference Schlagintweit and Gawlick2005), as well as a non-subdivided deuteroloculus.
Vercorsella arenata Arnaud-Vanneau, Reference Arnaud-Vanneau1980
Figure 5.6–5.7
- Reference Arnaud-Vanneau1980
Vercorsella arenata Arnaud-Vanneau, p. 519, pl. 71, figs. 1–3.
- Reference Chiocchini, Farinacci, Mancinelli, Molinari, Potetti and Mancinelli1994
Vercorsella arenata; Chiocchini et al., pl. 9, figs. 11, 12.
Holotype
Longitudinal section (ID 20 768) cutting the proloculus from the Urgonian platform, France (Arnaud-Vanneau, Reference Arnaud-Vanneau1980, pl. 71, fig. 1).
Remarks
Six specimens were examined in tangential sections. The biserial stage ranges from 5–7 pairs of chambers. Chambers are divided by 1–3, rarely 4 beams. Proloculus diameters are between 0.097 mm and 0.138 mm, which is similar to measurements (0.09–0.13 mm in Arnaud-Vanneau, Reference Arnaud-Vanneau1980) of the original description. The maximum height of the test (hmax.) is 0.73 mm in our specimens versus 0.725 mm in Arnaud-Vanneau (Reference Arnaud-Vanneau1980). Rafters are not observed in thin sections. The morphology and biometric parameters of these specimens correspond to the type specimens of Vercorsella arenata Arnaud-Vanneau, Reference Arnaud-Vanneau1980. The comparisons with other representatives of the genus Vercorsella are made in remarks for Vercorsella scarsellai (De Castro, Reference De Castro1963).
Vercorsella scarsellai (De Castro, Reference De Castro1963)
Figure 5.8–5.13
- Reference De Castro1963
Cuneolina scarsellai De Castro, p. 71, pl. 1, figs. 2–10, pl. 2, fig. 1.
- Reference Brönnimann and Conrad1968
Pseudotextulariella? scarsellai; Brönnimann and Conrad, p. 96, pl. 1, figs. 1–8, pl. 2, figs. 1–10.
- Reference Altıner and Decrouez1982
Pseudotextulariella? scarsellai; Altıner and Decrouez, pl. 3, figs. 15, 16.
- Reference Cvetko Tešović, Glumac and Bucković2011
Vercorsella scarsellai; Cvetko Tešović et al., fig. 11J.
- Reference Chiocchini, Pampaloni and Pichezzi2012
?Cuneolina scarsellai; Chiocchini et al., p. 68, pl. 46, figs. 1, 7.
Holotype
Specimen (767.33) in from the S. Maria la Face, Sarno, Italy (De Castro, Reference De Castro1963, pl. 1, fig. 1a–d).
Remarks
More than 25 specimens in mostly longitudinal, tangential, and a few transverse sections were examined. Test morphology and the absence of beams in the first chambers following the proloculus correspond to the types of Vercorsella scarsellai described from the Lower Cretaceous of Italy (De Castro, Reference De Castro1963). Beams are thicker (0.015–0.019 mm versus 0.006–0.013 mm in De Castro, Reference De Castro1963). The height of the test measured from longitudinal sections is up to 1.26 mm. While the last three or four pairs of chambers in most specimens are subdivided by beams (Fig. 5.10, 5.13), some specimens seem to have up to eight pairs of chambers subdivided by well-developed beams because tangential sections (e.g., Fig. 5.9) passed through both complete and incomplete (rudimentary) beams in the marginal zone of the test. In transverse sections passing through the adult stage, complete and incomplete beams are evident (Fig. 5.11, 5.12). The spiral initial stage is rarely observed (Fig. 5.10), followed by a biserial stage with up to 12 pairs of chambers. Some specimens (Fig. 5.10) may show a change from biserial to apparently uniserial chamber arrangement due to twisting of the test, as stated in Brönnimann and Conrad (Reference Brönnimann and Conrad1968, p. 97).
Vercorsella scarsellai differs from Vercorsella arenata Arnaud-Vanneau, Reference Arnaud-Vanneau1980 by its larger size (hmin. 0.7 mm in our specimens versus hmax. 0.72 mm in Arnaud-Vanneau, Reference Arnaud-Vanneau1980) and by having rare and incomplete beams. Vercorsella laurentii (Sartoni and Crescenti, Reference Sartoni and Crescenti1962) and Vercorsella camposaurii (Sartoni and Crescenti, Reference Sartoni and Crescenti1962) are distinguished from V. scarsellai by more flaring test shape and more regular beams. Velić and Gušić (Reference Velić and Gušić1973) stated that V. laurentii has very thick interchamber bands. Vercorsella tenuis (Velić and Gušić, Reference Velić and Gušić1973) can be distinguished easily from V. scarsellai by its very thin septa and beams. Vercorsella wintereri Arnaud-Vanneau and Sliter, Reference Arnaud-Vanneau and Sliter1995 and Vercorsella halleinensis Schlagintweit and Gawlick, Reference Schlagintweit and Gawlick2005 have smaller tests, with maximum heights are 0.30 mm and 0.72 mm, respectively.
Subfamily Sabaudiinae Brönnimann et al., Reference Brönnimann, Zaninetti and Whittaker1983
Genus Akcaya Özdikmen, Reference Özdikmen2009
Type species
Textulariella minuta (Hofker, Reference Hofker1965), p. 186, pl. 3, figs. 5, 6, pl. 4, figs. 1–9, Province of Santander, Puerto de Las Alisas, Spain.
Akcaya minuta (Hofker, Reference Hofker1965)
Figure 5.14–5.18
- Reference Hofker1965
Textulariella minuta Hofker, p. 186, pl. 3, figs. 5, 6, pl. 4, figs. 1–9.
- Reference Arnaud-Vanneau1980
Sabaudia minuta; Arnaud-Vanneau, p. 525, pl. 16, figs. 6–13.
- Reference Altıner and Decrouez1982
Sabaudia minuta; Altıner and Decrouez, pl. 3, figs. 3, 4.
- Reference Arnaud-Vanneau, Chiocchini, Schroeder and Neumann1985
Sabaudia minuta; Arnaud-Vanneau and Chiocchini, p. 29, pl. 10, figs. 1–14.
- Reference Chiocchini, Farinacci, Mancinelli, Molinari, Potetti and Mancinelli1994
Sabaudia minuta; Chiocchini et al., pl. 10, figs. 3, 4.
- Reference Özdikmen2009
Akcaya minuta; Özdikmen, p. 243.
- Reference Chiocchini, Pampaloni and Pichezzi2012
Sabaudia minuta; Chiocchini et al., pl. 38, figs. 2–6, 9, 10, 12.
- Reference Schlagintweit, Rosales and Najarro2016
Akcaya minuta; Schlagintweit et al., p. 124, fig. 6J.
Holotype
Longitudinal section oblique to the direction of the apertural slit (Nr. 115178) from Province of Santander, Puerto de Las Alisas, Spain (Hofker, Reference Hofker1965, pl. 3, fig. 5).
Remarks
The genus name Sabaudia Charollais and Brönnimann, Reference Charollais and Brönnimann1965 (type species: Textulariella minuta Hofker, Reference Hofker1965) preoccupied by Sabaudia Ghigi, Reference Ghigi1909 (type species: Sabaudia liguriae Ghigi, Reference Ghigi1909) was renamed as Akcaya by Özdikmen (Reference Özdikmen2009) and included in the classification of Kaminski (Reference Kaminski2014). A new name, Akcayinae, for the subfamily Sabaudiinae was proposed by Özdikmen (Reference Özdikmen2009) owing to the invalid type genus Sabaudia, however, the name Sabaudiinae was retained by Kaminski (Reference Kaminski2014).
About 15 specimens in longitudinal and transverse sections were examined. These specimens are similar to the types of Hofker (Reference Hofker1965) and Chiocchini et al. (Reference Chiocchini, Pampaloni and Pichezzi2012), with the exception of their thinner beams. The maximum height of the test measured from the longitudinal sections is 0.264 mm, and the maximum diameter measured from the circular/subcircular transverse sections is 0.281 mm, except for a few specimens (hmax = 0.328 mm). Juvenarium composed of a proloculus followed by two or rarely three globular chambers and surrounded by a layer of hyaline calcite is clear (Fig. 5.14, 5.15). Biserial arranged adult test in these specimens consists of mostly four or five pairs of chambers subdivided by beams.
Akcaya minuta differs from Akcaya capitata (Arnaud-Vanneau, Reference Arnaud-Vanneau1980) and Akcaya briacensis (Arnaud-Vanneau, Reference Arnaud-Vanneau1980) by the absence of rafters, as stated by Arnaud-Vanneau (Reference Arnaud-Vanneau1980, p. 525). It has a smaller test than Akcaya capitata and a less-flared cone test shape than Akcaya briacensis. Akcaya capitata also has a shape/section compressed perpendicularly to the plane of biseriality versus the nearly circular basal section of Akcaya minuta. Akcaya dinapolii (Chiocchini, Reference Chiocchini1984) from the upper Aptian of the Aurunci Mountains, Italy, is an infrequent form and has a less-pointed cone shape than Akcaya minuta. It also has constant quadrilocular embryos, whereas Akcaya minuta rarely has quadrilocular embryos (e.g., Arnaud-Vanneau and Chiocchini, Reference Arnaud-Vanneau, Chiocchini, Schroeder and Neumann1985; Schlagintweit et al., Reference Schlagintweit, Rosales and Najarro2016). Akcaya auruncensis (Chiocchini and Di Napoli-Alliata, Reference Chiocchini and Di Napoli-Alliata1966) is distinguished from Akcaya minuta by its more elongated and larger conical test and by the presence of rafters in the last chambers.
Akcaya auruncensis (Chiocchini and Di Napoli-Alliata, Reference Chiocchini and Di Napoli-Alliata1966)
Figure 5.19–5.23
- Reference Chiocchini and Di Napoli-Alliata1966
Textulariella auruncensis Chiocchini and Di Napoli-Alliata, pl. 4, figs. 1, 3, 5–7, pl. 5, figs. 3–6.
- Reference Chiocchini1984
Sabaudia auruncensis; Chiocchini, p. 16, text-fig. 2, pl. 1, figs. 1–6, 11, 12.
- Reference Chiocchini, Farinacci, Mancinelli, Molinari, Potetti and Mancinelli1994
Sabaudia auruncensis; Chiocchini et al., pl. 10, figs. 5, 6.
- Reference Chiocchini, Pampaloni and Pichezzi2012
Sabaudia auruncensis; Chiocchini et al., pl. 57, figs. 2, 3, 5–7.
Holotype
Longitudinal section (Chiocchini and Di Napoli-Alliata, Reference Chiocchini and Di Napoli-Alliata1966, pl. 4, fig. 1) and schematic reconstruction of the longitudinal section of the holotype, Aurunci Mountains, Southern Latium, Italy (Chiocchini, Reference Chiocchini1984, text-fig. 2).
Remarks
Akcaya auruncensis was described as Textulariella auruncensis by Chiocchini and Di Napoli-Alliata (Reference Chiocchini and Di Napoli-Alliata1966). Chiocchini (Reference Chiocchini1984) then revised the description of Akcaya auruncensis after it was regarded as synonymous with Akcaya minuta by some authors (e.g., Arnaud-Vanneau, Reference Arnaud-Vanneau1980). The measurements were made from ~10 specimens in longitudinal and transverse sections. These specimens exactly correspond to the Form A specimens of Chiocchini (Reference Chiocchini1984) in test diameters (height of 0.625–0.486 mm, from the subcircular/ovoid transverse sections and basal diameter of 0.468–0.421 mm). The rafters, which are rarely found in the last chambers of Chiocchini's (Reference Chiocchini1984) specimens, are more numerous in our specimens and observed in the last four or five pairs of chambers (Fig. 5.19–5.21). The juvenarium appears to be made up of a proloculus and two postembryonic chambers in some longitudinal sections (Fig. 5.20). The first larger postembryonic chamber may have been formed as a result of the melting of the wall between the two postembryonic chambers. Biserial arranged adult test consists of mostly eight or nine pairs of chambers. Comparison with other representatives of genus Akcaya is made in remarks for Akcaya minuta.
Suborder Orbitolinina Kaminski, Reference Kaminski, Bubík and Kaminski2004
Superfamily Orbitolinoidea Martin, Reference Martin1890
Family Orbitolinidae Martin, Reference Martin1890
Subfamily Dictyoconinae Moullade, Reference Moullade1965
Genus Cribellopsis Arnaud-Vanneau, Reference Arnaud-Vanneau1980
Type species
Orbitolinopsis? neoelongata Cherchi and Schroeder, Reference Cherchi and Schroeder1978, p. 162, Lower Cretaceous, France.
Cribellopsis moulladei (Saint-Marc, Reference Saint-Marc1974)
Figure 6.1–6.4
- Reference Saint-Marc1974
Simplorbitolina moulladei Saint-Marc, p. 225, pl. 2, figs. 3–8.
- Reference Chiocchini, Pampaloni and Pichezzi2012
Cribellopsis arnaudae; Chiocchini et al., pl. 69, figs. 2–10.
- Reference Schlagintweit2020
Cribellopsis moulladei; Schlagintweit, p. 37, fig. 1a–s.

Figure 6. Orbitolinid foraminifera from the Albian platform limestones, Geyik Dağı area. (1–4) Cribellopsis moulladei (Saint-Marc, Reference Saint-Marc1974): (1, 2) transverse sections (A9/1, A9/3), (3) subaxial section (A9/2), (4) oblique subaxial section (A9/4). (5–9) Paracoskinolina cf. P. sunnilandensis (Maync, Reference Maync1955): (5, 7, 8) subaxial sections (A9/1, A9/1, A9/3), (6) oblique transverse section (A9/2), (9) oblique subaxial section (A9/1). (10–16) Mesorbitolina gr. M. texana (Roemer, Reference Roemer1849): (10) axial section of a juvenile specimen in the embryonic stage (A19/5), (11) oblique subaxial section (A19/8), (12, 15, 16) axial sections (A19/6, A19/12, A22/1), (13) transverse section passing through the megalospheric embryo (A19/8), (14) axial section of a microspheric specimen (A19/1). Bar scale except for 14.
Holotype
Thin section view of the specimen from Albian, Dlebta Chenan Aaïr, Ghâzir, Lebanon (Saint-Marc, Reference Saint-Marc1974, pl. 2, fig. 5).
Remarks
The Geyik Dağı specimens (height ~0.73 mm, basal diameter 0.53–0.58 mm, adult chamber height 0.04–0.05 mm) are slightly larger than the Simplorbitolina moulladei specimens of Saint-Marc (Reference Saint-Marc1974), but they are within the range of morphological variation of Cribellopsis moulladei revised by Schlagintweit (Reference Schlagintweit2020) in test diameters. Triangular or inverted cone-shaped pillars in the central zone alternate in successive chambers (Fig. 6.3) and form an irregular and coarse central network (Fig. 6.1, 6.2). A specimen with rafter-like structures (Fig. 6.3) is tentatively included in this species.
Subfamily Dictyorbitolininae Schroeder in Schroeder et al., Reference Schroeder, Clavel and Charollais1990
Genus Paracoskinolina Moullade, Reference Moullade1965
Type species
Coskinolina sunnilandensis Maync, Reference Maync1955, p. 106, pl. 16, figs. 1, 2, 5–7, pl. 17, figs. 1–9, 12, southern Florida, USA.
Paracoskinolina cf. P. sunnilandensis (Maync, Reference Maync1955)
Figure 6.5–6.9
- ?Reference Maync1955
Coskinolina sunnilandensis Maync, p. 106, pl. 16, figs. 1, 2, 5–7, pl. 17, figs. 1–9, 12.
- Reference Arnaud-Vanneau and Premoli Silva1995
Paracoskinolina sp. cf. P. sunnilandensis; Arnaud-Vanneau and Premoli Silva, p. 208, pl. 5, figs. 5, 6.
Holotype
External view of the specimen from Humble Oil and Refining Company's No. 16 Gulf Coast Realties, core No. 23, Florida, USA (Maync, Reference Maync1955, pl. 16, fig. 1).
Remarks
This species is represented by subaxial and transverse sections of 10 specimens in only one sample (A9). They have a height of 0.35–0.67 mm and a basal diameter of 0.40–0.67 mm. The ratio of test height/diameter (h/d) is 1/1 (three specimens) or 1.5/1 (one specimen). Chamber height in the adult stage is 0.05–0.06 mm. Paracoskinolina sunnilandensis, originally described from the Albian of Florida and Venezuela, is the type of the genus. The Geyik Dağı specimens have a comparably reduced central zone and the boundary between central and marginal zones is not pronounced as in typical specimens of Paracoskinolina sunnilandensis. It is also difficult to distinguish the initial spire (Fig. 6.5), as Maync (1955, p. 107) stated.
Previously, the species was found in the northern margin of the Tethys and central America (Arnaud-Vanneau and Premoli Silva, Reference Arnaud-Vanneau and Premoli Silva1995), later in the Mesorbitolina subconcava (Leymerie, Reference Leymerie1878) taxon-range zone and “Valdanchella” dercourti Decrouez and Moullade, Reference Decrouez and Moullade1974 partial-range zone assigned to the upper-middle Albian of the Adriatic Carbonate Platform (Velić, Reference Velić2007). This species is associated with Cribellopsis moulladei, Pseudonummoloculina aurigerica Calvez, Reference Calvez1988, Vercorsella scarsellai, Akcaya minuta, and Glomospira urgoniana Arnaud-Vanneau, Reference Arnaud-Vanneau1980. This is the first record of the species from the Taurides.
Paracoskinolina fleuryi Decrouez and Moullade, Reference Decrouez and Moullade1974 was described from the upper Albian of the Gavrovo-Tripolitza Platform, Greece, and is differentiated from P. sunnilandensis (Maync, Reference Maync1955) by its smaller size (height of 0.4–0.5 mm) and shape of central pillars (Decrouez and Moullade, Reference Decrouez and Moullade1974). Later, it was recorded in the upper Albian limestones of the Adriatic Carbonate Platform (Husinec and Sokač, Reference Husinec and Sokač2006; Velić, Reference Velić2007; Husinec et al., Reference Husinec, Velić and Sokač2009). The central zone of our specimens is divided by “hemipillars” aligned in successive chambers, which is one of the diagnostic characteristics of the genus Paracoskinolina (Granier et al., Reference Granier, Clavel, Moullade, Busnardo, Charollais, Tronchetti and Desjacques2013), instead of alternated triangular pillars (Decrouez and Moullade, Reference Decrouez and Moullade1974, pl. 4, figs. 5–10).
Subfamily Orbitolininae Martin, Reference Martin1890
Genus Mesorbitolina Schroeder, Reference Schroeder1962
Type species
Orbitulites texanus Roemer, Reference Roemer1849, p. 392, not figured, Cretaceous, Texas, USA.
Mesorbitolina gr. M. texana (Roemer, Reference Roemer1849)
Figure 6.10–6.16
- Reference Roemer1849
Orbitulites texanus Roemer, p. 392, not figured.
- 1852
Orbitulites texanus; Roemer, p. 86, pl. 10, fig. 7a–d.
- Reference Schroeder1964
Orbitolina (Mesorbitolina) texana texana; Schroeder, p. 471; text-fig. 4b.
- Reference Moullade, Peybernès, Rey and Saint-Marc1985
Orbitolina (Mesorbitolina) texana; Moullade et al., pl. 1, fig. 9.
- Reference Schroeder, Schroeder and Neumann1985
Orbitolina (Mesorbitolina) texana; Schroeder, p. 77, pl. 36, figs. 1–13.
- Reference Schroeder, Buchem, Cherchi, Baghbani, Vincent, Immenhauser, Granier, van Buchem, Al-Husseini, Maurer and Droste2010
Mesorbitolina texana; Schroeder et al., fig. 8(5).
- Reference Chiocchini, Pampaloni and Pichezzi2012
Orbitolina (Mesorbitolina) texana; Chiocchini et al., pl. 59, figs. 2–6.
Holotype
Specimen(s) not designated as a holotype by Roemer (Reference Roemer1849, Reference Roemer1852) (p. 392) from Texas, USA. Several specimens were illustrated in Roemer (Reference Roemer1852) (pl. 10, fig. 7a–d).
Remarks
According to measurements from nearly axial sections of eight specimens, excluding juvenile ones, height and basal diameter of the test range between 0.75–2 mm and 1.70–4.55 mm, respectively. The embryonic apparatus is composed of an oval proloculus (0.12–0.20 mm, N = 6) capped by a deuteroloculus (0.4–0.5 mm) subdivided by beams and rafters and an equally developed subembryonic zone (Fig. 6.10–6.12). Despite their smaller size corresponding to Mesorbitolina texana, some of our specimens (Fig. 6.16) are compatible with Mesorbitolina subconcava described by Schroeder (Reference Schroeder, Schroeder and Neumann1985) in size and shape of embryonic apparatus. Therefore, we have identified the Mesorbitolina association as a group.
Order Textulariida Delage and Hérouard, Reference Delage and Hérouard1896
Suborder Textulariina Delage and Hérouard, Reference Delage and Hérouard1896
Superfamily Chrysalidinoidea Neagu, Reference Neagu1968
Family Chrysalidinidae Neagu, Reference Neagu1968
Genus Protochrysalidina Luperto-Sinni, Reference Luperto-Sinni1999
Type species
Protochrysalidina elongata Luperto-Sinni, Reference Luperto-Sinni1999, p. 251, pl. 1, 2, Puglia, Italy.
Remarks
This genus was not included in the classification of Loeblich and Tappan (Reference Loeblich and Tappan1988), Mikhalevich (Reference Mikhalevich2004), or Kaminski (Reference Kaminski2014). Protochrysalidina was assigned to Family Chrysalidinidae Neagu, Reference Neagu1968 by Luperto-Sinni (Reference Luperto-Sinni1999).
Protochrysalidina elongata Luperto-Sinni, Reference Luperto-Sinni1999
Figure 7.1–7.10
- Reference Luperto-Sinni1999
Protochrysalidina elongata Luperto-Sinni, p. 251, pl. 1, 2.
- Reference Megza, Cvetko Tešović, Bajraktarević and Bucković2007
Protochrysalidina elongata; Megza et al., p. 143, fig. 5 (E, F).
- Reference Taslı and Solak2019
Protochrysalidina elongata; Taslı and Solak, p. 203, fig. 10 (1–3).

Figure 7. Benthic foraminifera from the Albian platform limestones, Geyik Dağı area. (1–10) Protochrysalidina elongata Luperto-Sinni, Reference Luperto-Sinni1999: (1, 2, 5, 8, 9) subaxial sections (A12/2, A19/1, A19/1, A19/1, A9/1), (3, 4) axial sections (A19/2, A19/2), (6, 7, 10) transverse sections (A19/3, A19/1, A19/4). (11–14) Glomospira urgoniana Arnaud-Vanneau, Reference Arnaud-Vanneau1980, transverse sections (A8/1, A12/1, A8/1, A12/1). Bar scale except for (11–14).
Holotype
Subaxial section (GT3) from the Casino Chieco well, Puglia, Italy (Luperto-Sinni, Reference Luperto-Sinni1999, pl. 1, fig.1).
Remarks
Various oriented sections of >50 specimens were examined. Based on measurements from axial and subaxial sections of 27 specimens, they have a height of 1.0–2.91 mm and a basal diameter of 0.70–2.0 mm. The ratio of test height/diameter (h/d) is 1–2.09. The population of Protochrysalidina elongata composed of macrospheric specimens is characterized by low-conical forms (h/d = 1–1.16, eight specimens, and slightly above 1.27–1.45, six specimens) as well as the typical elongate conical forms (h/d = 1.5–2.09, 13 specimens). The pillars (forming endoskeleton) used as a distinguishing criterion do not exist in the early stage (at least in the first whorl). Incomplete pillars develop in the later stage, and complete pillars appear rarely in the last stage. The incomplete pillar development can be seen in earlier whorls (e.g., in the second whorl, Fig. 7.5, 7.9) in low-conical specimens compared to typical elongate-conical forms. The canaliculate (or pseudokeriothecal) wall microstructure is not evident. Aperture consisting of an interiomarginal slit in the juvenile stage, which is covered by a cribrate apertural plate in the later stages (also called ‘trematophore’), is distinct (Fig. 7.1, 7.5–7.7, 7.10). Proloculus spherical, followed by three chambers in a spiral coil (Fig. 7.4). We rarely observed in some sections that the triserial test then becomes biserial in the late ontogenic stage (Fig. 7.7).
These specimens are comparable to specimens of Protochrysalidina elongata illustrated by Luperto-Sinni (Reference Luperto-Sinni1999) from the Albian–lower Cenomanian of southern Italy and to specimens illustrated by Taslı and Solak (Reference Taslı and Solak2019) from the late Albian of the Bey Dağları, Turkey. We checked the accuracy of the dimensions (given in mm) by measuring the illustrations in Luperto-Sinni (Reference Luperto-Sinni1999) and noticed that the dimensions in centimeters are accidentally given in millimeters, as well as small deviations in the measurements of the shell diameter. The typical and low-conical specimens are similar in morphology and size (even if somewhat larger, hmax. = 2.91 mm versus 2.6 mm and 2.3 mm, respectively) to specimens of Luperto-Sinni (Reference Luperto-Sinni1999) (h/d = 1.5–2) and Taslı and Solak (Reference Taslı and Solak2019) (h/d = 1.6–2.25 of typical elongate-conical and 1–1.3 of low-conical forms). A specimen in Taslı and Solak (Reference Taslı and Solak2019) (fig. 10/1) is interpreted as a microspheric form (height 2.3 mm, diameter 1.96 mm). Banner et al. (Reference Banner, Simmons and Whittaker1991) described some specimens with complete and incomplete pillars from the Albian–Cenomanian of Iraq and Oman as “morphologically intermediate forms between Praechrysalidina infracretacea Luperto-Sinni, Reference Luperto-Sinni1979a and Chrysalidina gradata d'Orbigny, Reference d'Orbigny1839” (Banner et al., Reference Banner, Simmons and Whittaker1991, p. 110, 112, figs. 9, 12, 16, high-conical) and as “morphologically intermediate forms between Praechrysalidina infracretacea and Dukhania conica Henson, Reference Henson1948” (Banner et al, Reference Banner, Simmons and Whittaker1991, p. 110, 112, figs. 13, 14, low-conical), which are similar to the Geyik Dağı specimens.
Protochrysalidina elongata is distinguished from Praechrysalidina infracretacea, which can be found in a similar stratigraphic position (Albian), by its incomplete pillars in the adult stage and complete pillars in the final stage. Chrysalidina gradata, which is well known from the Cenomanian (e.g., De Castro, Reference De Castro1981, Reference De Castro, Schroeder and Neumann1985; Banner et al., Reference Banner, Simmons and Whittaker1991) differs by its stratigraphical position, its developed endoskeleton, and its higher and larger test. De Castro (Reference De Castro1981) stated that the height and diameter of Chrysalidina gradata specimens from Ile Madame (France) are between 2.4 and 4.5 mm; a limited number of specimens exceed 3.8 mm and 1.55–2.35 mm, respectively. De Castro (Reference De Castro1981) also noted that a specimen figured by Neumann (Reference Neumann1967) from Audignon (France) was 5 mm in height and the type specimen of Cushman (Reference Cushman1937) was 7 mm in height and 3 mm in diameter. The biseriality observed in sections of some specimens resembles Dukhania conica Henson, Reference Henson1948, but it has a more inflated and low-conical test (not elongate and high-conical), and better-developed pillars. Dukhania arabica Henson, Reference Henson1948 differs from Protochrysalidina elongata by its labyrinthic area.
Class Tubothalamea Pawlowski et al., Reference Pawlowski, Holzmann and Tyszka2013
Order Miliolida Delage and Hérouard, Reference Delage and Hérouard1896
Suborder Miliolina Delage and Hérouard, Reference Delage and Hérouard1896
Superfamily Milioloidea Ehrenberg, Reference Ehrenberg1839
Family Hauerinidae Schwager, Reference Schwager1876
Genus Pseudonummoloculina Calvez, Reference Calvez1988
Type species
Pseudonummoloculina aurigerica Calvez, Reference Calvez1988, p. 393, pl. 1, figs. 1–18, central Pyrenees, France.
Remarks
Loeblich and Tappan (Reference Loeblich and Tappan1988) included Pseudonummoloculina in foraminiferal genera of uncertain status.
Pseudonummoloculina aurigerica Calvez, Reference Calvez1988
Figure 8.1–8.6
- Reference Calvez1988
Pseudonummoloculina aurigerica Calvez, p. 393, pl. 1, figs. 1–18.
- Reference Mancinelli and Chiocchini2006
Pseudonummoloculina aurigerica; Mancinelli and Chiocchini, p. 100, pl. 6, figs. 8–15.
- Reference Chiocchini, Pampaloni and Pichezzi2012
Pseudonummoloculina aurigerica; Chiocchini et al., pl. 68, figs. 2–9.

Figure 8. Miliolid foraminifera from the Albian platform limestones, Geyik Dağı area. (1–6) Pseudonummoloculina aurigerica Calvez, Reference Calvez1988: (1, 2) equatorial sections (A9/1, A9/1), (3) oblique subaxial section (A9/1), (4, 5) oblique subequatorial sections (A9/1, A9/1), (6) axial section (A9/2). (7, 8) Pseudonummoloculina heimi (Bonet, Reference Bonet1956), equatorial and axial sections (A9/2, A9/1). (9–12) Paleocornuloculina sp.: (9) equatorial section showing the early cornuspirine stage (A30/1), (10, 11) subaxial sections (A30/1, A30/1), (12) tangential section oblique to the equatorial plane showing long and narrow chambers in the adult stage (A30/1). (13, 14) Peneroplis sp., subequatorial sections (A19/13, A27/1). (15, 16) Scandonea cf. S. pumila Saint-Marc, Reference Saint-Marc1974, equatorial sections (A9/3, A16/1). (17, 18) Cornuspiridae (cf. Fischerina? carinata), axial sections (A14/1, A8/1). (19) Spiroloculina sp., axial sections (A30/1). (20) Miliolidae, section perpendicular to the apertural axis (A9/3). (21, 22) Vidalina sp.: (21) axial section (A16/1), (22) equatorial section (A16/2).
Holotype
Oblique subaxial section (CLP 3) from the Cluse de Pereille, Pech de Foix, Central Pyrenees, France (Calvez, Reference Calvez1988, pl. l, fig. 5).
Remarks
The genus Pseudonummoloculina was distinguished from the genus Nummoloculina Steinmann, Reference Steinmann1881, with the type species N. contraria (d’Orbigny, Reference d'Orbigny1846) from the Miocene of the Vienna Basin, by a slit-like aperture bordered by a series of notches (Calvez, Reference Calvez1988). Based on this structural difference between the two genera, the widespread Cretaceous species Nummoloculina heimi Bonet, Reference Bonet1956, with a clear notched aperture (De Castro, Reference De Castro1987; Hottinger et al., Reference Hottinger, Drobne and Caus1989), was transferred to Pseodonummoloculina. Piuz and Vicedo (Reference Piuz and Vicedo2020) reviewed the other Cretaceous species of “nummoloculinas.” We could not observe the notched aperture due to the lack of available sections, however our specimens correspond to those of the type species (Calvez, Reference Calvez1988) in terms of the diameters and shape of the test, and the disposition and number of chambers per whorl.
Discussion and biostratigraphic remarks
In addition to the foraminiferal taxa discussed above (Figs. 3–8), the Albian platform limestones of the Geyik Dağı area contain Haplophragmoides globosus Lozo, Reference Lozo1944 (Fig. 4.1–4.7); Trochamminoides coronus Loeblich and Tappan, Reference Loeblich and Tappan1946, (Fig. 4.8–4.13); Phenacophragma oezeri Solak and Taslı, Reference Solak and Taslı2020 (Fig. 4.14, 4.15); Mayncina bulgarica Laug, Peybernès, and Rey, Reference Laug, Peybernès and Rey1980 (Fig. 4.16, 4.26); Reissella sp. (Fig. 4.17–4.25); Novalesia? sp. (Fig. 5.24); Glomospira urgoniana (Fig. 7.11–7.14); Pseudonummoloculina aurigerica (Fig. 8.1–8.6); Pseudonummoloculina heimi (Bonet, Reference Bonet1956) (Fig. 8.7, 8.8); Paleocornuloculina sp. (Fig. 8.9–8.12); Peneroplis sp. (Fig. 8.13, 8.14); Scandonea cf. S. pumila Saint-Marc, Reference Saint-Marc1974 (Fig. 8.15, 8.16); Cornuspiridae (cf. Fischerina? carinata Peybernès, Reference Peybernès1984) (Fig. 8.17, 8.18); Spiroloculina sp. (Fig. 8.19); Miliolidae (Fig. 8.20); Vidalina sp. (Fig. 8.21, 8.22); Nezzazata simplex Omara, Reference Omara1956; Dasycladalean algae (Fig. 9.1–9.6), including Triploporella cf. T. marsicana (Praturlon, Reference Praturlon1964) (Fig. 9.1, 9.2); Thaumatoporella sp. (Fig. 9.7); and Gahkumella huberi Zaninetti, Reference Zaninetti1978 (=Cretacicladus minervini Luperto-Sinni, Reference Luperto-Sinni1979b; e.g. Schlagintweit et al., Reference Schlagintweit, Kolodziej and Qorri2015) (Fig. 9.8). As seen in Figure 10, most of the benthic foraminifera identified range down into the Barremian–Aptian. Nevertheless, there is a consensus that Cuneolina parva and Pseudonummoloculina heimi do not exist at levels older than the Albian.

Figure 9. (1–6, 8) Dasycladalean algae and (7) incertae sedis from the Albian platform limestones, Geyik Dağı area. (1, 2) Triploporella cf. T. marsicana (Praturlon, Reference Praturlon1964) in Chiocchini et al. (Reference Chiocchini, Pampaloni and Pichezzi2012) (A19/3, A19/2), (3–6) (A19/6, A19/y5, A19/1, A19/y1), (8) Gahkumella huberi Zaninetti, Reference Zaninetti1978 (=Cretacicladus minervini Luperto-Sinni, Reference Luperto-Sinni1979b) (A31); (7) incertae sedis Thaumatoporella sp. (A8/1).

Figure 10. Stratigraphic distribution of some Early Cretaceous benthic foraminifera examined in this paper, from the literature. References: (1) Velić, Reference Velić2007; (2) Husinec et al., Reference Husinec, Velić and Sokač2009; (3) Chiocchini et al., Reference Chiocchini, Pampaloni and Pichezzi2012; (4) Calvez, Reference Calvez1988; (5) Conkin and Conkin, Reference Conkin and Conkin1958; (6) Hottinger et al., Reference Hottinger, Drobne and Caus1989; (7) Mancinelli and Chiocchini, Reference Mancinelli and Chiocchini2006; (8) Arnaud-Vanneau and Sliter, Reference Arnaud-Vanneau and Sliter1995; (9) Arnaud-Vanneau and Premoli Silva, Reference Arnaud-Vanneau and Premoli Silva1995; (10) Cvetko Tešović et al., Reference Cvetko Tešović, Glumac and Bucković2011; (11) Taslı and Solak, Reference Taslı and Solak2019; (12) Schroeder et al., Reference Schroeder, Buchem, Cherchi, Baghbani, Vincent, Immenhauser, Granier, van Buchem, Al-Husseini, Maurer and Droste2010; (13) BouDagher-Fadel et al., Reference BouDagher-Fadel, Hu, Price, Sun, Wang and An2017; (14) Arnaud-Vanneau, Reference Arnaud-Vanneau1980; (15) Maync, Reference Maync1955; (16) Saint-Marc, Reference Saint-Marc1974; (17) Schlagintweit, Reference Schlagintweit2020; (18) Chiocchini, Reference Chiocchini1984; (19) Hofker, Reference Hofker1965; (20) Henson, Reference Henson1948; (21) Chiocchini et al., Reference Chiocchini, Mancinelli and Romano1984. The geologic time duration in Myr is after Walker et al. (Reference Walker, Geissman, Bowring and Babcock2018).
Pseudonummoloculina aurigerica was first described from the Albian of the French and Spanish Pyrenees (Calvez, Reference Calvez1988). It was found in lower Albian of the Colle Santa Lucia succession (Mancinelli and Chiocchini, Reference Mancinelli and Chiocchini2006), and in Albian (to lower Cenomanian) strata of the Monte Cairo area, Central Italy (Chiocchini et al., Reference Chiocchini, Pampaloni and Pichezzi2012). It appears to be restricted to Albian, except for the Adriatic Carbonate Platform (Fig. 10) and the Southern Tibetan Plateau (BouDagher-Fadel et al., Reference BouDagher-Fadel, Hu, Price, Sun, Wang and An2017).
Protochrysalidina elongata was first described from the Albian–lower Cenomanian of southern Italy (Luperto-Sinni, Reference Luperto-Sinni1999), and has been used as an index taxon for the latest Albian in the Adriatic Carbonate Platform (e.g., Velić, Reference Velić2007; Husinec et al., Reference Husinec, Velić and Sokač2009) and in the Bey Dağları Platform (Taslı and Solak, Reference Taslı and Solak2019; Solak et al., Reference Solak, Taslı and Koç2020). Nevertheless, it is not a widely accepted taxon, probably due to difficulties in distinction among the chrysalidinid taxa.
There are a few studies on benthic foraminiferal biostratigraphy of the Lower–middle Cretaceous platform successions in the Tauride Carbonate Platform. Altıner et al. (Reference Altıner, Ömer Yılmaz, Özgül, Akçar, Bayazıtoğlu and Gaziulusoy1999) described the K3 Cuneolina gr. C. pavonia-Miliolidae Zone, which is upper Aptian to Cenomanian, from the Seydişehir, Akseki, and Hadim areas (Central Taurides) (Fig. 1.2). The studied benthic foraminiferal assemblage contains few species in common with the Protochrysalidina elongata-Cuneolina pavonia assemblage zone, which was defined by Taslı and Solak (Reference Taslı and Solak2019) from the upper Albian shallow-marine carbonate sequence of the Bey Dağları (Western Taurides) (e.g., Protochrysalidina elongata, Nezzazata isabellae, Nezzazata simplex, Pseudonummoloculina heimi, Peneroplis sp.). It can be correlated with the “Valdanchella” dercourti Taxon-Range Zone (Husinec and Sokač, Reference Husinec and Sokač2006) and Microfossil Assemblage VI (Cvetko Tešović et al., Reference Cvetko Tešović, Glumac and Bucković2011), described from the Adriatic Carbonate Platform and dated as late Albian. It fits well with the late Albian benthic foraminiferal assemblage of Husinec et al. (Reference Husinec, Velić and Sokač2009). An important exception is the absence of Mesorbitolina texana in the latter (Adriatic assemblages). Orbitolinid foraminifers are used for high-resolution biozonation of Early to mid-Cretaceous carbonate platform sediments (e.g., Simmons et al., Reference Simmons, Whittaker, Jones, Hart and Smart2000; Schroeder et al., Reference Schroeder, Buchem, Cherchi, Baghbani, Vincent, Immenhauser, Granier, van Buchem, Al-Husseini, Maurer and Droste2010). Although mesorbitolinids are of variable abundance, they occur frequently through the Geyik Dağı Albian succession (Fig. 2). The last occurrence of Mesorbitolina gr. M. texana is 20 m below the first occurrence of Sellialveolina viallii, indicative of Cenomanian. The stratigraphic range of Mesorbitolina species is extended up to middle Albian in numerous works (e.g., Velić, Reference Velić2007; Husinec et al., Reference Husinec, Velić and Sokač2009; Schroeder et al., Reference Schroeder, Buchem, Cherchi, Baghbani, Vincent, Immenhauser, Granier, van Buchem, Al-Husseini, Maurer and Droste2010), while an uppermost Aptian–upper Albian range of Orbitolina (Mesorbitolina) texana is given by Arnaud-Vanneau (Reference Arnaud-Vanneau, Hardenbol, Jacquin, Farley, de Graciansky and Vail1998). Consequently, the studied succession can be dated as late Albian based on the presence of Protochrysalidina elongata and on its stratigraphic position.
It seems that some taxa have not been reported previously from the Albian strata. Triploporella cf. T. marsicana (Fig. 9.1, 9.2) is a dasycladalean alga known from the lower Aptian (Arnaud-Vanneau, Reference Arnaud-Vanneau1980; Chiocchini et al., Reference Chiocchini, Pampaloni and Pichezzi2012) or Barremian–lower Aptian (Husinec et al., Reference Husinec, Velić and Sokač2009). If the taxon is confirmed, this will be its first record from younger strata. Some specimens (Fig. 8.17, 8.18) comparable with Fischerina? carinata have been described from the upper Albian of the Spanish Pyrenees (Peybernès, Reference Peybernès1984), in association with Mayncina bulgarica.
Conclusions
Although intervening unfavorable microfacies (e.g., ostracod-miliolid wackestones and mudstones) are present, the Urgonian-type limestones of the Geyik Dağı area are rich in rudists and benthic foraminifera. In this paper, the following taxa are identified: Nezzazata isabellae, Nezzazatinella sp., Cuneolina parva, Cuneolina sliteri, Vercorsella arenata, Vercorsella scarsellai, Akcaya minuta, Akcaya auruncensis, Cribellopsis moulladei, Paracoskinolina cf. P. sunnilandensis, Mesorbitolina gr. M. texana, Protochrysalidina elongata, and Pseudonummoloculina aurigerica. Most of these taxa are commonly known from the Barremian–Albian of the peri-Mediterranean platforms. The recently described Phenacophragma oezeri, a mayncinid taxon, seems to be restricted to the Albian and may be stratigraphically important. A spirocyclinid (Reissella sp.) and an ?ophthalmiidid (Paleocornuloculina sp.) are unknown benthic foraminifera from the Albian strata and probably new species. The late Albian age is based on the co-occurrence mainly of Protochrysalidina elongata, and Cuneolina parva, Cuneolina sliteri, Cribellopsis moulladei, Paracoskinolina cf. P. sunnilandensis, Pseudonummoloculina aurigerica, and Pseudonummoloculina heimi.
Acknowledgments
This study was supported by the Research Fund of Mersin University in Turkey with Project Number: 2020-1-AP1-3837. We are grateful to M. Septfontaine (Switzerland) and D. Altıner (Turkey) for valuable recommendations, which improved the manuscript. We thank S. Salar (Mersin University) for the preparation of thin sections.











