Introduction
The cosmopolitan family Polymorphidae Meyer, 1931 presently contains 13 genera and about 150 species (Amin, Reference Amin2013; García-Varela et al., Reference García-Varela, Pérez-Ponce de Léon, Aznar and Nadler2013). Phylogenetic analysis of the genera, based on sequences of two nuclear and one mitochondrial gene, supported the monophyly of the family and of nine of the genera analysed (Andracantha Schmidt, 1975, Corynosoma Lühe, 1904, Bolbosoma Porta, 1908, Profilicollis Meyer, 1931, Pseudocorynosoma Aznar, Pérez-Ponce de León & Raga, 2006, Southwellina Witenburg, 1932, Arhythmorhynchus Lühe, 1911, Hexaglandula Petrochenko, 1950 and Ibirhynchus García-Valera, Pérez-Ponce de León, Aznar & Nadler, 2011), but not the genus Polymorphus Lühe, 1911 (see García-Valera et al., 2013). The status of Andracantha as a separate genus had been strongly supported by genetic data (García-Varela et al., Reference García-Varela, Pérez-Ponce de León, Aznar and Nadler2009).
The genus Andracantha was erected by Schmidt (Reference Schmidt1975) to accommodate those species of Corynosoma that bear two fields of somatic spines on the fore-trunk, and have genital spines in at least one sex. The two genera were further distinguished by their use of definitive host; species of Corynosoma mature in pinnipeds, whereas species of Andracantha mature in piscivorous birds. Zdzitowiecki (Reference Zdzitowiecki1989) amended the diagnosis to include … ‘two fields of spines separated by a bare zone or by a zone of smaller spines’. More recently, Aznar et al. (Reference Aznar, Pérez-Ponce de León and Raga2006) re-examined the status of the genus Corynosoma. These authors confirmed the validity of the genus for marine species, erected the genus Pseudocorynosoma to encompass freshwater species of Corynosoma and placed the genus Andracantha in the same clade. A more recent analysis placed Andracantha together with Bolbosoma and Corynosoma in a clade, suggesting a shared a common ancestor (García-Varela et al., Reference García-Varela, Pérez-Ponce de Léon, Aznar and Nadler2013). The genus Andracantha now comprises seven valid species, namely: A. gravida (Alegret, 1941) (type species), A. baylisi (Zdzitowiecki, 1986), A. clavata (Goss, 1941), A. mergi (Lundström, 1941), A. phalacrocoracis (Yamaguti, 1939), A. tunitae (Weiss, 1914) and A. tandemtesticulata Monteiro et al., 2006 (Monteiro et al., Reference Monteiro, Amato and Amato2006).
The shags Leucocarbo chalconotus (Gray, 1843), the Stewart Island shag, and Phalacrocorax punctatus (Sparrman, 1786), the spotted shag, are locally common species, endemic to New Zealand. Individuals are often found dead on beaches or on roads close to the sea, particularly in Otago Harbour. Eudyptula minor (Forster, 1781), the little blue penguin, is found in colonies along the coastlines of southern Australia and New Zealand. Individuals may also be found occasionally washed up on shores other than their home beaches, along the Otago coastline.
Investigations into the intestinal parasites of these three species of piscivorous birds have revealed a number of polymorphids of the Corynosoma–Andracantha morphotype. Specimens of Corynosoma hannae Zdzitowiecki, 1984, were identified from all three of the bird species and have been reported elsewhere (Hernández-Orts et al., Reference Hernández-Orts, Smales, Pinacho-Pinacho, García-Varela and Presswell2017). The other acanthocephalans recovered, representing two morphotypes, corresponded to the diagnosis of Andracantha.
In this paper we present morphological and molecular data to characterize the new species of Andracantha, and provide a key to species of the genus. The significance of the patterns of distribution of trunk spines for species identification and the phylogeny of the family Polymorphidae are discussed briefly.
Materials and methods
Specimens
A total of nine individuals of P. punctatus (spotted shag), six of L. chalconotus (Stewart Island shag) and four of E. minor (little blue penguin) were found dead and were delivered to the first author between April 2012 and November 2014 from coastal sites between Otago Harbour (45°50′S, 170°37′E) and Waikouaiti Beach (45°37′S, 170°40′E) on the south-eastern coast of South Island. Birds were frozen upon receipt and later thawed for dissection of the intestines. Acanthocephalans recovered from the intestines were stored in 70% ethanol. Morphological types were separated into groups and one or more individuals were chosen as vouchers for molecular investigation. A small piece of the posterior end of each voucher was cut off for DNA extraction. For light microscopy, specimens were either cleared in beechwood creosote (temporary mounts), or stained using acetic acid iron carmine, dehydrated in an ethanol series, cleared in clove oil and permanently mounted in Canada balsam. Drawings were made using an Olympus drawing tube mounted on an Olympus compound microscope (Olympus, Tokyo, Japan). Measurements, presented as the range followed by the mean in parentheses, in micrometres unless otherwise stated, were made using Olympus DP2-BSW application software on an Olympus BX51 compound microscope mounted with DP25 camera attachment. Trunk length does not include proboscis length and width is taken at the widest part unless otherwise indicated. For scanning electron microscopy (SEM) the worms were washed for 2 h in distilled water before being fixed overnight in 2.5 % aldehyde in 0.1 m cacodylate buffer. They were then post-fixed in 1% osmium tetroxide for 1 h prior to being dehydrated through a gradient series of ethanols, critical-point dried in a CPD030 BalTec critical-point dryer (BalTec AG, Balzers, Liechtenstein) using carbon dioxide, mounted on aluminium stubs using double-sided adhesive carbon tape, and sputter coated with gold/palladium (60:40) to a thickness of 12 nm in an Emitech K575X Peltier-cooled high-resolution sputter coater (EM Technologies, Ashford, Kent, UK). The specimens were viewed with a JEOL 6700 F field emission scanning electron microscope (JEOL Ltd, Tokyo, Japan) at the Otago Centre for Electron Microscopy (OCEM, University of Otago, New Zealand).
Amplification and sequencing of DNA
Twelve adult worms from spotted shags, ten from Stewart Island shags and one from a little blue penguin were characterized molecularly (table 1). Genomic DNA was extracted from worm tissue in 200 μl of a 5% suspension of Chelex® in deionized water containing 0.1 mg/ml proteinase K, followed by incubation at 56°C for 5 h, boiling at 90°C for 8 min and centrifugation at 14,000 g for 10 min.
Table 1. List of taxa included in the phylogenetic analyses, with data on the host, locality, GenBank accession numbers for the sequences used, and the original reference.

a Accessioned as C. ‘australe’ (see Hernández-Orts et al., Reference Hernández-Orts, Smales, Pinacho-Pinacho, García-Varela and Presswell2017).
A partial fragment (655 bp) of the mitochondrial cytochrome c oxidase 1 gene (cox1) was amplified with forward primer 5′-AGTTCTAATCATAA(R)GATAT(Y)GG-3′ and reverse primer 5′-TAAACTTCAGGGTGACCAAAAAATCA-3′ (Folmer et al., Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994). A partial fragment (approximately 820 bp) of the large ribosomal subunit (LSU) that includes the domains D2 + D3 was amplified using forward primer 5′-CAAGTACCGTGAGGGAAAGTTGC-3′ and reverse primer 5′-GTCGATAGGACTCCCTTTG-3′ (García-Varela & Nadler, Reference García-Varela and Nadler2005). Polymerase chain reaction (PCR) amplifications (25 μl) consisted of 10 mm of each primer, 2.5 μl of buffer 10×, 50 mm MgCl2, 0.5 μl of mixed deoxyribonucleotides (dNTPs) (10 mm), 0.125 μl of Taq DNA polymerase (1 U/μl) (Platinum® Taq DNA, Invitrogen Corporation, Brazil) and 2 μl of DNA template. Thermocycling conditions were denaturation at 94°C for 3 min; followed by 35 cycles of 94°C for 1 min, annealing at 40°C (cox1) or 50°C (LSU) for 1 min and extension at 72°C for 1 min; followed by a post-amplification incubation at 72°C for 7 min. Each PCR product was cleaned and filtered using Millipore columns (Amicon®, Billerica, Massachusetts, USA). Sequencing reactions were performed using ABI BigDye® Terminator sequencing (PE Applied Biosystems, Boston, Massachusetts, USA) and reaction products were run on an ABI 310 capillary DNA sequencer. Contigs were assembled and base-called differences resolved using CodonCode version 5.1.5 (CodonCode Corporation, Dedham, Massachusetts, USA). All sequences were submitted to GenBank (LSU MF401623–24; cox1 MF527023–35).
Alignments and phylogenetic analyses
Cox1 and LSU fragments were aligned, together with published sequences of identified polymorphids from GenBank, using Clustal (Thompson et al., Reference Thompson, Higgins and Gibson1994) (see table 1). The cox1 and LSU alignments were analysed both independently and combined. The Akaike information criterion (AIC) was used to assess the fit of nucleotide substitution models for each alignment and the combined data using Modeltest version 3.0 (Posada & Crandall, Reference Posada and Crandall1998). The GTR model with invariable sites (+I) and rate heterogeneity (+G) (Yang, Reference Yang1994) was determined to be the best fit for the alignments. Maximum Likelihood analyses were conducted using PAUP* 4.0b10 (Swofford, Reference Swofford2002). Bayesian analyses were performed with the program MrBayes version 3.1.2 (Huelsenbeck & Ronquist, Reference Huelsenbeck and Ronquist2001) with the GTR + I + G model. The settings were two simultaneous runs with four Markov chains and 10 million MCMC generations, sampling every 1000 generations, a heating parameter value of 0.2. The outputs of MrBayes were examined with Tracer Version 1.4 (Rambaut & Drummond, Reference Rambaut and Drummond2007) to check for convergence of different parameters, and to determine the approximate number of generations at which log-likelihood values stabilized. A consensus tree was obtained after a conservative burn-in of 25% was applied for each dataset. Trees were drawn using FigTree version 1.3.1 (Rambaut, Reference Rambaut2009). The genetic divergence among taxa was estimated with the cox1 and LSU alignment using uncorrected ‘p’ distances with the program MEGA version 6 (Tamura et al., Reference Tamura, Stecher, Peterson, Filipski and Kumar2013).
Results
Andracantha leucocarboi n. sp.
Taxonomic summary
Type host. Leucocarbo chalconotus (Gray, 1843), the Stewart Island shag (Pelecaniformes: Phalacrocoracidae).
Other host. Phalacrocorax punctatus (Sparrman, 1786), the spotted shag (Pelecaniformes: Phalacrocoracidae).
Type locality. Otago Harbour, Dunedin, New Zealand (45°50′S, 170°37′E).
Type material. Holotype male AHC 36416; allotype female AHC 48080; voucher AHC 48081; hologenophore from L. chalconotus AHC 48086.
Material examined. Seventeen specimens from L. chalconotus (Otago Museum IV86232–IV86235); nine specimens from P. punctatus (IV86236–IV86238).
Site in host. Intestine.
Prevalence. Females more abundant than males in all hosts. Prevalence in P. punctatus, 5 out of 9 birds; in L. chalconotus, 3 out of 6 birds.
Intensity. Mean intensity in P. punctatus 19.2, (range 1–59); mean intensity in L. chalconotus 4.0 (range 1–7).
Etymology. The specific epithet refers to the type host.
Description
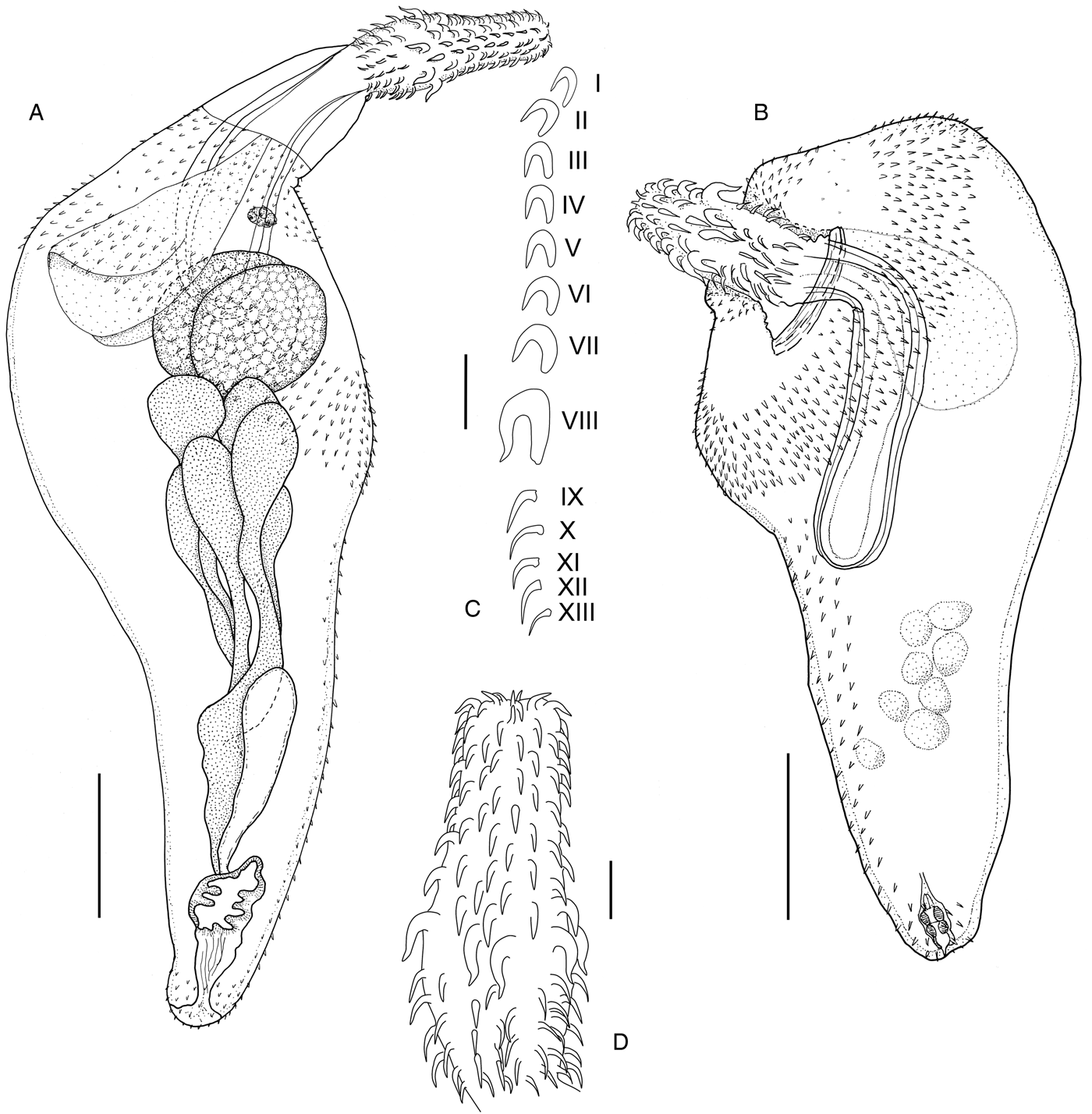
General. Small acanthocephalans (figs 1A–D, 2A–C). Trunk expanded anteriorly into bulb or disc; posterior trunk cylindrical or slightly tapering. Sexes not significantly different in size. Proboscis swollen just posterior to mid-region, armed with 18 longitudinal rows of 12 hooks each. Anterior 7–8 hooks with well-developed simple roots, shorter than thorn except for hook VIII; posterior 4–5 hooks spiniform with discoid roots. Hooks VIII always longest and most massive; hooks I most slender (table 2). Hooks VIII significantly larger in females. Proboscis length to width ratio 1:2.0–3.0. Proboscis receptacle double-walled; cerebral ganglion midway down length. Neck unarmed. Lemnisci leaf-shaped, half to two-thirds length of receptacle. Somatic spines on anterior disc in two fields with bare region between, posterior spine field more than three times deeper than anterior field; bare region widest on ventral surface; spines up to 38–44 (41) long. Small, sparse spines scattered dorsally (fig. 2A). Spines, somatic and genital, absent from hindbody in both sexes. Genital pore terminal.
Male. Based on four specimens. Trunk 2.8–3.5 (3.2) mm long × 1.1–1.5 (1.3) mm wide. Proboscis 644–769 (706) long × 242–296 (266) wide; neck not extended on any specimens. Proboscis receptacle 971–1212 (1118) long × 193–307 (252) wide. Lemnisci about 550 long. Testes oval, more or less side by side, extend slightly beyond proboscis receptacle; 418–506 (449) × 277–300 (290) and 399–545 (452) × 275–293 (285). Cement glands, six, pyriform, 220–240 long × 150 wide; cement ducts broad, convoluted. Saefftigen's pouch 400–450 long (n = 2). Bursa 305 long × 385 wide (n = 1); circumbursal papillae, about 40, 10 diameter (fig. 2C).
Female. Based on 17 specimens. Trunk 2.3–4.8 (3.4) long × 1.3–2.2 (1.8) mm wide. Proboscis 641–828 (711) long × 229–365 (307) wide; neck 110–225 (146) long × 246–360 (281). Proboscis receptacle 785–1362 (1182) long × 161–375 (300) wide. Lemnisci about 550 long. Reproductive system 1245–1640 (1432) long. Eggs with polar prolongations of middle membrane, 71–118 (83) × 27–36 (31) long. Genital pore terminal.

Fig. 1. Andracantha leucocarboi n. sp. ex Leucocarbo chalconotus. (A) Male, (B) female, (C) proboscis hook row, (D) proboscis of female. Scale bars: (A, B) 500 μm; (C, D) 100 μm.

Fig. 2. Scanning electron micrographs of Andracantha leucocarboi n. sp. (A) Female adult worm, (B) proboscis en face showing 18 hook rows, (C) interior of male copulatory bursa showing circumbursal papillae. Scale bars: (A) 500 μm; (B and C) 50 μm.
Table 2. Proboscis hook measurements for Andracantha leucocarboi n. sp. males and females.

Remarks
In having a cylindrical proboscis with swollen area, small tapering body with anterior swelling or disc, anterior trunk with two fields of spines separated by a zone of no spines or smaller dispersed spines, six cement glands and eggs with polar prolongations, these specimens may be placed in the genus Andracantha (see Schmidt, Reference Schmidt1975).
The seven known species of Andracantha (see Amin, Reference Amin2013) can be divided into those with ventral trunk spines and those without. Andracantha leucocarboi, in possessing no ventral trunk spines, is most similar to the cluster of species comprising A. baylisi, A. gravida, A. mergi and A. tandemtesticulata, which also have no ventral trunk spines.
Andracantha leucocarboi differs from A. baylisi in lacking ventral spines, having shorter lemnisci (550 compared with 900), smaller proboscis hooks, and hook VIII, the largest hook, 76–98 long, compared with hook V, the largest hook, 105–136 long in A. baylisi. In addition hook I is longer than hook VII in A. baylisi, a condition not found in A. leucocarboi (Zdzitowiecki, Reference Zdzitowiecki1989).
Andracantha leucocarboi, a smaller worm than A. gravida (females 2.3–4.8 mm long compared with females 4.2–6.0 mm long), differs from A. gravida in lacking genital spines (Schmidt, Reference Schmidt1975). Furthermore, A. leucocarboi differs from A. gravida in having a proboscis armature of 18 longitudinal rows of hooks compared with 16 (Schmidt, Reference Schmidt1975).
Andracantha leucocarboi does not display the sexual dimorphism seen in A. mergi, has a longer proboscis (644–769 as compared to 445–575 in males); a proboscis armature of 7/8 rooted and 4/5 spiniform hooks per longitudinal row, compared with 6/7 and 3/4 for A. mergi, and larger eggs (71–118 long compared with 44–50) (Schmidt, Reference Schmidt1975).
Andracantha leucocarboi is a smaller worm than A. tandemtesticulata (males 2.8–3.5 mm compared with 4.0–7.4 mm), has a proboscis armature with smaller, less uniformly sized hooks (34–98 versus 35–60) and fewer spiniform hooks (4–5 versus 6–9) in each longitudinal row. Furthermore, A. tandemtesticulata has orange pigmentation, tandem testes and two inflations of the body (Monteiro et al., Reference Monteiro, Amato and Amato2006).
Andracantha clavata, A. phalacrocoracis and A. tunitae can be distinguished from A. leucocarboi in having ventral trunk spines (Johnston & Best, Reference Johnston and Best1942; Johnston & Edmonds, Reference Johnston and Edmonds1953; Schmidt, Reference Schmidt1975; Zdzitowiecki, Reference Zdzitowiecki1986, Reference Zdzitowiecki1989, Reference Zdzitowiecki1991).
Andracantha sigma n. sp.
Taxonomic summary
Type host. Phalacrocorax punctatus (Sparrman, 1786), the spotted shag (Pelecaniformes: Phalacrocoracidae).
Other hosts. Leucocarbo chalconotus (Gray, 1843), the Stewart Island shag (Pelecaniformes: Phalacrocoracidae); Eudyptula minor (Forster, 1781), the little blue penguin (Sphenisciformes: Spheniscidae).
Type locality. Otago Harbour, Dunedin, New Zealand (45°50′S, 170°37′E).
Other localities. Waikouaiti beach, Otago, New Zealand (45°59′S, 170°67′E).
Type material. Holotype male AHC 48082; allotype female AHC 48083; voucher from E. minor AHC 36417; hologenophores from P. punctatus AHC 48084, 48087; from L. chalconotus AHC 48085.
Material examined. Twenty-nine specimens from P. punctatus (Otago Museum IV86239–IV86244); 14 specimens from L. chalconotus (not accessioned); 2 specimens from E. minor (not accessioned).
Site in host. Intestine.
Prevalence. Females more abundant than males in all hosts. Prevalence in P. punctatus 8 out of 9 birds; in L. chalconotus, 4 out of 6 birds; in E. minor, 3 of 4 birds.
Intensity. Mean intensity in spotted shags 21 (range 2–46); mean intensity in Stewart Island shags 7 (range 1–14); mean intensity in little blue penguins 3 (range 1–6).
Etymology. The specific epithet refers to the sigmoidal shape of the largest proboscis hook.
Description
General. Trunk (figs 3 and 4) expanded anteriorly into bulb or disc with two fields of spines, postero-ventral spines may not be continuous with second field of disc spines; posterior trunk tapering. Sexual dimorphism not evident. Proboscis swollen just posterior to mid-region, armed with 12–16 longitudinal rows of 12–13 hooks each. Anterior 8 hooks with well-developed simple roots, shorter than thorns; posterior 4–5 hooks spiniform with discoid roots. Hook VIII largest and most massive, about twice the size of next largest hook, with distinctive sigmoidal shape (fig. 4C); hook I more slender than hooks II–XIII (table 3). Proboscis length to width ratio 1:2.0–3.0. Neck unarmed. Proboscis receptacle double-walled, cerebral ganglion about midway down length. Lemnisci sub-circular, one-third to half length of receptacle, usually folded on ventral edge. Somatic spines on anterior disc in two fields with a bare region between; posterior spine field about as deep as anterior field; bare region about the same depth ventrally and dorsally; small, sparse spines partially filling bare region dorsally, in some specimens. Largest somatic spines in posterior disc field. Ventral spines smaller than disc spines, sparse, forming third field, not continuous with field on posterior disc, in narrow band extending to posterior end of worm (fig. 4B). Genital spines present in both sexes. Genital pore terminal.
Male. Based on 11 specimens. Trunk 2.3–3.1 (2.7) mm long × 1.0–1.4 (1.3) mm wide. Proboscis 612–744 (676) long × 215–320 (266) wide. Largest proboscis hook (VIII) up to 1.7 times longer than next largest (VII). Neck on most extended specimen 388 long × 426 wide. Proboscis receptacle 908–1193 (1039) long × 215–323 (254) wide. Lemnisci about 380 long (n = 1). Largest somatic spines on disc 34–49 (43) long; ventral trunk spines 19–30 (25) long. Testes about level with proboscis receptacle; one, 507–599 (560) long × 275–453 (358) wide, slightly more anterior than the other, which is 510–607 (562) long × 253–459 (344) wide. Six cement glands, pyriform, 310 long × 200 wide; cement ducts broad, convoluted. Saefftigen's pouch 360 long (n = 2). Genital spines 21–33 (25) long × 5–7 (6) wide; continuous with ventral spines; encircle genital pore. Bursa not everted in any specimens.
Female. Based on 11 specimens. Trunk 2.3–3.1 (2.8) mm long × 1.0–1.7 (1.3) mm wide. Proboscis 566–771 (647) long × 222–304 (261) wide. Largest proboscis hook (VIII) up to 1.7 times longer than next largest (VII). Neck not extended in any specimens. Proboscis receptacle 979–1134 (1070) long × 231–355 (272) wide. Lemnisci about 400 long (n = 1). Largest somatic spines 35–48 (42) long; ventral trunk spines 19–31 (23) long. Reproductive system 1260 long (n = 1). Eggs with polar prolongations of middle membrane; 65–88 (75) long × 17–31 (22) wide. Genital spines 24–34 (29) long × 7–11 (8) wide; continuous with ventral spines; encircle genital pore.

Fig. 3. Andracantha sigma n. sp. ex Phalacrocorax punctatus. (A) Male, (B) female, (C) proboscis hook row, (D) proboscis of female. Scale bars: (A, B) 500 μm; (C, D) 100 μm.

Fig. 4. Scanning electron micrographs of Andracantha sigma n. sp. (A) Male adult worm, (B) ventral body showing three distinct fields of spines, (C) distinctive sigmoid large hook of the proboscis, (D) proboscis (not fully everted). Scale bars: (A, B) 500 μm; (C) 10 μm; (D) 100 μm.
Table 3. Proboscis hook measurements for Andracantha sigma n. sp. males and females.

Remarks
Andracantha sigma conforms to the diagnosis of the genus given by Schmidt (Reference Schmidt1975) as discussed above. This new species is morphologically closest to those species of Andracantha that have ventral trunk spines, namely A. clavata, A. phalacrocoracis and A. tunitae.
The scattering of small spines between the fields of disc spines distinguishes A. sigma from A. clavata, A. phalacrocoracis and A. tunitae. Andracantha sigma can be further distinguished from A. clavata in having larger testes (507–607 compared with 250–400 long) and larger proboscis hooks (up to 134 long compared with up to 83 long) (Zdzitowiecki, Reference Zdzitowiecki1991); from A. phalacrocoracis by its smaller size (length 2.3–3.1 mm versus 4.0–6.0 mm), a proboscis armature of 12–16 versus 16–19 longitudinal rows of hooks, and 6 cement glands in the male, as opposed to 8 (Schmidt, Reference Schmidt1975); and from A. tunitae in having a proboscis armature of 12–16 compared with 16–19 longitudinal rows of hooks, and genital spines (Zdzitowiecki, Reference Zdzitowiecki1986).
The only two species available for molecular comparison, A. leucocarboi and A. gravida (see below), differ from A. sigma in lacking ventral trunk spines.
The large, S-shaped proboscis hook VIII, longer than the equivalent in any other species of Andracantha, sub-circular lemnisci (not elongated or leaf shaped) and the ventral spines separated from the posterior disc field by an aspinous gap, differentiate A. sigma from all other species in the genus.
A key to the species of Andracantha is given below.
Molecular results
Comparative sequence analysis of the newly characterized isolates revealed two genotypes for the cox1 region, which corresponded to the morphotypes A. leucocarboi and A. sigma as described above. Variability within each species was low: A. sigma 0.00–0.32% and A. leucocarboi 0.00–1.38%.
Including all taxa, divergence (uncorrected p-distances) within species for this region of cox1 ranged from 0.0 to 4.6%, between species it ranged from 11.3 to 22.2% and between genera it ranged from 18.3 to 33.3%. There is therefore a degree of overlap between genetic distinctiveness within and between genera as they stand at the moment.
The combined dataset (LSU + cox1) consisted of 2198 characters with 30 taxa. Figure 5 shows a phylogenetic hypothesis depicted by ML analysis of the combined cox1 and LSU dataset. Choice of outgroups was made based on a published molecular phylogeny (see García-Varela et al., Reference García-Varela, Pérez-Ponce de León, Aznar and Nadler2009, Reference García-Varela, Pérez-Ponce de Léon, Aznar and Nadler2013). This tree shows those genera of Polymorphidae included in the dataset to be monophyletic, with the exception of the genus Polymorphus. This supports the findings of García-Varela et al. (Reference García-Varela, Pérez-Ponce de León, Aznar and Nadler2009, Reference García-Varela, Pérez-Ponce de Léon, Aznar and Nadler2013) and reiterates the need for a taxonomic revision of this latter genus.

Fig. 5. Maximum Likelihood tree based on DNA sequences of cox1 and LSU genes, with –ln of 21391.48. Branch lengths are proportional to the inferred amount of nucleotide substitutions; numbers at nodes are ML bootstrap values based on 100 replicates (left of slash) and Bayesian probabilities (right).
The specimens of A. leucocarboi form a clade with A. gravida, and A. sigma is sister to these two Andracantha species, this clade being strongly supported by Bayesian probabilities. The sister clade to these genera consists of species of Corynosoma and Bolbosoma.
The LSU dataset (1543 characters: 30 taxa) gave the same overall topology (not shown), except that, within the Corynosoma/Andracantha clade, A. sigma was more closely related to species of Corynosoma than to those of Andracantha. However, support for this relationship was poor. In this tree the genus Corynosoma is seen to be paraphyletic with respect to the genus Bolbosoma. The cox1 dataset alone (655 characters; 47 taxa) produced a tree with the same arrangement of Corynosoma/Andracantha as the combined dataset, but showed variation in the placement of the outgroups Ibirhynchus dimorpha and Hexaglandula corynosoma and the Pseudocorynosoma clade, all of which were poorly supported (tree not shown).
Discussion
Species of Andracantha are usually associated with shags and cormorants, Phalacrocoracidae, as their definitive hosts. The only reported exception is A. mergi, which is found in the merganser, Mergus serrator L. 1758 and American bald eagle, Haliaeetus leucocephalus (L. 1766) (Nickol & Kocan, Reference Nickol and Kocan1982). Our record of A. sigma in the little blue penguin, E. minor, is the second report of a host other than a phalacrocoracid, and the first of a species of Andracantha from a penguin. Previous records of acanthocephalans from penguins are of species of Corynosoma, whose definitive hosts are usually marine mammals (Brandão et al., Reference Brandão, Moreira and Luque2014).
The intermediate arthropod and paratenic fish hosts for the two species reported herein are not known. The diet of the Stewart Island shag consists primarily of tripterygiid and pleuronectid fish, along with a quantity of crustacea, and to a lesser extent cephalopods and polychaetes. Spotted shag diets focus more on morid fish, with a smaller content of crustacea and cephalopods (Lalas, Reference Lalas1983). These two species of shag feed mainly on the bottom of the sea, often many kilometres offshore and in depths of up to 50 m (Lalas, Reference Lalas1983). The diet of little blue penguins is similar to that of the two shags, but with a higher proportion of squid (Flemming et al., Reference Flemming, Lalas and van Heezik2013). The cystacanth of the related acanthocephalan Corynosoma hannae is found in flatfish sole and brill (Hernández-Orts et al., Reference Hernández-Orts, Smales, Pinacho-Pinacho, García-Varela and Presswell2017), and it is likely that the intermediate host for the two new species is a sea-floor crustacean that is a key organism in the marine food web, with benthic fish being paratenic hosts (Kennedy, Reference Kennedy2006).
Trunk spination is often used in morphological characterization of this group of trematodes, but should be undertaken with care. In some instances, ventral spines may not be present on individual worms, may go unnoticed or be unreported. The original description of A. clavata by Goss (Reference Goss1940), for example, mentioned neither a ventral field of spines nor genital spines. Subsequent reports included both ventral and genital spines (Johnston & Best, Reference Johnston and Best1942; Zdzitowiecki, Reference Zdzitowiecki1991) and genital spines only (Johnston & Edmonds, Reference Johnston and Edmonds1953). This suggests that some individuals, or some populations, of A. clavata may have either very small ventral spines, or none at all. For genital spines in particular, there may be considerable individual variation between specimens. Nevertheless, the presence or absence of fields of genital and trunk spines, and their distribution, have been used in helping to characterize the species of Andracantha (see for example Schmidt, Reference Schmidt1975; Aznar et al., Reference Aznar, Pérez-Ponce de León and Raga2006; Monteiro et al., Reference Monteiro, Amato and Amato2006). The bare area on the ventral trunk of A. sigma between the posterior field of disc spines and the ventral field of spines may be an adaptive feature, improving attachment to the host (Aznar et al., Reference Aznar, Crespo, Raga and Hernández-Orts2016). The presence of a sparse scattering of small spines across otherwise bare fields of trunk spines has been noted as a distinguishing character for A. baylisi, A. leucocarboi and A. sigma (Zdzitowiecki, Reference Zdzitowiecki1986; this study). These spines are small and difficult to observe on some specimens and their presence may be subject to individual variation. Therefore, this character needs to be treated with caution as it may have been underreported in the descriptions of other species of Andracantha.
The circumbursal papillae observed on the bursa of A. leucocarboi (fig. 2C) appear to resemble closely the description given for the circumbursal papillae of Corynosoma hamanni by Holloway & Nickol (Reference Holloway and Nickol1970), both species being in the family Polymorphidae. These authors estimated that there were 35 papillae, each about 7.5 μm high and about the same in diameter, while our estimate was about 40, 10 μm in diameter. Such papillae appear to have been illustrated by SEM photographs only for Pomphorhynchus laevis (Zoega in Muller, 1776), family Pomphorhynchidae and Acanthocephalus ranae (Shrank, 1788), family Echinorhynchidae (Heckmann et al., Reference Heckmann, Amin, Tepe, Dusen and Oguz2011). The papillae of P. laevis and A. ranae appear to differ from those of A. leucocarboi and C. hamanni. More data are needed, however, before the significance at a taxonomic level, if any, of the morphology of the circumbursal papillae can be determined.
The ML and Bayesian trees derived from the combined data (cox1 + LSU) placed A. sigma n. sp. and A. leucocarboi n. sp. as sister taxa to other species of Andracantha from Middle America, suggesting that the genus is monophyletic and that the association of this genus with phalacrocoracid birds could represent a single event of colonization, with some exceptions such as A. mergi. However, to test this hypothesis it will be necessary to analyse the other congeneric species within a phylogenetic framework. With this description of two new species, the genus Andracantha now contains nine species parasitizing shags, cormorants and other piscivorous birds.
The genetic distance between A. leucocarboi and A. gravida was 22.2%, and between A. leucocarboi and A. sigma was 21.4%, both of which fall within the intergeneric range. However, at this stage there are no morphological characters to support the erection of new genera for the two new species described herein. New specimens of other Andracanda species should help to place the new species with more certainty.
Distances between taxa for the LSU marker generally supported those seen for cox1. Genetic distances within and between species and genera in this study largely corroborate those reported by García-Varela & Pérez-Ponce de León (Reference García-Varela and Pérez-Ponce de León2008) for the same region of cox1 within the Polymorphidae. Their study found that intraspecific distances ranged from 1 to 5%, interspecific distances ranged from 11 to 21% and intergeneric distances ranged from 22 to 30%. Our additional data therefore add strength to these genetic distance criteria for species delineation.
Acknowledgements
Thanks to Matthew Downes of the Department of Zoology and Liz Girvan of the OCEM for their assistance with scanning electron microscopy.
Financial support
This work has been supported indirectly by the Marsden Fund (Royal Society of New Zealand) and a Zoology Department PBRF Research Enhancement grant to Professor Robert Poulin.
Conflict of interest
None.
Ethical standards
The sampling in this work complies with the current laws and animal ethics regulations of New Zealand.
Key to the species of Andracantha
1 Trunk with two swellings, testes tandem………tandemtesticulata
Trunk without two swellings, testes parallel………2
2 With scattering of small spines between trunk fields………3
Without scattering of small spines between trunk fields………4
3 Largest hook VIII, 76–98 long; lemnisci 550 long………leucocarboi n. sp.
Largest hook V, 105–136 long; lemnisci 900 long………baylisi
4 With 8 cement glands, hook IX largest………phalacrocorax
With 6 cement glands, hooks V–VIII largest………5
5 With ventral field of trunk spines………6
Without ventral field of trunk spines………7
6 14–16 rows of 10–11 hooks, largest hook 78–83 long, with genital spines, testes 250–420………clavata
16–19 rows of 10–12 hooks, largest hook 60–70 long, without genital spines, testes 450–650………tunitae
7 12–13 hooks per row, hook VIII massive, 97–134 long, sigmoid shape, lemnisci subcircular………sigma n. sp.
9–12 hooks per row, hooks VI/VII largest, not sigmoid, lemnisci not subcircular………8
8 14–16 rows, 11–12 hooks per row, last 4–5 hooks without roots, genital spines displaced ventrally, eggs 68–80………gravida
15–17 rows, 9–10 hooks per row, last 2–3 hooks without roots, genital spines in female only, eggs 44–50………mergi