


Weaning is a critical period in swine production. In physiological conditions, piglets would be weaned gradually from week 11 to week 20–22, approximately (Newberry & Wood-Gush, Reference Newberry and Wood-Gush1986). This natural situation contrasts with the sudden weaning that piglets experience at week 3–4 in intensive systems. Probably, this is one of the leading causes responsible for the mortality and morbidity rates observed during this production stage since sow milk not only fulfils all the nutritional requirements of the rapidly-growing piglets, but also protects them against infectious diseases (Nagy et al. Reference Nagy, Mackenzie and Bharucha1976).
Similarly to other mammal species, microbiological studies focused on sow milk have been restricted to the identification of potential pathogenic microorganisms. In contrast, human breast milk has been recently shown to be an excellent and continuous source of probiotic lactic acid bacteria to the infant gut (Heikkilä & Saris, Reference Heikkilä and Saris2003; Martín et al. Reference Martín, Langa, Reviriego, Jiménez, Marín, Xaus, Fernández and Rodríguez2003, Reference Martín, Heilig, Zoetendal and Rodríguez2007), which may play an important role in the reduction of the incidence and severity of infections in the suckling infant. Interestingly, some of the lactic acid bacteria strains isolated from this biological fluid have the ability to inhibit the growth of a wide spectrum of pathogenic bacteria by competitive exclusion and/or through the production of antimicrobial compounds, such as bacteriocins, organic acids or hydrogen peroxide (Beasley & Saris, Reference Beasley and Saris2004; Martín et al. Reference Martín, Olivares, Marín, Fernández, Xaus and Rodríguez2005a).
More recently, it has been shown that such bacteria are transferred from the maternal gut to the mammary gland during late pregnancy and lactation through an immunologically-based internal route that involves dendritic cells and macrophages: the enteromammary pathway (Martín et al. Reference Martín, Langa, Reviriego, Jiménez, Marín, Olivares, Boza, Jimenez, Fernandez, Xaus and Rodriguez2004; Perez et al. Reference Perez, Doré, Leclerc, Levenez, Benyacoub, Serrant, Segura-Roggero, Schiffrin and Donnet-Hughes2007). Since many Lactobacillus (Lb.) species are common inhabitants of both the human and the porcine gut, the objective of this work was to elucidate if sow milk may be a source of lactic acid bacteria for the piglet gut and, in such case, to evaluate the probiotic potential of the isolates.
Materials and Methods
Isolation and enumeration of lactobacilli from sow milk
The 8 large white sows from which the biological samples were obtained fulfilled the following criteria: (a) healthy sows without present or past underlying conditions; (b) normal pregnancy; (c) no antibiotics were used in the feed or given as treatment to the sows during gestation/lactation, and (d) absence of piglet and/or sow perinatal problems. The protocol was approved by the Ethical Committee on Animal Experimentation of Universidad Complutense de Madrid (Spain). Milk samples were collected in sterile tubes by manual expression using sterile gloves. Previously, nipples and surrounding skin were cleaned with soap and sterile water, and soaked in chlorhexidine (Cristalmina, Salvat, Barcelona, Spain). The first drops (~1 ml) were discarded. Skin sampling (~4 cm2) was performed as described previously (Paulino et al. Reference Paulino, Tseng, Strober and Blaser2006). The milk and skin samples were obtained at day 7 after delivery and kept at 4°C until delivery to the laboratory, which happened within the first 3 h after collection.
Peptone water dilutions of the milk and skin samples were plated in triplicate onto de Man, Rogosa, and Sharpe (MRS, Oxoid, Basingstoke, UK) supplemented with L-cysteine (0·5 g/l) (MRS-Cys) agar plates, which were incubated anaerobically (85% nitrogen, 10% hydrogen, 5% carbon dioxide) in an anaerobic workstation (MINI-MACS, DW Scientific, Shipley, UK) at 37°C for 48 h. Parallel, and to evaluate faecal contamination, the samples were also cultured on Violet Red Bile Agar (VRBA; Difco, Detroit, MI) agar plates, which were aerobically incubated at 37°C for 24 h. In both growth media, the lower limit of detection was estimated to be approximately 50 CFU (colony-forming units)/ml.
Identification of lactobacilli from sow milk
A total of 104 colonies were selected from the MRS-Cys plates for further screening. The isolates were examined by phase-contrast microscopy to determine cell morphology and Gram-staining reaction, and tested for oxidase and catalase activities and, also, for good growth (>108 CFU/ml) in MRS broth incubated at 37°C for up to 24 h. Oxidase- and catalase-negative Gram-positive bacilli were genotyped by randomly amplified polymorphic DNA (RAPD) analysis. Genomic DNA was isolated from 10 ml of overnight MRS cultures using the DNeasy tissue kit (Qiagen, Hilden, Germany). The DNA was used in PCR amplifications that were performed using primers ArgDei (5′-ACCYTRGAAGGYGGYGATGTB-3′) and OPL5 (5′-ACGCAGGCAC-3′) (Veyrat et al. Reference Veyrat, Miralles and Pérez-Martínez1999) as described by Ruiz-Barba et al. (Reference Ruiz-Barba, Maldonado and Jiménez-Díaz2005).
One representative of each RAPD profile was identified by PCR amplification of a section of a 16S rRNA gene variable region using primers pbl16 (5′-AGAGTTTGATCCTGGCTCAG-3′) and mlb16 (5′-GGCTGCTGGCACGTAGTTAG-3′) (Kullen et al. Reference Kullen, Sanozky-Dawes, Crowell and Klaenhammer2000). PCR conditions were as follows: 96°C for 30 s, 50°C for 30 s and 72°C for 45 s (35 cycles) and a final extension at 72°C for 4 min. Amplified fragments were purified using the NucleoSpin Extract II (Macherey-Nagel Gmb; Düren, Germany) and sequenced using the primers cited above on an ABI 377A automated sequencer (Applied Biosystems, Foster City, USA). The sequences were compared with those deposited in the EMBL database using BLAST algorithm (http://www.ncbi.nlm.nih.gov/BLAST).
Final identification of the isolates that, on the basis of 16S rRNA sequencing, had been classified as belonging to the Lb. plantarum-Lb. paraplantarum-Lb. pentosus group, was performed by a multiplex species-specific PCR assay (Torriani et al. Reference Torriani, Felis and Dellaglio2001).
Determination of antimicrobial spectrum
An overlay method previously described (Magnusson & Schnürer, Reference Magnusson and Schnürer2001; Martín et al. Reference Martín, Olivares, Marín, Xaus, Fernández and Rodríguez2005b) was used to determine the ability of the lactobacilli strains to inhibit the growth of different bacteria, moulds and yeasts. The following bacteria were employed as indicator organisms: Enterococcus faecium P21, Ent. faecalis TAB28, Listeria monocytogenes ScottA, List. monocytogenes Ohio, List. innocua RdC, Staphylococcus aureus CECT 5191, Staph. epidermidis CECT 231, Salmonella cholerasuis CECT 4155, Sal. cholerasuis CECT 409, Sal. cholerasuis CECT 443, Sal. enteritidis 4396, Escherichia coli CECT 4076 (O157:H7), Esch. coli RJM1, Esch. coli RJM2, Klebsiella pneumoniae CECT 142, Kleb. oxytoca CECT 860T and Proteus vulgaris CECT 484. Aspergillus fumigatus CECT 2071, Kluyveromyces marxianus CECT 10357, and Debaryomyces hansenii CECT 10360 were provided by the Spanish Collection of Type Cultures (CECT, Burjassot, Spain), while the mould Penicillium roquefortii PB1 was provided by Puleva Biotech (Granada, Spain). The plates overlaid with bacterial indicators were incubated at 37°C for 48 h, while those overlaid with yeasts cells or fungal spores were incubated at 30°C for up to 120 h. The plates were examined for zones of inhibition around the lactobacilli streaks. All experiments assaying inhibitory activity were performed in triplicate.
Production of bacteriocins
Lactobacilli were grown in MRS broth at 37°C until early stationary phase (A620 ~1·0). Preparation of cell-free supernatants and analysis of their bacteriocinogenic activity were performed as described (Martín et al. Reference Martín, Olivares, Marín, Xaus, Fernández and Rodríguez2005b). The microorganisms employed as indicators of bacteriocinogenic activity were the Gram-positive bacteria previously used for determination of the antimicrobial spectrum: Ent. faecium P21, Ent. faecalis TAB28, List. monocytogenes ScottA, List. monocytogenes Ohio, List. seeligeri RdC, Staph. aureus CECT 5191 and Staph. epidermidis CECT 231.
Since Lb. salivarius CELA2 showed bacteriocinogenic activity against some of the indicators, PCR assays were performed to detect structural genes corresponding to known bacteriocins produced by this bacterial species. The primer pairs SalB-for (5′-TGATAAGAAAGAATTGGCACATATAATTG-3′) and SalB-rev (5′-TCTGTTTAACTACAAATATTTTGATTTGAATG-3′), and Abp118A-for (5′-AAACGTGGTCCTAACTGTGTAGG-3′) and Abp118B-rev (5′-AACGGCAACTTGTAAAACCACCAG-3′), were designed on the basis of the structural genes encoding salivaricin B (Çataloluk, Reference Çataloluk2001) and bacteriocin Abp-118 (Flynn et al. Reference Flynn, van Sinderen, Thornton, Holo, Nes and Collins2002), respectively. DNA was amplified in 25-μl reaction mixtures containing 2·5 mm-MgCl2, 1×reaction buffer, 0·2 μm of each of the deoxynucleotides triphosphates (dNTPs), 1 μm of each primer, and 1·25 U Taq DNA polymerase (Ecotaq; Ecogen, Barcelona, Spain). Amplification included denaturation at 94°C for 4 min, followed by 30 cycles of denaturation at 94°C for 30 s, annealing at 58°C for 30 s, polymerization at 72°C for 30 s, and a final polymerization step at 72°C for 5 min.
Production of reuterin
Production of reuterin by Lb. reuteri CR20 was investigated since it showed an increased antimicrobial activity when glycerol (which is required for reuterin biosynthesis) was present in the medium. For this purpose, cell-free supernatants were prepared as described by Martín et al. (Reference Martín, Olivares, Marín, Xaus, Fernández and Rodríguez2005b) and the presence of reuterin in the supernatants was detected by the colorimetric method of Smiley & Sobolov (Reference Smiley and Sobolov1962). Lb. reuteri CECT 925 and Lb. coryniformis CECT 5711 (Martín et al. Reference Martín, Olivares, Marín, Xaus, Fernández and Rodríguez2005b) were used as positive controls. In parallel, the inhibitory activity of purified reuterin extracts was determined and quantified with the MIC microtiter assay of Talarico et al. (Reference Talarico, Casas, Chung and Dobrogosz1988), using Esch. coli K12 CECT 433 as the indicator organism. One unit of reuterin was defined as the reciprocal of the highest dilution that did not allow the growth of the Esch. coli indicator strain. Reuterin previously purified from Lb. reuteri CECT 925 cultures was used as standard. All these assays were performed in triplicate.
PCR was used to detect a 279 bp DNA fragment belonging to the gene encoding the 60-kDa glycerol dehydratase subunit, which is essential for reuterin production, using the primers (GD1 and GD2) and PCR conditions described by Claisse & Lonvaud-Funel (Reference Claisse and Lonvaud-Funel2001).
Survival after transit through an in vitro gastrointestinal model
The survival of the strains was tested in an in vitro model of the porcine stomach and small intestine based on that described by Marteau et al. (Reference Marteau, Minekus, Havenaar and Huis In't Veld1997). UHT-treated porcine milk (25 ml) containing approximately 109 CFU/ml of the strain tested was diluted in 5 ml of a sterile electrolyte solution containing 6·2 g NaCl, 2·2 g KCl, 0·22 g CaCl2, and 1·2 g NaHCO3/l to simulate the in vivo dilution by saliva. Then, 5 ml porcine gastric juice was added and the mixture was incubated at 37°C with agitation. The pH curve in the stomach-resembling compartment was controlled to reproduce the values found in monogastrics after yogurt consumption (Conway et al. Reference Conway, Gorbach and Goldin1987): pH 5·0 at initiation, pH 4·1 at 20 min, pH 3·0 at 40 min, and pH 2·1 at 60 min. Fractions were successively taken from this compartment at 20, 40, 60, and 80 min, in a manner that simulates the normal gastric emptying (Marteau et al. Reference Marteau, Minekus, Havenaar and Huis In't Veld1997). After adjusting their pH to 6·5±0·2 with 1 m-NaHCO3, they were mixed with 10 ml of a sterile electrolyte solution containing 5 g NaCl, 0·6 g KCl and 0·3 g CaCl2/l, 4% porcine bile, and 7% pancreatin (Sigma), which simulates the content of the duodenal juice. After 120 min of successive exposure to these conditions, bacterial survival was determined by plating the samples onto MRS agar plates, which were anaerobically incubated at 37°C for 48 h. All these assays were performed in quadruplicate and the values were expressed as the mean±sd.
Adherence to and/or degradation of porcine mucin
The adhesion of bacteria was determined according to the method described by Cohen & Laux (Reference Cohen and Laux1995) with some modifications. Briefly, 100 μl of a solution (1 mg/ml) of porcine mucin (Sigma) in HEPES-buffered Hanks salt solution (HH) were immobilized in polystyrene microtiter plates (Maxisorp; Nunc, Roskilde, Denmark) after overnight incubation at 4°C. The wells were washed twice with 250 μl HH. In parallel, bacteria were grown overnight at 37°C in MRS broth and the bacterial pellets from 1 ml fractions were obtained by centrifugation and washed with HH. Then, 10 μl 10 mm-carboxyfluorescein (Sigma) were added to the pellets and the bacterial suspensions were incubated for 20 min at 37°C. Subsequently, the bacterial cells were washed 3 times with HH and, finally, resuspended in 1 ml HH. Then, a suspension of 50 μl of the fluorescent-labelled bacteria (~5×107 CFU) was added to each well. After incubation for 1 h at 37°C, the plates were washed twice with 250 μl HH to remove unattached cells, and incubated for 1 h at 60°C in the presence of 50 μl 1% SDS-0·1 m-NaOH to release and lyse bound microorganisms. Fluorescence was measured in a fluorescence microplate reader (Tecan Austria GMBH, Salzburg, Austria). Adhesion was assessed as the percentage of the fluorescence retained in the wells after the washing steps when compared with that present in the labelled bacterial aliquots originally added to the wells. The assays were performed in duplicate.
The potential of the lactobacilli strains to degrade hog gastric mucine (HGM; Sigma) in vitro was evaluated in duplicate following the procedure developed by Zhou et al. (Reference Zhou, Gopal and Hill2001).
Formation of biogenic amines
The ability to form biogenic amines (tyramine, histamine, putrescine and cadaverine) was assessed using the decarboxylase broth and the method described by Bover-Cid & Holzapfel (Reference Bover-Cid and Holzapfel1999). The precursor amino acids (tyrosine, histidine, ornithine and lysine, respectively) were purchased from Sigma.
Sensitivity to antibiotics
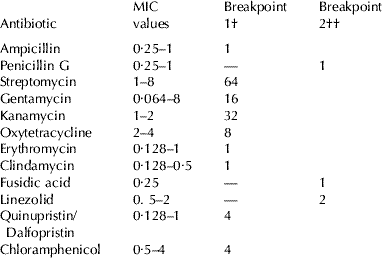
Minimum inhibitory concentrations (MICs) of 12 antimicrobial agents were determined by microdilution using the newly developed and standardized LAB susceptibility test medium (LSM) broth formulation essentially consisting of a mixture of Iso-Sensitest (IST; Oxoid) broth (90%) and MRS broth (10%) adjusted to pH 6·7 as previously described (Klare et al. Reference Klare, Konstabel, Müller-Bertling, Reissbrodt, Huys, Vancanneyt, Swings, Goossens and Witte2005). Ten of the antimicrobials tested (ampicillin, gentamicin, streptomycin, quinupristin/dalfopristin, erythromycin, clindamycin, oxytetracycline, chloramphenicol, kanamycin) were those for which the panel on additives and products or substances used in animal feed (FEEDAP) of the European Food Safety Authority (EFSA) has established microbiological breakpoints (cut-off values) that enable the distinction between lactobacilli strains harbouring acquired antimicrobial resistances and susceptible strains (EFSA, 2008). MICs of three additional antimicrobials agents for which tentative cut-off values for lactobacilli have been suggested (Klare et al. Reference Klare, Konstabel, Werner, Huys, Vankerckhoven, Kahlmeter, Hildebrandt, Müller-Bertling, Witte and Goossens2007) were also determined. The antimicrobials were tested in the concentration ranges (mg/l) given in parentheses: penicillin G (0·032-64), ampicillin (0·032-64), gentamicin (1-2048), streptomycin (2-4096), quinupristin/dalfopristin (tested as 30:70 ratio: 0·032-64), erythromycin (0·016-32), clindamycin (0·032-32), oxytetracycline (0·063-128), fusidic acid (0·063-128), linezolid (0·016-32), chloramphenicol (0·125-256), kanamycin (1-256).
Potential sow-to-piglet transfer of lactobacilli
Presence of the lactobacilli strains isolated from milk of sows 5, 7, and 8 was investigated in faecal swabs collected from the same sows and from two of their respective piglets. Among the isolates obtained on MRS-Cys plates from each sample, a total of 15 randomly selected oxidase- and catalase-negative Gram-positive bacilli were identified as described above. When isolates belonging to the same Lactobacillus species were present in the milk and faecal samples of a sow-piglets pair, they were submitted to pulsed-field gel electrophoresis (PFGE) genotyping following the method described by Jiménez et al. (Reference Jiménez, Fernández, Maldonado, Martín, Olivares, Xaus and Rodríguez2008).
Results and Discussion
In this study, presence of lactic acid bacteria in sow milk was investigated. Colonies were obtained from all the milk samples in MRS-Cys plates. The MRS-Cys counts ranged from 1·55×102 to 1·16×104 CFU/ml (Table 1). Such bacterial concentration values are similar to that reported from hygienically-obtained human milk (Heikkila & Saris, Reference Heikkilä and Saris2003; Martín et al. Reference Martín, Langa, Reviriego, Jiménez, Marín, Xaus, Fernández and Rodríguez2003; Perez et al. Reference Perez, Doré, Leclerc, Levenez, Benyacoub, Serrant, Segura-Roggero, Schiffrin and Donnet-Hughes2007). No growth was detected on VRBA plates inoculated with 6 samples while the counts in the remaining two samples were low: 5·70×101 (sow 1) and 3·30× 102 CFU/ml (sow 4) (Table 1), which confirmed the hygienic collection of the milk samples. Samples 1 and 4 were excluded from this study while a total of 104 colonies (17–18 per sample) were selected from MRS-Cys plates of the remaining milk samples. Among them, only 23 isolates were Gram-positive catalase- and oxidase-negative rods while the rest of the isolates were identified as enterococci, streptococci or staphylococci (data not shown). Subsequently, the 23 rod-shaped isolates were inoculated in MRS broth tubes, which were incubated aerobically to exclude those with fastidious incubation requirements (<107 CFU/ml after 24 h at 37°C). As a result, four additional isolates were discarded and the remaining 19 rod-shaped isolates were selected for identification and assessment of their probiotic potential. In contrast to milk samples, only enterobacteria (VRBA plates) or enterococci, streptococci or staphylococci (MRS-Cys plates) could be isolated from the skin samples (data not shown).
Table 1. Enumeration of the bacteria grown on MRS-Cys and VRBA plates, RAPD clustering of the rod-shaped isolates obtained from MRS-Cys plates and taxonomical identification

a The isolate underlined (one per each genotype) was selected for further characterization. Nd, not detected
RAPD profiling revealed the existence of 8 different genetic profiles among the rod-shaped isolates obtained from milk (Table 1). One representative of each profile (from now, “strain”) was selected for identification and characterization. Identification by 16S rDNA PCR sequencing revealed that all the strains belonged to the Genus Lactobacillus with the exception of strain CLC22, which was identified as Weissella paramesenteroides (Table 1). The results showed that this biological fluid contains lactobacilli and, therefore, is a natural source of such microorganisms for the piglet. Similarly, human milk constitutes a good source of maternal lactic acid bacteria for the infant gut (Heikkilä & Saris, Reference Heikkilä and Saris2003; Martín et al. Reference Martín, Langa, Reviriego, Jiménez, Marín, Xaus, Fernández and Rodríguez2003). Five species belonging to the genus Lactobacillus (Lb. reuteri, Lb. salivarius, Lb. plantarum, Lb. paraplantarum, Lb. brevis) were identified. Lactobacilli belonging to the same or closely related species have been frequently isolated or detected in porcine faeces and, particularly, in lactating piglets (Leser et al. Reference Leser, Amenuvor, Jensen, Lindecrona, Boye and Møller2002; Konstantinov et al. Reference Konstantinov, Awati, Williams, Miller, Jones, Stokes, Akkermans, Smidt and de Vos2006; De Angelis et al. Reference De Angelis, Siragusa, Caputo, Ragni, Burzigotti and Gobbetti2007). The lactobacilli pattern of sow milk was host-specific, a finding that has been reported for human milk (Martín et al. Reference Martín, Heilig, Zoetendal and Rodríguez2007) and piglet faeces (Simpson et al. Reference Simpson, McCracken, Gaskins and Mackie2000).
All the strains showed a clear inhibitory antimicrobial activity against all indicator organisms used in this study, with the exception of the moulds Asp. fumigatus CECT 2071 (no inhibition zone) and Pen. roquefortii PB1 (inhibition zone <2 mm). K. marxianus CECT 10357 and Deb. hansenii CECT 10360 were only inhibited by Lb. reuteri CR20, provided that glycerol was present in the MRS agar plates. Lb. paraplantarum CLB7, Lb. salivarius CELA2 and Lb. reuteri CR20 were the strains that displayed the strongest antimicrobial activity, which was particularly effective against the Listeria, Salmonella and Esch. coli strains used as indicator organisms (Table 2). In contrast, Lb. brevis CLC23 and Lb. plantarum CLC17 showed the narrower inhibition zones. Subsequently, the strains were screened for production of bacteriocins and/or reuterin. Only Lb. salivarius CELA2 showed bacteriocinogenic activity against some of the indicator bacteria (Table 2) although the strain did not harbour the structural genes encoding salivaricins B, OR-7 and Abp118. Therefore, most probably, it produces a novel bacteriocin, which will be characterized in a future study. Lb. reuteri CR20 produced reuterin and its inhibitory activity (~105 units/ml) was similar to that of Lb. reuteri CECT 925 (~100 units/ml). The gene encoding a subunit of glycerol dehydratase, an enzyme required for reuterin production, was detected in Lb. reuteri CR20.
Table 2. Antimicrobial activity of the lactobacilli isolates against several indicador organismsFootnote a

a +; halo around the streak <3 mm; ++: halo 3–6 mm; +++: >6 mm). bB: indicator organism inhibited by the bacteriocin produced by L. salivarius CELA2. cThe size of the halo increased when glycerol was added to the medium
The viability of the strains after exposition to conditions simulating those found in the gastrointestinal tract varied from 60–64% (Lb. reuteri CR20, Lb. paraplantarum CLB7, Lb. salivarius CELA2) to 38–40% (Lb. plantarum CLC17 and Lb. brevis CLC23) (Table 3). The lactobacilli strains tested showed a variable ability to adhere to porcine mucin (Table 4). Lb. reuteri CR20 was the strain that showed the highest adherence ability followed by Lb. salivarius CELA2. None of the strains were able to degrade gastric mucin in vitro. Production of biogenic amines by the lactobacilli tested could not be detected with the exception of Lb. brevis CLC23, which produced histamine and tyramine. According to the values recommended by European Food Safety Authority (EFSA, 2008), the lactobacilli were sensitive to all the antibiotics tested in this study (Table 5).
Table 3. Percentage (%) of the lactobacilli inoculated (109 CFU/ml) in sow milk that survived to conditions simulating those of the porcine gastrointestinal tract

a The different fractions were taken from the gastric-like compartment at 20, 40, 60, and 80 min and later submitted to the intestinal-like secretion, which did not affect the viability of the strains tested
Table 4. Ability of the lactobacilli to adhere to porcine mucin

a Values expressed as the percentage of the fluorescence retained in the wells after the washing steps of the assay (see Material and methods for details)
Table 5. MICs and breakpoint values (μg/mL) of the antimicrobial agents tested against the lactobacilli strains
PFGE typing of Lactobacillus isolates revealed the presence of Lb. paraplantarum CLB7 in the faecal swabs obtained from sow 5 (7 out of 15 colonies) and its two piglets (11 and 12 out of 15 colonies, respectively). Similarly, Lb. salivarius CELA 2 was isolated from the faecal swabs obtained from sow 7 (9 out of 15 colonies) and its two piglets (9 and 12 out of 15 colonies, respectively), and Lb. reuteri CR20 (11 out of 15 colonies) from the faecal samples collected from sow 8 and its piglets (9 and 11 out of 15 colonies, respectively) (Fig. 1). These results suggest that milk can be a source of lactobacilli for the piglet gut. In fact, the lactobacilli community in the piglet gut undergoes dramatic changes, including a severe decrease in its concentration, as a consequence of weaning (Konstantinov et al. Reference Konstantinov, Awati, Williams, Miller, Jones, Stokes, Akkermans, Smidt and de Vos2006; Su et al. Reference Su, Yao, Perez-Gutierrez, Smidt and Zhu2008). The possible existence of other species or strains in the milk samples cannot be excluded since they may have fastidious growth requirements and/or may be present in a low concentration in this biological fluid.

Fig 1. PFGE patterns of SmaI-digested genomic DNA. Lanes: 1, Lb. salivarius CELA2 (sow 7; milk); 2, Lb. salivarius CELA2 (sow 7; faecal swab); 3, Lb. salivarius CELA2 (piglet from sow 7; faecal swab); 4, Lb. reuteri CR20 (sow 8; milk); 5, Lb. reuteri CR20 (sow 8; faecal swab); 6, Lb. reuteri CR20 (piglet from sow 8; faecal swab); 7, Lb. paraplantarum CLB7 (sow 5; milk); 8, Lb. paraplantarum CLB7 (sow 5; faecal swab); 9, Lb. paraplantarum CLB7 (piglet from sow 5; faecal swab); L, LowRange PFGE marker (New England BioLabs).
The elucidation of the origin of the lactobacilli present in mammal milk will be an attractive research target in the next years. Traditionally, it was considered that they were acquired by skin or faecal contamination. Obviously, sampling of milk for microbiological analysis must take into account that skin contamination is almost unavoidable and that doubts about the original location (internal mammary gland, skin) of the isolated bacteria may arise; however, no lactobacilli could be isolated from skin swabs obtained from the sows. Previously, it has been reported that lactobacilli isolates present in human milk are genotypically different from those isolated in the skin within the same host (Martín et al. Reference Martín, Langa, Reviriego, Jiménez, Marín, Xaus, Fernández and Rodríguez2003). The suggestion that the origin of the live bacteria found in milk could be the maternal gut and the bacteria would arrive to the mammary gland through an endogenous route (the so-called entero-mammary pathway), involving maternal dendritic cells and macrophages (Martín et al. Reference Martín, Langa, Reviriego, Jiménez, Marín, Olivares, Boza, Jimenez, Fernandez, Xaus and Rodriguez2004), has been confirmed recently (Perez et al. Reference Perez, Doré, Leclerc, Levenez, Benyacoub, Serrant, Segura-Roggero, Schiffrin and Donnet-Hughes2007). These authors showed that fresh human milk contains a number of viable bacteria and a wide range of free bacterial DNA signatures, which may program the neonatal immune system. More studies will be required to confirm the existence of such bacterial transfer mechanism. In vivo assays are in progress to confirm the probiotic potential of some of the lactobacilli strains isolated from sow milk.
We are grateful to Eduardo González for giving us permission to collect the biological samples analyzed in this study. This study was supported by grants FUN-C-FOOD (Consolider-Ingenio 2010) and AGL2007-62042 from the Ministerio de Educación y Ciencia y Tecnología (Spain).






