



Introduction
Clam shrimp, or “conchostracans,” are a paraphyletic group of bivalved crustaceans known in fossil records from Devonian to recent times. They are sometimes considered as flagship groups in non-marine aquatic biotopes, with important stratigraphic, paleogeographic, and paleoenvironmental significance due to their rapid evolution, high environmental adaptability, and wide distribution. Carboniferous, Permian, and Mesozoic clam shrimp have been reported from all the present continents (e.g., Raymond, Reference Raymond1946; Kobayashi, Reference Kobayashi1954; Novojilov, Reference Novojilov1957, Reference Novojilov1970; Tasch, Reference Tasch and Moore1969, Reference Tasch1987; Zhang et al., Reference Zhang, Chen and Shen1976; Chen and Shen, Reference Chen and Shen1985; Kozur and Weems, Reference Kozur, Weems and Lucas2010; Ghosh, Reference Ghosh2011; Gallego et al., Reference Gallego, Monferrana, Stigall, Zacaríasa, Hegna, Jiménez, Bittencourt, Li and Barrios Calathaki2020).
In the last century, researchers have described the “clam shrimp” of the Cambrian and Ordovician (Ulrich and Bassler, Reference Ulrich and Bassler1931; Soot-Ryen, Reference Soot-Ryen1960; Howell, Reference Howell1963). However, all the records were proven to be of non-clam shrimp affinity (mollusks or stem-group crustaceans; see details in the discussion section). In other words, there are no credible records of any clam shrimp taxon prior to the Devonian. The Devonian saw the emergence and subsequent diversification of the early clam shrimp. Clam shrimp of the Devonian, including >20 genera, were mainly found in Europe (especially the Old Red Sandstone area), China, Central Asia (Kazakhstan), and North Asia (Siberia and adjacent areas in Russia). The earliest clam shrimp are several species from the Early Devonian: Belgolimnadiopsis stockmansi Maillieux, Reference Maillieux1939 (see also in Defretin, Reference Defretin1950; Cuvelier et al., Reference Cuvelier, Hennion, Gallego and Vachard2015); Concherisma eifelense Raymond, Reference Raymond1946; Pseudestheria (Estheria) diensti Gross, Reference Gross1934; and Pseudestheria (Tuvinopsis) arduennae Novojilov, Reference Novojilov1961. It is difficult to determine which one is the oldest because none of them is dated precisely (Hegna and Astrop, Reference Hegna and Astrop2020). Moreover, their morphological descriptions and illustrations are relatively ambiguous and imprecise. Further studies on their stratigraphic placement and detailed taxonomic information are needed.
Most fossil clam shrimp are comprised of carapaces only. Important features of their crustacean body and appendages are often missing or poorly preserved in fossil records. As a result, the key characteristics of body and appendages that are crucial in the morphological taxonomy of modern clam shrimp could not be used for morphological comparison between fossil and modern species. It is not known if these Early Devonian forms represent the crown or stem-group of Spinicaudata (the main taxon of clam shrimp that includes the largest number of modern species), although their carapace character (presence of growth lines) indicates that they look like modern spinicaudatans. Limnestheria arda Wright, Reference Wright1920, of the Moscovian (Westphalian, late Carboniferous) from Ireland was considered the oldest-known stem-group spinicaudatan because it possesses two pairs of claspers (Orr and Briggs, Reference Orr and Briggs1999), which is consistent with the characteristics of all modern spinicaudatan males (Hegna and Astrop, Reference Hegna and Astrop2020).
Regardless, discoveries and taxonomic studies of early forms, especially the Early Devonian species, are still essential to broaden our knowledge of the origin and early evolution of clam shrimp. In this study, we discovered and described four species (including one sp. in open nomenclature) of four genera of clam shrimp from the Lower Devonian Yuankou Formation (Emsian) in Jiangyong, Hunan Province, China. They represent the oldest clam shrimp records in China. This discovery indicates that highly diverse clam shrimp communities already appeared in suitable biotopes on Earth at that time and provides clues for the diversification that occurred after the Early Devonian.
Geological setting
The Yuankou Formation, mainly exposed in southwestern Hunan Province and northern Guangxi Province, China, is a set of continental-coastal clastic sediments. Before the 1970s, continental-coastal clastic sediments that were widely distributed in the region were collectively referred to as the “Tiaomajian Formation.” The age of the Tiaomajian Formation was considered to be Eifelian (early Middle Devonian). In the early 1970s, the regional geological survey team of Hunan Province divided the sediments into three members from bottom to top according to lithology and sedimentary rhythm (Bureau of Geology and Mineral Resources of Hunan Province, 1988, 1997). The lowest member of the three is a set of purple-red and gray to white-grayish clastic sediments, which consist of quartz sandstones, quartz sandstones with gravels, and shales (Fig. 1). The fossil plant Zosterophyllum Penhallow, Reference Penhallow1892, was found in the member and its age was considered to be the latest Early Devonian (Wang, Reference Wang1979). Subsequently, Risheng Yu measured the section near the river in southwestern Yuankou Town, Jiangyong County (24.9933°N, 111.0757°E) and named the member “Yuankou Formation” in 1979 (Yu in Bureau of Geology and Mineral Resources of Hunan Province, 1988, 1997). The section in southwestern Yuankou Town unconformably overlies a set of Cambrian slates. Although the top of the section is covered by Quaternary sediments, making its contact with the Tiaomajian Formation invisible, outcrops in other regions indicate that the Yuankou Formation is conformably overlain by the Middle Devonian Tiaomajian Formation (Bureau of Geology and Mineral Resources of Hunan Province, 1988, 1997). Fossil clam shrimp, bivalves, and plants have been found in the lower part of the Tiaomajian Formation (Wang, Reference Wang1979; Bureau of Geology and Mineral Resources of Hunan Province, 1988, 1997; Gao and Wang, Reference Gao and Wang1990). Fossil plants consist of Zosterophyllum cf. Z. yunnanicum Hsü, Reference Hsü1966; Emplectophycus yunnanensis Li and Cai, Reference Li and Cai1978; Hostimella; Aphyllopteris; Psilophylon; and Taeniocrada; similar to the Drepanophycus spinaeformis Göppert, Reference Göppert1852-Zosterophyllum yunnanicum flora, which was widely distributed in the late Early Devonian continental strata in southwestern China (Wang, Reference Wang1979; Gao and Wang, Reference Gao and Wang1990). Zosterophyllum yunnanicum is generally considered an index species of the Emsian in southwestern China (Li and Cai, Reference Li and Cai1977; Wang, Reference Wang1979). Gao and Wang (Reference Gao and Wang1990) studied spores of the Yuankou Formation and concluded that the formation was correlated to the Guijiatun Formation in Yunnan Province and the Nagaoling Formation in Guangxi Province. According to the latest stratigraphic age framework of the Devonian in China, the age of the Guijiatun Formation is latest Pragian to early Emsian, while the age of the Nagaoling Formation is Pragian (Qie et al., Reference Qie, Ma, Xu, Qiao, Liang, Guo, Song, Chen and Lu2019). Therefore, with all the information above, we think that the age of the Yuankou Formation is most likely early Emsian. The formation consists of continental-coastal clastic sediments, which indicate a coastal wetland environment. Most of the fossils, including plants, clam shrimp, and brackish bivalves, were found in very close horizons in the lower part of the section. Wang (Reference Wang1979) mentioned the existence of fossil brachiopods in the section, but the exact horizon and taxonomic information of these brachiopods were not reported. We assume the Devonian clam shrimp inhabited a coastal wetland with seawater sources and influences.
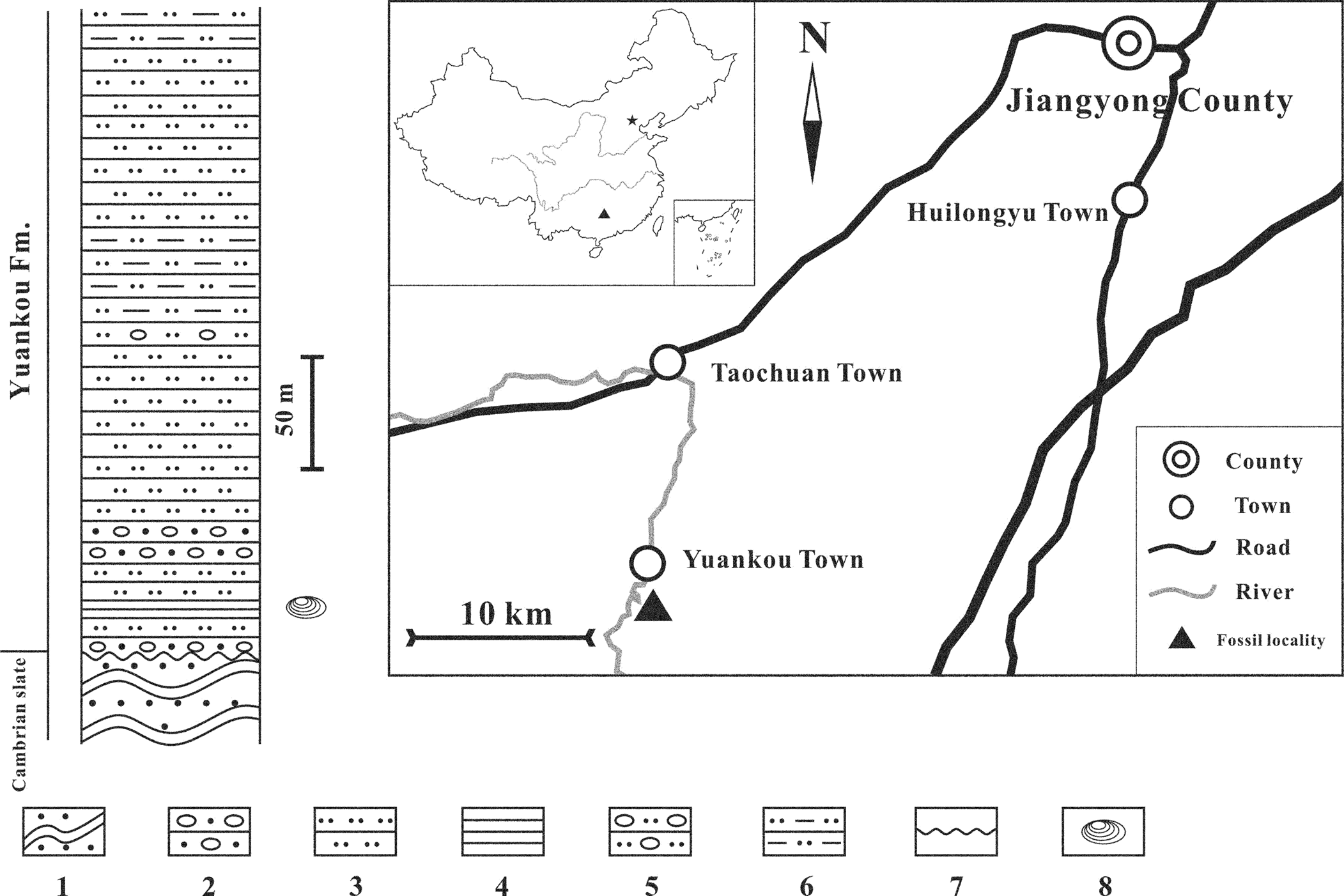
Figure 1. Stratigraphic column and locality of the Yuankou Formation at Yuankou Town, Hunan Province, China. 1. sandy slate; 2. quartz sandstone with gravels; 3. quartz sandstone; 4. shale; 5. Fine-grained quartz sandstones with gravels; 6. muddy quartz sandstones; 7. unconformity; 8. fossil clam shrimp.
Material and methods
Clam shrimp specimens in this study were collected by the second author on November 26, 1979, from the lower part of the Lower Devonian (Emsian) Yuankou Formation in southwestern Yuankou Town, Jiangyong County, Hunan Province, China (Fig. 1). Chen and Shen (Reference Chen and Shen1985) mentioned the presence of these clam shrimp in their published book “An Introduction to Fossil Conchostracans.” However, no further research has been conducted. The outcrop of the Yuankou Formation still exists, but some parts of the section, including the fossil-bearing beds, are covered by thick vegetation. This makes it very difficult to excavate fossils by hand and general tools. As a result, we rely on a very limited number of fossil specimens that were collected tens of years ago. The specimens were preserved in dark-grayish muddy shales. Twenty-one individuals with only carapaces preserved on several rock pieces were collected. All the rock pieces were from the same horizon. Individuals NIGPA174183 (Fig. 2.1) and NIGPA174186 (Fig. 2.4) occur on the same rock piece. Moreover, individuals NIGPA174185 (Fig. 2.3) and NIGPA174189 (Fig. 3.3) occur on the same piece.
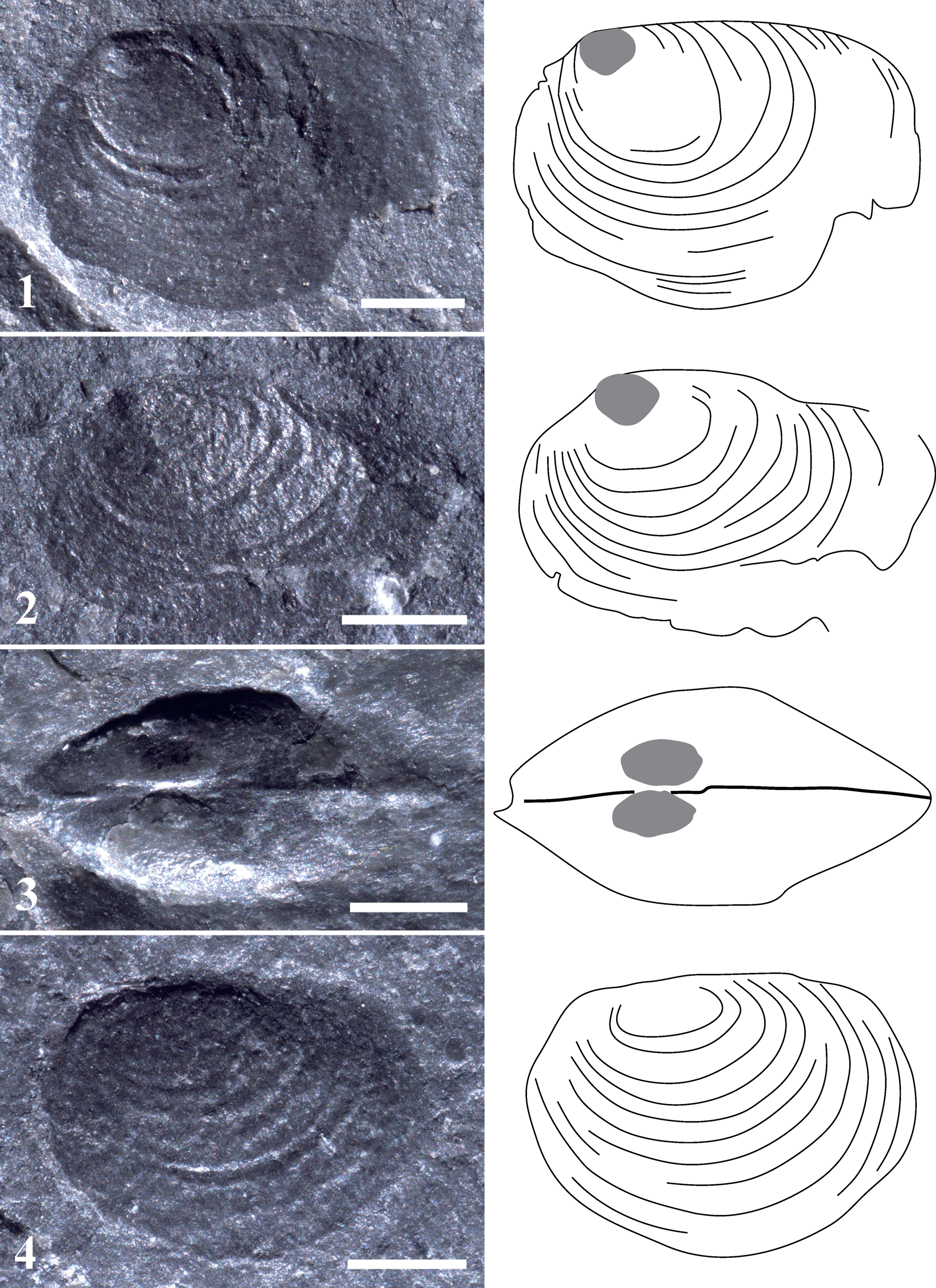
Figure 2. Photographs and sketches of Cornia cheni n. sp. (1–3) and Pseudestheria cf. P. diensti (4). (1) Cornia cheni n. sp., holotype, NIGP174183, left valve, the gray part on the larval valve shows the tubercle; (2) Cornia cheni n. sp., paratype, NIGP174184, left valve, the gray part on the larval valve shows the tubercle; (3) vertical view of the two valves of an individual of Cornia cheni n. sp., paratype, NIGP174185, the gray parts on the larval valve show a pair of tubercles; (4) Pseudestheria cf. P. diensti, NIGP174186, left valve. All scale bars = 1 mm.

Figure 3. Photographs and sketches of Pseudestheria cf. P. diensti (1), Palaeolimnadiopsis zhangi n. sp. (2), and ?Palaeolimnadia sp. (3) (1) Pseudestheria cf. P. diensti, NIGP174187, right valve; (2) Palaeolimnadiopsis zhangi n. sp., holotype, NIGP174188, left valve; (3) ?Palaeolimnadia sp., NIGP174189, right valve. All scale bars = 1 mm.
The fossils were observed and photographed under a Zeiss Discovery V16 microscope. Polarizing coatings were used for observation and photography to eliminate the redundant reflection of light on the surface of the specimens. Considering that the microstructures might be preserved on the surface, a LEO 1530 VP scanning electron microscope was also used for observation. However, no valuable microstructure was found on the surface owing to poor preservation. The invert function of the software Adobe Photoshop 7.0 was used to reverse images taken from the specimens to demonstrate their characteristics more clearly than previously. In the descriptions, we follow the recent classification of spinicaudatans by Martin and Davis (Reference Martin and Davis2001) and the fossil classification by Chen and Shen (Reference Chen and Shen1985), combining the proper terms and suggestions of Chen and Shen (Reference Chen and Shen1985) and Scholze and Schneider (Reference Scholze and Schneider2015).
Repository and institutional abbreviation
Types, figured, and other specimens examined in this study are deposited in Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGPAS), Nanjing, Jiangsu Province, China.
Systematic paleontology
Class Branchiopoda Latreille, Reference Latreille1817
Order Diplostraca Gerstaecker, Reference Gerstaecker and Bronn1866
Suborder Spinicaudata Linder, Reference Linder1945
Family Vertexiidae Kobayashi, Reference Kobayashi1954
Genus Cornia Lutkevich, Reference Lutkevich1937
Type species
Cornia papillaira Lutkevich, Reference Lutkevich1937 from the upper Permian (Lopingian) at Kuznetsk Basin, Siberia, Russia, by original designation.
Cornia cheni new species
Figure 2.1–2.3
Holotype
NIGP174183, a whole carapace from the Lower Devonian (Emsian) Yuankou Formation at Yuankou Town, Hunan Province.
Diagnosis
Carapace small in size, oval in outline; dorsal margin straight and long; larval valve small, bearing an oval node on its top; umbo in anterior and marginal position; anterior margin sharply curved; posterior margin very sharply curved.
Occurrence
Lower part of the Yuankou Formation in southwestern Yuankou Town, Jiangyong County, Hunan Province, China (Fig. 1).
Description
Carapace small in size, oval in outline, 3.2–4.1 mm in length (L), 2.0–2.9 mm in height (H); H/L ratio between 0.63–0.70; dorsal margin straight and long, ratio of dorsal-margin length/carapace length between 0.65–0.70; larval valve small; umbo in anterior and marginal position; an oval or round tubercle (or node) situated on top of the larval valve; anterior margin sharply curved; posterior margin very sharply curved; growth lines fine; 10–12 growth bands, relatively wide and sparsely distributed on the carapace.
Etymology
In memory of Professor Peiji Chen (1936–2019), who contributed significantly to the study of fossil branchiopods of the world and co-authored the book “Fossil Conchostracans of China” in 1976.
Remarks
This species differs from the type species of the genus, C. papillaira from the Upper Permian in Russia (Lutkevich, Reference Lutkevich1937), in its outline, tubercle feature, and position; the latter is round in shape and its tubercle is more like a spine instead of a round node, which is situated at a much lower position in the umbo. The genus Cornia is widely distributed. Species of this genus have often been reported in the Permian–Carboniferous and Triassic continental or continental-marine strata in Europe, China, Africa, North America, South America, India, and Antarctica (see details in Lutkevich, Reference Lutkevich1937; Raymond, Reference Raymond1946; Cardoso, Reference Cardoso1962; Zaspelova, Reference Zaspelova1965; Molin, Reference Molin1968; Novojilov, Reference Novojilov1970; Zhang et al., Reference Zhang, Chen and Shen1976; Kozur, Reference Kozur1983; Tasch, Reference Tasch1987; Kozur and Mock, Reference Kozur, Mock, Lucas and Morales1993; Kozur and Weems, Reference Kozur, Weems and Lucas2010).
Characteristics, such as carapace outline, size, and tubercle feature, among different species are diverse and distinctively different. Some species share similarities in outline and tubercle features with the new species, although the differences are still easy to observe. For example, C. angolata Tasch, Reference Tasch1987, from the Upper Triassic in Angola possesses a very similar outline and tubercle to the new species, but is smaller and possesses fewer growth bands compared to the new species. Cornia laminata Raymond, Reference Raymond1946, from the Permian in the USA also shares similarities in outline and tubercle features with the new species, while its growth bands are also much fewer in number. Based on the diagnosis of the genus, growth bands of corniids were ornamented with punctae or hachures (lirae) (Tasch, Reference Tasch1987). In addition, a specimen identified as “C. papillaira” (Lutkevich, Reference Lutkevich1937) from the Maltsevo Formation in the Babiy Kamen section of Siberia displayed a transition of reticulations and lirae on its carapace (Sabirova et al., Reference Sabirova, Zharinova, Scholze, Schneider and Kogan2019). Unfortunately, no useful ornamentation was found in our specimens due to poor preservation. Moreover, the limited number of our specimens makes it impossible to fully understand both the preservation variability and the intraspecific variability. Regardless, considering that it may be the oldest known Cornia with very significant implications for the origin and early evolution of the genus and correlation of Early Devonian continental strata in southern China, we established this new species. It is noteworthy that differentiating Cornia from Lioestheria Depéret and Mazeran, Reference Depéret and Mazeran1912, may be not easy when the fossils are not well preserved. Lioestheria shows a morphologically variable (conical to pillar-shaped or, especially, oval to elongate-shaped) tubercle on its larval valve. Well-preserved Cornia usually shows a spine-like tubercle on its larval valve. However, the spines on the larval valves of some corniids also could appear to be oval to round when the carapaces are heavily compacted. But it should be noted that the larval valves of lioestheriids are usually larger than one-third of the valves’ length (Holub and Kozur, Reference Holub and Kozur1981; Kozur and Sittig, Reference Kozur and Sittig1981; Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019). Larval valves of this new species are obviously not that large, hence we assign it to Cornia instead of Lioestheria.
Family Euestheriidae Defretin, Reference Defretin1965
Genus Pseudestheria Raymond, Reference Raymond1946
Type species
Pseudestheria brevis Raymond, Reference Raymond1946, from the lower Permian (Cisuralian) Wellington Formation at Noble County, Oklahoma, USA, by original designation.
Remarks
The genus Pseudestheria Raymond, Reference Raymond1946, is one of the most common late Paleozoic genera and has high biostratigraphic value (e.g., Martens, Reference Martens1983; Schneider and Scholze, Reference Schneider and Scholze2018; Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019). Its type species, P. brevis, is oval in outline and possesses a small larval valve with an umbo near the anterior end (Raymond, Reference Raymond1946). The genus diagnosis was amended by Holub and Kozur (Reference Holub and Kozur1981) and Martens (Reference Martens1983). Raymond mentioned that Pseudestheria and Euestheria Depéret and Mazeran, Reference Depéret and Mazeran1912, were much alike and that their difference was mainly in the ornamentation: polygonal reticulations could be observed on the growth bands of the latter under high-power microscopes (Raymond, Reference Raymond1946). He realized that it was impossible in many cases to make definite identifications of the genus, and that the genus inevitably would become a sort of dumping ground for many fossils that were not well preserved (Raymond, Reference Raymond1946). As expected, many poorly preserved fossils were assigned to Pseudestheria in the following decades. In terms of shape and size, Pseudestheria strongly resembles Euestheria, Lioestheria, and Polygrapta Novojilov, Reference Novojilov1946. This has resulted in many taxonomic and biostratigraphic issues and puzzles for researchers, especially in poorly preserved specimens with limited and ambiguous morphological characteristics. Fortunately, some constructive suggestions and distinguishing criteria have been proposed to differentiate Pseudestheria from other similar forms (e.g., Houlb and Kozur, Reference Holub and Kozur1981; Kozur and Sittig, Reference Kozur and Sittig1981; Kozur and Seidel, Reference Kozur and Seidel1983; Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019; for detailed discussions see Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019). However, it should be emphasized that differentiating the genus from other similar forms is still a difficult task when the specimens are poorly preserved. Recently, some studies have shown that different species of Pseudestheria possess different ornamentations on their carapaces. For example, P. novacastrensis Mitchell, Reference Mitchell1927, from the Babiy Kamen section in Siberia, and P. chatangensis Novojilov, Reference Novojilov1946, from the Obnora Formation in Russia, are ornamented with pits (punctae), whereas P. exigua Eichwald, Reference Eichwald1860, from the Monastery Ravine section and Pseudestheria cf. P. itiliana Novojilov, Reference Novojilov1950, from the Cheremushka Ravine section in Russia, are ornamented with reticulations (Zharinova et al., Reference Zharinova, Scholze, Silantiev, Schneider, Nurgaliev, Barclay, Nikolaeva, Silantiev, Zharinova and Vasilyeva2018; Sabirova et al., Reference Sabirova, Zharinova, Scholze, Schneider and Kogan2019).
Pseudestheria cf. P. diensti Gross, Reference Gross1934
Figures 2.4, 3.1
Occurrence
The same as above.
Description
Carapace small in size, oval in outline, 3.3–4.3 mm in length (L), 2.2–2.7 mm in height (H); H/L ratio between 0.63–0.73; dorsal margin straight or slightly arched, long to very long; ratio of dorsal-margin length/carapace length between 0.60–0.81; larval valve very small; umbo in anterior and marginal position; anterior margin sharply curved; posterior margin very sharply curved; 9–12 growth lines; fine growth bands relatively wide and sparsely distributed on the carapace.
Remarks
The specimens of Pseudestheria cf. P. diensti were poorly preserved, with limited visible characteristics: each of them possesses an oval or sub-oval carapace (single valve) with a small umbo, which is situated at the anterior position of the dorsal margin. There are two important characteristics that distinguish this species from Cornia cheni n. sp.: (1) it has no tubercle (or spine) on the larval valve, which is present in Cornia cheni n. sp.; and (2) its larval valve is obviously smaller than that of Cornia cheni n. sp.: ratio h/H (height of the larval valve/total height of the valve) of this species is ~0.2, while ratio h/H of Cornia cheni n. sp. is ≥0.4.
Our specimens share similarities in outline and umbo position with some individuals of P. diensti Gross, Reference Gross1934, from the Lower Devonian of Germany. However, Gross’ specimens possess more growth lines (15–18) than our specimens. In addition, as in Gross (Reference Gross1934), our specimens also show differences in outline among different individuals; some are oval and some are sub-oval to round. We consider that they all belong to one species and that their different shapes may be caused by compression deformation or sexual heteromorphism. Unfortunately, these speculations could not be confirmed due to a limited number of specimens and poor preservation. Further research is needed when better-preserved specimens are collected in the future.
Family Palaeolimnadiopseidae Defretin, Reference Defretin1965
Genus Palaeolimnadiopsis Raymond, Reference Raymond1946
Type species
Palaeolimnadiopsis carpenteri Raymond, Reference Raymond1946, from the lower Permian (Cisuralian) Wellington Formation at Noble County, Oklahoma, USA, by original designation.
Palaeolimnadiopsis zhangi new species
Figure 3.2
Holotype
NIGP174188, a whole carapace from the Lower Devonian (Emsian) Yuankou Formation at Yuankou Town, Hunan Province.
Diagnosis
Carapace small in size, oval in outline; dorsal margin straight to slightly arched and long; larval valve small; umbo in sub-medial and inframarginal position; anterior margin very sharply curved; posterior margin sharply curved; 3–5 growth lines slightly recurved at postero-dorsal area.
Occurrence
The same as above.
Description
Carapace small in size, oval in outline, 4.8 mm in length (L), 3.1 mm in height (H); H/L ratio 0.64; dorsal margin straight to slightly curved and long, ratio of dorsal-margin length/carapace length 0.71; larval valve small; umbo in sub-medial and inframarginal position; 11 fine growth lines; 3–5 growth lines slightly recurved at postero-dorsal area; growth bands wide and sparsely distributed on the carapace.
Etymology
In memory of Professor Wentang Zhang (1924–2013), who contributed significantly to the study of fossil branchiopods of the world and lead-authored the book “Fossil Conchostracans of China” in 1976.
Materials
Holotype: NIGP174188 (Fig. 3.2).
Remarks
This species differs from all the other individuals from the same locality by the presence of ~3–5 growth lines, which slightly recurved at the postero-dorsal area of the carapace, forming a small cape at the postero-dorsal end. Although only one poorly preserved specimen was found, the slight recurvature of growth lines provided a crucial basis for assigning the specimen to the genus Palaeolimnadiopsis. The genus is known from Devonian–Cretaceous (e.g., Webb, Reference Webb1978; Shen, Reference Shen1985; Tasch, Reference Tasch1987; Gallego, Reference Gallego2005; Carbonaro et al., Reference Carbonaro, Rohn and Ghilardi2013).
Unlike the small size of this new form, other species of the genus are usually very large. Some can be extremely large in carapace size (e.g., P. bassi Webb, Reference Webb1978, from the Middle Jurassic Hawkesbury Sandstone in Sydney, Australia is 17.5 mm in length). Moreover, P. sp. from the Upper Triassic Santa Maria Formation in Rio Grande do Sul State, Brazil, and P. carpenteri from the late Early Permian Wellington Formation in Oklahoma, USA both possess astonishingly large carapaces that reach 42.0 mm in length (Tasch, Reference Tasch1987). Palaeolimnadiopsis vileness Varentsov, Reference Varentsov1955, from the Permian/Triassic Moscow Syneclise in Russia, is similar in shape and size to our new species. However, it possesses many more growth bands (up to 20, see details in Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Sennikov, Schneider and Silantiev2015, Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019). It is noteworthy that Chu et al. (Reference Chu, Miao, Wu, Guo, Shu and Tong2018) argued that “two-line typed” growth lines were present on the carapace of P. vilujensis. However, after consulting a large number of taxonomic descriptions and fossil photographs, we found little evidence of this distinctive characteristic in any other specimens of Palaeolimnadiopsis. Based on our observations of the photographs of P. vilujensis in Chu et al. (Reference Chu, Miao, Wu, Guo, Shu and Tong2018), we believe the “two-line typed” growth lines were caused by superposition of two valves when the carapace was preserved as compacted cast. Considering the especially small size and potential of significance of this individual for correlation of continental strata in southern China, we chose to establish this new species, although only one individual was found.
Family Palaleolimnadiidae Tasch, Reference Tasch1956
Genus Palaeolimnadia Raymond, Reference Raymond1946
Type species
Estheria wianamattensis Mitchell, Reference Mitchell1927, from the Triassic Wianamatta Group near Glenlee Homestead on the Great Southern Railway, New South Wales, Australia, by original designation.
?Palaeolimnadia sp.
Figure 3.3
Occurrence
The same as above.
Description
Carapace small in size, oval in outline, ~4.7 mm in length; dorsal margin straight to slightly curved and long, ratio of dorsal-margin length/carapace length ~0.70; larval valve large to very large, no tubercle or rib visible; umbo in sub-medial and inframarginal position; growth lines fine, unknown in number.
Materials
NIGP174189 (Fig. 3.3).
Remarks
The carapace was incomplete (i.e., the ventral and anterior areas of the carapace were missing), making it impossible to count the number of growth lines or measure the height of the carapace. However, its large larval valve without a tubercle or node structure differentiates it from other species of the same locality. There are many forms of clam shrimp that possess large or very large larval valves, including Curvacornutus Tasch, Reference Tasch1961, and Tripemphigus Novojilov in Molin and Novojilov, Reference Molin and Novojilov1965, which possess large larval valves with morphologically variable tubercles (or nodes) (see details in Tasch, Reference Tasch1961; Novojilov, Reference Novojilov1970; Chen and Shen, Reference Chen and Shen1985), while Mesolimnadia Chen, Reference Chen1975, Yunmenglimnadia Chen, Reference Chen1975, and Eolimnadia Chen, Reference Chen1975, possess large larval valves with visible imprints of shell glands (see details in Zhang et al., Reference Zhang, Chen and Shen1976; Chen and Shen, Reference Chen and Shen1985). Larval valves of some species of Palaeolimnadiopsis, Palaeolimnadia, and Asmussia Pacht, Reference Pacht1849, are also relatively large (see details in Lutkevitch, Reference Lutkevich1937; Raymond, Reference Raymond1946; Varentsov, Reference Varentsov1955; Novojilov, Reference Novojilov1970; Zhang et al., Reference Zhang, Chen and Shen1976; Chen and Shen, Reference Chen and Shen1985; Schneider and Scholze, Reference Schneider and Scholze2018; Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019). The species is obviously different from Asmussia membranacea Pacht, Reference Pacht1849, the type species of Asmussia, which was found in the Givetian (Middle Devonian) of Latvia. The ratio h/H of A. membranacea is ~0.4, while we estimate the ratio h/H of this species to be >0.6. In addition, unlike the typical oval outline of this species, A. membranacea is round in outline and possesses a relatively straight dorsal margin, which makes its antero-dorsal and postero-dorsal angles very prominent (observation of characteristics of A. membranacea is based on photographs and sketch from Chen and Shen, Reference Chen and Shen1985). Therefore, the species is unlikely to belong to the genus Asmussia. We could not precisely determine the taxonomic placement of the species given its very limited morphological characteristics. Based on all its visible characteristics, it resembles some paleolimnadids. Thus, we questionably assigned it to the genus ?Palaeolimnadia at this stage, considering its incomplete carapace and poor preservation.
Discussion
To date, there are no credible records of any clam shrimp taxon prior to the Devonian. The small bivalved Cambrian “conchostracan” described by Ulrich and Bassler (Reference Ulrich and Bassler1931) and discussed by Howell (Reference Howell1963) was subsequently shown to be a stem-group crustacean (Bradoriida + Eucrustacea sister to Marellomorpha [Hou et al., Reference Hou, Williams, Siveter, Siveter, Aldridge and Sansom2010]). Eoasmussia Soot-Ryen, Reference Soot-Ryen1960, described as a ‘conchostracan,’ has been referred to as a pelecypod mollusk (Pojeta, Reference Pojeta1971). The late Silurian taxon Asmussia? buchoti Péneau, Reference Péneau1936 (see also in Bate et al., Reference Bate, Collins, Robinson, Rolfe, Harland, Holland, House, Hughes, Rebolds, Rudwick, Satterthwaite, Tarlo and Willey1967), was apparently made without consulting Novojilov (Reference Novojilov1961), who had previously reclassified Péneau's (Reference Péneau1936) taxonomy as Glyptoasmussia buchoti and found it to be Middle Devonian.
Although discerning the oldest clam shrimp is difficult, forms from the Early Devonian provide very important information on the origin and early evolution of clam shrimp. Belgolimnadiopsis Novojilov, Reference Novojilov1958, and Palaeolimnadiopsis both belong to the family Palaeolimnadiopseidae Defretin, Reference Defretin1965, which is the most representative posterior-recurved-type clam shrimp. Their presence in the Early Devonian evinces that clam shrimp with posterior recurvation of growth lines were already diverse and widely distributed at the time. The recurvation of growth lines occurs mainly in species among five families: Palaeolimnadiopseidae; Sinoestheriidae Chen and Shen, Reference Chen and Shen1982; Ipsiloniidae Novojilov, Reference Novojilov1958; Vertexiidae Kobayashi, Reference Kobayashi1954; and Pemphilimadiopseidae Tasch, Reference Tasch1961. Tasch (Reference Tasch1961) thought that the posterior recurvation of growth lines occurred at the genus level, independent of similar development in paleolimnadiopseids. Liao et al. (Reference Liao, Shen, Gao and Huang2019) agreed with Tasch's judgment and considered that the recurvation of growth lines occurred in many clam shrimp clades with diverse characteristics at different ages, indicating the consequence of convergent evolution. Therefore, we cannot say that the ancestral types of palaeolimnadiopseids—Belgolimnadiopsis and Palaeolimnadiopsis—have any inevitable connection with all the forms that possess recurved growth lines. Growth line recurvation has been observed in numerous species from Devonian to recent times (Daday, Reference Daday1925; Shen, Reference Shen1985). The cause of its formation and physiological significance are worth further exploration.
Concherisma eifelense (Raymond, Reference Raymond1946) from the Lower Devonian in Germany was first described and treated as an individual of “Estheria diensti” by Gross (Reference Gross1934). Raymond (Reference Raymond1946) discussed that the posterior end of this specimen (in Gross, Reference Gross1934, fig. 2) was not fully shown in the figure, the shape and recurved direction of some of the growth lines might be due to crushing, but still reassigned it into the genus Palaeolimnadiopsis. Novojilov (Reference Novojilov1961) reclassified this species as “C. eifelense” and displayed a sketch in his publication. According to Novojilov's (Reference Novojilov1961) sketches of five species of Concherisma Novojilov and Varentsov, Reference Novojilov and Varentsov1956, they all possess narrow and long (elongate oval) carapaces and very sharply curved posterior margins, but none show apparent recurved growth lines. Therefore, it should be evident that C. eifelense is not a posterior-recurved-type form.
The genus Pseudestheria, as mentioned above, with the “dumping ground” issue, was once considered to have no stratigraphic significance (Raymond, Reference Raymond1946). Fortunately, however, some constructive suggestions and characteristics have been proposed to differentiate Pseudestheria from other similar forms, such as Polygrapta, Euestheria, and Lioestheria (e.g., Houlb and Kozur, Reference Holub and Kozur1981; Kozur and Sittig, Reference Kozur and Sittig1981; Kozur and Seidel, Reference Kozur and Seidel1983; Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019; for detailed discussion see Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019). Its biostratigraphic value has begun to be considered. Pseudestheria is one of the most common late Paleozoic genera and has a high biostratigraphic value, particularly in the Permian (e.g., Martens, Reference Martens1983; Schneider and Scholze, Reference Schneider and Scholze2018; Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019). Moreover, based on the presently known records, it is also the most common genus in the Early Devonian. We think it is possibly related to the origin of Polygrapta and Euestheria. Holub and Kozur (Reference Holub and Kozur1981) suggested that Pseudestheria was possibly derived from Lioestheria. However, based on the presently known fossil record, Pseudestheria appeared earlier than Lioestheria. Lioestheria shows a morphologically variable (conical to pillar-shaped or especially oval to elongate-shaped) tubercle located on the external surface of the umbo or the larval valve, which distinguishes it from Pseudestheria. In poorly preserved specimens with broken or strongly compressed umbonal areas, it is practical to separate the two genera by the following convention: forms with a larval valve smaller than one-third of the total valve length belong to Pseudestheria, and forms with a larval valve larger than one-third of the valve length belong to Lioestheria (Holub and Kozur, Reference Holub and Kozur1981; Kozur and Sittig, Reference Kozur and Sittig1981; Scholze et al., Reference Scholze, Golubev, Niedźwiedzki, Schneider and Sennikov2019). We believe that Lioestheria may originate from or is closely related to Cornia, which also possesses a tubercle on the larval valve.
Based on the total fossil record, it can be seen that clam shrimp were diverse and widely distributed on Earth during the Early Devonian. In addition, different forms began to form communities with high diversity in suitable biotopes at the time. These earliest species include diverse types—forms with recurved growth lines (Palaeolimnadiopsis and Belgolimnadiopsis), forms with an oval tubercle on its larval valve (Cornia), forms with a very large larval valve (?Palaeolimnadia sp. in this study), and “normal” forms without distinctly special features, such as a tubercle, recurved growth lines, or very large larval valves (Pseudestheria and Concherisma eifelense). We assume that they had a profound effect on the origin and evolutionary trends of clam shrimp that later appeared on Earth. In the Middle and Middle/Late Devonian, global diversity of clam shrimp increased significantly. Some display morphological inheritance from forms of the Early Devonian. Ipsilonia Novojilov, Reference Novojilov1953, of Givetian age possesses recurved growth lines (Novojilov, Reference Novojilov1953). Concherisma and very similar forms, such as Ovjurium Novojilov and Varentsov, Reference Novojilov and Varentsov1956, Ubsanuria Novojilov and Varentsov, Reference Novojilov and Varentsov1956, and Dolichostheria Novojilov and Varentsov, Reference Novojilov and Varentsov1956, also were found in the Givetian strata (Novojilov and Varentsov, Reference Novojilov and Varentsov1956; Novojilov and Kapel'ka, Reference Novojilov and Kapel'ka1960). Some other forms of the Middle and Middle/Late Devonian from Russia, Europe, and China share distinct differences from those of the Early Devonian, possibly indicating the origin of new forms or emergence of new evolutionary trends.
Conclusions
A clam shrimp community that consists of four species, Cornia cheni n. sp., Palaeolimnadiopsis zhangi n. sp., Pseudestheria cf. P. diensti, and ?Palaeolimnadia sp., was found and described from the Early Devonian Yuankou Formation in Jiangyong County, Hunan Province, China. These species are the oldest-known fossil clam shrimp in China and are among the oldest in the world. The Devonian saw the emergence and subsequent diversification of the clam shrimp. This discovery indicates that highly diverse clam shrimp communities already had appeared in suitable biotopes on Earth by the Early Devonian. In addition, based on global records, Early Devonian forms may have had a profound effect on the origin and evolutionary trends of younger forms.
Acknowledgments
This study was supported by the Strategic Priority Research Program (B) (Grant No. XDB26000000 and XDB18000000) of the Chinese Academy of Sciences, the National Natural Science Foundation of China (Grant Nos. 41972007, 41688103, and 41925008), the Open-end Foundation of State Key Laboratory of Palaeobiology and Stratigraphy (NIGPAS; Grant No. 183118), and the Basic Research Plan of Yunnan Province (Youth Project; Grant No. 2019FD130). We are grateful to Z. Feng (Yunnan University) and D. Huang (NIGPAS) for their constructive suggestions, and J. Tang (NIGPAS) for her technical support. We are also grateful to T.A. Hegna (associate editor), F. Scholze, and D. Chu (reviewers of this article) for their constructive comments.



