Introduction
A balanced maternal diet during gestation and lactation is needed to maintain a full-term pregnancy and cover increased maternal and placental–fetal metabolism and growth. Reference Kominiarek and Rajan1 According to the “Recommended Dietary Allowances”, caloric intake should increase by approximately 300 kcal/day during pregnancy and recommended protein intake is 60 g/day (1.1 g of protein/kg/day during pregnancy as compared to 0.8 g of protein/kg/day for non-pregnant states). Reference Kominiarek and Rajan1 If these intakes are not attained, the resulting maternal malnutrition may impair placentation, with resulting changes in placental size, morphology, and blood flow. Reference Belkacemi, Nelson, Desai and Ross2 To the fetus, compromised supply of nutrients affects organogenesis, growth, and neural development resulting in intrauterine growth restriction (IUGR) and newborns with low birth weight, two conditions predisposing to development of metabolic disease, neural disturbancies, and behavioral disturbancies. Reference de Brito Alves, de Oliveira, Ferreira, Barros, Nogueira, Alves, Vidal, Leandro, Lagranha, Pirola and da Costa-Silva3–Reference Dos Santos, Moura Dos Santos, Almeida, Nobre, Nobre, Ferreira, Gomes, Antonio Ribeiro Maia and Leandro5 In the central nervous system (CNS), the existence of a close relationship between the maternal nutritional status, brain development, and cognitive functions has been proposed. Reference Veena, Gale, Krishnaveni, Kehoe, Srinivasan and Fall6
The maternal phenotype, or “maternal capital”, is the primary influence on early nutrition and developmental trajectory of offspring. Reference Wells7 In undernourished mothers (8% protein), a reduced body weight gain, anemia, and microalbuminuria have been observed during gestation. Reference Fragoso, Lira, Chagas, Lucena Cavalcanti, Beserra, de Santana-Muniz, Bento-Santos, Martins, Pirola, da Silva Aragao and Leandro4,Reference Fidalgo, Falcao-Tebas, Bento-Santos, de Oliveira, Nogueira-Neto, de Moura, Lisboa, de Castro and Leandro8,Reference de Brito Alves, Nogueira, de Oliveira, da Silva, Wanderley, Leandro and Costa-Silva9 Undernutrition results in placenta with low weight and size, reduced blood flow and proliferation of Langhan’s cell of the villi, and hypoplasia calcifications of villi. Reference Belkacemi, Nelson, Desai and Ross2,Reference Tarrade, Panchenko, Junien and Gabory10 In the fetus, maternal protein restriction can affect the growth and development, proliferation, migration, differentiation, synaptogenesis, myelination, and the expression of neurotrophic growth factors (neurotrophins). Reference Amaral, Jakovcevski, McGaughy, Calderwood, Mokler, Rushmore, Galler, Akbarian and Rosene11,Reference Gonzalez-Maciel, Romero-Velazquez, Reynoso-Robles, Uribe-Escamilla, Vargas-Sanchez, de la Garza-Montano and Alfaro-Rodriguez12 Most of the evidence suggests that, during pregnancy and lactation, the nutritional investment is mainly directed to the placenta–fetus axis, and to the offspring during suckling, at the expenses of the neurobiology of undernourished mothers. Thus, offspring are exposured to patterns of nutritional investment that may maximize their adaptation even in the presence of maternal nutritional constraints.
Brain-derived neurotrophic factor (BDNF) is an important neurotrophin influencing almost all aspects of the CNS development, such as neuronal proliferation, migration and survival, synapse formation, axonal and dendritic plasticity. Reference Garces, Sanchez, Torres-Sierra, Ruiz-Parra, Angel-Muller, Alzate, Sanchez, Gomez, Romero, Castaneda, Sanchez-Rebordelo, Dieguez, Nogueiras and Caminos13 BDNF also has been shown to exert an important role during embryo implantation, placental development, and control of fetal growth in mice. Reference Mayeur, Silhol, Moitrot, Barbaux, Breton, Gabory, Vaiman, Dutriez-Casteloot, Fajardy, Vambergue, Tapia-Arancibia, Bastide, Storme, Junien, Vieau and Lesage14 Maternal food restriciton (50% of the food intake of control mothers) induced IUGR of fetuses at term and decreased the placental BDNF messenger RNA (mRNA), and its functional receptor, the tyrosine kinase receptor B (TrkB/NTRK2) protein expression in the brain of growth-restricted rat offspring. Reference Mayeur, Silhol, Moitrot, Barbaux, Breton, Gabory, Vaiman, Dutriez-Casteloot, Fajardy, Vambergue, Tapia-Arancibia, Bastide, Storme, Junien, Vieau and Lesage14 Maternal low-protein (LP) diet (7 to 8% casein) is related to fewer synapses, synaptic structural changes, decreased dendritic spines and arborization (complexity of branching projections) and decreased insulin-like growth factor (IGF-1) and insulin-like growth factor receptor (IGF-1r) levels. Reference Gonzalez-Maciel, Romero-Velazquez, Reynoso-Robles, Uribe-Escamilla, Vargas-Sanchez, de la Garza-Montano and Alfaro-Rodriguez12,Reference Jones and Dyson15,Reference Ikeda, Shoji, Suganuma, Ohkawa, Kantake, Murano, Sakuraya and Shimizu16 However, little is known about the gene expression of neurotrophic factors in different areas of mother’s brain and placenta upon condition of protein restriction.
Many of the effects of maternal protein restriction are deemed to be permanent, though some degree of plasticity may be expected by the simultaneous exposure to a stimulating and enriched condition, such as physical activity. Previous studies have shown that maternal physical activity may mitigate or even prevent impairments of neuronal growth and development induced by maternal protein restriction. Reference Fragoso, Lira, Chagas, Lucena Cavalcanti, Beserra, de Santana-Muniz, Bento-Santos, Martins, Pirola, da Silva Aragao and Leandro4,Reference Fidalgo, Falcao-Tebas, Bento-Santos, de Oliveira, Nogueira-Neto, de Moura, Lisboa, de Castro and Leandro8,Reference Leandro, Fidalgo, Bento-Santos, Falcao-Tebas, Vasconcelos, Manhaes-de-Castro, Carpinelli, Hirabara and Curi17 Based on these findings, healthy pregnant women without medical contraindications should be encouraged to engage in regular physical activity at least 150 min per week (i.e., 20–30 min/day on most or all days of the week) of moderate aerobic intensity. Reference Ferrari and Graf18 In humans, infants born from active mothers during pregnancy (three times per week, at least 20 min at 55% of their maximal aerobic capacity) showed a better response to sound discrimination and auditory memory as measured by electroencephalography. Reference Labonte-Lemoyne, Curnier and Ellemberg19 In rats, pups born from dams which were active throughout pregnancy showed an increased amount of neuronal and non-neuronal cells in the hippocampus, improved cognitive functions (habituation behavior and spatial learning), and enhanced memory as tested using a novel object recognition paradigm. Reference Robinson and Bucci20,Reference Gomes da Silva, de Almeida, Fernandes, Lopim, Cabral, Scerni, de Oliveira-Pinto, Lent and Arida21
The concept of “spontaneous physical activity” refers to physical movement motivated by psychological, behavioral, or physical rewarding and may be related to a survival need or directly motivated by any other external factor. Reference Santana Muniz, Beserra, da Silva Gde, Fragoso, Lira Ade, Nascimento, Manhaes de Castro and Leandro22 On the other hand, “physical exercise” refers to physical activity performed on a voluntary and systematic way, with control of intensity (maximal consumption of oxygen or plasma lactate concentration), frequency, duration, and type (anaerobic or aerobic). Reference Leandro, Levada, Hirabara, Manhães-de-Castro, De-Castro, Curi and Pithon-Curi23
In a previous study, spontaneous physical activity on a running wheel before and throughout pregnancy attenuated the effects of a maternal LP diet on patterns of locomotor activity of offspring rats at 60 days of age. Reference Fragoso, Lira, Chagas, Lucena Cavalcanti, Beserra, de Santana-Muniz, Bento-Santos, Martins, Pirola, da Silva Aragao and Leandro4 However, molecular events in the brain and placenta such as the gene expression of neurotrophins in both mother and fetuses were not explored.
In line with the reported contribution of BDNF, neurothrophin 4 (NTF4/NT-4), TrkB/NTRK2, IGF-1, and IGF-1r in placental development and in the control of fetal growth, the present study tested the hypothesis that maternal neuroplasticity represents the first investment to which offspring are exposed, and hence is the primary influence of early nutrition and physical activity on neural development. In this study, two maternal environmental stimuli were tested (LP diet and physical activity). We posit that the variability in maternal investment is shaped by trade-offs that emerge during gestation. This may help explain the “placental commitment” to support the intrinsic quality of fetal development. Thus, the main goal of this study was to evaluate the effects of undernutrition and physical activity on the gene expression levels of BDNF, NTF4/NT-4, TrkB/NTRK2, IGF-1, and IGF-1r in the different areas of mother’s brain (hypothalamus, hippocampus, and motor cortex), placenta, and fetal’s brain of rats.
Material and methods
The experimental protocol was approved by the Ethical Committee of the Biological Sciences Center (protocol nº 23076.015984/2015-30), Federal University of Pernambuco, Recife, PE, Brazil, and we followed the Guidelines for the Care and Use of Laboratory Animals.
Animals and experimental diets
Twenty virgin female albino Wistar rats (Rattus norvegicus) aged 85–95 days were obtained from the Department of Nutrition, Federal University of Pernambuco, Brazil. Animals were maintained at a room temperature of 22 ± 1 °C with a controlled light–dark cycle (dark 06.00 am–6.00 pm). Food and water were given ad libitum throughout the experiment. The rats were individually housed in voluntary physical activity cages (cages equipped with a running wheel) for 4 weeks in order to monitor their spontaneous propensity to physical activity. After this period, the rats were classified as inactive (I; n = 11, animals doing < 1 km/day) and active (A; n = 9, animals doing > 1 km/day) and were placed into a standard cage and mated (1 female for 1 male) for a period of 1–5 days. Females had no access to the running wheel during mating. The day on which spermatozoa were present in a vaginal smear was designated as day 0 of gestation. Dams were transferred back to their original cages with free access to the running wheel throughout gestation. Part of the dams received a casein-based diet (AIN-93 G diet, containing 18% protein) and the other part received the same diet, but low amount of protein (8% protein). Thus, groups were formed as follows: inactive normoprotein diet (I-NP, n = 6); inactive LP diet (I-LP, n = 5); active normoprotein diet (A-NP, n = 4), and active LP diet (A-LP, n = 5). At day 20 of gestation, dams were euthanized by decapitation after a 6-h fasting period. Experimental analyses were performed in specific brain areas of mothers (hypothalamus, hippocampus, and motor cortex) and placenta. At sacrifice, one to two pups from each dam were collected for extraction of the fetus’ brain. Brains from male and female pups were immediately removed and frozen: I-NP (n = 11); I-LP (n = 8); A-NP (n = 11), and A-LP (n = 10). The tissues collected were stored at −80 °C until RNA extraction.
Voluntary physical activity measurements
Female Wistar rats were individually housed in voluntary physical activity cages (with running wheels – 27 cm diameter) for a 4-week period of adaptation. A wireless cyclocomputer (Cataye, model CC-VL820, Colorado, USA) was attached in the wheel to calculate and display trip information, such as distance traveled, duration of activity, and estimated calorie burned. These parameters were used to classify the rats according to the level of daily physical activity in: inactive (I) or active (A) according to previous studies. Reference Fragoso, Lira, Chagas, Lucena Cavalcanti, Beserra, de Santana-Muniz, Bento-Santos, Martins, Pirola, da Silva Aragao and Leandro4,Reference Santana Muniz, Beserra, da Silva Gde, Fragoso, Lira Ade, Nascimento, Manhaes de Castro and Leandro22 After mating, dams continued to have access to the running wheel during gestation.
Body weight and food intake
Mother’s body weight was recorded each 3 days throughout the experiment. Maternal food consumption was determined by the difference between the amount of food provided at the onset of the dark cycle (06.00 am) and the amount of food remaining 48 h later. Body weight of the pups and the placental weight were measured at the day of sacrifice (day 20 of gestation). Body weight was recorded using a Marte Scale (AS-1000) with 0.01 g accuracy.
Blood glucose measurements
Fasting glycemia levels were evaluated in the last day of adaptation and weekly during gestation using blood samples from the tail vein of the rats, using a glucometer (Accu Check Advantage and Accutrend GCT) and the glucose oxidase method. The animals were fasted 6 h prior to glycemia measurement.
RNA extraction
Total RNA was extracted from brain regions of mother’s brain (hypothalamus, hippocampus, and cortex), placenta, and fetus’ brain with TRI reagent® (SIGMA-ALDRICH T9424, St. Quentin Fallavier, FR) according to the manufacturer’s instructions. Briefly, 1 mL of TRI reagent® was added per 50–100 milligram of tissue, the resulting suspension was homogenized and incubated at room temperature for 5 min. Thereon, 0.2 mL of chloroform was added, samples were vortexed for 15 s, incubated for 5 min at room temperature and centrifuged at 12,000 g for 15 min at 4 °C. The upper aqueous phase was transferred to a fresh tube and 0.5 mL of isopropanol were added to precipitate RNA. Samples were incubated for 10 min at room temperature and centrifuged at 12,000 g for 15 min at 4 °C. The supernatant was removed and RNA-containing pellets were washed sequentially with 75% and 100% ethanol and dissolved in 100 μL Rnase-free water. RNA concentration and purity (defined by a 260/280 nm absorbance ratio > 1.8) was determined on a Nanodrop 2000 (Thermofisher).
Reverse transcription
Reverse transcription was performed using an PrimeScript RT reagent Kit-Perfect Real Time (TAKARA) using 0.5 μg of RNA for brain of mothers (hypothalamus, hippocampus, and cortex) and 1 μg of RNA for placenta and fetus’ brain following the manufacturer’s instructions. RNase-free H2O (3 μL), PrimeScript Buffer 5× (4 μL), Oligo dT – 50 μM (1 μL), Random hexamers – 100 μM (1 μL), and of PrimeScript RT Enzyme Mix (1 μL) were sequentially added, followed by a 15-min incubation at 37 °C and 15 s at 85 °C. Reverse transcription reactions were brought to 200 μL final volume by adding Rnase-free water and stored at −20 °C.
Quantitative PCR (qPCR)
Real-time quantitative PCR (qPCR) amplification was performed using a Rotor-Gene Real-Time PCR System (Labgene Scientific Instruments, Archamps, France). The sequences of primers used in this study are reported in Table 1. Reactions were incubated at 95 °C for 10 min, followed by 40 cycles of denaturation (95 °C, 10 s), annealing (58–65 °C depending on the primer sets, 30 s), and elongation (72 °C, 30 s). mRNA expression levels of insulin-like growth factor 1 (IGF-1), insulin-like growth factor 1 receptor (IGF-1 R), BDNF, and neurotrophic tyrosine kinase receptor type 2 (NTRK2, or TrkB) were performed. qPCR results from each gene (including the housekeeping genes) were expressed as arbitrary units derived from a standard calibration curve derived from a reference sample. qPCR for each sample was carried out in duplicate. The mRNA levels of the analysed genes were normalized using the mRNA levels of ribosomal protein L19 and beta actin.
Table 1. Primers sequence used to perform qRT-PCR

IGF-1, insulin-like growth factor 1; IGF-1r, insulin-like growth factor 1 receptor; BDNF, brain-derived neurotrophic factor; NTF4, neurothrophin 4; NTRK2, neurotrophic tyrosine kinase receptor type 2; RPL19, ribosomal protein L19; Actb, beta actin.
Statistical analyses
The Kolmogorov–Smirnov test was performed to determine the normal distribution of data. Measurements of distance traveled, estimated calories burnt, and time of activity were analyzed by two-way ANOVA, with physical activity and time as factors, followed by Bonferroni’s post hoc test. During the adaptation period, statistical analyses of body weight, food intake, and fasting glycemia were performed by Student’s t-test (active vs inactive). During gestation, statistical analyses were performed by using two-way ANOVA, with physical activity and diet as factors, followed by Bonferroni’s post hoc tests. All data are presented as means ± S.E.M. Significance was set at p < 0.05. Data analysis was performed using the statistical program GraphPad Prism 5 (GraphPad Software Inc., La Jolla, CA, USA).
Results
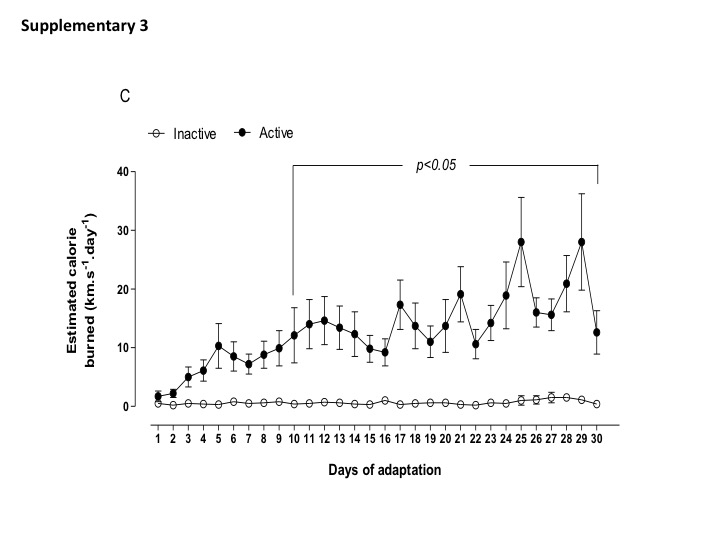
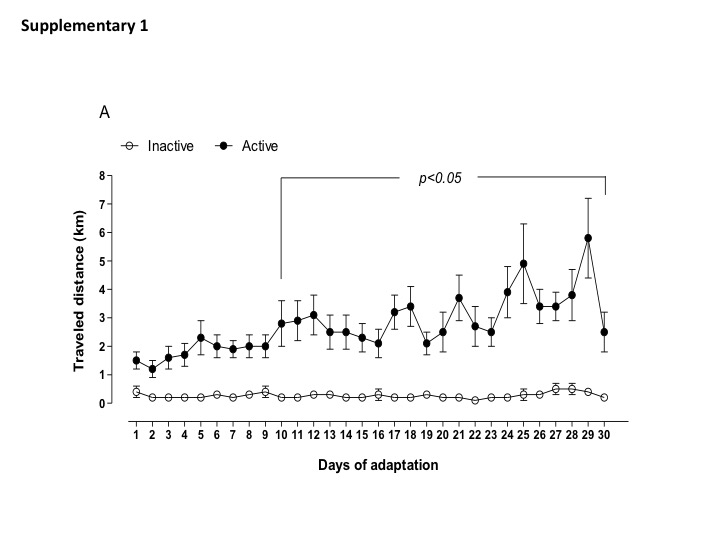
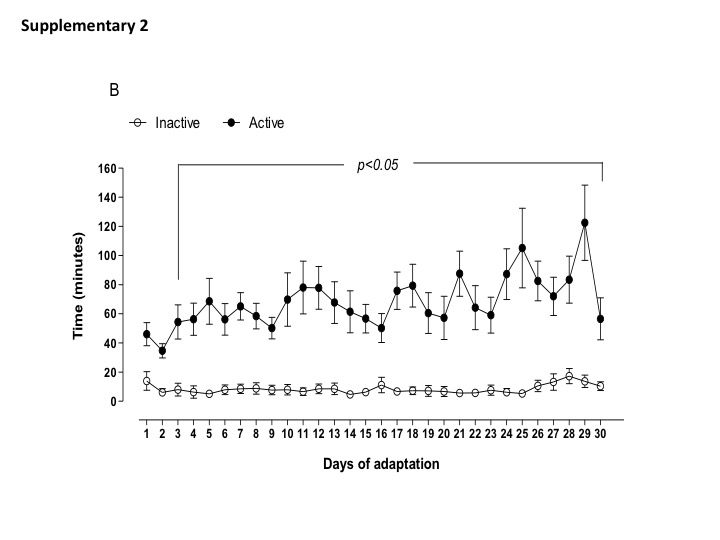
Daily physical activity (distance traveled, time spend in the wheel running, and estimated burned calories) of active and inactive dams are shown in Supplemental data (Figs. S1, S2, and S3). Of note, the most active dams spent 1 to 2 h in the wheel running, performing 4–6 km per day before breeding. Body weight, food intake, and fasting glycemia assessment were not different between active and inactive dams (Table 2). During gestation, maternal exposure to low protein diet did not change the amount of distance traveled and time in the wheel running (1–2 km per day) until the end of gestation. Inactive dams, irrespective to LP diet, continued performing less than 1 km per day (Fig. 1a–c). In addition, I-LP dams showed reduced food intake, while A-LP showed an increased food intake when compared to I-LP (Table 3).
Table 2. Maternal physiological parameters during adaptation (30 days before pregnancy). Groups: inactive (n = 11) and active (n = 9). Values expressed as mean ± S.E.M. using Student’s t-test


Fig. 1. Parameters of maternal voluntary physical activity for the experimental groups: inactive normoprotein diet (I-NP, n = 6); inactive low-protein diet (I-LP, n = 5); active normoprotein diet (A-NP, n = 4), and active low-protein diet (A-LP, n = 5). Traveled distance (A), time of activity (B), and estimated calorie burned (C) were recorded during the period of gestation. Values are presented as means.
Table 3. Maternal physiological parameters during gestation

NP = normoprotein and LP = low protein. Data were analyzed by two-way ANOVA, and Bonferroni’s post hoc test was used. *p < 0.05: A-LP vs I-LP.
We next asked whether changes in the observed maternal active phenotypes were secondary to changes in either physical activity or protein restriction on the expression of mRNA in different brain areas of mother’s brain, placenta, and fetus’ brain. In the hypothalamus, dams submitted to a LP diet during gestation showed upregulation of IGF-1r and BDNF mRNA (Fig. 2a). However, voluntary physical activity was able to attenuate the increase in IGF-1r but a higher upregulation of BDNF was seen in the hypothalamus of active dams (Fig. 2a). Likewise, in the hippocampus, maternal low protein diet-induced upregulation of IGF-1r and NTRK2 were attenuated by voluntary physical activity (Fig. 2b). In addition, BDNF was also increased in the hippocampus of active dams.

Fig. 2. mRNA expression of IGF1, IGF1r, BDNF, and NTRK2 in the hypothalamus (A), hippocampus (B), and motor cortex (C) of mothers. The groups were constituted by: inactive normoprotein diet (I-NP, n = 6), inactive low-protein diet (I-LP, n = 5), active normoprotein diet (A-NP, n = 4), and active low-protein diet (A-LP, n = 5). Values are presented as mean ± S.E.M. a P < 0.05 vs I-NP; b P < 0.05 vs I-LP. Statistical analysis was performed using two-way ANOVA with Bonferroni’s post hoc test.
Maternal protein restriction induced an upregulation of all genes that were evaluated in the motor cortex (Fig. 2c). In the placenta, we observed a downregulation of IGF-1 in inactive LP dams (Fig. 3). This effect was attenuated by maternal physical activity.

Fig. 3. mRNA expression of IGF1, IGF1r, BDNF, and NTF4 in the placenta. The groups were constituted by inactive normoprotein diet (I-NP, n = 6), inactive low-protein diet (I-LP, n = 5), active normoprotein diet (A-NP, n = 4), and active low-protein diet (A-LP, n = 5). Values are presented as mean ± S.E.M. a P < 0.05 vs I-NP. Statistical analysis was performed using two-way ANOVA with Bonferroni’s post hoc test.
In the fetus’ brain, there was a downregulation in IGF-1r in response to LP diet irrespective to maternal physical activity. BDNF mRNA was upregulated in offspring brain from active dams (Fig. 4). We observed a downregulation of NTRK2 in inactive LP dams (Fig. 4), and this effect was attenuated by maternal physical activity (Fig. 4).

Fig. 4. mRNA expression of IGF1, IGF1r, BDNF, and NTRK2 in the fetus’ brain. The groups were constituted by inactive normoprotein diet (I-NP, n = 6), inactive low-protein diet (I-LP, n = 11), active normoprotein diet (A-NP, n = 8), and active low-protein diet (A-LP, n = 10). Values are presented as mean ± S.E.M. a P < 0.05 vs I-NP; b P < 0.05 vs I-LP. Statistical analysis was performed using two-way ANOVA with Bonferroni’s post hoc test.
Discussion
The mother’s brain and placenta interact during pregnancy and mediate maternal adaptations to support eventual environmental disturbances. Reference Behura, Kelleher and Spencer24 In the context of developmental neuroplasticity, previous studies focused on the short- and long-lasting effects of nutrition on the offspring and on how the fetus adapts to diverse nutritional variations. Reference Ferreira, Liu, Fernandes and Lagranha25–Reference Thanos, Zhuo, Robison, Kim, Ananth, Choai, Grunseich, Grissom, George, Delis and Reyes28 Conversely, environmental stimuli before and during pregnancy are primarily sensed by mothers, showing that the maternal life history is highly relevant to any adaptations of the offspring. Reference Wells7 In this study, we applied maternal protein restriction during pregnancy to investigate the gene expression of neurotrophic factors. In LP mothers, IGF-1r and BDNF mRNA were upregulated in the hypothalamus. NTRK2 and IGF-1r were upregulated in the hippocampus, and all neurotrophic factors were upregulated in motor cortex of the LP mother. Changes in the neural structure, function, and molecular patterns at the end of gestation period can be expected to depend on certain brain systems that need to be plastic to specific circumstances and adaptable to environmental cues. Reference Kim, Strathearn and Swain29 Next, we found that placenta safeguards the fetal brain except for IGF-1r and NTRK2 mRNA that were downregulated, probably because of the reduced placental IGF-1 mRNA. The connectivity between the mother and the placental–fetus structure may be plastic according to the demands of different environmental stimuli. Indeed, maternal physical activity, regardless of the diet, did not affect the transcriptional response of neurotrophic factors, as supported by previous studies on phenotype plasticity. Reference Fragoso, Lira, Chagas, Lucena Cavalcanti, Beserra, de Santana-Muniz, Bento-Santos, Martins, Pirola, da Silva Aragao and Leandro4,Reference Santana Muniz, Beserra, da Silva Gde, Fragoso, Lira Ade, Nascimento, Manhaes de Castro and Leandro22,Reference Senna, Torres, Lopes, Alheiros-Lira, de Moura, Pereira, de Aguiar, Ferraz and Leandro30 However, in the present study, some of undernutrition-induced changes on the transcription levels of neurotrophic factors were permanent, and an active maternal phenotype was not able to revert the effects on motor cortex in mother’s brain. The findings provide new evidence for and insights into how the mother’s brain, placenta, and fetus’ brain interact during pregnancy in response to different environmental cues.
During gestation, mothers remained active although with a reduction in the amount of distance traveled (1–2 km/day) and time spent (30–40 min/day) in the running wheel, irrespective to the low-protein diet. This result is aligned with previous studies using the same experimental model. Reference Fragoso, Lira, Chagas, Lucena Cavalcanti, Beserra, de Santana-Muniz, Bento-Santos, Martins, Pirola, da Silva Aragao and Leandro4,Reference Santana Muniz, Beserra, da Silva Gde, Fragoso, Lira Ade, Nascimento, Manhaes de Castro and Leandro22 In addition, food intake, weight of placenta, and the body weight of the fetuses were similar among groups. Paradoxically, in energetic terms, the maternal dietary intake during pregnancy has relatively modest effects on fetal energy requirements even after addressing placental costs. Reference Wells7,Reference Tarrade, Panchenko, Junien and Gabory10,Reference Butte and King31 It is interesting to observe that active mothers performed around 1–2 h on the wheel running (4–6 km per day) before breeding. Active mothers probably allocated relatively more energy to gestation by increasing basal oxygen consumption and fat-free mass. Reference Fidalgo, Falcao-Tebas, Bento-Santos, de Oliveira, Nogueira-Neto, de Moura, Lisboa, de Castro and Leandro8 We speculate that dams with high levels of daily physical activity may increase the protein anabolism and promote nutritional investment to their fetus during pregnancy even when submitted to a LP diet. Reference Fidalgo, Falcao-Tebas, Bento-Santos, de Oliveira, Nogueira-Neto, de Moura, Lisboa, de Castro and Leandro8,Reference Leandro, Fidalgo, Bento-Santos, Falcao-Tebas, Vasconcelos, Manhaes-de-Castro, Carpinelli, Hirabara and Curi17 Thus, it is plausible to consider maternal voluntary physical activity as a “proactive prediction” by the mother in order to buffer an eventual injured growth trajectory of fetus due to environmental insults.
IGF-1r mRNA was upregulated in hippocampus, hypothalamus, and motor cortex of mother’s brain during gestation. Our data converge with previous studies. Reference Kim, Strathearn and Swain29,Reference Patz and Wahle32,Reference Coupe, Dutriez-Casteloot, Breton, Lefevre, Mairesse, Dickes-Coopman, Silhol, Tapia-Arancibia, Lesage and Vieau33 IGF-1r has a potent effect on cellular neuroplasticity in neuroepithelial cell types and shows a relatively stable pattern of expression from early development to maturity. Reference Dyer, Vahdatpour, Sanfeliu and Tropea34 The increased IGF-1r expression highlights the association between regions of increased neurogenesis and the active transport mechanism that allows peripheral circulating IGF-1 to cross the blood–brain barrier and induce the mother’s brain to be responsive to undernutrition. Physical activity mitigated the upregulation of IGF-1r except for the motor cortex. Peripheral IGF-1 concentration is quickly increased in humans in response to physical exercise, and adaptive responses of its receptor were seen in the brain. Reference Dyer, Vahdatpour, Sanfeliu and Tropea34,Reference Wrigley, Arafa and Tropea35 In turn, normalization of brain IGF-1r mRNA may be caused not only by increased circulating levels of IGF-1 but also by the increased brain–blood flow induced by exercise or IGF-1 itself. Reference Wrigley, Arafa and Tropea35
The placenta plays a foremost role in the development of the fetal brain and also influences maternal brain function, by a mechanism that may include neurotrophic and growth factors. Reference Behura, Kelleher and Spencer24,Reference Dyer, Vahdatpour, Sanfeliu and Tropea34 Our data showed that IGF-1r mRNA was downregulated in the placenta of LP mothers, but maternal physical activity was able to attenuate this reduction. This study focused on the evaluation of hormones gene expression in the placenta, and future histological analysis will allow determining whether IGF-1 protein expression is likewise modulated by LP diet and physical activity. A previous study showed that maternal exercise (treadmill, 20 m/min for 20 min/day, once per day for 19 days) significantly increased plasma IGF-1 concentration in the late period of pregnancy. Reference Turgut, Kaptanoglu, Emmungil and Turgut36 In fetus’ brain, there was a reduced IGF-1r gene expression in response to maternal LP diet and physical activity, which may be sex-independent as both female and male pups were analyzed. The pattern of changes in placental and fetal IGF-1 mRNA mirrors weight gain and neural development. Reference Kim, Strathearn and Swain29 Indeed, our recent data showed that physical features, reflex ontogeny, and somatic growth were affected in pups from protein-restricted mothers, and physical activity acted as a buffer for these effects. Reference Fragoso, Lira, Chagas, Lucena Cavalcanti, Beserra, de Santana-Muniz, Bento-Santos, Martins, Pirola, da Silva Aragao and Leandro4 The present study confirms the importance of both IGF-1 and IGF-1r as genes mediating the interaction between adaptive response to malnutrition and physical activity during embryonic development and placental growth.
In mother’s brain, BDNF mRNA was upregulated in response to protein restriction and physical activity in both hypothalamus and hippocampus. In adult, a close relationship between the nutritional status and BDNF level in different areas of the brain has been proposed to control neuronal proliferation, neurogenesis, axonal and dendritic plasticity, synapse formation, stimulation of neurotransmitter or neuropeptide synthesis and release. In addition, brain metabolic changes evoked by exercise strongly depend on the levels of BDNF and NTRK2 mRNA. Reference Dyer, Vahdatpour, Sanfeliu and Tropea34 BDNF is one of the neurotrophic factors supporting differentiation, maturation, and survival of neurons and showing a neuroprotective effect. Reference Garces, Sanchez, Torres-Sierra, Ruiz-Parra, Angel-Muller, Alzate, Sanchez, Gomez, Romero, Castaneda, Sanchez-Rebordelo, Dieguez, Nogueiras and Caminos13 In the present study, mothers submitted to a LP diet showed upregulation of BDNF in the hypothalamus and motor cortex, and their fetus were not growth-restricted in response to the LP diet. This neurothrophic factor plays a major role in energy homeostasis under adverse conditions such as the maternal undernutrition applied in this study and can protect the growth of fetus. Reference Mayeur, Silhol, Moitrot, Barbaux, Breton, Gabory, Vaiman, Dutriez-Casteloot, Fajardy, Vambergue, Tapia-Arancibia, Bastide, Storme, Junien, Vieau and Lesage14 Collectively, these observations indicate that maternal physical activity initiated in early pregnancy induces maternal–fetal adaptations that can be considered as an investment countering the effects of undernutrition. The underlying mechanism can be related to epigenetic modulation induced by physical activity that regulates gene expression. Reference Gomez-Pinilla, Zhuang, Feng, Ying and Fan38 We found that maternal physical exercise induces upregulation of BDNF and its receptor NTRK2. In the case of these changes occurring during the critical period of fetal development, nutrition and physical activity assume important roles in the control of gene transcription in the context of the long-term effects of developmental plasticity.
In accordance with previous observations, Reference Coupe, Dutriez-Casteloot, Breton, Lefevre, Mairesse, Dickes-Coopman, Silhol, Tapia-Arancibia, Lesage and Vieau33,Reference Lee, Duan and Mattson39,Reference Lee, Seroogy and Mattson40 our findings show that, in developing rats, BDNF mRNA levels are significantly augmented in the fetus’ brain during sensitive developmental windows. Both maternal undernutrition and physical activity enhanced BDNF mRNA in the fetus’ brain. The environmental stimuli acting in the critical period of CNS development may elicit molecular signals to protect brain development. Thus, BDNF could be implicated in the metabolic adjustment during food-restricted conditions and energetic expenditure that may also participate in the protection of CNS against excitotoxicity damage under these conditions. Reference Coupe, Dutriez-Casteloot, Breton, Lefevre, Mairesse, Dickes-Coopman, Silhol, Tapia-Arancibia, Lesage and Vieau33 BDNF modulates survival, differentiation, and activity of neurons by binding to its high-affinity receptor, NTRK2, which is responsible for the initiation of intracellular signaling cascades and the regulation of the local availability and responsiveness to BDNF. Reference De Assis, Gasanov, de Sousa, Kozacz and Murawska-Cialowicz37 We showed that the gene expression of NTRK2 is downregulated in the fetus’ brain. Maternal LP diet may especially have affected the neurotrophic BDNF/NTRK2 pathway during the perinatal life. However, pups from active LP mother did not change mRNA NTRK2. Thus, nutrition and physical activity can be considered important investments during pregnancy, and fetal life is a critical period for the structural and functional development of brain. Collectively, these traits underpin the maternal and placental capacity for safeguarding the magnitude of prenatal growth as a valuable marker of the intrinsic quality of the neuroplasticity.
Conclusion
In conclusion, maternal protein restriction and physical activity can influence the gene expression of BDNF, NTRK2, IGF-1, and IGF-1r in different areas of mother’s brain. There is an important interaction between mother and placental development in the control of fetal growth factors gene expression. In the present study, we demonstrated maternal neuroplasticity representing the first investment for offspring and the primary influence of early nutrition and physical activity on neural development. Additionally, IGF-1 gene expression alterations observed in the placenta suggest that this organ may also contribute to the maintenance of fetal neurodevelopment. Our investigation, performed on a tractable animal model, provides some valuable insights on the relevance of the hormonal control defects in pregnant women. Indeed, recent studies demonstrated that placentas from mothers with small from gestational age pregnancies have lower placental IGF-1 signaling Reference Lazo-de-la-Vega-Monroy, Mata-Tapia and Garcia-Santillan41 and in pre-eclampsia cases the expression of IGF-1 is lowered, Reference Dubova, Pavlov, Lyapin, Kulikova, Shchyogolev and Sukhikh42 via a hypermethylation mechanisms on the IGF-1 promoter control, Reference Ma, Zhou, Xiong, Li and Li43 which underscores the importance of epigenetic control mediating the gene–environment interactions during pregnancy.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S2040174420000756
Acknowledgments
This study was supported by the National Council for Scientific and Technological Development (CNPq, 302515/2015-1), Coordination for the Improvement of Higher Level (or Education) Personnel (CAPES-COFECUB Me797-14), and State of Pernambuco Science and Technology Support Foundation (FACEPE, APQ: 0797-4.05/14 and APQ-0201-4.05/14).
The authors’ contributions are as follows: J.F., J.H.C.S., L.P., and C. G. L. were the principal investigators, contributed to the study design, interpretation of the findings, and data analysis and wrote the manuscript; G. C. J.-S., H.T.S., E.L., and V.O.N.S. contributed to the preparation of diets and tissues and data analysis. R.C.A.G., R.S.A., and H.V. contributed to the data analysis and reviewed the manuscript. All authors have read and approved the final version of the manuscript.
The authors declare that there are no conflicts of interest.