Introduction
The genus Moniliformis Travassos, 1915 comprises 17 species that parasitize mammals and birds in different parts of the world (Amin et al., Reference Amin, Heckmann, Halajian, El-Naggar and Takavol2014, Reference Amin, Heckmann, Mohammed and Evans2016, Reference Amin, Heckmann, Sharifdini and Albayati2019; Martins et al., Reference Martins, Del Rosario Robles and Navone2017). Two species parasitize Brazilian mammals: Moniliformis moniliformis (Bremser, 1811) Travassos, 1915 and Moniliformis travassosi Meyer, 1932. Moniliformis moniliformis (the type species) is cosmopolitan and generalist, infecting humans, wild and domestic mammals, and is considered an agent of acanthocephaliasis in humans, which is a zoonotic disease (Travassos, Reference Travassos1917; Amin, Reference Amin, Crompton and Nickol1985; Meyers et al., Reference Meyers, Neafie, Marty and Wear2000; Berenji et al., Reference Berenji, Fata and Hosseininejad2007; Salehabadi et al., Reference Salehabadi, Mowlavi and Sadjjadi2008). In Brazil, it has been reported infecting the rodents Rattus rattus Linnaeus, 1758 and Rattus norvegicus Berkenhout, 1769, and the bat Phyllostomus hastatus Pallas, 1767, in different regions (Travassos, Reference Travassos1917; Machado Filho, Reference Machado Filho1946; Gibson and McCarthy, Reference Gibson and McCarthy1987; Tietz Marques and Scroferneker, Reference Tietz Marques and Scroferneker2003; Araújo et al., Reference Araújo, Mendes, Langone and Müller2014; Santos and Gibson, Reference Santos and Gibson2015; Simões et al., Reference Simões, Garcia, Costa-Neto, Santos, Faro and Maldonado2017). Moniliformis travassosi Meyer, 1932 has been reported infecting only R. norvegicus (Travassos, Reference Travassos1917; Machado Filho, Reference Machado Filho1946).
Necromys lasiurus (Lund, 1840) is a small terrestrial Sigmodontinae (<80 g) (Rodentia: Cricetidae), which is broadly distributed in South America, from the Atlantic coast through central Brazil to the southern Amazon basin, including north-eastern Argentina, extreme south-eastern Peru, Paraguay and Bolivia (Redford & Eisenberg, Reference Redford and Eisenberg1999; Bonvicino et al., Reference Bonvicino, Oliveira and D'Andrea2008). In Brazil, this sigmodontine rodent inhabits grasslands of Cerrado, Pantanal, Caatinga and open areas in the Atlantic Forest (Bonvicino et al., Reference Bonvicino, Oliveira and D'Andrea2008) and has a generalist diet (Redford & Eisenberg, Reference Redford and Eisenberg1999; Vieira et al., Reference Vieira, Baumgartem, Paise and Becker2010). Among the helminths described in N. lasiurus, nematodes are the most frequently reported (Vicente et al., Reference Vicente, Rodrigues, Gomes and Pinto1997; Anderson et al., Reference Anderson, Chabaud and Willmott2009). In general, studies of helminths from Brazilian rodents often reported nematodes (Vicente et al., Reference Vicente, Rodrigues, Gomes and Pinto1997; Anderson et al., Reference Anderson, Chabaud and Willmott2009; Simões et al., Reference Simões, Gentile, Rademaker, D'Andrea, Herrera, Freitas, Lanfredi and Maldonado2010, Reference Simões, Souza, Maldonado and Luque2011, Reference Simões, Maldonado and Luque2012, Reference Simões, Garcia, Costa-Neto, Santos, Faro and Maldonado2017; Cardoso et al., Reference Cardoso, Simões, Luque, Maldonado and Gentile2016; Tavares et al., Reference Tavares, Campião, Costa-Pereira and Paiva2017; Costa et al., Reference Costa, Simões, Vilela, Souza, Cardoso, Leiner, Gentile and Maldonado2018); however, most studies are case reports. Studies of acanthocephalans from rodents, including taxonomy, ecology, molecular biology and phylogeny, are still scarce.
Descriptions of Moniliformis species are based mainly on morphological features. However, molecular phylogenies including species of Moniliformis have been complementing conventional taxonomy in studies of integrative taxonomy, revealing new and cryptic species (Amin et al., Reference Amin, Heckmann, Halajian, El-Naggar and Takavol2014, Reference Amin, Heckmann, Mohammed and Evans2016, Reference Amin, Heckmann, Sharifdini and Albayati2019). During an investigation of hantavirus cases (Limongi et al., Reference Limongi, Moreira, Peres, Suzuki, Ferreira, Souza, Pinto and Pereira2013), rodents were captured in the municipality of Uberlândia (18°55'07″S, 48°17'19″W), in the state of Minas Gerais, south-eastern Brazil, within the Cerrado biome. Helminths were recovered for an ecological study of the metacommunity structure of the helminth community of the rodent N. lasiurus (Costa et al., Reference Costa, Simões, Vilela, Souza, Cardoso, Leiner, Gentile and Maldonado2018). From that study, we recovered specimens of the genus Moniliformis, which had never been previously reported for this host. Herein, we describe a new species of the genus Moniliformis recovered from the rodent N. lasiurus captured in the Brazilian Cerrado biome. The description is based on morphology and molecular phylogenetic analyses.
Material and methods
Field study and collection of acanthocephalan specimens
Specimens of N. lasiurus were captured with Sherman® traps (3 × 3.75 × 12 inches) and Tomahawk® traps (16 × 5 × 5 inches) (Hazelhurst, Wisconsin, USA) baited with a mixture of peanut butter, banana, oats and bacon. Trapping occurred between December 2011 and November 2012. Mammals were anaesthetized, euthanatized and necropsied, and the abdominal and thoracic cavities were examined for helminths. Rodent capture and handling were authorized by the Chico Mendes Institute for Biodiversity Conservation (Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio)), under license number 13373, and by the Ethics Committee on Animal Use of Oswaldo Cruz Institute (CEUA; Instituto Oswaldo Cruz (FIOCRUZ-RJ)), according to licences L-049/08 and 066/08.
Morphological analysis
Worms recovered were washed in saline solution to remove tissue debris, fixed in 70% ethanol and taken to the Laboratory of Biology and Parasitology of Reservoir Wild Mammals (Laboratório de Biologia e Parasitologia de Mamíferos Silvetres Reservatórios (LABPMR)). At the LABPMR, the acanthocephalan specimens used for morphological characterization were stained with acid carmine, destained in a solution of 2% hydrochloric acid (HCl) and 70% ethanol, dehydrated in a graded ethanol series (70 to 100%), cleared in 90% phenol (modified from Amato, Reference Amato1985) and analysed using an Axion Scope A1 light microscope and Zeiss Scope Z1 light microscope (Zeiss, Göttingen, Germany). Drawings were made with the aid of a camera lucida attached to a Nikon light microscope (Nikon Corporation, Tokyo, Japan). Measurement ranges are in millimetres unless otherwise stated, followed by mean within parentheses. One male and two female specimens, preserved in ethanol, were deposited in the Helminthological Collection of the Oswaldo Cruz Institute (Coleção Helmintológica do Instituto Oswaldo Cruz (CHIOC)), Rio de Janeiro, Brazil (CHIOC number 38594: (a) holotype; (b) allotype; and (c) paratype).
For scanning electron microscopy (SEM), two specimens, previously fixed in 70% ethanol, were dehydrated in an ascending ethanol series (80%, 90%, 100%), critical-point dried with CO2, mounted with silver adhesive tape on aluminium stubs and sputter-coated with a 20 nm layer of gold. Samples were examined using a JSM-6390 LV microscope (JEOL, Akishima, Tokyo, Japan) at an accelerating voltage of 15 kV, in the Electron Microscopy Platform of the Oswaldo Cruz Institute (Plataforma de Microscopia Eletrônica Rudolf Barth (IOC-FIOCRUZ)).
Molecular phylogenetic analyses
For genomic DNA recovery, acanthocephalan specimens preserved in 70% ethanol were washed in ultrapure water for 24 h at room temperature. Total genomic DNA was isolated from one individual worm using the QIAamp DNA Mini Kit according to the manufacturer's protocol (Qiagen, Venlo, Netherlands). DNA amplification by polymerase chain reaction (PCR) was conducted using two primer pairs: the partial nuclear large subunit ribosomal RNA (28S rRNA) gene was amplified using the primers C1 5′-ACCCGCTGAATTTAAGCAT-3′ and D2 5′-TGGTCCGTGTTTCAAGAC-3′ (Hassouna et al., Reference Hassouna, Michot and Bachellerie1984 – modified from Chisholm et al., Reference Chisholm, Morgan, Adlard and Whittington2001); and the partial mitochondrial cytochrome c oxidase subunit I (MT-COI) gene using the primers F 5′-CTAATCATAARGRTATYGG-3′ and R 5′-TAAACYTCAGGRTGACCAAARAAYCA-3′ (Falla et al., Reference Falla, Brieva and Bloor2015 – modified from Folmer et al., Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994). PCR amplifications were performed using Promega PCR Master Mix (Promega Corporation, Madison, Wisconsin, USA) in 25 µl reactions, following the manufacturer's protocol. The thermal cycling profiles were programmed on an Eppendorf Mastercycler Epsystem thermocycler (Eppendorf, Hamburg, Germany) and executed. For the28S rRNA gene: initial denaturation step at 95°C (2 min); followed by 40 cycles of denaturation at 94°C (60 s), annealing at 55°C (60 s), and extension at 72°C (60 s); followed by a final extension at 72°C (5 min); and hold at 4°C. For the MT-COI gene: initial denaturation step at 95°C (2 min); 35 cycles of 94°C (1 min), 40°C (1 min) and 72°C (1 min); followed by a final extension at 72°C (5 min); and hold at 4°C. PCR products were analysed, after electrophoresis, on a 1.5% agarose gel using GelRed Nucleic Acid Gel Stain (Biotium, Hayward, California, USA) by visualizing on a UV transilluminator.
Successful amplifications were purified using the QIAquick PCR Purification Kit (Qiagen, Venlo, Netherlands) following the manufacturer's protocol. Sequencing reactions using the Big Dye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, California, USA) were performed using the same primers mentioned above in a Gene Amp thermocycler (Applied Biosystems) and analysed using an ABI 3730 DNA analyser (Applied Biosystems). Both procedures and cycle-sequenced product precipitations were conducted at the subunit RPT01A – DNA sequencing platform of the Oswaldo Cruz Institute (Programa de Desenvolvimento Tecnológico em Insumos para Saúde/FIOCRUZ).
For each gene, chromatograms were initially assembled into contigs and manually edited for ambiguities using the software package Geneious 9.1 (Biomatters Ltd., Auckland, New Zealand; Kearse et al., Reference Kearse, Moir and Wilson2012). The resulting consensus sequences were compared for similarities with sequences of the GenBank database using the Basic Local Alignment Search Tool algorithm (https://blast.ncbi.nlm.nih.gov/Blast.cgi) from the National Centre for Biotechnology Information (https://www.ncbi.nlm.nih.gov/).
For molecular phylogenetic analyses using the 28S rRNA and the MT-COI datasets, we added representative sequences of the class Archiacanthocephala Meyer, 1931 retrieved from GenBank. Three families, representing the orders Gigantorhynchida Southwell & Macfie, 1925, Oligacanthorhynchida Petrochenko, 1956 and Moniliformida Schmidt, 1972, were included in our datasets. (1) Gigantorhynchidae Hamann, 1892: Mediorhynchus gallinarum (Bhalerao, 1937) and Mediorhynchus sp.; (2) Moniliformidae Van Cleave, 1924: M. moniliformis (Bremser, 1811), Moniliformis kalahariensis Meyer, 1931, Moniliformis saudi Amin et al., 2016, Moniliformis cryptosaudi Amin et al., 2019 and Moniliformis necromysi sp. n.; and (3) Oligacanthorhynchidae Southwell & Macfie, 1925: Oligacanthorhynchus tortuosa (Leidy, 1850), Macracanthorhynchus hirudinaceus (Pallas, 1781), Macracanthorhynchus ingens (von Linstow, 1879), Prosthenorchis elegans (Diesing, 1851), Prosthenorchis sp., Oncicola luehei (Travassos, 1917), Oncicola venezuelensis Marteau, 1977 and Oncicola sp. All these genera infect mammals, and Mediorhynchus may infect birds as well. As outgroup, we used representative sequences of the classes Palaeacanthocephala and Eoacanthocephala (table 1).
Table 1. Class, family, species, accession number and references of sequences from GenBank used in our phylogenetic analyses with 28S rRNA and MT-COI genes.

The 28S rRNA dataset was aligned using the MAFFT program under default parameters in the Geneious package and manually edited by removing non-complementary extremities. The matrix was subsequently realigned using the Geneious alignment algorithm with the following settings: global alignment with free end gaps, cost matrix of transition/transversion (5.0/1.0) and penalty of 6.0 for both gap opening and extension. The MT-COI dataset was aligned using the MUSCLE program under default parameters in the Geneious package and manually edited by removing non-complementary extremities, followed by a realignment of the sequences using the TranslatorX online software (Abascal et al., Reference Abascal, Zardoya and Telford2010). In both matrices, final manual editing of poorly aligned regions was done using the Mesquite 3.51 software package (Maddison & Maddison, Reference Maddison and Maddison2018).
For both matrices, substitution saturation was assessed using the program DAMBE, version 7.0.35 (Xia, Reference Xia2018) via the Xia test (Xia et al., Reference Xia, Xie, Salemi, Chen and Wang2003; Xia & Lemey, Reference Xia and Lemey2009), with analysis performed on fully resolved sites only, and a graph of transitions and transversions versus JC69 model genetic distances (Jukes & Cantor, Reference Jukes, Cantor and Munro1969). Substitution saturation testing and graphs were also performed separately for each codon position on the MT-COI matrix.
Phylogenetic reconstructions were carried out using maximum parsimony (MP), maximum likelihood (ML) and Bayesian inference (BI) methods, for each matrix (28S rRNA and MT-COI). MP was carried out using PAUP 4.0a164 (Swofford, Reference Swofford2003) heuristic search, with starting trees via stepwise addition, 100 random sequence addition replicates, holding ten trees at each step, and the tree bisection and reconnection branch-swapping algorithm. Node support in MP was assessed by nonparametric bootstrap percentages (MP-BP) after 10,000 pseudoreplications. ML was carried out using PhyML 3.0 (Guindon et al., Reference Guindon, Dufayard, Lefort, Anisimova, Hordijk and Gascuel2010) with tree heuristic search using subtree pruning and regrafting, with ten random starting trees and model selection by the Smart Model Selection (SMS) algorithm (Lefort et al., Reference Lefort, Longueville and Gascuel2017) under the Akaike information criterion (AIC). Node support in ML was assessed by the approximate likelihood-ratio test (aLRT) for branches (Anisimova & Gascuel, Reference Anisimova and Gascuel2006) and by nonparametric bootstrap percentages (ML-BP) after 1000 pseudoreplications. BI was carried out using MrBayes version 3.2.6 (Ronquist et al., Reference Ronquist, Teslenko and van der Mark2012) on the CIPRES Science Gateway platform, version 3.3 (Miller et al., Reference Miller, Pfeiffer and Schwartz2010) with two Markov chain Monte Carlo simulation runs of ten million generations, sampling every 100 generations, and with a burn-in removal of 25%. For the 28S rRNA matrix, the nucleotide substitution model was GTR + I+G. To account for differences between codon positions of MT-COI, independent GTR + I+G models were adopted for each codon position with unlinking of base frequencies and parameters. Node supports were assessed in BI by Bayesian posterior probabilities (BPP). Effective sample sizes (ESS) of parameters were estimated using Tracer, version 1.7.1 (Rambaut et al., Reference Rambaut, Drummond, Xie, Baele and Suchard2018) to assess sampling robustness. We considered values over 100 effectively independent samples to be sufficient. Additionally, the level of variation in MT-COI among sequences of the matrix of different taxa was determined using ML genetic distances calculated in the program PAUP* 4.0a164 (Swofford, Reference Swofford2003).
Results
Moniliformidae Van Cleave, 1924
Moniliformis Travassos, 1915
Moniliformis necromysi sp. n.
Description (based on nine specimens)
General. Pseudosegmented trunk. Proboscis cylindrical and small, compared to the trunk (figs 1a and 2a). Proboscis hooks similar in size and shape in both sexes. Hooks arranged in 12 rows of nine to ten hooks, curved posteriorly and inserted in the tegument (figs 1b and 2d, e). Length of hook blades from anterior in micrometres: 22–12 (17); 18–8 (16); 17–10 (14); 17–11 (15); 16–11 (15); 17–9 (15); 16–10 (14); 14–12 (7); 13–7 (11); 11–7 (10). Apical end of proboscis with no sensory pores (fig. 2b, c). Proboscis receptacle double-walled with muscle fibres spirally arranged in diagonal layers and similar in shape and size in both sexes (fig. 1a). Neck shorter (figs 1a and 2a). Lemnisci flat, reaching the middle of the trunk with some prominent nuclei (fig. 1c). Cephalic ganglion not visualized. Sexual dimorphism absent.

Fig. 1. Line drawing of Moniliformis necromysi sp. n. from Necromys lasiurus: (a) anterior region presents a cylindrical proboscis armed with small hooks, followed by a receptacle proboscis; (b) small hooks from proboscis; (c) leminisci flat, usually in the middle of the body and proboscis receptacle inverted; (d) male body with anterior and posterior testis, with eight cement glands; (e) posterior end of female body; (f) ellipsoid eggs with three membranes. Scale bars: 100 µm.

Fig. 2. External morphology of Moniliformis necromysi sp. n. via SEM: (a) proboscis armed with small hooks; (b) apical view of the proboscis without sensory pore in the apex of the proboscis; (c) apical view of the proboscis without sensory pore in the apex of the proboscis; (d) lateral view of anterior hooks of the proboscis; (e) lateral view of anterior hooks of the proboscis; (f) posterior end of adult female showing a terminal gonopore. Abbreviations: Pb, proboscis; Ho, hook; Gp, gonopore.
Male (based on four adult specimens). Trunk 16.11–43.45 (30.54) long and 0.92–1.21 (1.04) wide. Proboscis 0.30–0.45 (0.37) long and 0.14–0.24 (0.19) wide with 12 rows of nine to ten rootless hooks each. Proboscis receptacle 0.59–0.69 (0.64) by 0.21–0.26 (0.23). Lemnisci 7.95 (n = 1). Testes ellipsoid, in tandem: anterior testis 2.29–2.45 (2.35) by 0.53–0.61 (0.58), posterior testis 1.55–2.24 (2.01) by 0.53–0.66 (0.58) (fig. 1d). Eight ovoid cement glands in 0.64–0.48 (0.57) by 0.39–0.26 (0.32) (fig. 1d). Saefftigen's pouch 1.00–1.32 (1.18).
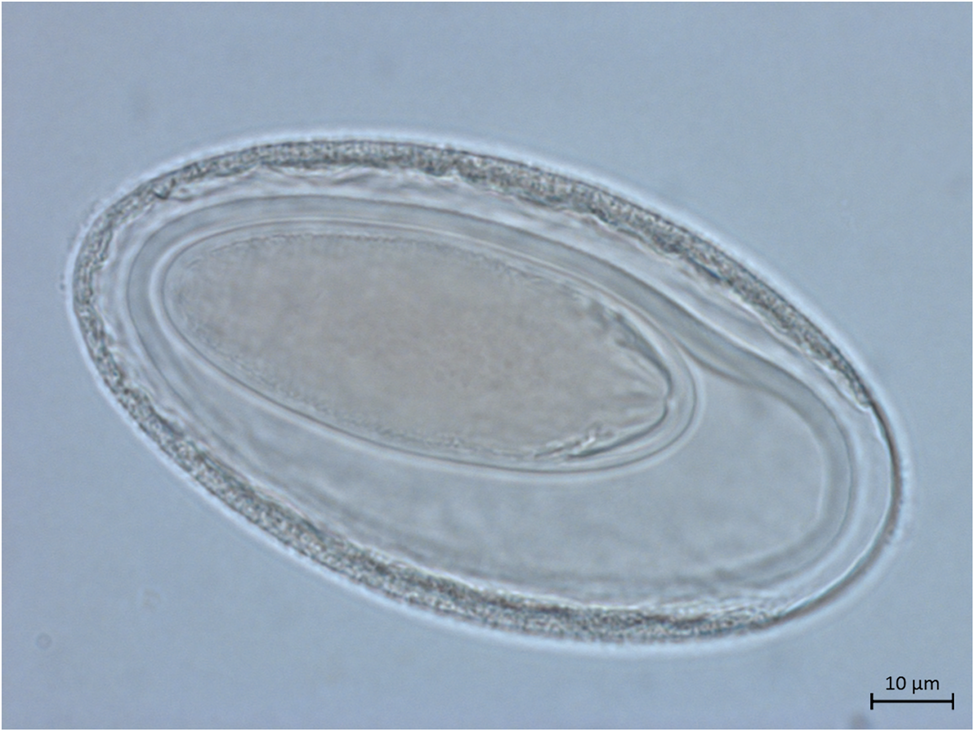
Female (based on five adult specimens). Trunk 26.08–40.84 (30.68) by 0.92–1.66. Proboscis 0.40–0.43 (0.41) by 0.11–0.16 (0.13) with 12 rows of nine to ten hooks each. Proboscis receptacle 0.66–0.71 (0.69) by 0.25–0.27 (0.26). Lemnisci 6.26 (n = 1), mostly covered by eggs. Distance from uterine bell to genital pore including the vagina, uterus and uterine bell 1.33–1.39 (1.36) (n = 2) (fig. 1e). In the lateral vaginal sphincter, two glands or cells associated. Eggs ellipsoid with three membranes showing no prolongation of outer membrane 0.084–0.103 (0.094) long and 0.043–0.070 (0.052) wide (n = 28; figs 1f and 3). Gonopore terminal and round (fig. 2f).

Fig. 3. Light microscopy of egg from adult Moniliformis necromysi sp. n. from Necromys lasiurus.
Taxonomic summary
Type host. Necromys lasiurus (Lund, 1840).
Type locality. Uberlândia (18°55′07″S, 48°17′19″W), Minas Gerais, Brazil.
Site of infection. Small intestine.
Type material. CHIOC 38594 a–c (a, holotype male; b, allotype female; c, paratypes).
Prevalence. 6.9%.
Intensity. 10.3.
Etymology. Species necromysi was named after the host genus.
Remarks
The new species found in the rodent N. lasiurus was identified as belonging to the genus Moniliformis due to the small size of the proboscis compared to the trunk, hooks arranged in longitudinal rows and pseudosegmented trunk, in agreement with Schmidt (Reference Schmidt1972) and Amin (Reference Amin1987). The new species is distinguished from the other moniliformid species by presenting 12 rows of nine to ten small rootless hooks, eight spherical cement glands in pairs and females with terminal rounded gonopore.
Moniliformis necromysi sp. n. is distinguished by the number of rows and hooks per row from M. gracilis (ten rows, 8–9 hooks each), M. tarsi (11–12 rows, 6–7 hooks each), M. convolutes (12 rows, 11–12 hooks each), M. kalahariensis (16 rows, 9–11 (10) hooks each), Moniliformis cestodiformis (16–18 rows, 7–8 hooks each), M. saudi (14 rows, eight hooks each), M. monoechinus (12 rows, 40 hooks each), M. cryptosaudi (13–14 rows, 7–8 hooks each) and M. echinosorex (12–15 rows, 11–13 hooks each). Moniliformis necromysi sp. n. is also distinguished by the host species, because the aforementioned moniliformid species does not parasitize rodents.
According to Amin et al. (Reference Amin, Heckmann, Mohammed and Evans2016) and Martins et al. (Reference Martins, Del Rosario Robles and Navone2017), only eight species have been recorded parasitizing rodents, mainly of the family Muridae, in different geographic regions of the world. The new species is distinguished from other moniliformid species from rodents, particularly by the number of rows and hooks per row, such as M. travassosi (14 rows, 15 hooks each), M. clarki (12–16 rows, 6–8 hooks each), M. spiralis (12 rows, 10–13 hooks each), M. aegyptiacus (12 rows, eight hooks each) and M. siciliensis (14 rows, eight hooks each). Although the range of the number of rows and hooks per row described in M. acomysi (11–16 (12–14) rows, 6–10 hooks each), M. moniliformis (11–14 (12) rows, 9–14 (10–11) hooks each) and Moniliformis amini (12–14 longitudinal rows, 8–12 hooks each) are similar to the new species, the size of the proboscis and eggs distinguished the new species from M. moniliformis (proboscis 320–640 × 130–210 µm and eggs 84–120 × 34–60 µm) and M. amini (proboscis 300–280 × 90–110 µm and eggs 60.73–62.65 × 26.62–27.10 µm). Moreover, M. necromysi sp. n. is distinguished from M. acomysi by the body size (male 30.64 mm and female 30.68 mm in M. necromysi, and male 12–76 mm and female 22–89 mm in M. acomysi), the host species and the geographic distribution, since this species occurs in Acomys cahirinus Geoffroy, 1803 in Cairo, Egypt.
Molecular analyses
Phylogenetic analyses of the 28S rRNA dataset
Our chromatograms assembly resulted in a partial 28S rRNA gene consensus sequence of 760 bp from one adult specimen of M. necromysi sp. n. The resulting matrix comprised 11 taxa and 520 characters, of which 189 characters were constant (proportion = 0.36), 141 were parsimony-uninformative and 190 were parsimony-informative variable characters. The test of Xia provided no evidence for substitution saturation in the 28S rRNA data matrix (supplementary table S1).
The MP analyses resulted in a single most-parsimonious tree of 658 steps length, 0.72 consistency index (CI), 0.28 homoplasy index (HI) and 0.41 rescaled consistency index (RC). The ML best-fit model chosen by SMS in PhyML according to AIC was the GTR + G, with four substitution rate categories and gamma shape parameter 1.009, resulting in a tree with score lnL = −3049.6743. The BI resulted in a mean estimated marginal likelihood of −2964.8606 (standard deviation = 4.0623; median = −2964.5210). Estimated sample sizes were robust for all parameters, above 744.
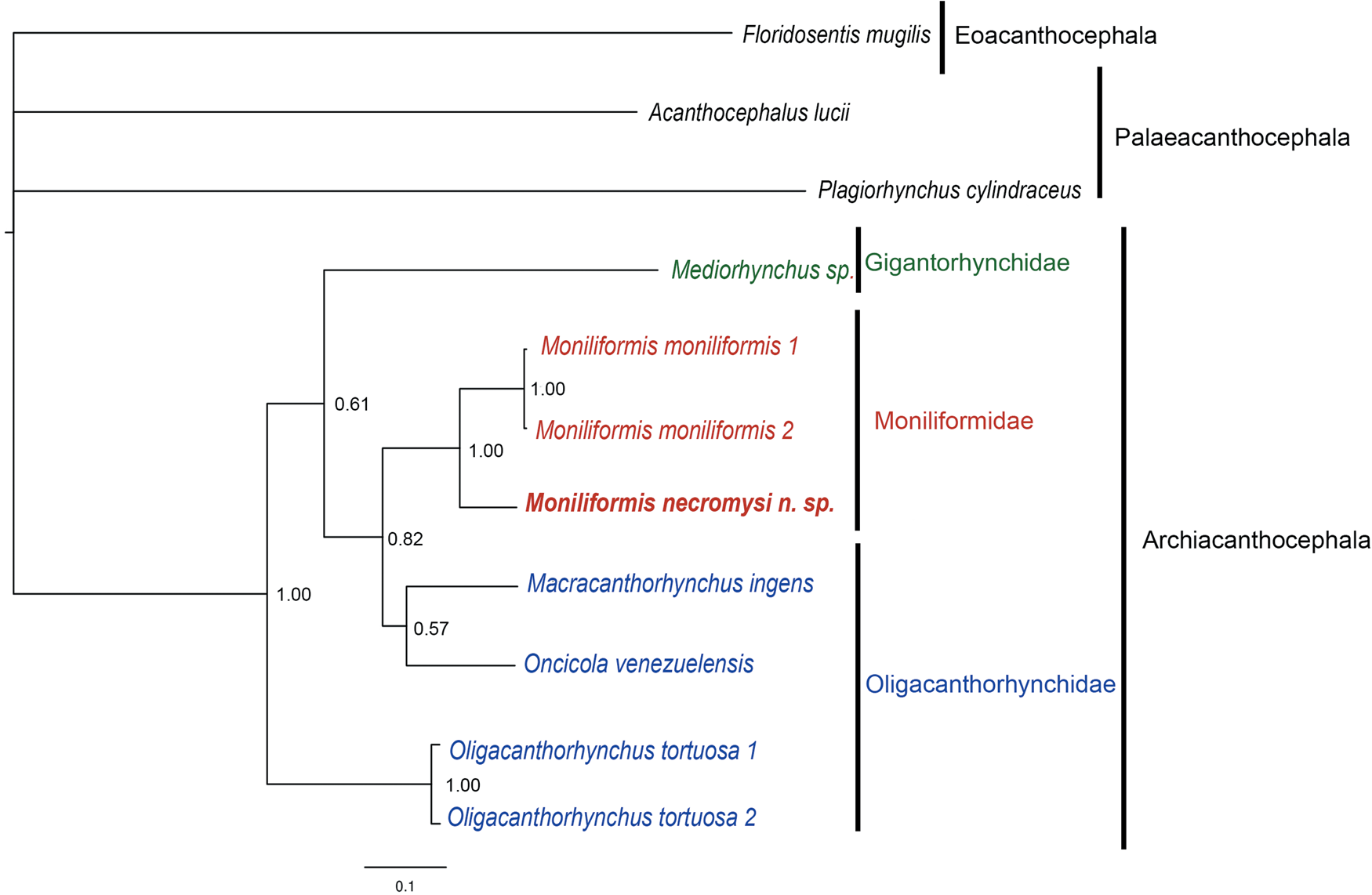
The 28S rRNA MP, ML and BI tree topologies were similar, with little variation in nodes and support values. The BI topology is shown in fig. 4. The class Archiacanthocephala sequences formed a well-supported monophyletic group (MP-BP = 1.00, aLRT = 0.95, ML-BP = 0.92, BPP = 1.00). All analyses also agreed that the 28S rRNA sequences of M. moniliformis grouped together, forming a well-supported monophyletic group (MP-BP = 1.00, aLRT = 0.99, ML-BP = 1.00, BPP = 1.00); and that the sequence of M. necromysi sp. n. was sister to this group with high support values (MP-BP = 1.00, aLRT = 0.98, ML-BP = 0.95, BPP = 1.00), thus conveying the monophyly of the sequences of Moniliformis in our sample. The family Moniliformidae was sister to a monophyletic group, with support values ranging from poor to moderate (MP-BP = 0.59, aLRT = 0.86, ML-BP = 0.45, BPP = 0.82). This group was formed by two oligacanthorhynchid sequences, M. ingens and O. venezuelensis, and was poorly-to-moderately supported (MP-BP = 0.33, aLRT = 0.8, ML-BP = 0.43, BPP = 0.57). The group formed by Moniliformidae and the two oligacanthorhynchids was sister to the family Gigantorhynchidae, represented by the sequence of Mediorhynchus sp., although with little support (MP-BP = 0.47, aLRT = 0.24, ML-BP = 0.37, BPP = 0.61). Finally, the sequences of O. tortuosa formed a well-supported monophyletic group (MP-BP = 1.00, aLRT = 0.99, ML-BP = 1.00, BPP = 1.00), sister to all the other archiacanthocephalans.

Fig. 4. BI phylogenetic reconstructions tree of 28S rRNA gene sequences of Moniliformis necromysi sp. n. from the present study (in bold) and of archiacanthocephalans from GenBank. The classes Palaeacanthocephala and Eoacanthocephala were added as outgroups. Node values are Bayesian posterior probabilities (BPP). Scale bar indicates expected substitutions per site.
Phylogenetic analyses of the MT-COI dataset
Our chromatograms assembly resulted in a partial MT-COI gene consensus sequence of 706 bp from one adult specimen of M. necromysi sp. n. Alignment of sequences resulted in a matrix comprising 23 taxa and 624 characters, of which 184 were constant (proportion = 0.29), 60 were parsimony-uninformative and 380 were parsimony-informative variable characters. The test of Xia & Lemey (Reference Xia and Lemey2009) for substitution saturation provided evidence of saturation only at the third codon positions, whereas, considering the other codon position, there was little saturation in the matrix (supplementary table S1).
The MP analyses resulted in a single most parsimonious tree of 2114 steps length, 0.41 CI, 0.59 HI and 0.19 RC. The ML best-fit model chosen by SMS in PhyML based on AIC was the GTR + G+I, with four substitution rate categories and gamma shape parameter of 0.641, resulting in a tree with score lnL = −8378.5516. For the BI analyses, the mean estimated marginal likelihood was −7954.7109, the standard deviation was 6.5085 and the median was −7954.3670. Estimated sample sizes were robust for all parameters, above 5725.
The ML and BI phylogenies resulted in similar topologies with little variation in nodes and support values, as shown in fig. 5 in the BI topology, whereas the MP tree topology was slightly different, possibly due to saturation. Despite that, the MT-COI sequences of the class Archiacanthocephala formed a well-supported monophyletic group in all topologies (MP-BP = 0.98, aLRT = 1.00, ML-BP = 1.00, BPP = 1.00). In ML and BI topologies, the sequences of the genus Moniliformis formed a poorly-to-moderately supported monophyletic group (aLRT = 0.83, ML-BP = 0.46, BPP = 0.84), being that the sequence of M. moniliformis was the most basal in the family Moniliformidae. However, in the MP topology, the sequence of M. moniliformis did not group with the other sequences of Moniliformis and, rather, was the most basal of all archiacanthocephalan sequences. The sequence of M. necromysi sp. n. was sister to the sequences of M. saudi and M. cryptosaudi, although poorly supported (MP-BP = 0.29, aLRT = 0.64, ML-BP = 0.54, BPP = 0.61). These last two sequences formed a strongly supported clade (MP-BP = 1.00, aLRT = 1.00, ML-BP = 1.00, BPP = 1.00). Nevertheless, in the MP tree topology, the sequences of M. necromysi sp. n. and M. kalahariensis formed a clade, although poorly supported (MP-BP = 0.44). In the ML and BI topologies, M. kalahariensis was sister to the group formed by M. saudi, M. cryptosaudi and M. necromysi sp. n., with support values ranging from poor to strong (MP-BP = 0.60, aLRT = 0.93, ML-BP = 0.80, BPP = 0.66).

Fig. 5. BI phylogenetic reconstructions tree of MT-COI gene sequences of Moniliformis necromysi sp. n. from the present study (in bold) and of archiacanthocephalans from GenBank. The classes Palaeacanthocephala and Eoacanthocephala were added as outgroups. Node values are Bayesian posterior probabilities (BPP). Scale bar indicates expected substitutions per site.
The family Moniliformidae was recovered as sister to the oligacanthorhynchid genera Prosthenorchis Travassos, 1915, Oncicola Travassos, 1916 and Macracanthorhynchus Travassos, 1917, although poorly supported (MP-BP = 0.10, aLRT = 0.54, ML-BP = 0.24, BPP = 0.68), in ML and BI, while not recovered in the MP tree topology. The sequences of Oncicola, represented by O. luehei and Oncicola sp., formed a well-supported monophyletic group (MP-BP = 1.00, aLRT = 0.99, ML-BP = 1.00, BPP = 1.00), sister of the genus Prosthenorchis (MP-BP = 0.99, aLRT = 1.00, ML-BP = 1.00, BPP = 1.00). Prosthenorchis similarly formed a well-supported monophyletic group represented by the sequence of Prosthenorchis sp. and two sequences of P. elegans (MP-BP = 1.00, aLRT = 0.87, ML-BP = 0.91, BPP = 0.99). Prosthenorchis elegans formed a strongly supported clade (MP-BP = 1.00, aLRT = 0.89, ML-BP = 0.91, BPP = 0.97). The group formed by the sequences of Oncicola and Prosthenorchis was recovered as sister to the sequences of Macracanthorhynchus, with support values ranging from poor to strong (MP-BP = 0.26, aLRT = 0.83, ML-BP = 0.48, BPP = 0.99). Macracanthorhynchus, represented by sequences of M. hirudinaceus and M. ingens, formed a poorly-to-strongly-supported clade (MP-BP = 0.37, aLRT = 0.90, ML-BP = 0.67, BPP = 0.99). The sequences of the oligacanthorhynchid O. tortuosa formed a well-supported monophyletic group (MP-BP = 1.00, aLRT = 1.00, ML-BP = 1.00, BPP = 1.00), sister to the group formed by the sequences of moniliformids and of the other oligacanthorhynchids. Additionally, the sequences of the genus Mediorhynchus Van Cleave, 1916, represented by the two sequences of Mediorhynchus sp. and one of M. gallinarum, formed a well-supported clade (MP-BP = 0.85, aLRT = 0.88, ML-BP = 0.86, BPP = 1.00), sister to all the other archiacanthocephalans, except in the MP tree topology.
The pairwise ML-corrected distances over the MT-COI gene sequences between representatives of three classes of acanthocephalans (Archiacanthocephala, Palaeacanthocephala and Eoacanthocephala) are provided in table 2. Pairwise ML distances ranged from 0.003, between M. cryptosaudi and M. saudi, to 0.844, between M. moniliformis (Archiacanthocephala) and Pallisentis celatus (Eoacanthocephala) (mean = 0.485).
Table 2. Pairwise maximum-likelihood genetic distances over MT-COI gene sequences between representatives of the Acanthocephala.

The MT-COI pairwise ML distances between ingroup (Archiacanthocephala) and outgroup (Palaeacanthocephala, Eoacanthocephala) ranged from 0.491, between Plagiorhynchus transversus and M. kalahariensis, to 0.845, between M. moniliformis and P. celatus (mean = 0.656). Within the class Archiacanthocephala, the ML genetic distances ranged from 0.003, between M. cryptosaudi and M. saudi, to 0.542, between O. tortuosa and Mediorhynchus sp. 1 (mean = 0.377). The ML genetic distance between the families Moniliformidae and Gigantorhynchidae ranged from 0.376, between M. kalahariensis and Mediorhynchus sp. 2, to 0.472, between M. moniliformis and M. gallinarum (mean = 0.419); between Moniliformidae and Oligacanthorhynchidae, ranged from 0.323, between M. kalahariensis and Prosthenorchis sp., to 0.454, between O. tortuosa 2 and M. moniliformis (mean = 0.388); and between Gigantorhynchidae and Oligacanthorhynchidae, ranged from 0.367 to 0.542 (mean = 0.437).
The MT-COI pairwise ML distances between species within each genus of archiacanthocephalans were: Mediorhynchus ranged from 0.320, between Mediorhynchus sp. 2 and M. gallinarum, to 0.382, between Mediorhynchus sp. 1 and Mediorhynchus sp. 2 (mean = 0.358); Macracanthorhynchus was 0.370 between M. ingens and M. hirudinaceus; Oncicola was 0.031 between Oncicola sp. and O. luehei; Prosthenorchis ranged from 0.016, between the two species of P. elegans, to 0.088, between Prosthenorchis sp. and P. elegans (mean = 0.060); Oligacanthorhynchus ranged from 0.042, between O. tortuosa 2 and O. tortuosa 3, to 0.269, between O. tortuosa 2 and O. tortuosa 1 (mean = 0.190). Among the sequences of Moniliformis, the interspecific MT-COI pairwise ML distances ranged from 0.003, between M. cryptosaudi and M. saudi, to 0.368, between M. moniliformis and M. necromysi sp. n. (mean = 0.267). The ML genetic distances between M. necromysi sp. n. and the other species of Moniliformis were: 0.243 with M. kalahariensis, 0.254 with M. saudi, 0.273 with M. cryptosaudi and 0.368 with M. moniliformis (mean = 0.285).
Discussion
The genus Moniliformis was proposed by Travassos (Reference Travassos1915) with the type species M. moniliformis (syn. Echinorhynchus moniliformis) (Bremser, 1811). Travassos (Reference Travassos1917) revised the family Gigantorhynchidae and allocated the genus Moniliformis to the subfamily Gigantorhynchinae with two species: M. moniliformis and M. cestodiformis. Southwell & Macfie (Reference Southwell and Macfie1925) validated the family Moniliformidae, as described by Van Cleave (Reference Van Cleave1924), and included the genus Moniliformis with the two valid species considered by Travassos (Reference Travassos1917). Van Cleave (Reference Van Cleave1953) and Yamaguti (Reference Yamaguti1963) agreed with Southwell and Macfie and both allocated the genus Moniliformis to the family Moniliformidae. Later, Schmidt (Reference Schmidt1972) revised the class Archiacanthocephala and proposed the new order Moniliformida. Thereafter, Amin (Reference Amin2013) updated the classification of Acanthocephala and validated the order Moniliformida, with a single family, Moniliformidae, encompassing three genera: Australiformis Schmidt & Edmonds, 1989; Promoniliformis Dollfus & Golvan, 1963; and Moniliformis Travassos, 1915, this last one having 18 valid species. However, Amin et al. (Reference Amin, Heckmann, Mohammed and Evans2016) reviewed the genus Moniliformis and recognized 14 valid species, and described the 15th species: M. saudi from the hedgehog Paraechinus aethiopicus Ehrenberg, 1832 in Saudi Arabia. Martins et al. (Reference Martins, Del Rosario Robles and Navone2017) added another new species to the genus: M. amini from the sigmodontine rodent Abrothrix olivaceus (Waterhouse, 1837) in Argentina. Finally, Amin et al. (Reference Amin, Heckmann, Sharifdini and Albayati2019) described another new species from the long-eared hedgehog Hemiechinus auritus (Gmelin, 1770) in Iraq. To date, the genus Moniliformis comprises 17 species and is characterized by the presence of cylindrical proboscis with numerous and small rootless hooks; pseudosegmented body; lemnisci long and filiform with prominent nuclei; ellipsoid testes; and eight cement glands of spherical shape (Travassos, Reference Travassos1917; Van Cleave, Reference Van Cleave1923, Reference Van Cleave1953; Southwell and Macfie, Reference Southwell and Macfie1925; Yamaguti, Reference Yamaguti1963). Species of Moniliformis are parasites of mammals and occasionally birds (Yamaguti, Reference Yamaguti1963; Amin et al., Reference Amin, Heckmann, Mohammed and Evans2016).
Despite the limited number of GenBank sequences available, we inferred the phylogenetic relationships of representatives of the genus Moniliformis based on the 28S rRNA and MT-COI gene sequences. Our molecular phylogenetic analyses suggested that M. necromysi sp. n. nested within other species of the genus Moniliformis, especially with the sequences of M. saudi and M. cryptosaudi, forming a monophyletic group, and agreed with our conclusion based on morphology. Furthermore, our phylogenetic analyses of the class Archiacanthocephala genera agreed with previous studies recovering the family Moniliformidae as sister to oligacanthorhynchids, with poor-to-strong support (García-Varela & Pérez-Ponce de León, Reference García-Varela, Pérez-Ponce de León, Morand, Krasnov and Littlewood2015; Amin et al., Reference Amin, Heckmann, Mohammed and Evans2016; Amin et al., Reference Amin, Heckmann, Sharifdini and Albayati2019). In addition, interspecific ML distances between the M. necromysi sp. n. sequence and the other sequences of Moniliformis, ranging from 0.243 to 0.368, suggested that it may represent a different species when compared to the inter- and intraspecific ML genetic distances of species within other genera of Archiacanthocephala.
Reports of Acanthocephala in wild rodents are rare, and M. necromysi sp. n. is the first moniliformid species described from a wild rodent host in Brazil. Moreover, this is the first species description of an acanthocephalan for the rodent N. lasiurus. Our findings contribute with morphological and molecular data of this new species and with a new occurrence record, adding more necessary information on the genus Moniliformis and phylum Acanthocephala.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0022149X20000188
Acknowledgements
We are grateful to Ricardo Baptista Schmidt of the image processing and treatment service of the Oswaldo Cruz Institute (FIOCRUZ) for assistance; to the curator of the Helminthological Collection of FIOCRUZ, Dr Marcelo Knoff, for making available the specimens of the collection; to Dr Paulo D'Andrea for providing us with the ICMBio license; to Dr Cibele Bonvicino for taxonomic identification of the rodent specimen; to the staff of the Centro de Controle de Zoonoses de Uberlândia, especially to Jean E. Limongi, and the Universidade Federal de Uberlândia, for assistance with collecting the rodents, and to Alexandro Guterres for helping in the fieldwork. We also thank the postgraduate program in Parasite Biology of the Oswaldo Cruz Institute (PGBP/IOC/Fiocruz).
Financial support
This study was funded by the Brazilian Coordination for the Improvement of Higher Education Personnel (CAPES – finance code 001 to N.A.C.); the Oswaldo Cruz Institute (IOC/FIOCRUZ); the Brazilian National Council for Scientific and Technological Development (CNPq); and the Rio de Janeiro Carlos Chagas Filho Research Support Foundation (FAPERJ – E-26/201.961/2017). R.G. received grants from the Brazilian National Council for Scientific and Technological Development (CNPq – 304355/2018-6).
Conflicts of interest
None.
Ethical standards
The permits for animal capture and handling were granted by the Chico Mendes Institute for Biodiversity Conservation (ICMBio) under authorization number 13373, and by the Ethics Committee on Animal Use of Oswaldo Cruz Institute (CEUA/IOC/FIOCRUZ-RJ) according to licenses L-049/08 and 066/08.