Introduction
Fishermen, managers and fishery scientists face a serious set of problems once the pseudophyllidean cestode Triaenophorus crassus Forel becomes established in a given water body. Although T. crassus does not present a hazard to human health, the parasite causes extensive financial damage to inland fisheries; plerocercoids of this parasite invade the muscle tissue of coregonid and salmonid fishes, which represent the second intermediate hosts (Kuperman, Reference Kuperman1981; Schäperclaus, Reference Schäperclaus1990). Thus, the normally highly valuable flesh cannot be marketed due to its unappetizing appearance. The commercial whitefish fishery in our study site, Lake Achensee, had to be closed due to this parasite. Apart from the considerable economic loss in the largest lake of Tyrol, Austria, T. crassus might also negatively affect survival and growth of these intermediate fish hosts (Rosen & Dick, Reference Rosen and Dick1984; Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999; Sichrowsky et al., Reference Sichrowsky, Schabetsberger, Gassner, Kaiser, Boufana and Psenner2013).
The life cycle of T. crassus involves two intermediate hosts and one definitive host. Copepods, the first intermediate hosts, are infected with procercoids of T. crassus for only one brief period each year (Pulkkinen et al., Reference Pulkkinen, Valtonen, Niemi and Poikola1999; Lahnsteiner et al., Reference Lahnsteiner, Kletzl and Weismann2009; Sichrowsky et al., Reference Sichrowsky, Schabetsberger, Gassner, Kaiser, Boufana and Psenner2013). Coregonid and salmonid fishes are the predominant second intermediate hosts of T. crassus plerocercoids and the parasite is transmitted to them via infected copepods (Petersson, Reference Petersson1971; Kuperman, Reference Kuperman1981; Schäperclaus, Reference Schäperclaus1990). The plerocercoids are able to live for several years in their second intermediate hosts (Reference MillerMiller, 1945b; Rosen & Dick, Reference Rosen and Dick1984) and the life cycle of T. crassus is completed when pike Esox lucius L. feed on infected fish. In the intestine of E. lucius the plerocercoids then develop into mature tapeworms, which release their eggs during spring (Reference MillerMiller, 1945a; Kuperman, Reference Kuperman1981; Lahnsteiner et al., Reference Lahnsteiner, Kletzl and Weismann2009). Triaenophorus nodulosus, a congener of T. crassus, has a wider range of second intermediate hosts and often occurs in the same water body as T. crassus (Reference MillerMiller, 1945a; Kuperman, Reference Kuperman1981; Lahnsteiner et al., Reference Lahnsteiner, Kletzl and Weismann2009; Sichrowsky et al., Reference Sichrowsky, Schabetsberger, Gassner, Kaiser, Boufana and Psenner2013). The life cycle of T. nodulosus is similar to that of T. crassus, with the exception that the plerocercoids of T. nodulosus encyst in the liver of the second intermediate host (Reference MillerMiller, 1945a; Kuperman, Reference Kuperman1981; Schäperclaus, Reference Schäperclaus1990) and thus the fish fillets can still be marketed. Lahnsteiner et al. (Reference Lahnsteiner, Kletzl and Weismann2009) reported that Coregonus sp. is vulnerable to both T. crassus and T. nodulosus.
Triaenophorus crassus infection has been studied in various types of lakes and in different countries (Reference MillerMiller, 1945a; Petersson, Reference Petersson1971; Kuperman, Reference Kuperman1981; Amundsen & Kristoffersen, Reference Amundsen and Kristoffersen1990; Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999; Achleitner et al., Reference Achleitner, Gassner and Schabetsberger2009). Comparing these studies, it is noticeable that the pattern of infection in second intermediate hosts can vary between lakes. It is well known that biotic, physico-chemical and morphometric characteristics of a water body play a fundamental role in structuring the parasite assemblage in coregonid and salmonid fishes (Marcogliese & Cone, Reference Marcogliese and Cone1991; Karvonen & Valtonen, Reference Karvonen and Valtonen2004; Goater et al., Reference Goater, Baldwin and Scrimgeour2005). Especially the trophic status of lakes (Kuperman, Reference Kuperman1981; Molzen, Reference Molzen2005; Brinker & Hamers, Reference Brinker and Hamers2007) and the strength of trophic interactions between the hosts (Petersson, Reference Petersson1971; Kuperman, Reference Kuperman1981; Amundsen & Kristoffersen, Reference Amundsen and Kristoffersen1990) are considered to influence the intensity of Triaenophorus spp. infections. Nevertheless, it is not possible to make reliable predictions of the pattern of T. crassus infection in a given lake, not least because the primary second intermediate host can differ between lakes (Pulkkinen et al., Reference Pulkkinen, Valtonen, Niemi and Poikola1999; Achleitner et al., Reference Achleitner, Gassner and Schabetsberger2009). Predictions are further impeded by the fact that only a few studies (Petersson, Reference Petersson1971; Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999) have investigated in depth the T. crassus infection in different age groups of coregonid and salmonid fishes. Therefore, more detailed information on the infection dynamics and the pattern of T. crassus infections is a prerequisite for further parasite control measures.
The primary aim of this study was to investigate and compare the prevalence, abundance and accumulation of T. crassus in different age groups of whitefish Coregonus lavaretus L. s.l. and Arctic charr Salvelinus umbla (L.) of Achensee. Achleitner et al. (Reference Achleitner, Gassner and Schabetsberger2009) showed that Arctic charr can also act as a primary second intermediate host; nevertheless, this study is the first to compare patterns of T. crassus infection in both fish species within a water body. Additionally we aimed to characterize the risk of T. crassus infection for S. umbla by analysing its feeding strategy during the occurrence of infected copepods. We also assessed whether or not the sympatric congener T. nodulosus, infects C. lavaretus and S. umbla in Lake Achensee.
Materials and methods
Study site
Lake Achensee (47°27′N, 11°42′E, 929 m above sea level, surface area = 6.8 km2, zmax= 133 m, zmean= 67 m) is a holomictic prealpine lake situated in the Limestone Alps of North Tyrol in Austria (Beiwl & Mühlmann, Reference Beiwl and Mühlmann2008). The lake underwent eutrophication in the 1970s and 1980s, but since 1991 total phosphorus concentration has been reduced to less than 3 μg l− 1, mainly due to sewage diversion (unpublished results). Hydropower generation causes water-level fluctuations of up to 6 m, and up to 11.5 m before 2004. Sixteen fish species currently exist in the lake (Gassner & Achleitner, Reference Gassner and Achleitner2006), including S. umbla, pike Esox lucius L., perch Perca fluviatilis L., and the commercially important C. lavaretus. Various stocking activities have been undertaken during the past decades, chiefly to enhance the angling experience (Kandler, fisheries manager of Lake Achensee, pers. comm., 2011).
Collection and examination of fish for Triaenophorus
Sampling of C. lavaretus and S. umbla was carried out 2–3 times a week between 25 May 2011 and 30 July 2011. Benthic and pelagic gillnets with mesh sizes from 10 to 43 mm (knot to knot) were deployed overnight at different locations and set at various depths. Additionally, nine S. umbla were obtained from recreational fishermen between June and July 2011. Fish were identified, measured to the nearest millimetre (total length), weighed ( ± 0.1 g) and their sex was determined. Subsequently livers of all fish and stomachs of S. umbla were fixed in 70% ethanol. In addition, alcohol was injected into each stomach to prevent further digestion. Fish were stored at 4°C for no more than 2 days before the number of parasites was ascertained.
The age of C. lavaretus was determined using scales sampled from the left body side between the dorsal fin and lateral line. The range of S. umbla age groups was estimated based on former length-at-age data for the Achensee population described by Gassner & Achleitner (Reference Gassner and Achleitner2006). A total of 168 C. lavaretus and 120 S. umbla were investigated for the abundance (number of parasites per fish) of T. crassus plerocercoids. Each cooled specimen was skinned and detected plerocercoids were removed, excysted if they were not free, and counted. Starting on one side, a thin fillet of approximately 2 mm thickness was cut along the anterioposterior axis and the newly discovered plerocercoids were removed, excysted and counted. Cutting and investigating the fillets was continued, until all muscle tissue of one side was examined. Subsequently, the same procedure was applied on the other side. To ensure that no plerocercoid was counted twice, their scolices were used to determine the abundance. In the end, the fillets were cut into 5 mm stripes to reveal very small cysts that may have been overlooked. In addition to counting, the condition of T. crassus was categorized into intact and degenerated. Plerocercoids were classified as degenerated if they were shrunken, fragmented or their integuments started to lyse. Cysts were termed as degenerated, if they were empty, solid or contained only remains of plerocercoids. A random sample of about 200 plerocercoids located in the muscles of the hosts was fixed in 70% ethanol and studied under a light microscope at 200-fold to 400-fold magnification. Prevalence of T. crassus was determined with the same set of specimens that were used for estimating abundance. For C. lavaretus, an additional 54 individuals were investigated. Furthermore, livers of all specimens were studied for infection with T. nodulosus. For this purpose the livers were cut into strips of approximately 2 mm thickness and examined.
In Achensee, C. lavaretus can be separated into a pelagic and a benthic morph. The pelagic whitefish represent the predominant portion of the whitefish stock and feed primarily on zooplankton, while the few benthic whitefish mainly feed on macrozoobenthos (Kandler, fisheries manager of Lake Achensee, pers. comm., 2011). The accumulation of T. crassus with fish age was studied by analysing 160 pelagic and 8 benthic whitefish specimens from 1 to 8 years in age. Benthic and pelagic whitefish of the same age were pooled, as the benthic specimens had similar T. crassus burdens as the pelagic ones. The newly incorporated plerocercoids of the season 2011 were not included in the fish-age-specific abundance of T. crassus. Thus, the reported abundance of plerocercoids reflects the state after the last Triaenophorus infection in 2010. As the investigation for the T. crassus abundance of C. lavaretus was performed during a period when most plerocercoids of 2011 were not yet developed or at an early stage of development, it was straightforward to separate them from plerocercoids of previous years. The annual T. crassus accumulation in C. lavaretus was computed by dividing the plerocercoid abundance by the number of times fish had been exposed to T. crassus invasions (Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999). The number of annual invasions is indicated by fish age increased by one.
Dietary analysis of fish
The stomach samples of each month were pooled, as no remarkable differences were apparent between the different fish length classes. After the stomachs were opened, their total fullness was estimated visually on a percentage scale of 0% (empty) to 100% (full). Stomachs with no identifiable prey were excluded from further analyses, so that 68 remaining samples were investigated in detail. Prey were identified, counted and their numerical contribution (per cent abundance) to the diet was determined. The zooplankton items were identified to the species level and counted. If necessary, subsampling was applied. The other prey items were grouped into chironomids, other insects, fish and molluscs.
In order to assess the feeding strategy of S. umbla, a graphical analysis suggested by Amundsen et al. (Reference Amundsen, Gabler and Staldvik1996) was used. Therefore, the prey specific abundance (P i) was plotted against the frequency of occurrence (F i) on a two-dimensional graph. The two indices can be described by the equations:
 $$\begin{eqnarray} P _{ i } = (\sum S _{ i }/\sum S _{ ti })\times 100 \end{eqnarray}$$
$$\begin{eqnarray} P _{ i } = (\sum S _{ i }/\sum S _{ ti })\times 100 \end{eqnarray}$$
 $$\begin{eqnarray} F _{ i } = N _{ i }/ N \end{eqnarray}$$
$$\begin{eqnarray} F _{ i } = N _{ i }/ N \end{eqnarray}$$
where S i is the number of prey i found in the stomach contents, S ti is the total amount of food items found in only those predators with prey i in their stomachs, N i is the number of predators with prey i in their stomachs, and N is the total amount of predators with food items in their stomachs (Amundsen et al., Reference Amundsen, Gabler and Staldvik1996). The position of prey data points in the graph produced indicates the prey importance and provides insight into the feeding strategy of the predator. A prey data point positioned in the upper left indicates that some individual predators are specialized on that particular prey. A prey data point located in the upper right, however, points out that the whole population is specialized on a given prey. Prey data points positioned in the lower part of the graph reflect a more generalistic feeding strategy. Points will be located in the lower right part of the graph if the corresponding prey are eaten occasionally by most predators. By contrast, points will be in the lower left if only a few predators sporadically consume the given prey. Amundsen et al. (Reference Amundsen, Gabler and Staldvik1996) provide a more precise description of the method.
Data analysis
Mean length-at-age for the pelagic C. lavaretus was modelled using the von Bertalanffy growth model. Bootstrap methods were used to construct 95% confidence bounds and 95% prediction bounds for the model. Due to the small sample size, the growth of the benthic C. lavaretus was not modelled. The relationship between abundance of T. crassus plerocercoids and age of C. lavaretus was analysed with Spearman's rank correlation coefficient. Differences between age groups of C. lavaretus in annual accumulation of T. crassus were tested using a one-way analysis of variance (ANOVA; general linear model, 95% confidence). Data were tested for normal distribution and equal variance using the Kolmogorov–Smirnov test and the Levene's test, respectively. A Fisher's exact test was conducted to reveal differences in prevalence of T. crassus between C. lavaretus and S. umbla. For the test, the age groups were pooled within each species. The parasitological terminology of prevalence and abundance follows Bush et al. (Reference Bush, Lafferty, Lotz and Shostak1997).
Results
Infection levels of Triaenophorus in fish
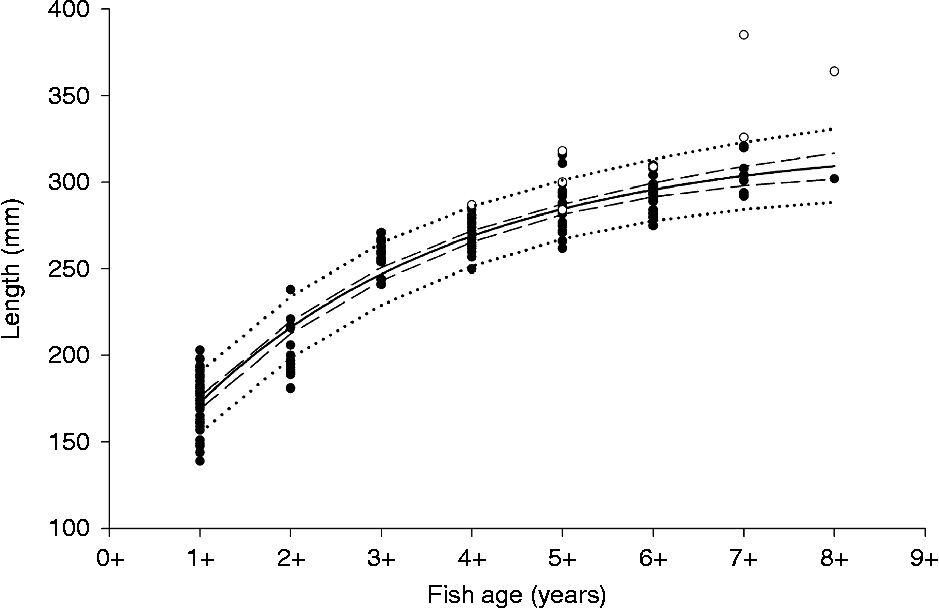
All examined C. lavaretus were infected with T. crassus (n= 222). In S. umbla, however, only 15.8% of the investigated individuals harboured T. crassus (n= 120). The prevalence of this parasite differed significantly between the two species (Fisher's exact test: df = 1, P< 0.001). The congener T. nodulosus was not detected in C. lavaretus (n= 222) or S. umbla (n= 120). The mean length of examined pelagic C. lavaretus ranged from 174.6 mm (1-year-old fish) to 302.0 mm (8-year-old fish). Pelagic C. lavaretus showed rapid growth during the first 3 years of life, until they reached maturity at the age of 3+ (fig. 1). In C. lavaretus older than 3+, yearly length increments were small and growth was substantially reduced.

Fig. 1 Fish length/age values of pelagic (●) and benthic (○) Coregonus lavaretus from Lake Achensee during June/July 2011; the growth of pelagic fish was modelled using the von Bertalanffy growth model (f= 323.5(1 − e− 0.34(x − ( − 1.26)) and 95% confidence (![]() ) and 95% prediction intervals (
) and 95% prediction intervals (![]() ) were computed using bootstrap methods.
) were computed using bootstrap methods.
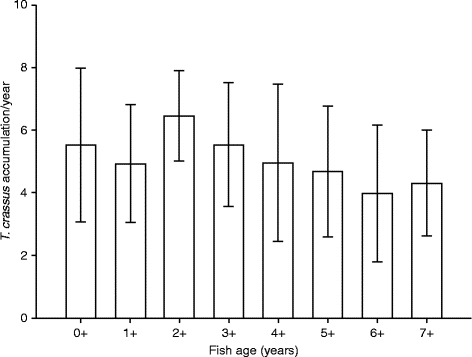
Mean abundance of T. crassus plerocercoids in C. lavaretus (pelagic and benthic morph pooled) significantly increased with fish age (fig. 2; Spearman K s= 0.74, df = 166, P< 0.001). The increase was distinct in the range of age groups 0+ to 3+, while it levelled off in older fish. Fish belonging to the age group 2+ were already infected with 19.4 plerocercoids on average, while older specimens frequently harboured about 30 plerocercoids. The maximum individual infection with T. crassus consistently increased with fish age, until it culminated at a maximum of 50 plerocercoids in the age groups 4+ and 5+. The mean annual accumulation of T. crassus plerocercoids (intake of plerocercoids per year) in C. lavaretus did not significantly differ between age groups (fig. 3; one-way ANOVA (GLM): F= 1.65, df = 167, P= 0.125). The mean accumulation in all age groups was 5.2 (SD = 2.2) plerocercoids per year. No increase of T. crassus with fish age or total length was apparent in S. umbla. The majority of infected fish harboured one or two parasites, while the maximum individual infection remained at a low level of three plerocercoids (table 1). The length of S. umbla ranged from 92.0 to 463.0 mm, which corresponded approximately to age groups 2+ and 9+.
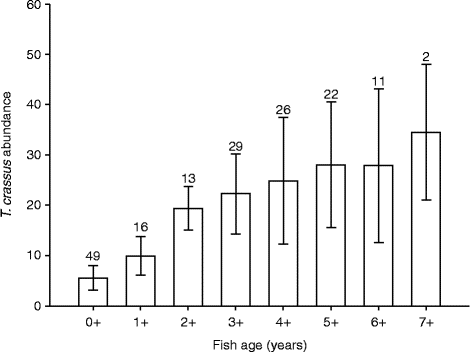
Fig. 2 Mean abundance values ( ± SD) of Triaenophorus crassus in Coregonus lavaretus from Lake Achensee, relative to fish age. Numbers of fish studied within each age group are indicated above the bars.
Fig. 3 Mean annual accumulation of T. crassus plerocercoids ( ± SD) for the age groups of Coregonus lavaretus from Lake Achensee. The numbers of investigated fish are as in fig. 2.
Table 1 The frequency of occurrence (%) of plerocercoids of Triaenophorus crassus as encysted or free and intact or degenerated in 168 Coregonus lavaretus and 120 Salvelinus umbla from Lake Achensee in June/July 2011.
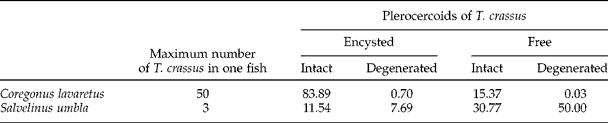
In C. lavaretus a large proportion of T. crassus plerocercoids were encysted, whereas free (not encysted) plerocercoids were scarce (table 1). Overall, most plerocercoids were intact and only a small portion of them were degenerated. The former were frequently about 20 cm long, whereas the latter were much shorter, mainly encysted and often had a mushy appearance. Some degenerated cysts were yellowish, solid and contained only remains of plerocercoids. In infected S. umbla, however, both intact and degenerated plerocercoids were of the same size and rarely exceeded 12 cm. Moreover, degenerated specimens made up more than half of all harboured plerocercoids in S. umbla (table 1). In contrast to C. lavaretus, the vast majority of degenerated plerocercoids were not encysted.
Feeding strategy of fish
No major differences in the diet between length classes of S. umbla were detected in June and July. On the other hand, the proportion (per cent abundance) of food items in S. umbla varied strongly over time (table 2). Chironomids, mainly pupae, dominated the diet in June and became rare in July. Bosmina sp. was the most consumed zooplankton species in June, whereas Daphnia sp. made up more than 90% of the entire diet in July. Cyclops sp. was completely missing from the stomach content in June and not very abundant in July. Likewise Cyclops, fish, molluscs and insects (excluding chironomids) were not frequent in the diet of S. umbla. It is likely that oligochaetes were not detected because they are digested rapidly. The feeding strategy graphs (fig. 4) reveal a specialization at the population level of S. umbla in both June and July. This population specialization was reflected by one prey point for each month positioned in the upper right part of the diagram. The population specialized on chironomids and Daphnia sp. in June and July, respectively. The other food items, including Cyclops sp., with their prey points in the lower left part, were only eaten occasionally at a predominantly individual level. Besides the specialization at the population level, a slight individual feeding specialization on Bosmina sp. existed in June.
Table 2 The proportion of food contents (%) in the stomach of 40 (June) and 28 (July) S. umbla from Lake Achensee.
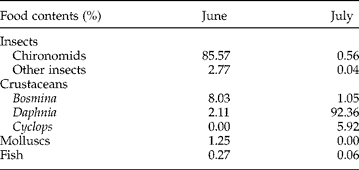
Fig. 4 Prey specific abundance (%) against the frequency of occurrence within the stomach of (a) 40 Salvelinus umbla in June and (b) 28 S. umbla in July 2011 from Lake Achensee; Bosmina sp. (▲), Cyclops sp. (△), fish (■), Daphnia sp. (□), chironomids (●), other insects (○) and molluscs (●).
Discussion
One of the main goals of the present study was to investigate the pattern of T. crassus infection in C. lavaretus and S. umbla of Achensee. Both fish species are known to be second intermediate hosts for T. crassus (Reference MillerMiller, 1945b; Kuperman, Reference Kuperman1981; Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999; Achleitner et al., Reference Achleitner, Gassner and Schabetsberger2009). Coregonus lavaretus of Achensee exhibited extremely heavy T. crassus burdens. The prevalence of T. crassus plerocercoids was 100% in all examined age groups. Such a high prevalence has only been recorded for older age groups, while in fish aged 0+ and 1+ the prevalence never exceeded 45% (Reference MillerMiller, 1945b; Petersson, Reference Petersson1971; Amundsen & Kristoffersen, Reference Amundsen and Kristoffersen1990; Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999). The high prevalence of T. crassus in younger C. lavaretus of Achensee is, therefore, exceptional. The high burden of T. crassus is also reflected in a remarkably high load of plerocercoids in all age groups. The mean abundance of T. crassus in C. lavaretus significantly increased with fish age, which is in line with the results of other studies (Petersson, Reference Petersson1971; Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999). However, we showed much higher fish-age-specific abundances of T. crassus than reported in previous work. Dick & Rosen (Reference Dick and Rosen1982) observed that young-of-the-year whitefish contained one or two plerocercoids and the mean abundance of T. crassus for 1- to 4-year-old whitefish was described to be in the range of 0–8 plerocercoids (Reference MillerMiller, 1945b; Pulkkinen et al., Reference Pulkkinen, Valtonen, Niemi and Poikola1999). In Achensee, however, 0+ aged whitefish already harboured 5.5 T. crassus on average. The mean abundance rapidly increased to 19.4 plerocercoids per fish in the age group 2+ and reached more than 27 plerocercoids in older fish. Pulkkinen & Valtonen (Reference Pulkkinen and Valtonen1999) noticed such high loads of T. crassus only in whitefish aged 6 years and older, whereas mean abundances in 1- to 5-year-old fish in their samples stayed below nine plerocercoids.
This mass infection of T. crassus may be the result of several factors. First, the immune defence of C. lavaretus against this parasite seems to be weak in Achensee. According to Pulkkinen & Valtonen (Reference Pulkkinen and Valtonen1999), the annual accumulation of T. crassus in whitefish of Lake Puruvesi remained at a low level for young age groups, until the intake of parasites almost doubled in older age groups. One possible explanation for the increased accumulation is a breakdown of immune defence, once a critical number of parasites per fish is exceeded (Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999). In contrast, the intake of T. crassus in C. lavaretus of Achensee constantly remained at a very high level throughout all age groups, underscoring a potentially weak immune defence. However, an experimental infection study would have been needed to clarify this claim, as it is unknown how many infected copepods had been consumed by the fish and, therefore, the efficiency of the immune defence cannot be rated.
Second, the ultra-oligotrophic conditions of Lake Achensee very likely contribute to the high burdens of T. crassus. It is known that the transmission of Triaenophorus sp. increases in water bodies with low nutrient concentrations because of the relative dominance of (infected) copepods compared to cladocerans (Watson & Lawler, Reference Watson and Lawler1965; Molzen, Reference Molzen2005). Furthermore, the zooplankton biomass is low in oligotrophic lakes, so that planktivorous fish have to prey additionally on copepods beside the preferred cladocerans to meet their energy demands (Watson & Lawler, Reference Watson and Lawler1965; Kuperman, Reference Kuperman1981; Molzen, Reference Molzen2005; Brinker & Hamers, Reference Brinker and Hamers2007).
Third, the mass infection of T. crassus may be related partly to the water-level fluctuations in Achensee. Petersson (Reference Petersson1971) highlighted that in lakes with artificial water-level fluctuations both pelagic and benthic whitefish species had a high infection with T. crassus, although the macrozoobenthos-dominated diet of the benthic species should have counteracted a high invasion. He argued that the altered environment of regulated lakes favours especially benthic whitefish to evolve a pronounced planktivorous feeding habit and thus increases the infection with T. crassus. In fact, pelagic and benthic C. lavaretus of Achensee had similar T. crassus burdens, which reinforces the argument of Petersson (Reference Petersson1971) that water-level fluctuations may favour a shift from benthic to planktonic feeding behaviour. Nevertheless, Gassner & Achleitner (Reference Gassner and Achleitner2006) and our own observations in 2011 illustrate that the vast majority of C. lavaretus in Achensee reside in the pelagic zone and thus mainly utilize crustacean zooplankton (Schulz, Reference Schulz1979; Anegg et al., Reference Anegg, Psenner and Tartarotti2014). It remains unclear to what extent the water-level fluctuations enhance the degree of plankton feeding and the risk of T. crassus infection. Moreover, it is reasonable to assume that E. lucius in Achensee, as definitive hosts, are co-responsible for the high T. crassus burdens in C. lavaretus. In fact, it seems that over the past few decades the population has increased (Kandler, fisheries manager of Lake Achensee, pers. comm., 2011; authors' observations, 2011) which facilitates the transmission of the parasite.
In S. umbla the prevalence of T. crassus plerocercoids was significantly lower than in C. lavaretus. Less than 16% of S. umbla were infected and most infected fish only harboured one or two plerocercoids. Furthermore, no increase of T. crassus with fish age or total length was apparent. These findings are quite surprising in the light of the high burdens of T. crassus in C. lavaretus, especially because Achleitner et al. (Reference Achleitner, Gassner and Schabetsberger2009) documented a mass infection of T. crassus in S. umbla of Lake Grundlsee, another Austrian lake. Both fish species are vulnerable to infection with T. crassus and, although the parasite is heavily represented in Lake Achensee, S. umbla is only marginally infected. Infection with T. crassus plerocercoids requires a trophic relationship between the two intermediate hosts. Due to the strong dietary plasticity of Arctic charr (Hofer & Medgyesy, Reference Hofer and Medgyesy1997; Amundsen et al., Reference Amundsen, Knudsen and Klemetsen2008; Eloranta et al., Reference Eloranta, Kahilainen and Jones2010, Reference Eloranta, Siwertsson, Knudsen and Amundsen2011) trophic interactions can vary strongly between and within populations. Some populations exert a pronounced preference for crustacean zooplankton, which implicates a higher risk for infections with copepod-transmitted parasites (Bérubé & Curtis, Reference Bérubé and Curtis1986; Knudsen et al., Reference Knudsen, Amundsen, Nilsen, Kristoffersen and Klemetsen2008, Reference Knudsen, Amundsen and Klemetsen2010), particularly as some Arctic charr populations or individuals specialize on copepods (Dahl-Hansen et al., Reference Dahl-Hansen, Rubach and Klemetsen1994; Knudsen et al., Reference Knudsen, Klemetsen and Staldvik1996).
In order to assess the risk of T. crassus infection for S. umbla in Achensee, their feeding strategy was studied in June and July. This time slot was selected because in Achensee copepods are infected by Triaenophorus spp. between June and the beginning of August (Anegg et al., Reference Anegg, Psenner and Tartarotti2014). The detected procercoids in August were most likely T. nodulosus (with respect to the findings of Reference MillerMiller (1945a), Lahnsteiner et al. (Reference Lahnsteiner, Kletzl and Weismann2009) and Sichrowsky et al. (Reference Sichrowsky, Schabetsberger, Gassner, Kaiser, Boufana and Psenner2013)). Copepods were completely missing from the stomach contents of S. umbla in June. In July, only a small amount of copepods, in particular Cyclops sp., was consumed by a few individuals, while the vast majority of the population specialized on Daphnia sp. Thus, it can be suggested that the feeding habit of the population is probably a major cause for their marginal infection with T. crassus. Although the diet of S. umbla was only studied during one period of T. crassus transmission, there is evidence that copepods were also avoided in other years (unpublished results). This is congruent with Knudsen et al. (Reference Knudsen, Klemetsen and Staldvik1996), whose findings indicate that Arctic charr maintain dietary specializations over years. Besides feeding strategy, habitat choice of the S. umbla population may protect this species from high infection with T. crassus. According to Watson & Lawler (Reference Watson and Lawler1965), copepods infected with Triaenophorus spp. are most abundant in shallow waters near the shore. Salvelinus umbla of Achensee, however, primarily inhabits the benthal area of the deep-water zone (Gassner & Achleitner, Reference Gassner and Achleitner2006; authors' observations, 2011).
Apart from the low T. crassus infection in S. umbla, the high rate of degenerated plerocercoids is noteworthy. More than half of all harboured plerocercoids in S. umbla were degenerated, whereas in C. lavaretus more than 99% of the plerocercoids were intact. It appears that T. crassus is disturbed in its development in S. umbla, as many young plerocercoids began to degenerate before they were encysted. One possible explanation for the low infectivity of T. crassus for S. umbla in Achensee could be a high host specificity of the local parasite population for C. lavaretus and not for S. umbla. Pulkkinen et al. (Reference Pulkkinen, Valtonen, Niemi and Poikola1999) confirmed that the host specificity of this parasite can vary even locally between Coregonus species. Another reason might be an effective immune defence of the S. umbla population against this parasite.
In conclusion, this study demonstrates that a remarkably high prevalence and abundance of T. crassus in C. lavaretus already occurs in younger age groups. In contrast, previous studies have merely revealed high burdens of T. crassus in older age groups (Reference MillerMiller, 1945b; Dick & Rosen, Reference Dick and Rosen1982; Amundsen & Kristoffersen, Reference Amundsen and Kristoffersen1990; Pulkkinen & Valtonen, Reference Pulkkinen and Valtonen1999). Even though C. lavaretus exhibited an exceptionally high T. crassus infection status, the S. umbla population of Achensee was marginally infected. With respect to the mass infection of S. umbla in another Austrian lake (Achleitner et al., Reference Achleitner, Gassner and Schabetsberger2009), the present results corroborate the great variability in the pattern of T. crassus infection. Thus, one should be particularly cautious in generalizing infection characteristics of this parasite without detailed investigations. Based upon our findings, we recommend reducing the amount of older C. lavaretus in Achensee. These age groups have an extremely high abundance of T. crassus plerocercoids, hence they pose a high risk for transmission of this parasite to Esox lucius, the definitive host. Simultaneously, a strong decimation of pike will be required to reduce the Triaenophorus spp. populations (Amundsen & Kristoffersen, Reference Amundsen and Kristoffersen1990; Achleitner et al., Reference Achleitner, Gassner and Schabetsberger2009). Future research should study in depth the effects of such high T. crassus burdens on growth, behaviour and survival of second intermediate hosts under natural conditions.
Acknowledgements
We thank A. Kandler, fisheries manager of Lake Achensee, for field assistance and background information. We are also grateful to R. Konecny for his valuable comments on the draft.
Financial support
This work was supported financially by the Tiroler Wasserkraft AG, the Tyrolean Government and the city of Innsbruck.
Conflict of interest
None.