


Efforts have been made in research to increase milk unsaturated fatty acids (FA). Feeding oilseeds rich in polyunsaturated fatty acids (FA) such as soybean, sunflower and flaxseed can increase the proportion of total unsaturated FA in bovine milk fat (Glasser et al., Reference Glasser, Ferlay and Chilliard2008), however, milk with increased unsaturated FA is more prone to lipoperoxidation (De Lima et al., Reference De Lima, Santos, Schogor, de Marchi, de Souza, Santos, Santos and Petit2015). Thus, many studies have focused on supplying lactating dairy cows with antioxidants in order to prevent milk lipoperoxidation. However, effects of dietary antioxidants supplementation on milk antioxidants content and milk oxidative stability have been variable. Although some studies (Lima et al., Reference Lima, Palin, Santos, Benchaar, Lima, Chouinard and Petit2014; De Lima et al., Reference De Lima, Santos, Schogor, de Marchi, de Souza, Santos, Santos and Petit2015) have shown no effect of dietary antioxidants on milk lipoperoxidation others have shown positive effects that encourage further studies of dietary strategies to improve the oxidative stability of milk enriched with unsaturated fatty acids. Yerba Mate (Ilex paraguariensis) is a plant of the Aquifoliaceae family widely produced in southern Latin American countries. This plant is rich in polyphenols with antioxidant properties (Heck and de Mejia, Reference Heck and de Mejia2007). When fed to dairy cows, Yerba Mate increased the antioxidant activity in milk which was measured via reducing power analysis (Santos et al., Reference Santos, Zeoula, dos Santos, Lima, Dias, Rufino, Schogor, De Marchi and Petit2017).
Additionally, results from a previous study (Gobert et al., Reference Gobert, Martin, Ferlay, Chilliard, Graulet, Pradel, Bauchart and Durand2009) have suggested a preventive role of antioxidant mixtures (with lipophilic and hydrophilic properties) against lipoperoxidation damage generated by high oxidative stress. In this study, authors tested the effects of dietary vitamin E (lipophilic) and a plant extract, rich in polyphenol (hydrophilic), on plasma lipoperoxidation and this association slightly protected cow plasma against lipoperoxidation damage when they were fed flaxseed.
Thus, we hypothesized that the association of dietary vitamin E and Yerba Mate could prevent or decrease oxidation of milk enriched in unsaturated fatty acids (FA). This study was conducted to evaluate the distinct effects of dietary Yerba Mate and vitamin E and their interaction on: (1) milk composition, (2) milk concentration of polyphenols and (3) milk oxidative stability addressed by TBARS, conjugated diene and reducing power analyses.
Material and methods
Four multiparous lactating Holstein cows were used in a 4 × 4 Latin square design with four 28-day periods balanced for residual effects. Cows were housed in individual tie stalls and had free access to water. Treatments (Table 1) were planned according to a 2 × 2 factorial arrangement: (1) control diet (CO) with no Yerba Mate or vitamin E; (2) diet containing 375 IU/kg vitamin E on DM basis (VE); (3) diet containing 30 g/kg Yerba Mate on DM basis (YM); and (4) diet containing 375 IU/kg vitamin E and 30 g/kg Yerba Mate on DM basis (YE). Experimental doses of Yerba Mate and vitamin E were chosen based on previous studies (Gobert et al., Reference Gobert, Martin, Ferlay, Chilliard, Graulet, Pradel, Bauchart and Durand2009; Santos et al., Reference Santos, Zeoula, dos Santos, Lima, Dias, Rufino, Schogor, De Marchi and Petit2017; respectively) where these ingredients were used. To increase unsaturated fatty acids in milk, cows were fed 172 g/kg soybean seeds (on a dry matter basis). Soybean was ground through a 10-mm screen using a Nogueira dpm-2 chopper (Irmão Nogueira S.A. Máquinas Agrícolas e Motores, Itapira, São Paulo, Brazil). The vitamin E (DL-α-tocopherol acetate) was incorporated in the concentrate and then in the total mixed diet. Yerba Mate was mixed in TMR and it was a commercially available product (Laranjeiras, Laranjeiras do Sul, PR, Brazil) mainly composed of leaves and containing (on DM basis) 146 g/kg crude protein, 27 g/kg ether extract and 460 g/kg neutral detergent fibre (aNDF). The polyphenolic profile of Yerba Mate consisted of 216.33 mg/g gallic acid equivalent of chlorogenic acid, 35.49 mg/g quercetin equivalent of rutin, 6.33 mg/g gallic acid equivalent of caffeine, and 0.51 mg/g of caffeic acid. At the beginning of the experiment, the cows averaged (mean ± sd) 62 ± 15 d in milk and 549 ± 57 kg body weight. Diets were offered twice a day (0700 and 1500 h) for ad libitum intake (100 g/kg refusals on as fed basis) and milking times were 0600 and 1600 h. Diets were formulated to meet requirements for cows producing 25 kg/d of milk containing 3.8% of fat (NRC, 2001). The Animal Care and Use Committee of Universidade Estadual de Maringá approved all animal procedures.
Table 1. Ingredients and chemical composition of total mixed diets of Holstein cows fed no vitamin E and no Yerba Mate (CO), vitamin E and no Yerba Mate (VE), no vitamin E with Yerba Mate (YM) or a mixture of vitamin E and Yerba Mate (VY)
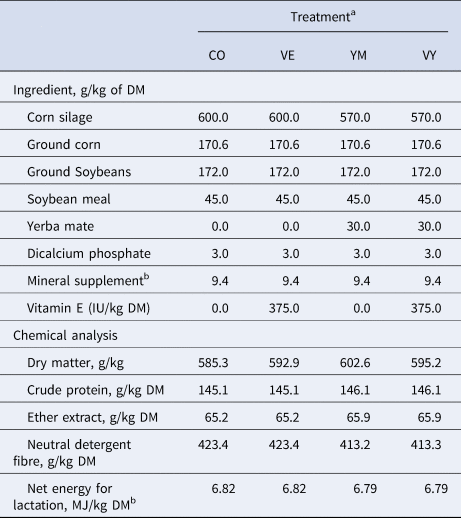
a Contained (per kg of product): Ca 240 g; P 60 g; Mg 15.0 g; S 18.0 g; Na 78.0; Fe 2.200 mg; Zn 3.800 mg; Cu 680 mg; Mn 1.105 mg; I 40 mg; Co 10 mg; Se 25 mg; vitamin A 100.000 IU; vitamin D3 66.700 IU; and vitamin E 1.000 IU.
b Calculated using published values of feed ingredients (NRC, 2001).
Each experimental period consisted of 28 d and feed intake and milk production were recorded daily throughout the experiment. Total mixed diets, refusals and Yerba Mate were sampled daily from d 15 to 21 and stored at −20 °C for subsequent drying at 55 °C. Milk samples were obtained from the four consecutive milkings on day 27 and 28 and pooled on a yield basis. Two aliquots of each milk samples were taken. One aliquot was kept frozen at −20 °C with Na azide (0.2 g/kg) for determination of antioxidant-related analyses. The second aliquot of milk samples was stored at +4 °C with bronopol-B2 until analysis of normal composition (fat, protein, and lactose).
Dry matter, total N, ether extract and ashes in feed and orts were analysed according to AOAC (1990) methods. Determination of neutral detergent fibre (aNDF) was performed according to Van Soest et al. (Reference Van Soest, Robertson and Lewis1991). Protein, fat, lactose, and urea N concentrations in milk were determined by infrared spectroscopy (Bentley model 2000; Bentley Instrument, Inc., Chaska, MN, USA) following procedure 972.16 of AOAC (1990). Milk somatic cell counts (SCC) were obtained using an electronic counter (Somacount 500®, Chaska, MN, USA).
The polyphenolic profile in Yerba Mate was analysed as previously described by Santos et al. (Reference Santos, Zeoula, dos Santos, Lima, Dias, Rufino, Schogor, De Marchi and Petit2017). Total polyphenols, production of conjugated diene hydroperoxides, and total reducing power were determined as previously reported by De Lima et al. (Reference De Lima, Santos, Schogor, de Marchi, de Souza, Santos, Santos and Petit2015). The TBARS concentration in milk was performed following the procedures described by Payá et al. (Reference Payá, Halliwell and Hoult1992).
Results were analysed using the MIXED procedure of SAS (2000) following the general model:
 $$Y_{ijk} = {\rm \mu} + C_i + P_j + T_k + e_{ijk}$$
$$Y_{ijk} = {\rm \mu} + C_i + P_j + T_k + e_{ijk}$$where Y ijk, the dependent variable; μ, overall mean; C i, random effect of cow (i = 1 to 4); P j, fixed effect of period (j = 1 to 4); T k, fixed effect of treatment (k = 1 to 4); and e ijk, random residual error. Treatments were compared to provide factorial contrasts: (1) with vs. without vitamin E, (2) with vs. without Yerba Mate, and (3) the interaction between vitamin E and Yerba Mate. Results are reported as least square means and significance was declared at P ≤ 0.05.
Results and discussion
In the present study, we were primarily interested in investigating the effects of combined dietary supplementation of vitamin E and Yerba Mate on milk antioxidant properties. With the purpose of changing milk oxidative stability, cows were fed ground soybeans in order to provide 34 g of oil/kg DM. Previous studies have shown that abomasal infusion of 8.3 g (Lima et al., Reference Lima, Palin, Santos, Benchaar, Lima, Chouinard and Petit2014) and 11.8 g (De Lima et al., Reference De Lima, Santos, Schogor, de Marchi, de Souza, Santos, Santos and Petit2015) of soybean oil/kg DM was able to increase lipoperoxidation in plasma and milk of dairy cows.
We did not observe interaction (P > 0.05) between vitamin E and Yerba Mate supplementation for milk antioxidant-related factors (polyphenols, reducing power, conjugated dienes, and TBARS) (Table 2). A previous study (Gobert et al., Reference Gobert, Martin, Ferlay, Chilliard, Graulet, Pradel, Bauchart and Durand2009) has shown that the combination of lipophilic and hydrophilic antioxidants (vitamin E and plant extract rich in polyphenol, respectively) reduced conjugated diene oxidation rate suggesting a protective effect against plasma lipoperoxidation damage when cows were fed with flaxseed as a source of polyunsaturated FA. We expected that feeding cows vitamin E and Yerba Mate as a source of lipophilic and hydrophilic antioxidants respectively would produce similar effects in milk. However, this was not observed in the present study. In fact, we did not observe any effect of vitamin E even when it was supplemented without Yerba Mate, which disagrees with previous effects observed in plasma of dairy cows by Gobert et al. (Reference Gobert, Martin, Ferlay, Chilliard, Graulet, Pradel, Bauchart and Durand2009). Although we did not analyse vitamin E in plasma and in milk, it is possible that dietary vitamin E was not transferred to milk which could have accounted for the lack of effect we observed. Further study would be necessary to investigate this possibility.
Table 2. Dry matter intake, milk production, and composition of Holstein cows fed no vitamin E and no Yerba Mate (CO), vitamin E and no Yerba Mate (VE), no vitamin E with Yerba Mate (YM) or a mixture of vitamin E and Yerba Mate (VY)

GAE, gallic acid equivalent; MDA, malonaldehyde.
a Experimental dose: Yerba Mate = 30 g/kg DM; vitamin E = 375 UI/kg DM.
b Somatic cell counts (log10).
On the other hand, milk reducing power was increased (P < 0.05) when cows were supplemented with Yerba Mate. This agrees with our previous finding (Santos et al., Reference Santos, Zeoula, dos Santos, Lima, Dias, Rufino, Schogor, De Marchi and Petit2017). In the very same study, authors reported no effects on milk antioxidants other than for reducing power which also agrees with our observations as no effects (P > 0.05) for polyphenols, conjugated dienes and TBARS were observed. This is interesting because as reducing power was changed, other antioxidant related data are expected to be affected as well. This is not completely understood, but as previously reported by Parejo et al. (Reference Parejo, Codina, Petrakis and Kefalas2000) antioxidant properties of single compounds within a group can vary markedly, suggesting that similar levels of phenolics do not necessarily correspond to the same antioxidants responses. Furthermore, total polyphenol could not precisely express the radical-scavenging capacity as the response of phenolics in the Folin-Ciocalteu assay depends on their chemical structure (Parejo et al. Reference Parejo, Viladomat, Bastida, Rosas-Romero, Flerlage, Burillo and Codina2002). Perhaps, future studies should look at new techniques to better approach and understand antioxidants in milk.
There was no interaction (P > 0.05) between vitamin E and Yerba Mate supplementation for dry matter intake, milk production, milk composition (fat, protein, lactose, and total solids), urea N, and somatic cell score (Table 2). Intake of dry matter was reduced (P < 0.05) when vitamin E was supplemented. However, milk composition in fat and lactose were not affected (P > 0.05) by vitamin E supplementation. This suggests that such statistical differences for dry matter intake had no physiological importance in this study.
Milk concentration of protein and total solids were decreased (P < 0.05) when cows were supplemented with Yerba Mate. This may suggest that diets containing Yerba Mate provided mammary gland with fewer amino acids for milk protein synthesis. It is well known that Yerba Mate contains tannins and that these compounds are able to decrease the proportion of rumen degradable protein (Bekele et al., Reference Bekele, Clément, Kreuzer and Soliva2009). Although microbial protein synthesis was not addressed in the present study, we hypothesized that tannins could have reduced microbial protein synthesis in the rumen, and thus possibly reduced the availability of amino acids for mammary protein synthesis.
In conclusion, our results suggest that the association of dietary vitamin E and Yerba Mate does not help to prevent or decrease oxidation of milk enriched in unsaturated FA. However, we have confirmed that supplementation with Yerba Mate enhances milk oxidative stability.
Acknowledgements
The present study was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq (Brasília, DF, Brazil). The authors express their gratitude to the Associação de Criadores de Bovinos da Raça Holandesa – APCBRH (Curitiba, PR, Brazil) for assistance in milk analyses.


