


INTRODUCTION
Intercropping can be defined as the simultaneous cultivation of two or more crops on the same area of land. The crops can be sown together or at different times, but they are usually grown simultaneously for a considerable proportion of their growing periods. Intercropping has been shown to increase yield compared with sole crops in low-input systems (Ofori & Stern Reference Ofori and Stern1987), reduce the chance of crop failure (Anil et al. Reference Anil, Park, Phipps and Miller1998) and can reduce nitrous oxide and leaching losses both during the growth of the intercrop and during following seasons (Pappa et al. Reference Pappa, Rees, Walker, Baddeley and Watson2011). Intercropping has the potential to improve yield stability (Hauggaard-Nielsen et al. Reference Hauggaard-Nielsen, Andersen, Jørnsgaard and Jensen2006) and plant resource utilization of water, light and nutrients (Willey Reference Willey1990; Jensen Reference Jensen1996a ; Whitmore & Schroder Reference Whitmore and Schroder2007; Xu et al. Reference Xu, Li and Shan2008; Zhang et al. Reference Zhang, Van Der Werf, Bastiaans, Zhang, LI and Spiertz2008). Where legumes are included as an intercrop, nitrogen transfer from biological nitrogen fixation (BNF) has also been reported (Jensen Reference Jensen1996b ; Hauggaard-Nielsen & Jensen Reference Hauggaard-Nielsen and Jensen2001; Corre-Hellou et al. Reference Corre-Hellou, Brisson, Launay, Fustec and Crozat2007). Intercropping of barley (Hordeum vulgare L.) with pea (Pisum sativum L.) has been shown to improve the use of plant growth resources when compared with the associated sole crops due to complementary morphological and physiological characteristics (Willey Reference Willey1979; Hauggaard-Nielsen & Jensen Reference Hauggaard-Nielsen and Jensen2001).
Intercropping is of significant interest in low-input and organic systems (Jones & Clements Reference Jones and Clements1993; J. C. Pridham & E. Martin, personal communication). However, there is an urgent global challenge in providing sufficient primary production to sustain a growing population with growing demands for foods, feeds and fuels without exacerbating climate change and other environmental impacts of agriculture (Tilman et al. Reference Tilman, Fargione, Wolff, D'antonio, Dobson, Howarth, Schindler, Schlesinger, Simberloff and Swackhamer2001); issues that drive farmers to consider alternative sources and approaches to N management such as intercropping (Tilman et al. Reference Tilman, Cassman, Matson, Naylor and Polasky2002; Erisman et al. Reference Erisman, Sutton, Galloway, Klimont and Winiwarter2008)/mixtures (Bebawi & Naylor Reference Bebawi and Naylor1978; Kaut et al. Reference Kaut, Mason, Navabi, O'donovan and Spaner2009); minimizing external inputs (e.g. fertilizers and pesticides) and more efficient use of nitrogen (N), e.g. from BNF, can increase the economic, environmental and social sustainability of agricultural systems.
In developing more sustainable cropping systems it is important to assess the stability of crop yield and N dynamics over time (whole rotations or periods of several years as well as within growing seasons). To date, most studies of intercrops have based their conclusions only on measurements of final yield (Connolly et al. Reference Connolly, Wayne and Bazzaz2001) and data from only one growing season (Hauggaard-Nielsen & Jensen Reference Hauggaard-Nielsen and Jensen2001; Andersen et al. Reference Andersen, Hauggaard-Nielsen, Ambus and Jensen2005). More recently, Hauggaard-Nielsen et al. (Reference Hauggaard-Nielsen, Gooding, Ambus, Corre-Hellou, Crozat, Dahlmann, Dibet, Von Fragstein, Pristeri, Monti and Jensen2009) found that overall use of N resources was 30–40% more efficient by pea-barley compared with the respective sole crop across Europe using the same cultivars. Few studies (e.g. Hauggaard-Nielsen et al. Reference Hauggaard-Nielsen, Ambus and Jensen2003; Pappa et al. Reference Pappa, Rees, Walker, Baddeley and Watson2011) have considered the dynamics over a longer period (greater than one year) and none has studied the carry-over effect of intercropping on subsequent crops within an arable rotation.
The main objectives of the present experiment were: (a) to evaluate the effects of different legumes (species and varieties) and barley on grain yield, dry matter production and N uptake of the intercrop treatments compared with the associated cereal sole crop; (b) to assess the effects on yield of the next grain crop and (c) to determine the accumulation of N in harvested crops in a low-input rotation.
MATERIALS AND METHODS
Site description and experimental design
The field experiment was sited at the Bush Estate (55°51′N, 3°12′W), near Edinburgh, Scotland, UK. This drained-plot experimental facility was established in 1990 and consists of 12 hydrologically isolated plots (25×9 m) (Vinten et al. Reference Vinten, VIVIAN and HOWARD1992). Prior to the current experiment, the plots had been left fallow for 3 years (2003–06). The soil is a sandy loam (Macmerry Series). In 2006, the water content at field capacity was 19±0·5% (v/v), the soil bulk density was 1·19±0·01 t/m3 (mean±s.e., n=5), the 1M KCl extractable N in the soil was 4·8 mg NH4 +–N/kg and 21·9 mg NO3 −–N/kg in 0–200 mm soil depth. The rainfall and air temperature for the 3 years of the current study are shown in Table 1. The average annual precipitation for the last 25 years (1984–2009) was 676·2 mm.
Table 1. Monthly total rainfall (mm) and monthly average air temperature (°C) for Bush Estate, Edinburgh, UK

The treatments were established in a complete randomized design: spring barley (H. vulgare cvar Westminster) as a sole crop; spring barley/spring pea (P. sativum cvar Zero4) intercrop; spring barley/spring pea (P. sativum cvar Nitouche) intercrop; spring barley/white clover (Trifolium repens cvar Alice) intercrop in 2006. Sole crops of pea cvars Nitouche and Zero4 were sown at the side of the plots for only the final grain yield comparisons and land equivalent ratio (LER) calculations. The sowing date for these treatments was 24 April 2006. In the second growing season (2007), all the plots were sown with spring oat (Avena sativa cvar Firth) on 3 April. In the third growing season, perennial ryegrass (PRG) was sown in all plots in a 50:50 mixture of cultivars Aberavon and Aberdart at a total seed rate of 35 kg/ha. The plots were tilled using a mouldboard plough followed by cultivating (rotary hoe), seeding and rolling. No fertilizers, herbicides or pesticides were used and although weed, pests and diseases were monitored, they did not constitute a serious problem. During the winter prior to sowing the oats (2006/2007), the clover continued to grow well after harvest due to mild winter temperatures and was incorporated into the soil by ploughing in the spring (3 April 2007).
Each treatment was replicated three times. In the intercrop treatments, the seed rates followed a 50:50 replacement design. This means that the target intercrop density was 0·50 of the monoculture density of each crop. Based on sole cropping seed rates of 200 kg/ha for barley (c. 350 germinable seeds/m2), 250 kg/ha for pea (c. 75–110 germinable seeds/m2) and 250 kg/ha oat (c. 450–500 germinable seeds/m2) and 5 kg/ha of white clover. These were based on typical seed rates for the specified crops grown in the region where the trial is located (McBain Reference Mcbain2010).
The varieties that were used had contrasting characteristics: Westminster is a medium-tall barley variety that is widely grown for malting and feed (HGCA 2005/06). Nitouche is a large blue pea with a consistent performance, good agronomic characters; long straw and good standing ability. Zero4 is a small blue combining pea with a unique combination of agronomic characters; short straw, excellent standing ability and very early maturity. Alice is a tall, large leaved white clover used in pasture mixtures for exceptional yields of palatable, high-quality and high-protein forage. Firth has a high kernel content and good resistance to mildew (HGCA 2005/2006).
Harvest and analysis
Above-ground plant material was collected just prior to crop harvest (cereals/peas on 11 September 2006, oats on 11 September 2007 and grass for silage on 26 June 2008) by cutting the plants 50 mm above the soil surface from a 1 m2 area (four 0·25 m2 quadrats) randomly placed in each plot. Biomass samples were separated into barley, pea, clover and weeds dried at 70°C for 24 h and weighed. For grain yield, crop plants were hand threshed at the time of crop maturity and grain yield was calculated at 0·85 dry matter. Each part (stems, ears and pods) of each crop was weighed and sub-samples were taken for determination of the total N concentration (mg/g) of the above-ground material. The oven-dried samples were initially prepared using a Glen Creston hammer mill (London, UK) with a 1 mm mesh. A sub-sample of this was then ball milled to a flour-like consistency using a Retch ball mill (West Yorkshire, UK). These samples were analysed for total N by combustion using a Carbo Erba NA 1500 analyser (Erba Science UK). Nitrogen accumulation and grain N yield were calculated by multiplying dry matter and grain yield, respectively, by their corresponding N concentration. The same procedure was used for the spring oat crop (2007), which was separated into straw and grain, and for the ryegrass crop (2008).
Calculations
LER
An important tool for the study and evaluation of intercropping systems is the LER (Willey Reference Willey1979; Dhima et al. Reference Dhima, Lithourgidis, Vasilakoglou and Dordas2007). LER provides a measure of the yield advantage obtained by growing two or more crops or varieties as an intercrop compared to growing the same crops or varieties as a collection of separate sole crops. The LER for the barley/pea intercrop is calculated using the following formula:

where L B and L L are the land for barley and legumes, respectively, Y Bint and Y Bmon are the grain yield of barley in intercrops and sole crop, respectively, and Y Lint and Y Lmon are the grain yields of the legume in the intercrop and sole crop, respectively. An LER value of 1·0 indicates no advantages in yield between the intercrop and sole crop. Any value greater than 1·0 shows a yield advantage for intercropping. An LER of 1·3, for example, indicates that the area planted to monocultures would need to be 30% greater than the area planted to intercrop to produce the same combined yield. The calculation of LER is the most common method adopted in intercropping studies, in particular in tropical regions where intercropping is commonly practised.
For barley plants (2006), the cereal growth stages GS23 (main shoot and three tillers, Zadoks et al. Reference Zadoks, Chang and Konzak1974), GS65 (flowering half-way), GS77 (late milk) and GS92 (grain hard (not dented by nail)) were 30, 70, 100 and 140 days from sowing, respectively. For oat plants (2007), the growth stages GS16 (six leaves unfolded), GS23, GS65, GS77 and GS92 were 40, 65, 100, 135 and 150 days from sowing, respectively.
Statistical analysis
All variables measured were normally distributed (checked using the Anderson-Darling test at P⩽0·05). Statistical analyses, using ANOVA, regarded the treatments as fixed effects in the completely randomized plot layout. The software programs used for the analysis were Genstat 8 and Minitab 15. In all cases, significant differences were defined as those with P⩽0·05.
RESULTS
Growth stages and height
Barley plants
During the 2006 intercropping season, barley height was influenced by companion variety choice and agronomic practice. Barley height was influenced by treatment, being tallest in the barley/pea cvar Nitouche treatment followed by barley in the barley/clover treatment. The height of the barley intercropped with the Nitouche pea was 0·29 m taller than barley grown with the pea cvar Zero4 (P<0·05) at harvest (GS92) (Fig. 1(a)).

Fig. 1. Height of spring barley in 2006 (a) and spring oat in 2007 (b) plotted against growth stages. All values are means (n=3)±s.e. (bars).
Spring oat plants
Oats grown on the previous year's barley/clover treatment were observed to be the strongest and tallest plants. The two pea intercrops were 0·14 m taller than the barley/pea cvar Nitouche grown in the previous year (P<0·05) (Fig. 1(b)).
Above-ground biomass
Spring barley, spring oat, weeds, legumes and ryegrass
The barley grew rapidly in 2006 between GS39 and GS87 after the canopy had closed, generating almost 0·70 of its total final dry matter in this phase. At the end of grain filling (GS85), when the maximum crop dry weight occurred, the stems and leaves started losing weight due to falling leaf tissue and senescence varied by 7 days between sole crop and intercropped treatments. The barley had the maximum dry biomass in the barley/clover treatment. The senescence started earlier in the barley sole crop and its dry biomass reduced much quicker than the intercrops (Fig. 2(a)). In 2006, the biomass of weeds in the barley treatment was significantly greater (P<0·05) than that in the barley/pea cvar Zero4 and barley/clover treatments (Fig. 2(c)). The greatest above-ground legume dry biomass was in pea cvar Nitouche (0·7 t/ha) (P<0·05) and the least was in clover (0·2 t biomass/ha) in June 2006. The dry biomass of the two pea varieties was similar in the 2006 growing season, even though the pea cvar Nitouche was double the height of pea cvar Zero4 (Fig. 2(e)).

Fig. 2. Above-ground biomass of spring barley in 2006 (a), spring oat in 2007 (b), weeds in 2006 (c) and 2007 (d) and legumes in 2006 (e) plotted against growth stages and PRG in 2008 (f) at the first cut. All values are means (n=3)±s.e. (bars) within the treatment.
In 2007, the growth of oat plants was significantly affected by the previous crop treatment. Oat grown following barley/clover started to grow earlier than the other treatments and had the greatest dry biomass (17 t/ha) at GS92 (Fig. 2(b)). The biomass of weeds growing in the previous treatment planted with barley as a sole crop, was greater than that in other treatments throughout the second growing season (P<0·05) (Fig. 2(d)).
In 2008, the above-ground dry biomass for PRG was similar in all treatments. All the treatments had close to 1·3 t dry matter/ha at the summer harvest (Fig. 2(f)).
Grain yields
Spring barley and pea intercrops during the first year of the cropping sequence (2006)
The pea cultivars differed in time of flowering/maturity and stem length, with Zero4 being the earliest maturing and shortest (nearly 0·40 m tall). The composition of the plant population of the established intercrops was close to the target of 50:50 with 162 plants/m2 for barley (intercropped), 47 pea cvar Nitouche (intercropped), 56 pea cvar Zero4 (intercropped) and 261 barley (sole crop). The grain yield of the barley intercropped with clover was significantly different (P<0·01) reaching 127% of the average barley sole crop yield. However, there was no significant difference between the two barley/pea treatments for the grain yields of barley and pea (Fig. 3(a)).

Fig. 3. Grain yields of spring barley and pea in 2006 (a) and spring oat in 2007 (b). All values are means (n=3)±s.e. (bars) within the treatment.
Spring pea sole crop during the first year (2006)
There were no significant differences in the yield of the pea sole crop during the first growing season (pea cvar Nitouche: 3·1 t grain/ha and pea cvar Zero4: 2·9 t grain/ha) at the harvest stage.
Spring oat in the second year of the cropping sequence (2007)
The spring oat grain yield was influenced by the previous year's treatments with a significant difference (P<0·05) between the oat grown after the clover/barley (6·7 t grain/ha) treatment and the other treatments (Fig. 3(b)).
Production efficiency of intercrops
The LER as an indicator of biological efficiency in intercropping systems were always greater than 1 with the intercropping treatments used in the present study. The average LER for the clover/barley treatment was 1·27 compared to 1·48 for barley/pea cvar Zero4 and barley/pea cvar Nitouche, but these differences were not statistically different (Fig. 4). However, it should be noted that the LER of barley/clover in a 50:50 proportion had a higher production than the sole crop.

Fig. 4. LER for the intercrops (2006) based on the grain yield. All values are means (n=3)±s.e. (bars).
Nitrogen accumulation in crops
Spring barley
The total above-ground N accumulation in barley was not significantly different between treatments in 2006 until harvest (P<0·05), when barley/pea cvar Nitouche reached 52 kg N/ha in the above-ground biomass (Fig. 5(a)). Barley straw biomass did not differ between treatments, but there were significant differences (P<0·05) in total N in the barley grain yield with the barley/pea cvar Zero4 treatment having the highest value (46 kg N/ha) (Fig. 6).

Fig. 5. Accumulation of N in above-ground biomass at different growth stages for spring barley (2006) (a) and spring oat (2007) (b). All values are means (n=3)±s.e. (bars).

Fig. 6. Accumulation of N in above-ground parts (straw and grain) of spring barley and legumes (clover and peas) expressed as kg N/ha in 2006. All values are means (n=3)±s.e. (bars).
Legumes (clover and pea)
The pea in the barley/pea cvar Nitouche treatment had the highest levels of total N (98 kg N/ha) at harvest with 79 (P<0·01) and 19 kg N/ha in grain and straw, respectively (Fig. 6). However, the pea in the barley/pea cvar Zero4 treatment had a total N uptake of 83 kg N/ha (63 kg N/ha in grain and 20 kg N/ha in straw) followed by the clover plants with 79 kg N/ha (Fig. 6). Early in the 2006 growing season, the total N accumulation in the legumes was significantly different (cereal GS10 and GS23) (Fig. 7).

Fig. 7. Accumulation of N in above-ground biomass of the legumes according to cereal (barley) growth stages in 2006. All values are means (n=3)±s.e. (bars).
Spring oat
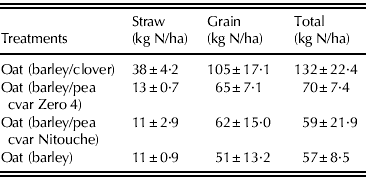
Total above-ground N accumulation was not significantly different between treatments at the beginning of the second growing season (2007). However, 8 weeks after sowing, the oat plants growing on the previous season's barley/clover plots had significantly (P<0·001) more total N in the above-ground parts (143 kg N/ha) than the other treatments. This difference continued until the end of the growing season (Fig. 5(b)). The total N in the oat growing in the previous two barley/pea plots was significantly different at GS23 (P<0·05), containing 71 and 53 kg N/ha on the plots growing barley/pea cvar Nitouche and barley/pea cvar Zero4, respectively. At the end of the growing season, the total N in straw for the spring oats grown in the previous barley/clover treatment was significantly different from the rest of the treatments, reaching almost 38 kg N/ha. The total N contained in the grain did not differ between the plots where pea crops and monoculture barley were grown in the previous year. The only difference was between plots in which the previous crop had been clover/barley and the barley sole crop (Table 2).
Table 2. Accumulation of N in above-ground parts (straw and grain) of spring oats (2007) expressed as kg N/ha. All the values are the mean (n=3)±s.e.
DISCUSSION
The interactions between intercropped cereals and legumes are affected both by differences in the morphology and physiology of the species and through environmental controls. Barley plants intercropped with clover had the highest biomass and yield (Figs 2(a) and 3), and the barley grown in the two barley/pea treatments was equal in biomass, but both had smaller yields than the barley grown in the barley/clover treatment (Fig. 2(a)). This was probably a result of the taller pea plants increasing the interspecific competition for light during vegetative growth compared with the lower growing clover. Barley and legume plants respond differently to light and cereal plants can cause shading, which leads to growth restrictions in legume plants (Fujita et al. Reference Fujita, Ofosu-Budu and Ogata1992). In addition, competition for soil water during grain filling of the barley and pea (Thorsted et al. Reference Thorsted, Olesen and Weiner2006), as well as possible higher levels of available N in the soil and/or the effect of shading of the weed understorey during the growing period from the clover (Grashoff & d'Antuono Reference Grashoff and D'antuono1997), may have influenced this outcome. However, the average temperature and rainfall during the growing season were within the range for the last 25 years (1985–2010) for the area.
The competition between the barley and pea reduced the amount of above-ground biomass in the barley intercropped with pea cvar Nitouche, with the barley being more shaded than when grown with the shorter pea cvar Zero4. It is probable that the amount of photosynthetically active radiation (PAR) received by the legumes was decreased as a result of competition for this resource with barley, which would reduce the rate of photosynthesis and probably also result in decreased BNF. Pea cvar Zero4 was the shorter of the two pea cultivars and it is possible that its growth and physiological activity was affected by shading, more so than the pea cvar Nitouche. Less shading of the legume component in an intercropped system may increase both the photosynthesis rate of the legume as well as the rate of BNF (Fujita et al. Reference Fujita, Ofosu-Budu and Ogata1992).
The nutrient dynamics of both cereals and legumes would be expected to be influenced when they are grown as intercrops. The legume has the potential to provide nitrogen (N) to the non-legume directly through mycorrhizal links, root exudates, or decay of roots and nodules (Jensen Reference Jensen1996a ; Vandermeer Reference Vandermeer1999; Webb et al. Reference Webb, Jensen, Heywood, Morris, Linton and Hooker2010). Another possible mechanism is that legumes can ‘bank’ large quantities of soil N, which might otherwise have leached out of the system, and release it through soil organic matter turnover to the non-legume companion crop later during the growing season, or to the following crops (Vinten et al. Reference Vinten, VIVIAN and HOWARD1992). In pea, there is evidence to indicate that the process of BNF begins approximately a month after sowing and lasts for up to 2 months (Balandreau & Dommergues Reference Balandreau and Dommergues1973) depending on inoculants, climate and soil conditions (Ledgard & Steele Reference Ledgard and Steele1992). If this were the case in the present study then it would suggest that for an early variety such as pea cvar Zero4, BNF would have continued during seed development while for a later variety (e.g. Nitouche), it would have stopped after flowering (Cousin Reference Cousin1997). Anil et al. (Reference Anil, Park, Phipps and Miller1998) found that the amount of N2 fixed by legumes generally declines with increasing soil N availability, and if the legumes are continuously shaded their ability to fix N2 is further impaired (Willey Reference Willey1979). The two pea cultivars had different growth rates and biomass even though at the harvest stage they had similar grain yields (Figs 2(a) and 4).
The LER values indicate good resource use efficiency by the intercrops included in the present study (Vandermeer Reference Vandermeer1999; Hauggaard-Nielsen & Jensen Reference Hauggaard-Nielsen and Jensen2001). Intercropping can also provide improvements in soil quality and more stable yields (Yildirim & Guvenc Reference Yildirim and Guvenc2005). The LER values were also used to compare cultivar performance in the intercrops, relative to the barley/pea and barley sole crop, with gains of up to 22% (Hauggaard-Nielsen & Jensen Reference Hauggaard-Nielsen and Jensen2001). The present results indicate comparable values up to 27 and 42% for the barley/pea and barley/clover intercrop, respectively, compared to the barley alone, representing a significant yield benefit for these treatments (Fig. 4). The majority of previous studies have also shown LERs of greater than one from intercropping experiments. For instance, Dariush et al. (Reference Dariush, Ahad and Meysam2006) recorded LERs of 1·19 when working with maize and kale. Research on wheat (Triticum aestivum) and field bean (Vicia faba) by Bulson et al. (Reference Bulson, Snaydon and Stopes1997) estimated LERs of significantly greater than 1 in situations where crop densities were sufficiently high, whereas Newman (Reference Newman1986), studying vegetables and fruit, found LERs more than 2 under some circumstances. The increased LERs that are observed in intercropping experiments are likely to be a consequence of a number of interacting factors. However, given that the companion species inevitably occupy different ecological niches, it is likely that there will be some increased exploitation of resources (light, water and nutrients).
There were significant differences in the accumulation of N in the barley plants between treatments during the first growing season (2006), indicating a possible effect of available N in the soil and N inputs by fixation (Bandyopadhyay & De Reference Bandyopadhyay and De1986). Many studies have been conducted using barley to elucidate the factors controlling N uptake. It has been shown that different yield response in barley is linked to the crop's varying N uptake and its N use efficiency (Perby & Jensén Reference Perby and Jensén1983; Tillman et al. Reference Tillman, Pan and Ullrich1991; Delogu et al. Reference Delogu, Cattivelli, Pecchioni, De Falcis, Maggiore and Stanca1998). Throughout plant growth, nitrogen has a key role in dry matter formation and accumulation. Barley plants grown in intercrops have an opportunity to increase N use efficiency and therefore yield through improved soil exploitation of N resources and N transfer from companion legume species (Hauggaard-Nielsen & Jensen Reference Hauggaard-Nielsen and Jensen2001). Additionally, crop N concentration is largely determined by crop growth rate and biomass due to a ‘dilution’ phenomenon of plant nitrogen by carbon assimilates as the plant grows (Greenwood et al. Reference Greenwood, Lemaire, Gosse, Cruz, Draycott and Neeteson1990; Justes et al. Reference Justes, Mary, Meynard, Machet and Thellier-Huché1994; Gastal & Lemaire Reference Gastal and Lemaire2002).
In most cases, the crop mixture contained more N than the component sole crops, indicating improved N use by the intercrops. Other studies confirm the N uptake and efficiency benefits of growing a legume with a non-legume (Martin & Snaydon Reference Martin and Snaydon1982; Szumigalski & Van Acker Reference Szumigalski and Van Acker2008). Spatial arrangements of receiver and donor plants can be important in influencing the processes of resource use. A number of mechanisms exist that enable mixed crops to utilize growth resources more efficiently than associated sole crops. For example, different spatial arrangements can not only influence nutrient transfer directly, but also have an effect on weed and disease pressure as well as competition for light and water (Anil et al. Reference Anil, Park, Phipps and Miller1998).
The most important result from the present study is the quantification of the substantial affect the previous legume treatments had on the following oat crop. Oat growing in the previous barley/clover plots had the highest biomass, grain yield and accumulation of N in straw and grain in comparison with the other treatments (Figs 2(b), 3(b) and 5(b) and Table 2). It has been reported in other studies that where clover continues to grow after the harvest of the accompanying plant, there is the potential for reduced N loss from the soil and possible N transfer through legume residue decomposition and turnover. It has also been suggested that there are additional benefits on soil structure during the growing season and into subsequent seasons (Mytton et al. Reference Mytton, Cresswell and Colbourn1993; Papadopoulos et al. Reference Papadopoulos, Mooney and Bird2006). In addition to the grain benefits, the intercrop treatments have a positive effect on nitrogen losses. Nitrogen leaching was reduced under the legume intercrops when compared with the barley monocrop (cumulative values of 0·67 and 3·80 kg NO3–N/ha, respectively) in 2006. Nitrous oxide losses were also significantly different between the treatments and especially the two barley/pea cultivars (cumulative values of 6·02 and 2·14 kg N2O–N/ha for intercrops cvars Nitouche and Zero4, respectively) in 2006 (Pappa et al. Reference Pappa, Rees, Walker, Baddeley and Watson2011).
The management practices employed in the present study might be appropriate for the manipulation of nitrate leaching and N2O losses from agricultural ecosystems (Pappa et al. Reference Pappa, Rees, Walker, Baddeley and Watson2011). The rate and morphology of barley root development in combination with the available N in the soil might be responsible for the different N uptakes (Eghball & Maranville Reference Eghball and Maranville1993; Mengel et al. Reference Mengel, Hütsch and Kane2006; Herrera et al. Reference Herrera, Stamp and Liedgens2007). Root biomass can control the amount of N uptake by plants, as increased root biomass will enable plants to exploit a larger volume of soil (Rees et al. Reference Rees, Bingham, Baddeley and Watson2005).
Agricultural systems can benefit from improved soil structure and an associated soil microbiological community structure (Ball et al. Reference Ball, Bingham, Rees, Watson and Litterick2005). In particular, low input and organic farming systems have a high potential to benefit from the use of legumes as a part of a rotation in order to improve soil fertility and structure. Subsequent crops can then benefit by using the N stored in the soil, which is released by mineralization (Watson et al. Reference Watson, Atkinson, Gosling, Jackson and Rayns2002). The present experiment has highlighted the higher potential grain yields from the use of intercrops with additional benefits of improved yields in subsequent crops. It is likely that in addition to the environmental and agronomic benefits of intercropping systems reported in the present paper that there will be positive economic effects (Rao & Willey Reference Rao and Willey1980; Hauggaard-Nielsen & Jensen Reference Hauggaard-Nielsen and Jensen2001).
The main conclusions of the present work are that the choice of legume cultivar or species is a key factor influencing the amount of N available to the system in the year of use and/or the following year, with this impacting significantly on the final grain yield in these years. The two barley/pea intercrops had similar above-ground biomass and grain yield for both component species (barley and pea). However, the accumulation of N in the above-ground biomass of barley differed between these two treatments with significantly more N accumulated in barley/pea cvar Nitouche than the barley/pea cvar Zero4. Undersowing cereals, even at low seed rates, with clover can contribute significantly to accumulation of N in the cereal plants in the year of growth and the subsequent year. If the residues remaining after the harvest of a barley/clover crop are incorporated into the soil by ploughing in the spring, it can provide significant amounts of available N for the following crop.
Improved understanding of different species/varietal combinations and ratios will help to develop management recommendations for optimizing the productivity and environmental impacts of intercropping systems and rotations in practice.
The work was funded by the Greek State Scholarship foundation (I.K.Y) and Scottish Government RERAD through a PhD studentship grant. We thank Colin Crawford and John Parker for technical advice and support.









