


Introduction
The Batocrinidae are iconic crinoids from the Lower Mississippian “Age of Crinoids” (Kammer and Ausich, Reference Kammer and Ausich2006). They were first described in some of the earliest studies of North American crinoid faunas, including Troost (Reference Troost1849, Reference Troost1850a, Reference Troost1850b), Casseday (Reference Casseday1854), Hall (Reference Hall1858, Reference Hall1859a), Lyon and Casseday (Reference Lyon and Casseday1859), and Casseday and Lyon (Reference Casseday and Lyon1862). Because of the typical high abundance of batocrinids in many faunas and taxonomic practices of the late 19th century, seemingly innumerable batocrinid species were named. For example, Miller and Gurley (Reference Miller and Gurley1893, Reference Miller and Gurley1894a, Reference Miller and Gurleyb, Reference Miller and Gurley1895a, Reference Miller and Gurleyb, Reference Miller and Gurley1896a, Reference Miller and Gurleyb, Reference Miller and Gurleyc, Reference Miller and Gurley1897) described more than 100 species of Batocrinus from the Mississippian of Iowa, Missouri, Illinois, Indiana, and Kentucky. Bassler and Moodey (Reference Bassler and Moodey1943) listed more than 130 valid species of only Batocrinus. This taxonomic morass was resolved in part by Lane (Reference Lane1958, Reference Lane1963a, Reference Laneb) and Ausich and Kammer (Reference Ausich and Kammer2010). Today, the Batocrinidae remain a very high-diversity family (15 genera, 154 species). The species remain over-split, but the generic diagnoses are well resolved (Ausich and Kammer, Reference Ausich and Kammer2010). The next phase of batocrinid research will be work on contemporaneous faunas in order to delineate diagnosable species within genera, which is one of the objectives of the present study.
Most Mississippian crinoid families with high richness and abundance are cosmopolitan, occurring on at least the present-day North American and European continents. Two notable exceptions are known among North American crinoids: the Batocrinidae (Ausich and Kammer, Reference Ausich and Kammer2010) and the Coelocrinidae (Agaricocrinus Hall, Reference Hall1858; Aorocrinus Wachsmuth and Springer, Reference Wachsmuth and Springer1897; Coelocrinus Meek and Worthen, Reference Meek and Worthen1865; and Dorycrinus, Roemer, Reference Roemer1854) (Webster and Webster, Reference Webster and Webster2013). Further, with one notable exception (Eretmocrinus sawdoi Itano et al., Reference Itano, Webster, Houck and Bateman2003; Middle Pennsylvanian, Colorado), the Batocrinidae is confined to the Tournaisian and Viséan of North America.
In the present study, species-level systematics is evaluated for the Fort Payne Formation Batocrinidae that occur in the outcrop belt from north-central Kentucky to northeastern Alabama (Fig. 1). Revisions resulting from this study include description of one new species, one species is reassigned to a different genus, and thirteen species are designated as junior synonyms.
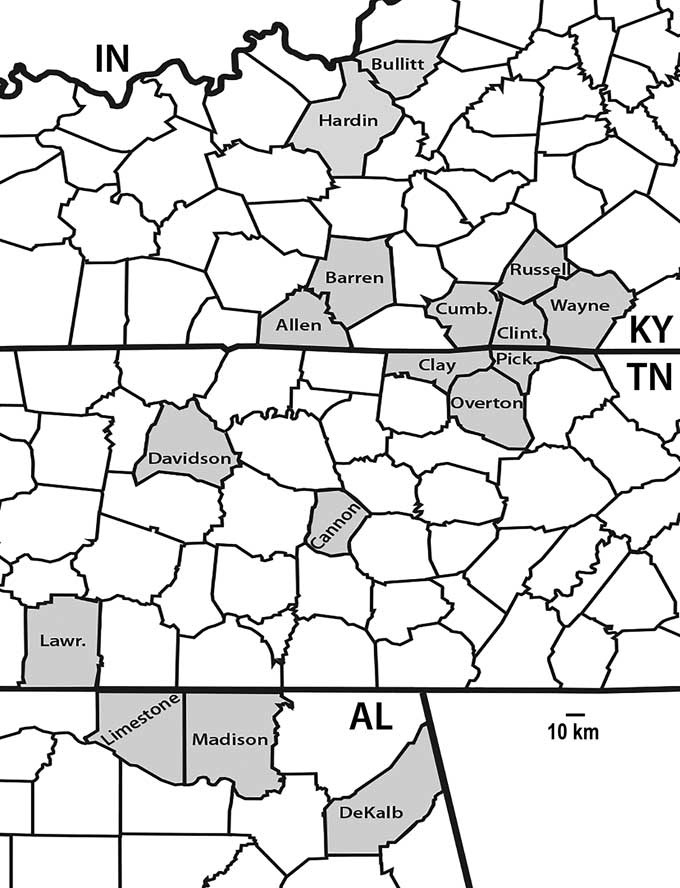
Figure 1 County occurrences of crinoids from the Fort Payne Formation and coeval strata from north-central Kentucky to northern Alabama (Cumb.=Cumberland; Lawr.=Lawrence; Pick.=Pickett).
Geologic setting
For being part of the ‘stable’ craton, the Eastern Interior Seaway of Laurentia was a dynamic setting during the early and middle Mississippian. The Tournaisian was punctuated by a glacial epoch that resulted in the draining of epicontinental seas from much of middle and eastern Laurentia (Kammer and Matchen, Reference Kammer and Matchen2008). Following the glacial epoch (by at least the late Tournaisian) epicontinental seas were re-established from the continental arch to the emerging Appalachian highlands (Matchen and Kammer, Reference Matchen and Kammer2006). The Eastern Interior Seaway, an epicontinental basin, ran north-south encompassing much of present-day Tennessee, Kentucky, Illinois, and Indiana. Maximum water depths in this epicontinental basin are estimated to have exceeded 90 m (Sedimentation Seminar, 1972; Ausich and Meyer, Reference Ausich and Meyer1990). The western margin of this basin was flanked by the Burlington Limestone-Keokuk Limestone carbonate ramp (H.R. Lane, Reference Lane1978), and the eastern margin was dominated by two separate and sometimes overlapping, prograding sediment wedges: the mixed carbonate-siliciclastic Fort Payne Formation and the siliciclastic Borden Group (or Formation). The Borden Delta prograded from present-day east and northeast, and the Fort Payne Formation prograded from present-day east and southeast. Both advanced as clinoforms with recognizable platform, slope, toe-of-slope, and basinal facies. (Swann et al., Reference Swann, Lineback and Frund1965; Lewis and Potter, Reference Lewis and Potter1978; Ausich and Meyer, Reference Ausich and Meyer1990; Greb et al., Reference Greb, Potter, Meyer and Ausich2008). The time-transgressive nature of the prograding Borden and Fort Payne formations has been documented in several studies (e.g., Ausich et al., Reference Ausich, Kammer and Lane1979; Ausich and Meyer, Reference Ausich and Meyer1990; Leslie et al., Reference Leslie, Ausich and Meyer1996; Richardson and Ausich, Reference Richardson and Ausich2004), as has the sequence stratigraphy and facies architecture (Lewis and Potter, Reference Lewis and Potter1978; Ausich and Meyer, Reference Ausich and Meyer1990; Khetani and Read, Reference Khetani and Read2002; Krause et al., Reference Krause, Meyer and Ausich2002; Krause and Meyer, Reference Krause and Meyer2004; Greb et al., Reference Greb, Potter, Meyer and Ausich2008).
The time of interest for the present study is the early Viséan (late Osagean) (Ausich and Meyer, Reference Ausich and Meyer1990; Leslie et al., Reference Leslie, Ausich and Meyer1996; Krivicich et al., Reference Krivicich, Ausich and Keyes2013) and is represented by a variety of epicontinental basinal and toe-of-slope facies (Lewis and Potter, Reference Lewis and Potter1978). During this time, recognized in present-day outcrops and subsurface stratigraphy, the toe-of-slope and basinal facies of the Fort Payne Formation extended along a line from north-central Alabama to south-central Kentucky (with the most significant exposures in south-central Kentucky and north-central Tennessee) (Krivicich et al., Reference Krivicich, Ausich and Keyes2013) (Fig. 1). The Muldraugh Member of the Borden Formation in central and north-central Kentucky represents the platform facies that is time-equivalent of the toe-of-slope Fort Payne facies in south-central Kentucky (Lewis and Potter, Reference Lewis and Potter1978; Greb et al., Reference Greb, Potter, Meyer and Ausich2008) (Fig. 2). Farther north, the early Viséan Borden delta is represented by prodeltaic sediments of the New Providence Shale in north-central Kentucky and southern Indiana (Kammer, Reference Kammer1984, Reference Kammer1985), delta slope facies of the Spickert Knob Formation in southern Indiana (Ausich et al., Reference Ausich, Kammer and Lane1979; Rexroad and Lane, Reference Rexroad and Lane1984) and the delta platform facies (Edwardsville Formation) in southern (Monroe County) and central (Montgomery County) Indiana (Lane, Reference Lane1973; Ausich and Lane, Reference Ausich and Lane1980; Ausich, Reference Ausich1983) (Fig. 2). On the western margin of the basin, the early portion of the early Viséan is represented by the Keokuk Limestone, whereas siliciclastic sediments overrode the northern reaches of the western basin margin by later early Viséan time, yielding the lower part of the Warsaw Formation (Kammer et al., Reference Kammer, Brenckle, Carter and Ausich1990).

Figure 2 North-South cross section of early and middle Viséan strata from southern Indiana to north-central Kentucky. Tournaisian conodonts that were recovered from the basal meter of the Fort Payne Formation indicate a highly condensed interval (Leslie et al., Reference Leslie, Ausich and Meyer1996). Tournaisian strata are designated (although not to scale) with a T.
New collections reported here are largely from south-central Kentucky and north-central Tennessee, which are particularly instructive because in most cases the depositional settings of fossiliferous localities are well constrained (Ausich and Meyer, Reference Ausich and Meyer1990; Meyer et al., Reference Meyer, Ausich, Bohl, Norris and Potter1995; Greb et al., Reference Greb, Potter, Meyer and Ausich2008). Batocrinids and other faunal elements occur in both autochthonous and allochthonous Fort Payne Formation facies (Ausich and Meyer, Reference Ausich and Meyer1988, Reference Ausich and Meyer1990, Reference Ausich and Meyer1992; Meyer et al., Reference Meyer, Ausich and Terry1989; Ausich et al., Reference Ausich, Kammer and Meyer1997; Meyer and Ausich, Reference Meyer and Ausich1997; Rhenberg et al., Reference Rhenberg, Ausich and Meyer2016; Thompson and Ausich, Reference Thompson and Ausich2016). The Fort Payne Formation is a mixed carbonate-siliciclastic depositional system with contemporaneous, laterally contiguous facies. The most prominent autochthonous facies are carbonate buildups, crinoidal packstone buildups, and wackestone buildups (Ausich and Meyer, Reference Ausich and Meyer1990) (Fig. 3). An autochthonous green shale facies and the fauna that lived in incised channels (the sediment fill of the channels was allochthonous) were autochthonous.

Figure 3 Distribution of Fort Payne Formation localities with batocrinid occurrences in the Lake Cumberland region, with sedimentary facies indicated. (1) Kentucky and Tennessee; (2) general locality map of south-central Kentucky and north-central Tennessee: BF, Blacks Ferry; CE, Celina; 61N, Kentucky Highway 61 north of Burkesville, Kentucky; (3) roadcut localities along Kentucky Highway 61 south of Burkesville, Kentucky; location on (2) is south of the intersection of Highway 61 and the Cumberland River; 61B, 61D, 61DW, 61R, see Meyer et al. (Reference Meyer, Potter, Thies, Ausich and Leslie1997); (4) localities along the shoreline of Lake Cumberland; see (2): BT, Big Turbidite; BW, Bugwood; CSN, Cave Springs North; CSS, Cave Springs South; GC, Gross Creek; GCW, Gross Creek West, GR, Greasy Creek; HC, Harmon Creek; MGC, Mouth of Gross Creek; LC, Lily Creek; OB, Owens Branch; OC, Otter Creek; PH, Pleasant Hill; SSF, Seventy-Six Falls; WCS, Wolf Creek South; WCCF, Wolf Creek/Caney Fork. Key to symbols: square, wackestone buildup; diamond, fossiliferous geen shale; circle, crinoidal packstone buildup; upside down triangle, channel fill facies; X, allochthonous facies.
Allochthonous facies were the background siltstone facies, sheetlike packstones, deposition that filled in the channel-form packstones, and the Jabez Sandstone Member. The distinction between autochthonous and allochthonous facies is based on a combination of sedimentologic, stratigraphic, and taphonomic criteria. These interpretations were strengthened by the demonstration that each of the autochthonous facies supported statistically different crinoid and blastoid faunas (Krivicich et al., Reference Krivicich, Ausich and Meyer2014). Further, partially complete echinoids are only present in facies interpreted to have been autochthonous (Thompson and Ausich, Reference Thompson and Ausich2016).
The siltstone and sheet packstone facies comprise the vast majority of the volume of the Fort Payne sediments. The autochthonous facies are a small fraction of the total volume of the Fort Payne Formation and tend to co-occur in restricted geographic clusters, presumably along the toe-of-slope near the mouths of submarine canyons (Ausich and Meyer, Reference Ausich and Meyer1990; Greb et al., Reference Greb, Potter, Meyer and Ausich2008).
Regional distribution of crinoids
As noted above, early Viséan facies vary from carbonate ramps in Iowa, Illinois, and Missouri, to deltaic settings in Indiana, to mixed carbonate-siliciclastic settings of Kentucky, Tennessee, and Alabama. Because early Viséan crinoids have been well documented across this entire region (Hall, Reference Hall1858, Reference Hall1859a, Reference Hallb, Reference Hall1860; Lane, Reference Lane1973; Ausich and Lane, Reference Ausich and Lane1982; Kammer, Reference Kammer1984; Ausich and Kammer, Reference Ausich and Kammer1990, Reference Ausich and Kammer1991a, Reference Ausich and Kammerb, Reference Ausich and Kammer1992; Ausich and Meyer, Reference Ausich and Meyer1992; Kammer and Ausich, Reference Kammer and Ausich1992, Reference Kammer and Ausich1993, Reference Kammer and Ausich1994, Reference Kammer and Ausich1996; Ausich et al., Reference Ausich, Kammer and Meyer1997, Reference Ausich, Goldstein and Yates2000; Meyer and Ausich, Reference Meyer and Ausich1997; Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Reference Krivicich, Ausich and Meyer2014; Rhenberg et al., Reference Rhenberg, Ausich and Meyer2016), the paleogeographic distribution of batocrinds can be evaluated with a considerable degree of confidence.
During the early Viséan (late Osagean), the distribution of individual genera and species through this region was variable. Abatocrinus, Alloprosallocrinus, Eretmocrinus, Macrocrinus, and Uperocrinus occurred through the region in both siliciclastic and carbonate facies and in both shallow- and deep-water settings. As noted by Krivicich et al. (Reference Krivicich, Ausich and Meyer2014), one of the most striking aspects of crinoid distributions is that all species of the genus Dizygocrinus were confined to shallower water facies in the Eastern Interior Basin. Dizygocrinus was present in the carbonate ramp setting of the Keokuk Limestone as well as the same basic setting when siliciclastics overtopped the Keokuk ramp (lower part of the Warsaw Formation). Dizygocrinus also occurred in various delta platform facies of the Borden Delta (Edwardsville Formation; Van Sant, Reference Van Sant1964; Ausich, Reference Ausich1983), as well as in the Muldraugh Member of the Borden Formation, which was the platform facies equivalent of the Fort Payne Formation (Ausich et al., Reference Ausich, Goldstein and Yates2000). However, Dizygocrinus was absent from toe-of-slope facies of the New Providence Shale (Kammer, Reference Kammer1984) and the Fort Payne Formation.
Other genera also had more restricted distributions. Eutrochocrinus is only known from the shallow-water carbonates of the Keokuk Limestone on the western portion of the basin. Magnuscrinus only occurred in deeper water settings in the eastern portion of the Eastern Interior Seaway, where it was present in carbonate and siliciclastic settings. Gongylocrinus is only known from three specimens, so any statements about its environmental preferences are tentative. However, the only early Viséan species of this genus occurred in a channel sandstone facies in the shallow-water delta platform facies of the Edwardsville Formation in Indiana.
At the species level, only Abatocrinus steropes (Hall, Reference Hall1859a), Alloprosallocrinus conicus Casseday and Lyon, Reference Casseday and Lyon1862, Macrocrinus mundulus (Hall, Reference Hall1859a), and Uperocrinus nashvillae (Hall, Reference Hall1858) occurred throughout the region in both shallow and deep settings. Alloprosallocrinus conicus and Macrocrinus mundulus are the most ubiquitous taxa, occurring in siliciclastic and carbonate facies and in both shallow- and deep-water settings. Alloprosallocrinus conicus had a preference for carbonate settings, but Macrocrinus mundulus was locally abundant in both facies types. Abatocrinus steropes and Uperocrinus nashvillae occurred only in carbonate facies.
The four widespread species noted above are atypical, as the distribution of most species is more restricted (Table 1). For example, although Abatocrinus steropes is present throughout the region, A. clavigerus is only present in the western, shallow-water carbonates of the Keokuk Limestone; and A. grandis is only known from the eastern side of the Eastern Interior Seaway but is in both shallow- and deep-water settings and in carbonate and siliciclastic facies. A second example illustrating variable distribution within a genus is Uperocrinus. As noted above, Uperocrinus nashvillae is ubiquitous with the exception that it is only present in carbonate facies. However, Uperocrinus marinus (Miller and Gurley, Reference Miller and Gurley1890) is only in the shallow-water, siliciclastic facies in the Edwardsville Formation in central Indiana; and Uperocrinus robustus (Wachsmuth and Springer, Reference Wachsmuth and Springer1897) only occurs in the deep-water, carbonate facies of the Fort Payne Formation. Macrocrinus casualis (Miller and Gurley, Reference Miller and Gurley1895a) is restricted to deep-water Fort Payne facies. Although both Macrocrinus mundulus and Macrocrinus strotobasilaris Ausich and Lane, Reference Ausich and Lane1982 are present in the deeper-water facies, they are much more common in the shallow-water facies of northern Kentucky and southern Indiana. The known distribution of early Viséan batocrinid species is listed in Table 1.
Table 1 Facies distribution of early Visean Batocrinidae in eastern North America.

Facies distribution of Fort Payne Formation Batocrinidae
Batocrinids occurred predominantly in carbonate facies of the Fort Payne Formation; although as noted below and in Table 2 and Figure 3, a few were also present in the autochthonous green shale facies. Summed across all Fort Payne localities, Eretmocrinus magnificus Lyon and Casseday, Reference Lyon and Casseday1859, Alloprosallocrinus conicus, and Uperocrinus robustus were the dominant batocrinids in the Fort Payne Formation; Abatocrinus grandis and Macrocrinus casualis were of intermediate abundance; and the other batocrinids were relatively rare by comparison (Table 2). In a study of Fort Payne crinoids, Krivicich et al. (Reference Krivicich, Ausich and Meyer2014) documented that the autochthonous Fort Payne crinoid assemblages (all crinoid taxa) were comprised of statistically distinct crinoid assemblages, despite many shared taxa. Analysis of only batocrinid Fort Payne assemblages demonstrates the same patterns documented in Krivicich et al. (Reference Krivicich, Ausich and Meyer2014). Batocrinids from the wackestone buildup facies are distinct from those of the crinoidal packstone buildups (Fig. 4). The single exception in Krivicich et al. (Reference Krivicich, Ausich and Meyer2014) remained with an evaluation of only batocrinids, which is Owens Branch wackestone buildup that has exceptionally large flank beds similar to crinoidal packstone buildups. In both analyses, the Owens Branch wackestone fauna was more similar to crinoidal packstone buildups rather than other wackestone buildups (Fig. 4.1, 4.2). If compared to other coeval crinoid-bearing facies across eastern North America, these deep-water carbonate buildups and the allochthonous facies that were sourced by them are distinct from other crinoid assemblages (Krivicich et al., Reference Krivicich, Ausich and Meyer2014; Fig. 4.3, 4.4).

Figure 4 Results for Principal Coordinates Analysis of Fort Payne Formation batocrinids (presence-absence data evaluated). (1, 2) Coordinate axes 1 and 2 and Coordinate axes 2 and 3; analysis of the batocrinid component of Fort Payne Formation localities; filled circles, wackestone buildups; filled rectangles, crinoidal packstone buildups; open rectangles, allochthonous facies; BUG, Bugwood Wackestone Buildup; CSN, Cave Springs North Crinoidal Packstone Buildup; CSS, Cave Springs South Crinoidal Packstone Buildup; GC, Gross Creek Crinoidal Packstone Buildup; GCW, Gross Creek West Crinoidal Packstone Buildup; LC, Lily Creek Wackestone Buildup; OB, Owens Branch Wackestone Buildup; PH, Pleasant Hill Wackestone Buildup; SSF, Seventy-Six Falls; WCCF, Confluence of Wolf Creek and Caney Fork; 301, Lawrence County, Tennessee; 61DW, Highway 61 D West; 61SR, Highway 61 South Ramp (see Supplementary Data 3); (3, 4) Ed-IC, Edwardsville Formation, Indiana Creek, Indiana; Ed-PB, Edwardsville Formation crinoidal packstone facies; Ed-Sh, Edwardsville Formation, shale facies, Indiana; Ed-Silt, Edwardsville Formation siltstone facies, Indiana; Ed-Ss, Edwardsville Formation sandstone facies, Indiana; FP-GS, Fort Payne Formation green shale facies; FP-PB, Fort Payne Formation crinoidal packstone buildup facies; FP-SP, Fort Payne Formation sheetlike packstone facies; FP-WB, Fort Payne Formation wackestone buildup facies; KeoL, Keokuk Limestone, Illinois, Iowa, and Missouri; LoWar, Lower Warsaw Formation, Illinois, Iowa, and Missouri; Mul, Muldraugh Formation, Kentucky; NPSh, New Providence Shale Member of the Borden Formation, Kentucky (see Krivicich et al., Reference Krivicich, Ausich and Meyer2014 for further details on localities).
Table 2 Distribution and relative abundance of Batocrinidae among facies and localities Kentucky and Tennessee with 25 or more batocrinid specimens known; total number of species per locality is based on Krivicich et al., Reference Krivicich, Ausich and Meyer2014.

The wackestone buildup facies had Alloprosallocrinus conicus as one of the three most dominant species, Eretmocrinus magnificus of secondary importance, and other batocrinids as a minor component. In contrast, in the crinoidal packstone buildup facies, both Eretmocrinus magnificus and Alloprosallocrinus conicus were of primary importance, and many other batocrinid species were only a minor component (Krivicich et al., Reference Krivicich, Ausich and Meyer2014).
Alloprosallocrinus conicus and all of the species of Eretmocrinus (E. magnificus, E. ramulosus, and E. spinosus) not only occur in carbonate buildup facies, but also are present in low abundance in the autochthonous green shale facies. Other genera and species are restricted to carbonate facies. Abatocrinus grandis, Macrocrinus casualis, Magnuscrinus cumberlandensis n. sp., Uperocrinus nashvillae, and Uperocrinus robustus all occur on both types of carbonate buildups, although their abundances on each type are variable. The only facies-restricted batocrinids are Abatocrinus steropes and Magnuscrinus kammeri, which were present on crinoid packstone buildups.
Most batocrinids were also known from the allochthonous sheet-like packstone facies, but batocrinids are absent from the channel-form packstone facies. Macrocrinus is unusual in that the majority of Fort Payne specimens are only known from allochthonous facies (Table 2). In fact, only Macrocrinus casualis is known from autochthonous facies, where it is present only in both types of buildups (Table 2).
We cannot speculate with confidence about the reasons for the strikingly different distributional patterns among some genera and species of the Batocrinidae. However, the combination of larval types, aerosol suspension feeding parameters (Ausich, Reference Ausich1980), and holdfast types (largely unknown) may have played important roles. Presumably, crinoids in platform settings would have experienced higher current velocities on average for suspension feeding and would have been subjected to more frequent and more intense storm events. Alternatively, conditions at the toe-of-slope basinal settings should have been more predictable, although the presence of sheet packstone facies indicates that this setting was periodically disrupted by storms and/or by down-slope sediment slides. The two contrasting Fort Payne buildups may have offered contrasting sediments for holdfast anchoring: coarse, poorly sorted skeletal debris on carbonate packstone buildups versus a semi-consolidated carbonate mud on wackestone buildups (Ausich and Meyer, Reference Ausich and Meyer1990; Meyer et al., Reference Meyer, Ausich, Bohl, Norris and Potter1995). It is also possible that the presumed microbial nature of mud accumulation on wackestone buildups may have offered a food resource that contrasted with that on crinoidal packstone buildups. However, differences in the substratum conditions between these two buildup types did not restrict most batocrinid species.
Materials and methods
As outlined in Supplemental Data 1, crinoid assemblages from specific localities and facies were evaluated using Principal Coordinate Analysis in order to examine the possibility of specific associations of batocrinids in each facies. Further explanation of methods is given in Supplemental Data 1.
Repositories and institutional abbreviations
CMC IP, Cincinnati Museum Center, Cincinnati, Ohio; FMNH, Field Museum of Natural History, Chicago, Illinois; IGS, Illinois State Geological Survey, Champaign, Illinois; IU, Department of Earth and Atmospheric Sciences, Indiana University, Bloomington, Indiana; OSU, Orton Geological Museum, The Ohio State University, Columbus, Ohio; UI, Department of Geology, University of Illinois, Champaign, Illinois; USNM, Springer Room, U.S. National Museum of Natural History, Washington, D.C.
Systematic paleontology
The synonymies listed include only key references and references with representative illustrations. Complete synonymies for taxa discussed here (Supplemental Data 2), detailed locality information (Supplemental Data 3), and tables of measurements (Supplemental Data 4) are companions to the present paper. In addition, most of the citations listed in Supplemental Data 2 will also be listed in Webster and Webster (Reference Webster and Webster2013).
Morphological terminology follows Ubaghs (Reference Ubaghs1978a) and Ausich et al. (Reference Ausich, Brett, Hess, Simms, Hess, Ausich, Brett and Simms1999). Abbreviations used in designating measurements include CaH, calyx height; CaW, calyx width; BH, basal plate height; BW, basal plate width; BRW, basal rim width; TH, tegmen height (from arm openings to base of anal tube). An * indicates a measurement of an incomplete or of a compressed specimen. Class- and order-level classification follows Wright et al. (Reference Wright, Ausich, Cole, Peter and Rhenberg2017).
Class Crinoidea Miller, Reference Miller1821
Subclass Camerata Wachsmuth and Springer, 1885
Infraclass Eucamerata Cole, Reference Cole2017
Order Monobathrida Moore and Laudon, Reference Moore and Laudon1943
Suborder Compsocrinina Ubaghs, Reference Ubaghs1978b
Superfamily Carpocrinoidea de Koninck and LeHon, Reference Koninck and Le Hon1854
Family Batocrinidae Wachsmuth and Springer, 1881
Remarks
Both Azygocrinus decoris (Miller, Reference Miller1892c) and Glannearycrinus spergenensis (Miller, Reference Miller1892a) have been listed as occurring in the Fort Payne Formation (e.g., Webster and Webster, Reference Webster and Webster2013). Both of these taxa were originally described from the “Warsaw Formation” of Kentucky, and we regard the Fort Payne occurrences as erroneous.
Genus Abatocrinus Lane, Reference Lane1963a
Type species
Actinocrinus turbinatus Hall, Reference Hall1858, by original designation.
Other species
See Ausich and Kammer (Reference Ausich and Kammer2010) for full list of species currently assigned to Abatocrinus. Early Viséan species of Abatocrinus include A. clavigerus (Hall, Reference Hall1859a), A. grandis, and A. steropes. Abatocrinus clavigerus is only known from the Keokuk Limestone in Illinois, but the latter two species have more widespread geographic distributions, as discussed below.
Diagnosis
Calyx shape low to medium cone or expanding with concave sides, or rarely low bowl; basal concavity absent; calyx as high as or higher than tegmen; calyx plates low, convex, nodose, very nodose, or rarely with ray ridges; plates commonly with distinct sutures; basal plates low or high, truncate or with proximal expansion formed by nodes; radial plates high; first primibrachial tetragonal; rays not lobate; regular interrays not in contact with tegmen; CD interray not in contact with tegmen; tegmen flat to low inverted bowl, flat to low cone, or rarely concave sides; tegmen plates smooth, nodose, or spinose; anal tube cylindrical; arm facets face outward (subvertical); free arms 18–26, unbranched, and rarely spatulate distally (from Ausich and Kammer, Reference Ausich and Kammer2010).
Occurrence
Abatocrinus is confined to Laurentia, where constituent taxa range in age from the Tournasian to Viséan (early Kinderhookian to Meramecian).
Remarks
Species characters for Abatocrinus are relative calyx size, degree of widening of the calyx at the position where the arms become free, calyx plate sculpturing, number of secundibrachials, number of interradial plates in regular interrays, and number of free arms.
Abatocrinus grandis (Lyon and Casseday, Reference Lyon and Casseday1859)

Figure 5 Species of Abatocrinus from the Fort Payne Formation: (1–4) Abatocrinus grandis; (1, 2) C-ray lateral view and basal view, respectively (USNM 639900); (3, 4) oral view of tegmen and D-ray oral view of calyx, respectively (USNM 639901); (5, 6) Abatocrinus steropes; (5) C-D interray lateral view (USNM 639904); (6) basal view of a partially geodized and compacted specimen, most of aboral cup missing to reveal geode within the calyx (USNM 639906). Scale bars, as indicated.
1849 Actinocrinites Urna Reference TroostTroost, p. 419 (nomen nudum).
1859 Actinocrinus sp. nobis (grandis) Reference Lyon and CassedayLyon and Casseday, p. 240.
1880 Actinocrinus wachsmuthi Reference WhiteWhite, p. 162, pl. 40, fig. 1a, b.
1895a Batocrinus casula Reference Miller and GurleyMiller and Gurley, p. 8, pl. 1, figs. 7, 8.
1895a Batocrinus honorabilis Reference Miller and GurleyMiller and Gurley, p. 11, pl. 1, figs. 9, 10.
1897 Batocrinus grandis (Lyon and Casseday); Reference Wachsmuth and SpringerWachsmuth and Springer, p. 381, pl. 1, fig. 2, pl. 27, figs. 1b, 2a, b.
1958 Abatocrinus grandis (Lyon and Casseday); Lane, p. 102, pl. 2, fig. 7.
1963a Abatocrinus grandis (Lyon and Casseday); Lane, p. 697.
1964 Abatocrinus grandis (Lyon and Casseday); Van Sant, p. 108, pl. 7, figs. 5–8, figs. 17.1, 19.4, 36.
2013 Abatocrinus grandis (Lyon and Casseday); Reference Webster and WebsterWebster and Webster, p. 682.
Complete synonymy list in Supplemental Data 2.
Holotype
USNM S 586.
Diagnosis
Calyx medium to large, slightly expanded at position of arm openings; transverse or circular nodes on calyx plates; two secundibrachials; three or four interradial plates in regular interrays; 20–26 arms.
Occurrence
Abatocrinus grandis was originally reported from the Edwardsville Formation (early Viséan) at Crawfordsville, Indiana. The type specimen(s) of junior synonyms are from the following: Batocrinus casula holotype from Little Barren River, Kentucky (presumably the Fort Payne Formation); Batocrinus honorabilis holotype from “Keokuk Group” of Tennessee (presumably the Fort Payne Formation); Actinocrinites urna (nomen nudum) holotype from the “Keokuk horizon” Whites Creek Spring, Tennessee (Fort Payne Formation); and Actinocrinites wachsmuthi holotype from the Edwardsville Formation at Crawfordsville, Indiana.
This species is now recognized from the following early Viséan formations: (1) the Fort Payne Formation in Kentucky (Clinton, Cumberland, Russell, and Wayne counties), in Tennessee (Davidson and Lawrence [Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 3] counties), and in Alabama in Limestone County (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 5); (2) the Edwardsville Formation in Indiana (Monroe and Montgomery counties); and (3) the Muldraugh Member of the Borden Formation at Elizabethtown, Hardin County, Kentucky.
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Abatocrinus grandis is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, Gross Creek, and Gross Creek West; wackestone buildup facies at Pleasant Hill, Owens Branch, Harmon Creek, and Otter Creek; and the sheetform packstone facies at Big Turbidite, Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, 61DW, and 61N (Fig. 3).
Description
Calyx medium cone shape, medium to large in size, slightly expanded at level of arm openings (Fig. 5.1), straight-sided from base to position where expansion to level of arm openings begins; plates on aboral cup with central circular or elongate transverse node.
Basal circlet truncate proximally, high, ~20% of cup height, shallow basal concavity formed in part by proximal extension of nodes on basal plates, basal-basal sutures indented; basal plates three, equal in size, elongate transverse nodes on basal plates form proximal part of calyx (Fig. 5.2). Radial circlet ~20% of cup height (Fig. 5.1, 5.4); radial plates five, hexagonal, ~1.5 times wider than high, central elongate transverse node, supports first primibrachial above and first interradials laterally above. Regular interrays not in contact with tegmen, all plates with central circular node; first interradial plate hexagonal, slightly wider than high, smaller than radial plates but larger than first primibrachial. Second range with two additional small plates, third range with a single plate (rarely additional or fewer plates may be present in regular interrays).
Primanal heptagonal, as wide as high, central elongate transverse node, larger than or equal in size to adjacent radial plates, interrupts the radial circlet; second range with three plates; third range with three plates; at least one additional plate in fourth range; CD interray not in contact with tegmen, all plates with central circular node.
Fixed brachials with central circular nodes that nearly form ray ridges, especially in distal fixed brachials. Distal-most fixed brachials in tertibrachitaxis or quartibrachitaxis. First primibrachial as wide as high, much smaller than primaxil, with central elongate transverse node; second primibrachial axillary, pentagonal, large, elongate transverse node. Additional fixed brachials approximately as high as wide; typically two secundibrachials; either third or fourth tertibrachial with arm opening or second tertibrachial axillary; if fixed quartibrachials present, two in number; fixed tertaxil and fixed quartibrachials common in C and D rays, rare in other rays.
Tegmen very low inverted cone from arm openings to base of anal tube; plates large, circular or elongate transverse nodes (Fig. 5.3); anal tube high, eccentric, but no preferential radial or interradial position.
Free arms 20–26, biserial; facets nearly vertical.
Proximal column circular with pentalobate lumen, other details of column not known from Fort Payne specimens.
Materials
The holotype of A. grandis is USNM S 586. Type material of junior synonyms include the following: Batocrinus casula Miller and Gurley, Reference Miller and Gurley1895a, holotype, FMNH UC 6434; Batocrinus honorabilis Miller and Gurley, Reference Miller and Gurley1895a, holotype, FMNH UC 6432; Actinocrinites urna Troost (nomen nudum), specimen from Troost collection, USNM 39896; Actinocrinus wachsmuthi White, Reference White1880, syntypes, CMC IP 3256 and FMNH UC 6343. Additional specimens include the lots USNM S 587, USNM S 588, and numerous complete and partial specimens collected from the Fort Payne exposures in south-central Kentucky. The following are new Fort Payne Formation specimens from this study USNM 639900–USNM 639903, OSU 54478–OSU 54481, and CMC IP76351–CMC IP76354.
Measurements
See Supplemental Table 3.
Remarks
Specimens of Abatocrinus grandis from the Fort Payne Formation display variability in arm number and calyx shape, which is well within the range of variability described for this species by Lane (Reference Lane1958). Typically A, B, and E rays have four arms; and the C and D rays may have four to seven arms. The tegmen may be either high and nearly equal in geometry to the calyx, or low and a considerably less significant proportion of the theca. Plating of the rays and half-rays adjacent to the CD interray may deviate slightly from that described above with either addition or elimination of plates from the primi- or secondibrachitaxis. The description given above is based only on material from the Fort Payne Formation. Additional details of the arms and column are known on specimens from Crawfordsville, Indiana.
Lane (Reference Lane1963a) placed Actinocrinites urna Troost (nomen nudum) and Actinocrinus wachsmuthi White in synonymy with Abatocrinus grandis. Herein, we designate Batocrinus casula Miller and Gurley, Reference Miller and Gurley1895a and Batocrinus honorabilis Miller and Gurley, Reference Miller and Gurley1895a as junior synonyms of Abatocrinus grandis. These two species were not considered in the Lane (1958, Reference Lane1963a) revisions. In all respects, B. honorabilis corresponds with the morphology of Abatocrinus grandis. Batocrinus casula has more pronounced aboral cup sculpturing than typical, but its morphology is well within the range present in known specimens of Abatocrinus grandis.
Abatocrinus grandis is distinct from other early Viséan Abatocrinus species by differences in calyx shape, plate sculpturing, nature of the basals and basal circlet, the number of plates in the regular and CD interrays, number of secundibrachials, and arm number (Lane, 1958, Reference Lane1963a). Abatocrinus grandis has a medium cone-shaped calyx shape with a straight-sided cup through the secundibrachitaxis, very nodose calyx plates, a high basal circlet, three to four plates in regular interrays, eight to ten total plates in the CD interray, two secundibrachials, and 21–26 arms. Abatocrinus grandis differs from Abatocrinus steropes, which also occurs in the Fort Payne Formation, in that Abatocrinus steropes has fewer and larger plates in the calyx, larger basals, one interradial plate in regular interrays, the primanal and only four or five additional plates in the CD interray, a single axillary secundibrachial, and 18–20 arms. Abatocrinus clavigerus (Hall), which is known only from the Keokuk Limestone at Nauvoo, Illinois, is characterized by having a low, wide calyx that is cylindrical from the base to the first primibrachial above which it widens to the arm openings, low tegmen, low basals, calyx plates that lack sculpturing except the radial plates, one interradial plate in regular interrays, primanal, and a total of four additional plates in other ranges, and 20 arms (Lane, Reference Lane1958).
Abatocrinus steropes (Hall, Reference Hall1859a)
1849 Actinocrinites gibbosus Reference TroostTroost, p. 419 (nomen nudum).
1850b Actinocrinites? Sayi Reference TroostTroost, p. 28, fig. 31 (nomen nudum).
1859a Actinocrinus steropes Hall, p. 43.
1909 Batocrinus gibbosus (Troost); Reference WoodWood, p. 64, pl. 15, fig. 9.
1925 Batocrinus springeranus Reference BasslerBassler, fig. 26.
1958 Abatocrinus steropes (Hall); Lane, p. 111, pl. 2, fig. 8.
1963a Abatocrinus steropes (Hall); Lane, p. 697.
2013 Abatocrinus springeranus Bassler; Reference Webster and WebsterWebster and Webster, p. 685.
2013 Abatocrinus steropes (Hall); Reference Webster and WebsterWebster and Webster, p. 685.
Complete synonymy list in Supplemental Data 2.
Holotype
UI X-840.
Diagnosis
Calyx medium to large, expanded slightly to moderately from above the secundibrachitaxis to the position of arm openings; calyx plates large and relatively few; circular or elongate transverse nodes on all calyx plates; one secundibrachial; one interradial plate in regular interrays; three to five plates in one or two ranges above the primanal; 18–20 arms.
Occurrence
The holotype of Abatocrinus steropes was described from the Keokuk Limestone at Hamilton, Hancock County, Illinois. The type specimen(s) of junior synonyms are from the following: Actinocrinites gibbosus and Actinocrinites sayi (nomina nuda) holotypes, Whites Creek Springs, Davidson Co., Tennessee (Fort Payne Formation); and Batocrinus springeranus holotype, “lower Keokuk”, Overton County, Tennessee (presumably the Fort Payne Formation).
This species is now recognized from early Viséan formations: (1) Keokuk Formation at Hamilton, Hancock County, Illinois; and (2) the Fort Payne Formation in the Lake Cumberland Region in Kentucky (Clinton and Russell Counties) and in Tennessee (Whites Creek Springs, Davidson County, and in Overton County).
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Abatocrinus steropes is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, and Gross Creek; and the sheetlike packstone facies at Seventy-Six Falls.
Description
Calyx medium cone shaped (Fig. 5.5), medium to large in size, slightly to moderately expanded from above the secundibrachitaxis to the level of arm openings; all calyx plates relatively large in size, with large central circular node or elongate transverse node (Fig. 5.6). Basal circlet truncate proximally, high, ~20% of cup height, very shallow basal concavity formed only by proximal extension of transverse nodes on basal plates, basal-basal sutures indented; basal plates three, equal in size, large elongate transverse node proximally. Radial circlet ~25% of calyx height; radial plates nearly as wide as high, five, hexagonal, ~1.5 times wider than high, supporting the first primibrachial above and the only interradial laterally above on either side. In regular interrays a single large interradial plate, octagonal, large central circular node, as high as wide, not in contact with tegmen, smaller than radials.
Primanal heptagonal, slightly wider than high, slightly smaller than adjacent radial plates, interrupts radial circlet, with large central circular node; three plates in second range; if third range present one or two plates.
Fixed brachials with central elongate transverse node. First primibrachial wider than high; second primibrachial axillary; a single, axillary secundibrachial; two to four (typically three) fixed tertibrachials.
Tegmen medium inverted cone from arm openings to base of anal tube, all plates with central circular nodes.
Free arms 18–20, 2–4 arms in A ray; facets large, nearly vertical.
Proximal column circular with pentalobate lumen, other details not known.
Materials
The holotype of A. steropes is UI X-840. Type material of junior synonyms include Actinocrinites gibbosus Troost (nomen nudum), Troost specimen USNM 39892; Actinocrinites? sayi Troost (nomen nudum), Troost specimen USNM 39899; B. springeranus, holotype, USNM S 3870. Specimens collected for this study include USNM 939904–USNM 939906.
Measurements
See Supplemental Table 3.
Remarks
Numerous specimens of Abatocrinus steropes are present in collections of the Fort Payne from the Lake Cumberland area. These specimens differ in relative degree of calyx size at the level of arm openings, as well as distortion of the entire theca through expansive silicification during diagenesis. Undistorted, smaller specimens have a much smaller diameter at the level of the arm openings. Additionally, these specimens may be preserved as an inner calcareous core of a partially silicified specimen (Fig. 5.6). The result of this style of preservation is a very subdued degree of plate sculpturing. In areas where the outer silicified surface is preserved, the plate sculpturing is consistent with other specimens. Most of the larger specimens are enlarged with expansive silicification that displaces thecal plating, yet retaining the relative positions of plates. Expansive silicification of Abatocrinus steropes specimens contrasts with Abatocrinus grandis that commonly has silica replacement of plates, but not expansive silicification.
See the discussion of A. grandis for comparison of A. steropes with other early Viséan species of Abatocrinus.
Previous authors have regarded two of Troost’s nomina nuda taxa to belong to Abatocrinus steropes: Actinocrinites gibbosus Troost, Reference Troost1849 and Actinocrinites? sayi Troost, Reference Troost1850b. Here, Batocrinus springeranus is also designated as a junior synonym of Abatocrinus steropes, which has a typical morphology for Abatocrinus steropes.
Genus Alloprosallocrinus Casseday and Lyon, Reference Casseday and Lyon1862
Type species
Alloprosallocrinus conicus Casseday and Lyon, Reference Casseday and Lyon1862, by monotypy.
Diagnosis
Calyx shape flat bowl or cone; basal concavity present, small or absent; calyx much lower than tegmen; calyx plates smooth to convex; no median ray ridges; plates commonly with indistinct sutures; basal plates low, in basal concavity, no proximal expansion; radial plates low; first primibrachial tetragonal or pentagonal (axillary); rays not lobate; regular interrays not in contact with tegmen; CD interray in contact with tegmen; tegmen high inverted conical; tegmen plates smooth or nodose; anal tube cylindrical; arm facets face outward (subvertical); and free arms 10–20, unbranched, not spatulate distally (from Ausich and Kammer, Reference Ausich and Kammer2010).
Occurrence
Alloprosallocrinus is only known from the early Viséan (late Osagean) of the central United States.
Alloprosallocrinus conicus Casseday and Lyon, Reference Casseday and Lyon1862

Figure 6 Alloprosallocrinus conicus from the Fort Payne Formation: (1) DE-interray lateral view of a specimen with a slightly convex calyx and broadly convex tegmen plates (USNM 639907); (2) CD-interray lateral view of a specimen with a flat calyx and more sharply nodose tegmen plates (USNM 639909); (3) basal view of a calyx with plate sutures visible due to weathering (USNM 639908). Scale bar, as indicated.
1849 Conocrinites Leæ Reference TroostTroost, p. 419 (nomen nudum).
1849 Conocrinites tuberculosus Reference TroostTroost, p. 419 (nomen nudum).
1862 Alloprosallocrinus conicus Reference Casseday and LyonCasseday and Lyon, p. 29.
1862 Alloprosallocrinus depressus Reference Casseday and LyonCasseday and Lyon, p. 31.
1897 Alloprosallocrinus conicus Casseday and Lyon; Reference Wachsmuth and SpringerWachsmuth and Springer, p. 407, pl. 42, figs. 14a–c.
1958 Alloprosallocrinus conicus Casseday and Lyon; Lane, p. 127, pl. 9, figs. 11.
1964 Alloprosallocrinus conicus Casseday and Lyon; Van Sant, p. 110, pl. 7, figs. 2–4; figs. 17.5, 18, 37 (non fig. 11 as given).
1978 Alloprosallocrinus conicus Casseday and Lyon; N.G. Lane, p. T467, fig. 275.3a–c.
2013 Alloprosallocrinus conicus Casseday and Lyon; Reference Webster and WebsterWebster and Webster, p. 800.
Complete synonymy list in Supplemental Data 2.
Lectotype
USNM S 783a.
Diagnosis
See genus diagnosis.
Occurrence
Alloprosallocrinus conicus is a widespread, early Viséan crinoid species. The holotype of Alloprosallocrinus conicus was described from Clear Creek, Hardin County, Kentucky (Muldraugh Formation). The type specimens of junior synonyms, all presumably from the Fort Payne Formation, are from the following: Conocrinus leae and Conocrinus tuberculosus (nomina nuda) holotypes, Whites Creek Springs, Davidson Co., Tennessee; Alloprosallocrinus depressus holotype, Clear Creek, Hardin County, Kentucky; and Alloprosallocrinus gurleyi holotype, “Keokuk Group” Brown County, Kentucky (this must be a mistaken location).
This species is now recognized from the following early Viséan formations: (1) Muldraugh Member of the Borden Formation (Elizabethtown, Hardin County, Kentucky); (2) Fort Payne Formation in Kentucky (Clinton, Cumberland, Russell, and Wayne counties, Kentucky); in Tennessee at Whites Creek Springs (Davidson County), Clay County, Lawrence County (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Localities 5 and 6), and Pickett County; and in Alabama in Dekalb (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 1), Limestone (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 3), and Madison (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 2) counties. In Indiana, Alloprosallocrinus conicus is from the Edwardsville Formation at the Crawfordsville lower quarry, Indian Creek, and Walnut Fork in Montgomery County and at Allens Creek Bank, Monroe County.
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Alloprosallocrinus conicus is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, Gross Creek, and Gross Creek West; wackestone buildup facies at Bugwood, Harmon Creek, Lily Creek, Owens Branch, Pleasant Hill; sheetlike packstone facies at Cove Creek, Obey Creek, Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, 61B, 61D, and 61DW, 61N, and 61RS; and the autochthonous green shale facies at Blacks Ferry and Celina. Old localities lacking precise coordinates include Glasgow and Metcalf counties, Kentucky, Eagle Mill near Livingston, and at Browns Ford, in Tennessee.
Description
Calyx flat cone (Fig. 6.2) or flat bowl shaped (Fig. 6.1), widest at level of arm openings, shallow basal concavity entirely within basal circlet or including as much as one-half of radial circlet; calyx plates flat to gently convex. Outline of calyx at level of arm openings variable, either subcircular, slightly indented interradially or moderately indented interradially.
Basal circlet part of basal concavity; basals three, equal in size, wider than high (Fig. 6.3). Radials five, twice as wide as high, hexagonal. Regular interrays with one to three plates, typically one; commonly first interradial somewhat higher than wide, not in contact with tegmen but in sutural contact on both sides above with the first secundibrachial (which is axillary) and the first tertibrachial, rarely first interradial approximately two and one-half times higher than wide and in contact with tegmen.
Primanal heptagonal, as wide as high, slightly higher and conspicuously narrower than radial plates; three plates in second range, medial plate at least twice as high as wide, in contact with tegmen, and in contact with plate above at approximately the level of arm opening with an oblique suture; two lateral plates of second range variable, either as high as wide or higher than wide.
Primibrachitaxis variable within an individual and between individuals, either two normal primibrachials (first quadrangular, second axillary), two primibrachials with the first not full width of ray, or the first primibrachial axillary. Arm openings in secundibrachials or tertibrachitaxis; arm openings large, vertical or inclined slightly upward.
Tegmen low inverted cone from arm openings to base of anal tube, symmetrical or asymmetrical; plates convex or spinose. Anal tube central to subcentral, high. One thecal pore on either side of arm openings (Fig. 6.1, 6.2).
Free arms 10–12; robust, aborally rounded, and relatively wide and deep.
Materials
Numerous specimens of A. conicus are present at the U.S. National Museum of Natural History, the Field Museum of Natural History, and at Indiana University. The type suite of A. conicus from Clear Creek, Hardin County, Kentucky (USNM S 783) includes four specimens. A lectotype is designated as USNM S 783a, and paratypes are USNM S 783b–d. A specimen from Troost’s work is USNM 39923 (Conocrinites Leæ), but the specimen for Conocrinites tuberculosus is missing (Wood, Reference Wood1909). Type specimens from other junior synonyms include the cotypes of A. depressus (USNM S 1062) and the holotype of Alloprosallocrinus gurleyi (FMNH UC 6275). The following are new Fort Payne Formation specimens from this study USNM 639907–USNM 639914, OSU 54482–OSU 54489, and CMC IP76355–CMC IP76361.
Measurements
See Supplemental Table 4.
Remarks
Variable characters include calyx shape in lateral profile, grouping or non-grouping of arms in each ray, nature of primibrachials, plating of interrays, shape of tegmen, and nature of tegmen plates. In lateral profile, the base of the calyx may be flat, slightly convex, or quite convex. Arms within a ray may be grouped, in which case the maximum calyx width has a pentagonal basal view outline; or arm openings are not grouped, yielding a subcircular outline at the maximum calyx width. As discussed by Van Sant (Reference Van Sant1964, p. 110, 111), the first primibrachial may be present as a typical tetragonal brachial plate, present but some shape so as not to occupy the entire distal suture of the radial plate, or the typical first primibrachial is completely absent so that the first primibrachial is axillary. This variability exists within rays of a single individual or among individuals.
Contrary to discussions in Lane (Reference Lane1958) and Van Sant (Reference Van Sant1964), specimens of Alloprosallocrinus conicus from the Lake Cumberland area may have interrays in contact with the tegmen. In specimens with distinct calyx plate sutures, ~10% of regular interrays have a very long first interradial that is in contact with the tegmen. The CD interray is clearly displayed in three individuals from the Fort Payne Formation, and in every case the middle plate in the second range is elongate and in contact with the tegmen. The tegmen shape may be either conical or have interradial depressions extending vertically along the tegmen. Finally, the plates of the tegmen vary from smooth to nodose.
The morphological variation recognized in Alloprosallocrinus conicus is relatively broad for a Paleozoic crinoid. Newly discovered specimens expand further this disparity with recognition that rarely the regular interrays and/or CD interray are/is in contact with the tegmen in some specimens. However, there are no co-varying sets of characters suggestive of different species, and the variation is regarded as intraspecific, as suggested by Lane (Reference Lane1958) and Van Sant (Reference Van Sant1964).
In 1897, Wachsmuth and Springer questionably reassigned Alloprosallocrinus depressus Casseday and Lyon, Reference Casseday and Lyon1862 to Agaricocrinus. This specimen has never been illustrated, but the cotypes (USNM S 1062) are available and are considered herein a junior synonym of Alloprosallocrinus conicus.
Genus Eretmocrinus Lyon and Casseday, Reference Lyon and Casseday1859
Type species
Eretmocrinus magnificus Lyon and Casseday, Reference Lyon and Casseday1859, by monotypy.
Other species
Early Viséan species of Eretmocrinus are E. remibrachiatus expansus Wachsmuth and Springer, Reference Wachsmuth and Springer1897; E. granuliferous Wachsmuth and Springer, Reference Wachsmuth and Springer1897; E. magnificus, E. ramulosus, and E. spinosus (see Ausich and Kammer, Reference Ausich and Kammer2010 for full list of species assigned to Eretmocrinus). Eretmocrinus cassedayanus is known only from the Edwardsville Formation, Indian Creek, Indiana, and E. remibrachiatus expansus is only known from the Montrose Chert Member of the Keokuk Limestone in Illinois and Iowa. Eretmocrinus spinosus is only recognized from the Fort Payne Formation, but E. magnificus and E. ramulosus have wider geographic distributions, as noted below.
Diagnosis
Calyx shape low to flat cone or expanding with concave sides; basal concavity absent; calyx as high as or lower than tegmen; calyx plates smooth, nodose, spinose, stellate, with or without ray ridges; plates commonly with distinct sutures; basal plates low, truncate, or with wide to very wide lateral expansion at base; radial plates low; first primibrachial tetragonal; rays not lobate; regular interrays not in contact with tegmen; CD interray not in contact with tegmen; tegmen low to medium cone to medium inverted bowl, commonly asymmetric; tegmen plates smooth, convex, or nodose; anal tube cylindrical; arm facets face outward (subvertical); free arms 12–20, unbranched, spatulate distally (from Ausich and Kammer, Reference Ausich and Kammer2010).
Occurrence
Eretmocrinus is recognized from Tournaisian to the early Viséan (early to late Osagean) of North America and has a single anomalous occurrence in the Atokan of Colorado.
Remarks
Early Viséan crinoid faunas in Indiana, Kentucky, and Tennessee are typically well represented by individuals assigned to Eretmocrinus. The Fort Payne Formation is not an exception with three species currently known from the Fort Payne: E. magnificus, E. ramulosus, and E. spinosus. In addition, hybrid specimens assigned to Eretmocrinus magnificus×Eretmocrinus spinosus are recognized (Ausich and Meyer, Reference Ausich and Meyer1994).
Species characters in Eretmocrinus include calyx width to height ratio, calyx height to tegmen height ratio, calyx plate sculpturing, basal circlet percent of calyx height, radial circlet percent of calyx height, dimensions of radial plates, plating in regular interrays, connection or non-connection of CD interray with the tegmen, plating in the CD interray, number of secundibrachials, plate sculpturing of anal tube plates, and arm number.
Eretmocrinus magnificus Lyon and Casseday, Reference Lyon and Casseday1859

Figure 7 Eretmocrinus magnificus from the Fort Payne Formation: (1) A-ray lateral view of a calyx with pustulose calyx plates (USNM 639917); (2, 3) specimen in matrix with a poorly preserved calyx but well-preserved anal tube; (2) enlargement of anal tube showing the contrast between the regenerated portion of the anal tube and the original (USNM 639918); (4–6) specimen with smooth calyx plates; oral view of tegmen, B-ray lateral view of calyx, and basal view of calyx, respectively (USNM 693316). Scale bars, as indicated.
1859 Eretmocrinus magnificus Lyon and Casseday, p. 241.
1895a Batocrinus wetherbyi Reference Miller and GurleyMiller and Gurley, p. 11, pl. 1, figs. 11, 12.
1895a Batocrinus commendabilis Reference Miller and GurleyMiller and Gurley, p. 25, pl. 2, fig. 16.
1897 Eretmocrinus magnificus Lyon and Casseday; Reference Wachsmuth and SpringerWachsmuth and Springer, p. 386, pl. 37, fig. 3.
1958 Eretmocrinus magnificus Lyon and Casseday; Lane, p. 155.
1964 Eretmocrinus commendabilis Miller and Gurley; Van Sant, p. 116, pl. 7, fig. 12.
1978 Eretmocrinus magnificus Lyon and Casseday; N.G. Lane, p. T469, fig. 275.8a.
2013 Eretmocrinus magnificus Lyon and Casseday; Reference Webster and WebsterWebster and Webster, p. 1420.
Complete synonymy list in Supplemental Data 2.
Holotype
USNM S 744a.
Diagnosis
Calyx high cone shaped, width to height ratio 1.66; calyx height to tegmen height ratio 1.18; calyx plate sculpturing variable, most common morphs with apparent smooth calyx plate sculpturing; basal circlet 23% of calyx height; radial circlet 15% of calyx height; radial plates 2.7 times as wide as high; regular interray plating commonly 1-2, but also 1-1, 1-1-1, 1-2-1; CD interray not in contact with tegmen, plating P-3-3 or P-4; two secundibrachials; anal tube with either low circular or horizontally elongate spines; free arms commonly 20 (range 18–21).
Occurrence
The holotype of Eretmocrinus magnificus was described from Hardin County, Kentucky (probably the Muldraugh Member of the Borden Formation). The type specimen(s) of junior synonyms are from the following: Eretmocrinus commendabilis holotype, “Keokuk Group” (Edwardsville Formation) Crawfordsville, Indiana; Batocrinus wetherbyi holotype, Whites Creek Springs, Davidson County, Tennessee (Fort Payne Formation); Batocrinus curiosus holotype, Allen County, Kentucky (probably Fort Payne Formation); Batocrinus lyonanus holotype, Little Bear River, Kentucky (present location not known); and Actinocrinites agassizi holotype, probably Whites Creek Springs, Davidson County, Tennessee (Fort Payne Formation).
This species is now recognized from the following early Viséan formations: (1) the Edwardsville Formation at Monroe County (Allens Creek Bank, Boy Scout Camp, and Waldrip Site) and Montgomery County (Indian Creek and Crawfordsville upper quarry), Indiana; (2) the Fort Payne Formation (Allen, Clinton, Cumberland, Hardin, Russell, and Wayne counties), Kentucky; Clay, Davidson (Whites Creek Springs), Lawrence (Krivicich et al., Reference Krivicich, Ausich and Keyes2013 Locality 5 and 6), and Pickett counties, Tennessee; and Dekalb (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 1) and Madison (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 2) counties, Alabama; and (3) the Muldraugh Member of the Borden Formation (Hardin County, Kentucky).
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Eretmocrinus magnificus is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, Gross Creek, Gross Creek West; wackestone buildup facies at Bugwood, Greasy Creek, Harmon Creek, Lily Creek, Owens Branch, and Pleasant Hill; sheetlike packstone facies at Big Turbidite, Cove Creek, Obey Creek, Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, 61B, 61D, 61DW, 61RS; and authochthonous green shale facies at Blacks Ferry and Celina.
Description
Calyx widest at arm openings, medium to high cone calyx shape, sides concave from base to arm openings, medium to large size for genus. Calyx width to height ratio 1.66 (range 1.14–1.96), calyx height to tegmen height (to base of anal tube) 1.18 (range 0.88–1.48) (Fig. 7.1, 7.5) (Supplemental Table 5). Proximal calyx (before extreme widening to arm bases) medium to high cone shape. Outline of calyx at level of arm openings circular with slight indentations between rays and half rays.
Calyx plate sculpturing variable, but dominant morphology with flat or modestly convex plates with apparent smooth plate sculpturing, but at high magnification, finely pustulose. Rarely some forms with circular to vertically elongate nodes that are not coalesced into ray ridges or other forms that have a weak to strong ray ridge. Basal rim formed from elongate transverse nodes on basal plates extended horizontally, complete or indented at basal-basal sutures, broadly rounded and narrow to sharp and wide (Fig. 7.1. 7.5), width ~50% of the maximum calyx width. Basal circlet high, ~18–30% (mean=23%) of calyx height, shallow basal concavity for proximal columnal. Basal plates three, equal in size, sculpture as noted above. Radial circlet 10–23% of calyx height (mean=15%); radial plates five, hexagonal, as wide as high; plate sculpturing as noted above. Radial plates 1.9–3.7 times as wide as high (mean=2.7), may be higher marginally than medially. Regular interrays not in contact with tegmen. First interradial large, higher than wide, variously shaped; plating typically 1-2, but the following also occur: 1-1, 1-1-1, 1-2-1; plates above first interradial plate may be equal or unequal in size.
Primanal heptagonal, wider than high; plating P-3-3 or P-4; not in contact with tegmen.
First primibrachials, tetragonal wider than high; approximately equal in size to second primibrachial; second primibrachial axillary; additional fixed brachials wider than high; two secundibrachials; last fixed brachials in tertibrachitaxis; free arm facet typically not vertically elongate; surficial plate sculpturing as noted above.
Tegmen medium inverted cone from arm openings to base of anal tube; tegmen plates with elongate circular spines (Fig. 7.4); one thecal opening on either side of arm openings. Anal tube long; plates spinose, either low circular or low horizontally elongate nodes (Fig. 7.2, 7.3).
Free arms commonly 20, range from 18–21. Distal arms not known from Fort Payne material.
Proximal columnal circular with crenularium, lumen pentalobate.
Materials
The cotypes of E. magnificus are USNM S 744; and designated herein from these specimens are a lectotype, USNM S 744a, and paralectotypes USNM S 744b–d. Type material of junior synonyms are as follows: Eretmocrinus commendabilis, holotype, FMNH UC 6475; Batocrinus weatherbyi, holotype, FMNH UC 6430.
More than 400 complete or incomplete specimens of E. magnificus were collected from the Fort Payne Formation in the Lake Cumberland area. Representative collections from this study are USNM 639915–USNM 639920, OSU 54490–OSU54499, and CMC IP76362–CMC IP76367.
Measurements
See Supplemental Table 4.
Remarks
Eretmocrinus magnificus is the most common Eretmocrinus species in the Fort Payne Formation. It is present in nearly every facies, but it is especially common on crinoid packstone buildups, such as Gross Creek and Cave Springs.
As indicated in the description above, E. magnificus is quite variable morphologically, although distinctive morphs exist. The two most common morphs in the Lake Cumberland area are one with a high calyx, convex and smooth calyx plates, and a somewhat expanded basal rim, and another with a shorter calyx, circular to vertically elongate nodes on ray plates and perhaps interradial plates, and a more expanded basal rim.
Lane (Reference Lane1958) regarded B. weatherbyi Miller and Gurley, E. lyonanus Miller, and B. laterna Miller and Gurley as junior synonyms of E. magnificus. We agree with the placement of E. wetherbyi, but assign E. lyonanus and B. laterna as junior synonyms of E. ramulosus, below. We also place E. commendabilis Miller and Gurley in synonymy with E. magnificus. Eretmocrinus commendabilis is an example of the E. magnificus morph with a high calyx, convex and smooth plates, and a somewhat expanded basal rim. Van Sant (Reference Van Sant1964) differentiated E. commendabilis from E. magnificus because of the number of arms, plate sculpturing on fixed brachials, and highly nodose tegmen. The arm number of E. commendabilis is 20, and that of E. magnificus is 20–26. Van Sant (Reference Van Sant1964) noted that E. magnificus has nodes on the fixed brachial plates; however, the most common morph of E. magnificus in the Fort Payne Formation lacks nodes on fixed nodes on fixed brachial plates. Finally, the character of the anal tube is not known on the type specimen of E. commendabilis. Thus, the morphology of E. commendabilis is the same as the common morph of E. magnificus.
Eretmocrinus magnificus is distinct among Fort Payne species of Eretmocrinus because it has a high cone-shaped calyx, width to height ratio 1.66; calyx height to tegmen height ratio 1.18; calyx plate sculpturing variable, most common morphs with apparent smooth calyx plate sculpturing; basal circlet height 23% of calyx height; radial circlet height 15% of calyx height; radial plates 2.7 times as wide as high; regular interray plating commonly 1-2, but also 1-1, 1-1-1, 1-2-1; CD interray not in contact with tegmen, plating P-3-3 or P-4; two secundibrachials; anal tube with either low circular or horizontally elongate spines; free arms commonly 20 (range 18–21). In contrast, E. ramulosus has a calyx width to height ratio 2.54; calyx height to tegmen height ratio 0.66 calyx plate sculpturing variable, radial plates typically with horizontally elongate node, ray plates always with prominent ray ridges or aligned nodes; basal circlet height 9.5% of calyx height; radial circlet height 7.7% of calyx height; radial plates 6.7 times as wide as high; regular interray plating commonly 1-2, but also 1-3 and 1-2-1; CD interray plating P-3-3-2-1; two secundibrachials; anal tube with horizontally elongate spines; free arms commonly 23 (range 20–26). Calyx width to height ratio 2.2; calyx height to tegmen height ratio 0.74; prominent horizontally elongate spines or thick circular spines on calyx plates; basal circlet 13% of calyx height; radial circlet 18% of calyx height; radial plates 3.0 times as wide as high; commonly a singular interray plate but plating may be 1-1; CD interray plating P-3-1; one secundibrachials; anal tube with small plates lacking nodes or spines; free arms commonly 12–15. Eretmocrinus spinosus has a calyx width to height ratio 2.38; calyx height to tegmen height ratio 0.74; prominent horizontally elongate spines or thick circular spines on calyx plates; basal circlet height 13% of calyx height; radial circlet height 18% of calyx height; radial plates height 3.0 times as wide as high; commonly a singular interray plate but plating may be 1-1; CD interray plating P-3-1; one secundibrachials; anal tube with small plates lacking nodes or spines; free arms commonly 12–15.
Eretmocrinus ramulosus (Hall, Reference Hall1858) (non Wachsmuth and Springer, Reference Wachsmuth and Springer1897)

Figure 8 Species of Eretmocrinus from the Fort Payne Formation. (1, 2, 4, 7) Eretmocrinus ramulosus: (1, 2) lateral view and basal view of thecae, respectively of specimen, note spinose tegmen, anal sac plates, and relatively low calyx (USNM 639921); (4) lateral view of calyx, note somewhat higher calyx comparted to specimen illustrated in (1) (USNM 639922); (7) compressed theca from basal view, large specimens are typically crushed in the oral-aboral direction depicted here (USNM 639923). (3, 5, 6, 8, 9) Eretmocrinus spinosus: (3) basal view of calyx in matrix (USNM 639930); (5, 6) CD-interray lateral and basal views of a theca, respectively, note large, broad spines on all calyx plates and circular spines on tegmen (USNM 639928); (8, 9) basal and lateral views of a partial theca, this larger specimen has a higher calyx than the specimen in (5) (USNM 639929). Scale bars, as indicated.
1849 Actinocrinites Agassizi Reference TroostTroost, p. 419 (nomen nudum).
1858 Actinocrinus ramulosus Reference HallHall, p. 615, pl. 15, fig. 7.
1891 Eretmocrinus lyonanus Reference MillerMiller, p. 59, pl. 10, fig. 3.
1895a Batocrinus curiosus Reference Miller and GurleyMiller and Gurley, p. 6, pl. 1, figs. 5, 6.
1895a Batocrinus laterna Reference Miller and GurleyMiller and Gurley, p. 12, pl. 1, figs. 13, 14.
1897 Eretmocrinus ramulosus (Hall); Reference Wachsmuth and SpringerWachsmuth and Springer, p. 387, pl. 37, figs. 4a, b, 5a–d.
1958 Eretmocrinus ramulosus (Hall); Lane, p. 177.
2013 Eretmocrinus ramulosus (Hall); Reference Webster and WebsterWebster and Webster, p. 1421.
Complete synonymy list in Supplemental Data 2.
Holotype
UI X-818.
Diagnosis
Calyx low cone shaped, width to height ratio 2.54; calyx height to tegmen height ratio 0.66; calyx plate sculpturing variable, radial plates typically with horizontally elongate node, ray plates always with prominent ray ridges or aligned nodes; basal circlet 9.5% of calyx height; radial circlet 7.7% of calyx height; radial plates 6.7 times as wide as high; regular interray plating commonly 1-2, but also 1-3 and 1-2-1; CD interray plating P-3-3-2-1; two secundibrachials; anal tube with horizontally elongate spines; free arms commonly 23 (range 20–26).
Occurrence
The holotype of Eretmocrinus ramulosus was described from the Keokuk Limestone, Nauvoo, Hancock County, Illinois. The type specimen(s) of junior synonyms are from the following: Batocrinus curiosus holotype, presumably the Fort Payne Formation, Allen County, Kentucky; Batocrinus lyonanus holotype, presumably the Fort Payne Formation, Little Barren River, Kentucky; Batocrinus laterna holotype, “Keokuk Group” (Fort Payne Formation), Whites Creek Spring; Actinocrinites agassizi holotype, Whites Creek Spring (Fort Payne Formation).
This species is now recognized from the following early Viséan formations: (1) Edwardsville Formation, Montgomery County (Indian Creek), Indiana; (2) Fort Payne Formation in Allen, Barren, Clinton, Cumberland, and Russell counties, Kentucky and in Davidson County (Whites Creek Spring), Tennessee; (3) the Keokuk Limestone at Nauvoo, Hancock County, Illinois and Augusta, Des Moines County, Iowa; and (4) the Montrose Chert Member of the Keokuk Limestone at Keokuk, Lee County, Iowa.
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Eretmocrinus ramulosus is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, and Gross Creek; wackestone buildup facies at Bugwood, Lily Creek, Owens Branch, and Pleasant Hill; and sheetlike packstone facies at Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, and 61RS.
Description
Calyx widest at arm openings, very low to low calyx shape, sides of calyx either straight or concave, medium to large size for genus. Calyx width to height ratio 2.54 (range 2.20–3.11), calyx height to tegmen height (to base of anal tube) ratio 0.66 (range 0.57–0.81). Outline of calyx at level of arm openings circular or circular with slight indentations between rays and half rays (Fig. 8.2). Surficial plate sculpturing variable.
Basal rim extremely variable; formed from elongate transverse nodes on basal plates that extend horizontally or slightly proximally, varies from complete and three times as wide as proximal column to isolated elongate nodes; in larger forms the basal rim is proportionally much narrower (Fig. 8.1, 8.4). Radial plate typically with prominent horizontally elongate node, but smaller circular to vertically elongate node may be present. Rays always with prominent ray ridges formed by vertically coalesced ridges. Regular interradial plates with or without circular node. In addition to nodes and ridges noted above, plates coarsely pustulose at high magnification. Basal circlet low, 5–17% of calyx height (mean=9.5%), very shallow basal concavity for proximal columnal. Basal plates three, equal in size, sculpture as noted above. Radial circlet 6–11% of calyx height (mean=7.7%); radial plates five, hexagonal or heptagonal, 4.8–10.3 times as wide as high (mean=6.7); sculpture as noted above; may be higher marginally than medially.
Regular interrays not in contact with tegmen. First interradial large, as high as wide, hexagonal; plating typically 1-2, but also 1-3 and 1-2-1; plating above first interradial may be equal. Plate sculpturing as noted above.
Primanal heptagonal, approximately as high as wide, similar in size and sculpturing to radial plates; plating P-3-3-2-1 in only example known; not in contact with tegmen.
First primibrachial wider than high; approximately equal in size or smaller to second primibrachial; second primibrachial axillary; additional fixed brachials wider than high; two secundibrachials; last fixed brachials at tertibrachial three; free arm facet vertically elongate, proximal elongation due to ray ridges; surficial plate sculpturing as noted above.
Tegmen medium inverted bowl from arm openings to the base of anal tube; tegmen plates with elongate circular spines; one thecal opening on either side of arm facets (Fig. 8.1). Anal tube high; plates with horizontally elongate spines.
Free arms 20–26 (mean=23). Distal arms not preserved on calyx of Fort Payne specimens.
Proximal-most column circular, narrow crenularium, wide areola, pentalobate lumen, distal column not known.
Materials
The holotype of E. ramulosus is UI X-818. Type material of junior synonyms include the following: Batocrinus curiosus, holotype FMNH UC 6436; Batocrinus lyonanus holotype, FMNH UC 6276; Batocrinus laterna, holotype FMNH UC 6431; and Actinocrinites agassizi Troost, the specimen that would have become a type, USNM 39893. The following Fort Payne material is deposited in the USNM: USNM S 752 (12 specimens), USNM S 753 (two specimens), USNM S 754 and USNM S 756 (two specimens). New specimens from the present study include USNM 639921–USNM 639927, OSU 54500–OSU 54501, and CMC IP76368–CMC IP76371.
Measurements
See Supplemental Table 5.
Remarks
Eretmocrinus ramulosus is a relatively rare species in the Fort Payne Formation. Like other Eretmocrinus, it is most common on crinoid carbonate buildups and wackestone buildups. The strong ray ridges, high calyx width to height ratio, and high radial plate width to height ratio are the most distinctive features of this species.
Lane (Reference Lane1958) placed B. curiosus, B. casula, and A. agassizi in synonymy with E. ramulosus. We agree with these assignments, and regard E. lyonanus and B. laterna to be junior synonyms of E. ramulosus.
The size range distribution of specimens assigned to E. ramulosus is bimodal. Only smaller specimens (e.g., Fig. 8.1) are listed in Supplemental Table 5, because these smaller specimens presumably had more firmly cemented calyx plate sutures so that they were more readily preserved in their original three-dimensional shape. Alternatively, the larger specimens (Fig. 8.7) are all compressed due to compaction so that they cannot be accurately compared biometrically to the smaller specimens. The smaller specimens are in the same basic size range as specimens of E. magnificus, whereas the calyx width of the larger morph is as much as twice as wide. In addition to the size distinction, the larger morph has pronounced nodes on interradial plates, rays may be more defined by nodes than ridges, and the basal rim is relatively narrower and indented at basal-basal plate sutures. In contrast, the smaller morph has inconspicuous nodes or no nodes on interradial plates, rays are defined by ridges, and the basal rim is relatively larger and typically is a continuous rim. We consider these differences to be ontogenetic until a time when numerous well-preserved thecae of all sizes can be compared biometrically. Both morphs appear to have the same basic overall geometry (although this cannot be positively confirmed), the calyces of both are dominated by ray ridges (although the style may be different), and they both have modestly sized pustulose sculpturing in addition to the conspicuous nodes and ridges.
For comparison to other Fort Payne species of Eretmocrinus, see remarks of E. magnificus.
Eretmocrinus spinosus Miller and Gurley, Reference Miller and Gurley1895a
Figure 8.3, 8.5, 8.6, 8.8, 8.9
1850b Actinocrinites Marineri Reference TroostTroost, p. 25, fig. 27 (nomen nudum).
1895a Batocrinus spinosus Reference Miller and GurleyMiller and Gurley, p. 5, pl. 1, figs. 1–3.
2013 Magnuscrinus spinosus (Miller and Gurley); Reference Webster and WebsterWebster and Webster, p. 1795.
2014 Magnuscrinus praegravis (Miller, Reference Miller1892a); Krivicich et al., p. 1155.
Complete synonymy list in Supplemental Data 2.
Holotype
FMNH UC 6435.
Diagnosis
Calyx low cone shaped, width to height ratio 2.38; calyx height to tegmen height ratio 0.60; prominent horizontally elongate spines or thick circular spines on calyx plates; basal circlet 13% of calyx height; radial circlet 18% of calyx height; radial plates 3.0 times as wide as high; commonly a singular interray plate but plating may be 1-1; CD interray plating P-3-1; one secundibrachial; anal tube with small plates lacking nodes or spines; free arms commonly 12–15.
Occurrence
The holotype of Eretmocrinus spinosus was described from Little Barren River, (probably) Barren County, Kentucky (presumably the Fort Payne Formation). Actinocrinites marineri was originally described from Cannon County, Tennessee and Little Barren River, Barren County, Kentucky (both presumably the Fort Payne Formation).
This species is now recognized from the Fort Payne Formation (early Viséan) in Barren, Clinton, Cumberland, and Russell counties, Kentucky and in Cannon, Lawrence (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 5), and Pickett counties, Tennessee; and in Dekalb (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 1) and Limestone (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 3) counties, Alabama.
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Eretmocrinus spinosus is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, Gross Creek, and Gross Creek West; wackestone buildup facies at Bugwood, Lily Creek, and Owens Branch; and sheet-form packstone facies at Cove Creek, Obey Creek, Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, 61B, 61D, and 61RS.
Description
Calyx widest at arm openings, sides straight from base to arm openings, low cone shaped (Fig 8.5, 8.9), large size for genus. Calyx width to height ratio 2.38 (range 1.87–2.49), calyx height to tegmen height (to base of anal tube) ratio 0.60 (range 0.33–0.86). Calyx medium to low cone shape, height to width ratio 1.7–3.3 (mean=2.5), medium to large size for genus, widest at level of arm openings, straight sided from base to level of arm openings. Outline of calyx at level of arm openings circular. All calyx plates nodose; nodes commonly large, horizontally elongate nodes, terminations sharp, rounded, or fluted (Fig. 8.3, 8.8); proximal elongate nodes may project downward (Fig. 8.9); plate sculpturing dominates appearance of calyx.
Basal circlet truncate proximally, high, 6–16% of calyx height (mean=13%), visible in side view; very shallow basal concavity formed by nodes. Basal plates three (Fig. 8.6, 8.7), equal in size, with an elongate transverse extension of basal plates forming proximal rim of calyx, nodes project horizontally or slightly proximally, but not beyond radials; basal rim always deeply indented at basal-basal sutures. Radial circlet 12–25% of calyx height (mean=18%); radial plates five, hexagonal or heptagonal, 2.2–4.1 times wider than high (mean=3.0). Regular interrays not in contact with tegmen, all plates nodose, typically a single interradial plate, may have one additional plate above; first interradial heptagonal to nonagonal, higher than wide, either smaller or larger than radials.
Primanal heptagonal, approximately as wide as high, central node prominent, but less transversely elongate than node on radials; plating P-3-1, not in contact with tegmen.
Fixed brachials all nodose. First primibrachial very small, wider than high, may not extend full width of ray so that the radial plate is in partial sutural contact with second primibrachial, much reduced transverse elongate node; second primibrachial axillary, large, pentagonal, central node. Only one secundibrachial, axillary. Additional fixed brachials slightly wider than high, last fixed brachial commonly tertibrachial three. Free arm facets elongate, inclined slightly upward.
Tegmen low inverted cone from arm openings to base of anal tube; plates large, spinose; anal tube long eccentric toward anterior; one thecal opening on either side of arm facet. Anal tube tall, narrow, small plates without nodes or spines.
Free arms 12–15; characters of the free arms are not known.
Proximal columnals circular, wide crenularium, very narrow aureola or absent, wide trilobate or pentalobate lumen; one nudinodal separated by one internodal. Character of remainder of column not known.
Materials
The holotype of B. spinosus is FMNH UC 6435, and the holotype of A. marineri is USNM 39895. Additional specimens from the Lake Cumberland area are USNM 639928–USNM 639932. OSU 54502–OSU54505, and CMC IP76372–CMC IP76375.
Measurements
See Supplemental Table 5.
Remarks
Eretmocrinus spinosus is a Miller and Gurley species that has not been considered for more than one century. Re-examination of Fort Payne Formation specimens from the Lake Cumberland region using the generic definitions of Ausich and Kammer (Reference Ausich and Kammer2010) has revised our understanding of this taxon. In many previous studies the crinoid identified as Magnuscrinus praegravis (né Eretmocrinus praegravis) is now recognized as Eretmocrinus spinosus (né Batocrinus spinosus). Magnuscrinus praegravis and E. spinosus are convergent in overall thecal shape with a low calyx, high tegmen, and dominant calyx and tegmen plate sculpturing. A key diagnostic character that separates Magnuscrinus and Eretmocrinus is the morphology of the interradial regions. In Magnuscrinus, the plating of the interradial regions is continuous with plating on the tegmen, whereas in Eretmocrinus it is not. The taxon in question lacks a connection between interradial and tegmen plating; thus it should be assigned to Eretmocrinus. Further, the horizontal, elongate, very large, downward-projecting nodes on the specimens now assigned to E. spinosus are consistent with the holotype of E. spinosus and not with that of Magnuscrinus praegravis. The only previous study that included both of these species was Wood (Reference Wood1909). She regarded them as distinct species, and we agree.
For comparison to other Fort Payne species of Eretmocrinus, see remarks of E. magnificus.
Eretmocrinus magnificus Lyon and Cassseday, 1859 ×
Eretmocrinus spinosus Miller and Gurley, Reference Miller and Gurley1895a
1895a Batocrinus laciniosus Reference Miller and GurleyMiller and Gurley, p. 14, pl. 1, figs. 15, 16.
1994 Eretmocrinus magnificus Lyon and Cassseday×Eretmocrinus praegravis Miller; Ausich and Meyer, p. 362, fig. 1b, c.
2013 Eretmocrinus laciniosus Miller and Gurley; Reference Webster and WebsterWebster and Webster, p. 1419.
Occurrence
All specimens currently recognized as Eretmocrinus magnificus×E. spinosus are from the Fort Payne Formation. The original specimens were described from localities along Lake Cumberland, but with the addition of Batocrinus laciniosus to Eretmocrinus magnificus×E. spinosus, this hybrid is also recognized from what is interpreted to be the Fort Payne Formation from an unspecified site in Tennessee.
Materials
The specimens assigned to this hybrid by Ausich and Meyer (Reference Ausich and Meyer1994) are USNM 463329 and USNM 463330. The holotype of Batocrinus laciniosus, also regarded as this hybrid, is FMNH UC 6433.
Remarks
As noted above, Lake Cumberland region crinoids previously identified in Ausich and Meyer (Reference Ausich and Meyer1994) as Eretmocrinus praegravis Miller, Reference Miller1892a are now regarded as Eretmocrinus spinosus; thus, changing the name of the hybrid specimens recognized by Ausich and Meyer (Reference Ausich and Meyer1994) to Eretmocrinus magnificus Lyon and Cassseday, 1859×E. spinosus Miller and Gurley, Reference Miller and Gurley1895a. Only three specimens of this unusual morphology are known and the morphology is variable; therefore, we regard these as hybrid specimens rather than a distinct species with a hybrid origin. As argued by Ausich and Meyer (Reference Ausich and Meyer1994), the three hybrid specimens occurred at localities (Gross Creek Buildup and Cave Springs Buildup) that supported both Eretmocrinus magnificus and Eretmocrinus spinosus. The hybrid specimens have an intermediate calyx shape and calyx plate sculpturing between these two parent species. Using Stepwise Discriminant Functional Analysis, Ausich and Meyer (Reference Ausich and Meyer1994) demonstrated that the two parent species were well resolved where the hybrid specimens were excluded; whereas if analyses included the hybrids, species separation in multidimensional space became blurred resulting in several “misclassified” specimens. Also, we regard Batocrinus laciniosus Miller and Gurley, Reference Miller and Gurley1895a to be more correctly described as Eretmocrinus magnificus×E. spinosus, rather than a separate valid species.
Hybridization between two species of the same genus is relatively common in nature among both plants and animals, including echinoderms (e.g., Swan, Reference Swan1953; Hyman, 1955; Hinegardner, Reference Hinegardner1975; Menge, Reference Menge1986; Kwast et al., Reference Kwast, Foltz and Stickle1990; Byrne and Anderson, Reference Byrne and Anderson1994). Hybridization occurs among a three-species complex in the extant echinoid Strongylocentrous. It is reasonable to assume that hybrids occurred during the geological past. In addition to the Eretmocrinus example, Nichols (Reference Nichols1959) reported hybrids in the fossil echinoid Micraster. We suggest further, that hybridization in the geological past is a vastly underappreciated phenomenon (see Goodfriend and Gould, Reference Goodfriend and Gould1996).
Genus Macrocrinus Wachsmuth and Springer, Reference Wachsmuth and Springer1897
Type species
Actinocrinus konincki Shumard, Reference Shumard1855.
Other species
Early Viséan species of Macrocrinus are M. casualis, M. mundulus, and M. strotobasilaris. See Ausich and Kammer (Reference Ausich and Kammer2010) for full list of Macrocrinus species.
Diagnosis
Calyx shape medium to low cone or low bowl, or rarely slightly expanding with concave sides; basal concavity absent; calyx as high as or higher than tegmen; calyx plates smooth, convex, or nodose with radial plate and first interradial plate commonly nodose; median ray ridges absent; plate sutures commonly distinct; basal plates low or high, may be proximally expanded by nodes; radial plates high; first primibrachial tetragonal; rays not lobate; regular interrays not in contact with tegmen; CD interray in contact with tegmen; tegmen flat to low inverted bowl, flat to low cone, or flat to low expanding cone with concave sides; tegmen plates smooth, convex, nodose, or spinose; anal tube cylindrical, may be curved or otherwise asymmetrical; arm facets face outward (subvertical); free arms 12–20, unbranched, rarely spatulate distally (from Ausich and Kammer, Reference Ausich and Kammer2010).
Occurrence
Macrocrinus occurs in the Tournaisian to early Viséan (Kinderhookian to late Osagean) of North America.
Remarks
Species characters in Macrocrinus are calyx shape, height of calyx relative to the height of the tegmen, relative height of basal circlet, tegmen shape, anal tube plate sculpturing, relative sizes of anal tube plates, and orientation of arm facets.
Macrocrinus casualis (Miller and Gurley, Reference Miller and Gurley1895a)

Figure 9 Species of Macrocrinus from the Fort Payne Formation. (1, 2) Macrocrinus mundulus; E-ray lateral view and oral views of theca, respectively (USNM 639939). (3, 4) Macrocrinus strotobasilaris; lateral view and oral views of thecae, respectively (USNM 639940). (5, 6) Macrocrinus casualis; oral and lateral views of theca in matrix, respectively (USNM 639933). (7) Macrocrinus casualis with anal tube preserved (USNM 639935a). Scale bar same for all.
1895a Batocrinus casualis Reference Miller and GurleyMiller and Gurley, p. 15, pl. 1, fig. 17.
1896c Batocrinus rudis Reference Miller and GurleyMiller and Gurley, p. 52, pl. 4, fig. 4.
2013 Macrocrinus casualis (Miller and Gurley); Reference Webster and WebsterWebster and Webster, p. 1782.
2013 Macrocrinus rudis (Miller and Gurley); Reference Webster and WebsterWebster and Webster, p. 1785.
Complete synonymy list in Supplemental Data 2.
Holotype
FMNH UC 6498.
Diagnosis
Calyx low to medium cone, higher than tegmen; basals high; tegmen shape low inverted cone with few large plates with large nodes; arm facets subvertical.
Occurrence
The holotype of Macrocrinus casualis was described from what was most probably the Fort Payne Formation at an undisclosed locality in Tennessee. In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, this taxon is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, and Gross Creek; wackestone buildup facies at Owens Branch; and sheetlike packstone facies at Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, and 61DW.
Description
Calyx low to medium cone shaped, height to width ratio 0.51–0.95 (mean=0.73), widest at arm openings, slight concavity where basals meet radials, straight from base of radials to arm openings, outline of calyx at arm openings circular (Fig. 9.5).
Basal rim formed from elongated transverse nodes of basal plates that extend horizontally, sutures flush with surrounding plates. Plate sculpturing variable; types include smooth, transverse nodes on all plates, central nodes on all plates, and any combination of the above. Basal circlet 18–45% of calyx height (mean=29%). Basal plates three, equal in size, sculpture as noted above. Radial circlet 27–53% of calyx height (mean=40%). Radial plates five, hexagonal, generally wider than high, although a few specimens have radial plates higher than wide. Sculpture variable. Regular interrays not in contact with tegmen, consists of a single plate. Interradial plate slightly higher than wide, hexagonal.
Primanal hexagonal, slightly higher than wide; plating P-3-3-1, in contact with tegmen.
First primibrachial wider than high; slightly smaller than second primibrachial, second primibrachial axillary, additional fixed brachials wider than high, two secundibrachials, last fixed brachials in tertibrachitaxis, free arm facets subvertical.
Tegmen very low inverted cone from arm openings to base of anal tube, with few plates leading to anal tube, tegmen plates with small spines (Fig. 9.6), anal tube shape tapering inverted cone, ~2.0 times higher than the calyx, comprised of large plates with a central node.
Free arms 14–17, distal arms unknown in Fort Payne Formation.
Materials
The holotype of E. casualis Miller and Gurley is FMNH UC 6498, and the holotype of Batocrinus rudis Miller and Gurley, Reference Miller and Gurley1896c, a junior synonym, is FMNH UC 8777. The following are new Fort Payne Formation specimens from this study USNM 639933–USNM 639937, OSU 54506–54521, and CMC IP76376–CMC IP76404.
Measurements
See Supplemental Table 6.
Remarks
Macrocrinus casualis has a low to medium cone-shaped calyx; basals high; tegmen shape low inverted cone with few large plates with large nodes; arm facets subvertical. In contrast Fort Payne species are characterized as follows: M. mundulus with a medium cone-shaped calyx, basals high, tegmen shape flat inverted cone with many small plates with small nodes, arm facets subvertical; and M. strotobasilaris with a medium cone-shaped calyx, higher than tegmen, basals low, tegmen shape flat inverted cone with few large plates with medium nodes, arm facets subhorizontal.
Macrocrinus mundulus (Hall, Reference Hall1859a)
1859a Actinocrinus mundulus Hall, p. 39.
1859a Actinocrinus similis Hall, p. 40.
1859a Actinocrinus lagunculus Hall, p. 41.
1890 Batocrinus jucundus Miller and Gurley, p. 20, pl. 4, figs. 5, 6.
1891 Actinocrinus agnatus Reference MillerMiller, p. 53, pl. 8, figs.1, 2.
1895b Batocrinus procerus Miller and Gurley, p. 33, pl. 1, figs. 40–42
1897 Macrocrinus jucundus (Miller and Gurley); Reference Wachsmuth and SpringerWachsmuth and Springer, p. 451, pl. 4, fig. 15, pl. 30, figs. 13, 14.
1897 Batocrinus mundulus (Hall); Reference Wachsmuth and SpringerWachsmuth and Springer, p. 382, pl. 30, figs. 4a–5.
1897 Macrocrinus lagunculus (Hall); Reference Wachsmuth and SpringerWachsmuth and Springer, p. 453, pl. 35, fig. 4.
1958 Macrocrinus mundulus (Hall); Lane, p. 204, pl. 7, fig. 3, pl. 8, fig. 4, pl. 9, fig. 9.
1964 Macrocrinus mundulus (Hall); Van Sant, p. 117, pl. 8, figs. 6, 13.
1978 Macrocrinus jucundus (Miller and Gurley); N.G. Lane, p. T471, fig. 276.2a, b.
1991b Macrocrinus mundulus (Hall); Reference Ausich and KammerAusich and Kammer, p. 936, figs. 1.1–1.3.
1999 Macrocrinus mundulus (Hall); Reference AusichAusich in Hess et al., figs. 158, 161.
2013 Macrocrinus mundulus (Hall); Reference Webster and WebsterWebster and Webster, p. 1784.
Complete synonymy list in Supplemental Data 2.
Holotype
IGS 1848.
Diagnosis
Calyx medium cone shaped, higher than tegmen; basals high; tegmen shape flat inverted cone with many small plates with small nodes; arm facets subvertical.
Occurrence
Macrocrinus mundulus was originally described from the Keokuk Limestone at Warsaw, Illinois. This species is now recognized from the following early Viséan formations: Keokuk Limestone at Warsaw and Nauvoo, Illinois and Keokuk, Iowa; the Edwardsville Formation in Monroe County (Allens Creek Bank, Boy Scout Camp, and Waldrip Site) and Montgomery County (Indian Creek and Crawfordsville upper quarry), Indiana; the upper Warsaw Formation in Missouri; the Harrodsburg Limestone in Indiana the Muldraugh Member of the Borden Formation (Hardin County, Kentucky).
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Macrocrinus mundulus is known from sheetlike packstone facies at Wolf Creek/Caney Fork Confluence and 61DW.
Description
Calyx medium cone shaped, height to width ratio 0.62–0.86 (mean=0.75), widest at arm openings, slight concavity where basals meet radials, straight from base of radials to arm openings, outline of calyx at level of arm openings circular (Fig. 9.1).
Basal rim formed from elongated transverse nodes of basal plates that extend horizontally, sutures flush with surrounding plates. Plate sculpturing variable: types include smooth, transverse nodes only discernable on radials, and transverse nodes discernable on all calyx plates. Basal circlet 21–29% of calyx height. Basal plates three, equal in size, sculpture as noted above. Radial circlet 36–41% of calyx height. Radial plates five, hexagonal, wider than high, sculpture as noted above. Regular interrays not in contact with tegmen; consists of a single plate. Interradial higher than wide, hexagonal.
Primanal septagonal, slightly wider than high; plating P-3-2-1, in contact with tegmen.
First primibrachial wider than high, approximately equal in size to second primibrachial; second primibrachial axillary; additional fixed brachials wider than high; two secundibrachials; last fixed brachials in tertibrachitaxis, free arm facets subvertical.
Tegmen low inverted cone from arm openings to base of anal tube, with many small plates leading to anal tube (Fig. 9.2); tegmen plates nodose; anal tube shape unknown from Fort Payne material.
Free arms 14–15; distal arms unknown from Fort Payne material.
Materials
The holotype of Actinocrinus mundulus Hall, Reference Hall1859a is IGS 1848. The type specimens of junior synonyms are Actinocrinus agnatus Miller, Reference Miller1892a, holotype, USNM S 795; Batocrinus jucundus Miller and Gurley, Reference Miller and Gurley1890a, syntypes, FMNH UC 6259; Actinocrinus lagunculus Hall, Reference Hall1859a, holotype, UI X-815, referred specimen, USNM S 798; Batocrinus procerus Miller and Gurley, Reference Miller and Gurley1895b, holotype, CMC IP 3238; and Batocrinus similis Hall, Reference Hall1859a, syntype, UI X 1104.
The following are new Fort Payne Formation specimens from this study USNM 639938–USNM 639939, and CMC IP 76405.
Measurements
See Supplemental Table 7.
Remarks
For comparison to other Fort Payne species of Macrocrinus, see remarks of E. casualis.
Macrocrinus strotobasilaris Ausich and Lane, Reference Ausich and Lane1982
1982 Macrocrinus strotobasilaris Ausich and Lane, p. 1355, pl. 2, figs. 1–8; fig. 3.
2013 Macrocrinus strotobasilaris Ausich and Lane; Reference Webster and WebsterWebster and Webster, p. 1786.
Complete synonymy list in Supplemental Data 2.
Types
The holotype is IU 15126-221.
Diagnosis
Calyx medium cone shaped, higher than tegmen; basals low; tegmen shape flat inverted cone with few large plates with medium nodes; arm facets subhorizontal.
Occurrence
The holotype of Macrocrinus strotobasilaris was described from the Edwardsville Formation, Monroe County, Indiana, where it occurred on the Allens Creek delta platform carbonate packstone bank and other delta-platform facies. In the Fort Payne Formation of south-central Kentucky, this taxon is known from the sheetlike packstone facies at Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, 61B, and 61RS.
Description
Calyx medium cone shape, height to width ratio 0.40–0.80 (mean=0.71), widest at arm openings, slight concavity where basals meet radials, straight from base of radials to arm openings, outline of calyx at level of arm openings circular (Fig. 9.3).
Basal rim formed from elongated transverse nodes of basal plates that extend horizontally; sutures flush with surrounding plates. Plate sculpturing variable; types include smooth and transverse nodes on radial plates and central nodes on other calyx plates. Basal circlet 10–40% of calyx height (mean=24%). Basal plates three, equal in size, sculpture as noted above. Radial circlet 32–52% of calyx height (mean=42%). Radial plates five, hexagonal, generally wider than high with a few higher than wide, sculpture as noted above. Regular interrays not in contact with tegmen, consists of a single plate. Interradial as high as wide, hexagonal.
Primanal septagonal, as high as wide, plating P-3-1, in contact with tegmen.
First primibrachial wider than high, approximately equal in size to second primibrachial; second primibrachial axillary; two secundibrachials, last fixed brachials in tertibrachitaxis; free arm facets subhorizontal.
Tegmen low inverted cone from arm openings to base of anal tube, few plates leading to anal tube; tegmen plates with small central nodes (Fig. 9.4), anal tube shape unknown from Fort Payne material.
Free arms 14–17, commonly 16. Distal arms unknown from Fort Payne material.
Materials
In addition to the holotype, numerous paratypes were designated (Ausich and Lane, Reference Ausich and Lane1982). The following are new Fort Payne Formation specimens from this study USNM 639940–USNM 639944, OSU 54522–OSU 54527, and CMC IP76406–CMC IP76408.
Measurements
See Supplemental Table 7.
Remarks
For comparison to other Fort Payne species of Macrocrinus, see remarks of E. casualis.
Genus Magnuscrinus Ausich and Kammer, Reference Ausich and Kammer2010
Type species
Magnuscrinus yandelli (Shumard, Reference Shumard1858).
Other species
All species of Magnuscrinus are early Viséan and include Magnuscrinus cumberlandensis n. sp.; Magnuscrinus kammeri Krivicich et al., Reference Krivicich, Ausich and Keyes2013; Magnuscrinus praegravis (Miller, Reference Miller1892a); and Magnuscrinus yandelli (Shumard, Reference Shumard1858).
Diagnosis.—Calyx shape low bowl or flat cone; basal concavity absent; calyx lower than tegmen; calyx plates very nodose; median ray ridges absent; plates commonly with distinct sutures; basal plates low, with large elongate proximal nodes; radial plates low; first primibrachial tetragonal; rays lobate; regular interrays in contact with tegmen (although may be variable on a single specimen); CD interray in contact with tegmen; tegmen low to medium inverted cone; tegmen plates very nodose or spinose; anal tube cylindrical; arm facets face outward (subvertical); free arms 12–25, other aspects of free arms unknown (from Ausich and Kammer, Reference Ausich and Kammer2010).
Occurrence
Magnuscrinus is known from the early Viséan (late Osagean) of Indiana, Kentucky, and Tennessee.
Remarks
Ausich and Kammer (Reference Ausich and Kammer2010) placed four species into Magnuscrinus: M. yandelli (Shumard, Reference Shumard1858), M. celsus (Miller and Gurley, Reference Miller and Gurley1894b), M. praegravis (Miller, 1892), and M. spinosus (Miller and Gurley, Reference Miller and Gurley1895a). Since 2010, M. kammeri Krivicich et al., Reference Krivicich, Ausich and Keyes2013 and M. cumberlandensis n. sp. are added to the genus. Given systematic revisions in this contribution, Magnuscrinus celsus is placed in synonymy with Magnuscrinus praegravis, and Magnuscrinus spinosus is reassigned to Eretmocrinus. Consequently, four species of Magnuscrinus are currently recognized, all from the early Viséan: M. cumberlandensis n. sp.; M. kammeri Krivicich et al., Reference Krivicich, Ausich and Keyes2013; M. praegravis (Miller, Reference Miller1892a), and M. yandelli (Shumard, Reference Shumard1858).
Species-level characters diagnostic for species include calyx size, shape of the calyx sides, amount of indentation in the interradial positions around arm periphery, relative sizes of radial plates, and calyx plate sculpturing.
Magnuscrinus yandelli (Shumard, Reference Shumard1858)

Figure 10 Magnuscrinus from the Fort Payne Formation. (1, 2) Magnuscrinus yandelli holotype, basal and CD-interray lateral view, respectively (USNM S 929a); (3, 4) Magnuscrinus kammeri holotype, lateral and basal views, respectively (USNM 546039). Scale bar same for all.
1858 Actinocrinus yandelli Reference ShumardShumard, p. 76, pl. 1, figs. 4a, b.
1895b Batocrinus prodigialis Miller and Gurley, p. 39, pl. 3, figs. 4–6.
1897 Lobocrinus yandelli (Shumard); Reference Wachsmuth and SpringerWachsmuth and Springer, p. 441, pl. 30, figs. 7a, b.
2010 Magnuscrinus yandelli (Shumard); Reference Ausich and KammerAusich and Kammer, p. 47, fig. 2.1–2.3.
2013 Magnuscrinus yandelli (Shumard); Reference Webster and WebsterWebster and Webster, p. 1795.
Complete synonymy list in Supplement Data 2.
Lectotype
USNM S 929a.
Diagnosis
Relatively large in size, very low cone-shaped calyx, interradial positions around arm periphery greatly indented, radial plates relatively low, calyx plates with pronounced elongate nodes that dominate calyx appearance.
Occurrence
The holotype of Magnuscrinus yandelli was described from the New Providence Shale at Button Mold Knob, Bullitt County, Kentucky (Fig. 10.1, 10.2), and it was also described from the “same horizon” at Whites Creek Spring, Davidson County, Tennessee. Laudon (Reference Laudon1957) recorded that M. yandelli from other “Borden bioherms” in Kentucky, but we have not located these collections. No specimens of M. yandelli were collected from the Lake Cumberland area. One specimen, USNM 123338, is labeled as from the Fern Glen Formation (lower Tournaisian) at Fern Glen, Missouri; but this latter occurrence is considered erroneous, herein.
Materials
Ten specimens, USNM S 929, constitute the syntype suite from which a lectotype, USNM S 929a, and paralectotypes, USNM S 929b–j, are designated. Three additional specimens USNM S 930, USNM 777, and USNM 123338 were also examined.
Remarks
Although not known at this time to occur in the Fort Payne Formation, this is the type species for the genus. In addition, this taxon is illustrated (Fig. 10.1, 10.2) because it is known from coeval deep-water, toe-of-slope facies in north-central Kentucky.
For comparison to other species of Magnuscrinus discussed here, see remarks of M. cumberlandensis.
Magnuscrinus kammeri Krivicich, Ausich, and Keyes, Reference Krivicich, Ausich and Keyes2013
2013 Magnuscrinus kammeri Reference Krivicich, Ausich and KeyesKrivicich et al., p. 140, fig. 2A–E.
Holotype
Holotype: USNM 546039.
Diagnosis
Relatively small in size, very low cone-shaped calyx, interradial positions around arm periphery slightly indented, radial plates relatively low, calyx plates with gnarled or highly pustulose sculpturing.
Occurrence
The holotype of Magnuscrinus kammeri is from Lawrence County (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 5), Tennessee, and paratypes were from the Fort Payne Formation at Gross Creek, Russell County, Kentucky and Lawrence County (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 5), Tennessee. All occurrences are early Viséan in age. The Gross Creek locality along Lake Cumberland is an example of the crinoidal packstone buildup facies.
Description
Calyx size relatively small for genus, very low cone shape (Fig. 10.3), base with continuous ridge formed by nodes (Fig. 10.4), shallow basal concavity, arms grouped, but not lobate (Fig. 10.4), calyx plates with numerous small nodes (Fig. 10.4) that may be coalesced into short ridges, plate sutures distinct.
Basals three, wider than high, equal in size, lower than radials. Radials five, hexagonal or heptagonal in shape, wider than high, higher than basals. Primanal hexagonal, larger than radial plates, interrupts radial circlet, second range in the posterior with two much smaller plates; posterior plating P-2-1 or P-2-1-1; posterior interray in contact with tegmen. Normal interrays slightly narrower than CD interray, first interradial 10-sided; plating 1-2, may or may not be in contact with tegmen. First primibrachial tetragonal, much wider than high; second primibrachial axillary, pentagonal, slightly larger than first primibrachial, wider than high; first and second secundibrachials approximately the same size as second primibrachials; last fixed brachial second or third tertibrachial (Fig. 10.4); arm openings subelliptical, higher than wide, directed obliquely upward (from Krivicich et al., Reference Krivicich, Ausich and Keyes2013).
Tegmen low inverted cone shape (Fig. 10.3), plates with very large nodes; anal tube high and slender, central, plates with large elongate nodes. Twenty facets for free arms, other aspects of arms not known. Column unknown.
Materials
Holotype: USNM 546039 (locality 5); paratype: USNM 546040 (locality 5); paratypes: USNM 546041–USNM 546043 (Gross Creek West).
Measurements
USNM 546039 (holotype): calyx height, 9.3 mm; calyx width, 35.0* mm, tegmen height, 22.0* mm. Paratypes: USNM 546040: calyx height, 10.5 mm; calyx width, 31.5 mm; USNM 546041: calyx height, 13.4 mm; calyx width, 34.8 mm; USNM 546043: calyx height, 9.1* mm; calyx width, 17.5* mm; anal tube height, 37.2* mm.
Remarks
Magnuscrinus kammeri is distinct from all congeners by having unique calyx plate sculpturing, high calyx width to height ratio, and high radial plate width to height ratio. In terms of overall calyx shape, M. kammeri is closest to E. ramulosus.
For comparison to other species of Magnuscrinus discussed here, see remarks of M. cumberlandensis.
Magnuscrinus praegravis Miller, Reference Miller1892a
1849 Actinocrinites cornutus Reference TroostTroost, p. 419 (nomen nudum).
1892b Eretmocrinus prægravis Reference MillerMiller, p. 678.
1892c Eretmocrinus praegravis Reference MillerMiller; Miller, p. 37, pl. 6, figs. 5, 6.
1894b Alloprosallocrinus celsus Miller and Gurley, p. 47, pl. 4, figs. 9–11.
1958 Eretmocrinus praegravis Miller; Lane, p. 173.
2013 Magnuscrinus praegravis Miller; Reference Webster and WebsterWebster and Webster, p. 1795.
Complete synonymy list in Supplemental Data 2.
Holotype
USNM S 5747.
Diagnosis
Relatively small in size, cone-shaped calyx, interradial positions around arm periphery not indented, radial plates relatively low, calyx plates with pronounced commonly elongate nodes that dominate the central part or the entire plate.
Occurrence
The holotype of Magnuscrinus praegravis was described from Pilot Knob, near Louisville, Kentucky (exact location and stratigraphic horizon not known). The type specimens of junior synonyms are from the following: Actinocrinites cornutus holotype: Cannon Co., Tennessee; and Alloprosallocrinus celsus: “Warsaw Group” Tennessee, both presumed to be from the Fort Payne Formation.
Materials
In addition to the holotype (USNM S5747), the holotype of the junior synonym, Actinocrinites cornutus is USNM 39891. The holotype for Alloprosallocrinus celsus has not been located.
Remarks
No specimens of M. praegravis were collected from the Fort Payne Formation of south-central Kentucky, Tennessee, or north-central Alabama during this study.
For comparison to other species of Magnuscrinus discussed here, see remarks of M. cumberlandensis.
Magnuscrinus cumberlandensis new species

Figure 11 Magnuscrinus and Uperocrinus from the Fort Payne Formation. (1–4) Magnuscrinus cumberlandensis n. sp.; (1) paratype, basal view of partial large theca in matrix, note some calyx plates are missing, but plate sutures outlined by silicification (USNM 639946); (2–4) holotype, of partially compressed, medium-sized specimen; oral, lateral, and basal views, respectively (USNM 639945). (5, 6) Uperocrinus nashvillae; oral and E-ray lateral views, respectively, note orientation of arm facets upward (USNM 639952). (7, 8) Uperocrinus robustus; B-ray lateral and oral views, respectively (USNM 639955). Scale bars, as indicated.
2014 Abatocrinus n. sp. Krivicich et al., p. 1155.
Holotype
USNM 639945.
Diagnosis
Relatively large in size, very low vase-shaped calyx, interradial positions around arm periphery slightly indented, radial plates relatively high, calyx plates with broad, circular to elongate nodes.
Occurrence
All specimens are from early Viséan strata along the shores of Lake Cumberland. The holotype is from the Gross Creek carbonate packstone buildup. Other localities include Cave Springs South (carbonate packstone buildup facies), Seventy-Six Falls (allochthonous sheetlike packstone facies), and Rowena wackestone buildup facies.
Description
Calyx large, very low vase shaped, greatly expanded at level of arm openings, straight-sided from base to position where expansion begins in primibrachitaxis; plates on aboral cup with broad, circular or elongate nodes; arms grouped slightly into rays at level of arm openings (Fig. 11.1, 11.3).
Basal circlet truncate proximally, high, ~20% of cup height, very shallow basal concavity formed by nodosity of basal plates; basal-basal sutures indented; basal plates three, equal in size, transverse nodes on basal plates forming basal portion of calyx (Fig. 11.3).
Radial circlet ~25% of calyx height; radial plates slightly wider than high, five, hexagonal, ~1.5 times wider than high, supporting the first primibrachial above and an interradial laterally above on each side. In regular interrays a single large interradial plate, octagonal, a large elongate node in center, higher than wide, not in contact with tegmen, smaller than radial plates, concave in profile because situated at position of calyx where expansion to arm openings begins.
Primanal heptagonal, slightly higher than wide, equal in width to adjacent radials, higher than adjacent radials, interrupts radial circlet; large, central, slightly elongate node; three plates in second range; third range not preserved.
Fixed brachials gently convex. First primibrachial wider than high; second primibrachial axillary; second secundibrachial axillary; three fixed tertibrachials, as known.
Tegmen large, flat inverted cone from arm openings to base of anal tube, height of anal tube not known; tegmen plates large with central circular nodes or spines (Fig. 11.2).
Free arms as many as 40, but ~20 in juveniles; facets large, vertical. Characteristics of free arms not known.
Proximal column circular with a pentalobate lumen; other details of column not known.
Etymology
The trivial name for this species acknowledges the area of origin of this species, Lake Cumberland, in south-central Kentucky.
Materials
Seven specimens of Magnuscrinus cumberlandensis n. sp. have been collected from the southern Lake Cumberland area. A fairly complete sub-adult specimen (USNM 639945) is designated as the holotype, and the six paratypes are USNM 639946–639951. USNM 639946 is partially preserved adult specimen, and USNM 639948–639950 are partially preserved juveniles.
Measurements
USNM 639945, holotype: CaH, 14.9 mm; CaW, 46.8 mm; proximal BW, 11.2 mm; TH, 12.6* mm. USNM 639946, paratype: CaH, 23.5 mm; CaW, 86.7 mm; proximal BW, 10.5* mm.
Remarks
Magnuscrinus cumberlandensis n. sp. is unique among Fort Payne Formation batocrinids in the considerable flattening and expansion of the calyx at the level of the arm openings. The calyx diameter at the arm openings is nearly three times the diameter of the calyx at the level of the radials. In contrast, in A. steropes the diameter at the arm openings is only 1.5 times greater than that at the radials.
Two basic sizes of specimens are assigned to M. cumberlandensis n. sp. The smaller specimens have a calyx diameter of 5–6 cm, whereas the larger have a diameter that is 28% larger (as much as 8.5 cm). These specimens also differ in the height of the aboral cup (higher in larger specimens), robustness of radial plate sculpturing (more robust in larger specimens), thickness of calyx plates (thicker in larger specimens), and in the number of free arms (more in larger specimens). These two morphotypes either represent juvenile versus adult specimens of a single species or two different species. The basic overall geometry of M. cumberlandensis n. sp., unique among early Viséan batocrinids, is similar for these two morphotypes, and these two morphologies are regarded as ontogenetic variation.
The unique morphology of this new taxon makes its generic assignment a challenge. With the revised diagnosis of the Batocrinidae (Ausich and Kammer, Reference Ausich and Kammer2010), the combination of a greatly expanded tegmen at the position of the arm facets, slightly grouped arms, and plating in all interrays in contact with the tegmen ally this species with Magnuscrinus.
Extensive calyx flattening and expansion at the level of arm openings is an iterative convergent design that developed in Mississippian crinoids, including other batocrinids (e.g., Eutrochocrinus), and actinocrinitids (e.g., Strotocrinus). In comparison to these other examples, M. cumberlandensis n. sp. has a much shorter calyx with expansion developed on more proximal plates.
Magnuscrinus cumberlandensis n. sp. is relatively large in size, with a very low vase-shaped calyx; interradial positions around arm periphery slightly indented; radial plates relatively high; and calyx plates with broad, circular to elongate nodes. Other species of Magnuscrinus have the following characters: M. yandelli, relatively large in size, very low cone-shaped calyx, interradial positions around arm periphery greatly indented, radial plates relatively low, calyx plates with pronounced elongate nodes that dominate calyx appearance; M. kammeri is relatively small in size, very low cone-shaped calyx, interradial positions around arm periphery slightly indented, radial plates relatively low, calyx plates with gnarled or highly pustulose sculpturing; and M. praegravis relatively small in size, cone-shaped calyx, interradial positions around arm periphery not indented, radial plates relatively low, calyx plates with pronounced commonly elongate nodes that dominate the central part or the entire plate.
Genus Uperocrinus Meek and Worthen, Reference Meek and Worthen1865
Type species
Actinocrinites pyriformis Shumard, Reference Shumard1855; by original designation.
Other species
Early Viséan species of Uperocrinus include U. marinus (Miller and Gurley, Reference Miller and Gurley1890), U. nashvillae, and U. robustus. Uperocrinus marinus is confined to the Edwardsville Formation of Montgomery County, Indiana; and U. robustus is only recognized from the Fort Payne Formation, as detailed below. Uperocrinus nashvillae is more geographically widespread, as noted below. See Ausich and Kammer (Reference Ausich and Kammer2010) for full list of Uperocrinus species.
Diagnosis
Calyx shape expanding with concave sides or low to medium cone shaped; basal concavity absent; calyx as high as or higher than tegmen; calyx plates smooth or rarely nodose; median ray ridges absent; plate sutures commonly distinct; basal plates high (rarely low), truncate or with a proximal expansion; radial plates high; first primibrachial tetragonal; rays lobate, slightly to well developed; regular interrays typically in contact with tegmen (but rarely and primitively not in contact); CD interray in contact with tegmen; tegmen flat to low inverted cone or low inverted bowl; tegmen plates smooth, convex, or nodose; anal tube cylindrical; arm facets face upward (<30˚) or obliquely upward (30˚–60˚); free arms 14–20, unbranched, not spatulate distally (from Ausich and Kammer, Reference Ausich and Kammer2010).
Occurrence
Uperocrinus is known only from the late Tournaisian to early Viséan (early to late Osagean) of North America.
Remarks
Characters used to diagnose species of Uperocrinus are width of basal sutures relative to radial plates, relative number of many plates in the regular and posterior interrays, intraray plating between halfrays, and orientation of arm facets.
Uperocrinus nashvillae (Hall, Reference Hall1858)
1849 Actinocrinites Nashvillæ Reference TroostTroost, p. 419 (nomen nudum).
1858 Actinocrinus nashvillæ, Reference HallHall, p. 609, pl. 15, fig. 4, pl. 6, figs. 4a, b, fig. 92.
1897 Actinocrinites Nashvillæ (Troost); Reference Wachsmuth and SpringerWachsmuth and Springer, p. 435, pl. 31, fig. 1.
1958 Uperocrinus nashvillae (Troost); Lane, p. 235, pl. 7, fig. 2.
1978a Uperocrinus nashvillae (Troost); Reference UbaghsUbaghs, fig. 155.1.
2013 Uperocrinus nashvillae (Hall, Reference Hall1858); Reference Webster and WebsterWebster and Webster, p. 2581.
Complete synonymy list in Supplemental Data 2.
Holotype
The holotype of Uperocrinus nashvillae is UI X-1207.
Diagnosis
Basal sutures flush with surrounding plates; many plates in regular and posterior interrays; intraray plate between half-rays; arm facets horizontal.
Occurrence
The holotype of Uperocrinus nashvillae was described from the Keokuk Limestone, Warsaw, Hancock County, Illinois. This species is now recognized from the following early Viséan formations: (1) Fort Payne Formation in Clinton, Cumberland, and Russell counties, Kentucky; Davidson (Whites Creek Springs), Lawrence (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Localities 5 and 6), and Pickett counties, Tennessee; and Limestone (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 3) and Madison (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Locality 2) counties, Alabama; and (3) the Keokuk Limestone at Hamilton, Nauvoo, and Warsaw, Hancock County, Illinois, and at Keokuk, Lee County, Iowa.
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Uperocrinus nashvillae is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, Gross Creek, and Gross Creek West; wackestone buildup facies at Owens Branch; sheet-form packstone facies at Cove Creek, Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, 61D, 61DW, and 61RS.
Description
Calyx medium cone shape, height to width ratio 0.63–1.0 (mean=0.86), widest at arm openings, concave from basals to arm openings (Fig. 11.6), outline of calyx at level of arm openings moderately pentalobate.
Basal rim formed from elongated transverse nodes of basal plate that extend horizontally; sutures between basals and all other plates flush. Plate sculpturing smooth. Basal circlet 15–39% of calyx height (mean=24%). Basal plates three, equal in size, sculpture as noted above. Radial circlet 23–36% of calyx height (mean=28%). Radials five, wider than high, sculpture as noted above. Regular interrays in contact with tegmen, plating 1-2-2-2; first interradial plate slightly higher than wide, shape commonly hexagonal, rarely septangonal.
Primanal septagonal, as wider than high, plating commonly P-3-3-2, rarely P-3-5-4, in contact with tegmen.
First primibrachial wider than high, approximately equal in size to second primibrachial; second primibrachial axillary; two secundibrachials, last fixed brachials in tertibrachitaxis; intraradial plate between half-rays, higher than wide, hexagonal in shape; free arm facets horizontal.
Tegmen very low inverted cone from arm openings to base of anal tube, plates nodose, anal tube shape unknown from Fort Payne material (Fig. 11.5).
Free arms commonly 20, distal arms unknown from Fort Payne material.
Measurements
See Supplemental Table 8.
Materials
Troost’s specimen that would have been a type is USNM 39894. The following are new Fort Payne Formation specimens from this study USNM 639952–USNM 639954, OSU 54528–OSU 54539, and CMC IP76409–CMC IP76418.
Remarks
Uperocrinus nashvillae is characterized by basal sutures flush with surrounding plates; many plates in regular and posterior interrays; intraray plate between half-rays; arm facets horizontal. This contrasts with U. robustus, which has basal sutures impressed between basals, flush with radials; fewer plates in regular and posterior interrays; no intraray plate between halfrays; arm facets subvertical.
Uperocrinus robustus (Wachsmuth and Springer, Reference Wachsmuth and Springer1897)
1850b Actinocrinites Holbrooki Reference TroostTroost, p. 29, fig. 33 (nomen nudum).
1897 Lobocrinus robustus Reference Wachsmuth and SpringerWachsmuth and Springer, p. 436, pl. 30, figs. 8a, b.
1958 Uperocrinus robustus (Wachsmuth and Springer); Lane, p. 237.
2013 Uperocrinus robustus (Wachsmuth and Springer); Reference Webster and WebsterWebster and Webster, p. 2583.
Complete synonymy list in Supplemental Data 2.
Holotype
USNM S 908.
Diagnosis
Basal sutures impressed between basals, flush with radials; fewer plates in regular and posterior interrays; no intraray plate between half-rays; arm facets subvertical.
Occurrence
The holotype of Uperocrinus robustus was described from the Fort Payne Formation at Whites Creek Spring, Davidson County, Tennessee. The provenance of the Actinocrinites holbrooki is not known, but presumably also from Whites Creek Springs.
This species is now recognized from only the Fort Payne Formation (early Viséan), where it is known from Clinton, Cumberland, and Russell counties, Kentucky, and Davidson (Whites Creek Springs), Lawrence (Krivicich et al., Reference Krivicich, Ausich and Keyes2013, Localities 5 and 6), and Pickett counties (Cove Creek), Tennessee.
In the Fort Payne Formation of south-central Kentucky and north-central Tennessee, Uperocrinus robustus is known from the following facies: crinoidal packstone buildup facies at Cave Springs North, Cave Springs South, Gross Creek, and Gross Creek West; wackestone buildup facies at Lily Creek and Owens Creek; and sheet-form packstone facies at Cove Creek, Seventy-Six Falls, Wolf Creek/Caney Fork Confluence, and 61RS.
Description
Calyx medium cone shape, height to width ratio 0.33–0.80 (mean=0.56) (Fig. 11.7), widest at arm openings, straight from basals to arm openings, outline of calyx at level of arm openings strongly pentalobate.
Basal rim formed from elongated transverse nodes of basal plate that extend horizontally; sutures between basals impressed in groove, sutures with radials flush or with slight impression. Plate sculpturing includes faint central nodes on radials with other calyx plates smooth. Basal circlet 17–25% of calyx height (mean=21%). Basal plates three, equal in size, sculpture as noted above. Radial circlet 28–51% of calyx height (mean=38%). Radial plates five, wider than high, sculpture as noted above. Regular interrays in contact with tegmen, plating 1-1-1, first interradial plate higher than wide, shape variable.
Primanal septagonal, wider than high, plating P-3-2-1, in contact with tegmen.
First primibrachial wider than high, approximately equal in size to second primibrachial; second primibrachial axillary; two secundibrachials, last fixed brachials in tertitaxis; free arm facets subvertical.
Tegmen very low inverted cone from arm openings to base of anal tube, plates with central spine (Fig. 11.8), anal tube shape unknown from Fort Payne material.
Free arms commonly 20, distal arms unknown from Fort Payne material.
Materials
The type specimen that would have been the type of Actinocrinites holbrooki Troost, Reference Troost1850b is USNM 39901. The following are new Fort Payne Formation specimens from this study: USNM 639955–USNM 639957, OSU 54540–OSU54542, and CMC IP76419–CMC IP76421.
Measurements
See Supplemental Table 9.
Remarks
For comparison with the other Uperocrinus in the Fort Payne Formation discussed here, see remarks of U. nashvillae.
Summary of systematic and nomenclatoral changes
Revisions resulting from this study include the following: (1) Batocrinus honorabilis Miller and Gurley, Reference Miller and Gurley1895a and Batocrinus casula Miller and Gurley, Reference Miller and Gurley1895a are designated junior synonyms of Abatocrinus grandis (Lyon and Casseday, Reference Lyon and Casseday1859); (2) Batocrinus springeranus Bassler, Reference Bassler1925 is designated a junior synonym of Abatocrinus steropes (Hall, Reference Hall1859a); (3) Agaricocrinus? depressus Casseday and Lyon, Reference Casseday and Lyon1862 is designated a junior synonym of Alloprosallocrinus conicus Casseday and Lyon, Reference Casseday and Lyon1862; (4) Batocrinus commendabilis Miller and Gurley, Reference Miller and Gurley1895a and Batocrinus wetherbyi Miller and Gurley, Reference Miller and Gurley1895a are designated junior synonyms of Eretmocrinus magnificus (Lyon and Casseday, Reference Lyon and Casseday1859); (5) Batocrinus curiosus Miller and Gurley, Reference Miller and Gurley1895a, Batocrinus laterna Miller and Gurley, Reference Miller and Gurley1895a, and Eretmocrinus lyonanus Miller, Reference Miller1891 are designated as junior synonyms of Eretmocrinus ramulosus (Hall, Reference Hall1858); (6) Magnuscrinus spinosus is reassigned to Eretmocrinus; (7) the hybrid specimens previously recognized as Eretmocrinus magnificus×E. praegravis should now be recognized as Eretmocrinus magnificus×E. spinosus, and Batocrinus laciniosus (Miller and Gurley, Reference Miller and Gurley1895a) is now considered to represent this hybrid; (8) Batocrinus rudis Miller and Gurley, Reference Miller and Gurley1896c is designated a junior synonym of Macrocrinus casualis (Miller and Gurley, Reference Miller and Gurley1895a); (9) Alloprosalloscrinus celsus Miller and Gurley, Reference Miller and Gurley1894b is designated a junior synonym of Magnuscrinus praegravis (Miller, Reference Miller1892a); and (10) finally, Azygocrinus decoris Miller, Reference Miller1892c and Glannearycrinus spergenensis (Miller, Reference Miller1892a) are not considered taxa from the Fort Payne Formation.
Acknowledgments
We thank the following (in alphabetical order) for assistance with curated specimens: D.B. Blake, University of Illinois; D.M. Gnidovec, Orton Geological Museum, The Ohio State University; K. Hollis, U.S. National Museum of Natural History; B. Hunda, Cincinnati Museum Center; P. Mayer and S. Lidgard, Field Museum of Natural History; and R. Norby, Illinois State Geological Survey. Many students from both the University of Cincinnati and Ohio State University helped with fieldwork. T.W. Kammer, F.J. Gahn, and S. Zamora improved an earlier version of this manuscript. Ohio State University and the University of Cincinnati provided partial support for Fort Payne Formation research. This research was also supported by the National Science Foundation (WIA: EAR-8407516, EAR-8706930, EAR-8903486; and DLM: EAR-8407744 and EAR-8705531).
Accessibility of supplemental data
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.c9p26.













