



Management Implications
Undertaking restoration projects that focus on invasive plant removal without taking into consideration the broader ecological picture is a mistake. Natural ecosystems supporting predominantly native species are often characterized by a myriad of complex biotic and abiotic interactions, many of which can be substantially altered by invasion by nonnative plants. As results of many prior projects have shown, just removing invasive plants is not always enough to reverse aboveground and belowground changes effected by plant invasion. If habitat restoration projects are ultimately to be successful, resource managers need to determine what relationships may have been impacted before moving ahead with invasive removal and to include measures to remediate these invasion effects. Some adaptive restoration measures that have been discussed include soil amelioration, topsoil removal, dead biomass or litter removal, inoculation with fungal or bacterial taxa, overseeding with native plant seed, carbon addition, or selective removal of nonnative species or species with strong biogeochemical feedbacks. The restoration approach also needs to be carefully considered, as the method used to remove weeds influences legacy and treatment effects. Even if removal efforts should initially appear successful, restored systems continue to be vulnerable to invasions by new weeds or secondary invaders, so project planning should include contingency plans to address these issues as well. While the focus for many restoration projects is often on re-creating natural ecosystems that, once restored, will need no maintenance, some type of long-term maintenance will probably always be needed to control opportunistic secondary invaders that often flourish in systems disturbed by restoration activities and to potentially implement post-project “tweaks” that will maintain the project on a successful restoration trajectory. Until restoration approaches are refined to integrate measures that address both aboveground and belowground invasion effects, managers would be best served by focusing on restoring areas with fewer invasion effects or a lower potential for legacy effects or focusing on areas where these effects are more easily countered.
Introduction
Many habitat restoration projects that focus on removal of invasive plants end up foundering due to unexpected ecological complications (Corbin and D’Antonio Reference Corbin and D’Antonio2012; Konlechner and Lord Reference Konlechner and Lord2015; Suding et al. Reference Suding, Gross and Houseman2004). Either long-term maintenance is needed to eradicate the primary invaders targeted, or primary invaders are eliminated, only to be replaced by new weeds or secondary invaders (Pearson et al. Reference Pearson, Ortega, Runyon and Butler2016). Restoration is complicated by the fact that, while efforts are generally “aboveground centric,” focusing on the highly visible displacement of native plant species by non-native ones, the effects of invasives are often not, effecting equally as pernicious effects on belowground conditions (Wolfe and Klironomos Reference Wolfe and Klironomos2005). Invasives’ impacts on soils can either be chemical or microbial or a combination of both (Corbin and D’Antonio Reference Corbin and D’Antonio2012; Inderjit and van der Putten Reference Inderjit2010), with changes in soil chemistry influencing microbial communities and vice versa (Badalamenti et al. Reference Badalamenti, Gristino, Laudicina, Novara, Pasta and La Mantia2016; Ehrenfeld Reference Ehrenfeld2004). Soil changes can exert a positive feedback effect on established invasives, enhancing their competitiveness at natives’ expense (Corbin and D’Antonio Reference Corbin and D’Antonio2012; Inderjit and van der Putten Reference Inderjit2010; Reinhart and Callaway Reference Reinhart and Callaway2006), or invasives can have direct, negative feedbacks on natives by introducing or amplifying pathogens (de la Peña et al. Reference de la Peña, de Clercq, Bonte, Roiloa, Rodríguez-Echeverría and Freitas2010; Eppinga et al. Reference Eppinga, Rietkerk, Dekker, De Ruiter and van der Putten2006; Vilà et al. Reference Vilà, Tessier, Suehs, Brundu, Carta, Galanidis, Lambdon, Manca, Médail, Moragues, Traveset, Troumbis and Hulme2006), producing allelochemicals (Inderjit et al. Reference Inderjit, Seastedt, Callaway, Pollock and Kaur2008; Novoa et al. Reference Novoa, González, Moravcová and Pyšek2012; Vivrette and Muller Reference Vivrette and Muller1977), or disrupting or altering symbiotic mutualistic relationships (Gornish et al. Reference Gornish, Franklin and Rowe2020; Lekberg et al. Reference Lekberg, Gibbons, Rosendahl and Ramsey2013; Pringle et al. Reference Pringle, Rillig, Gardes and Klironomos2009; Stinson et al. Reference Stinson, Campbell, Powell, Wolfe, Callaway, Thelen, Hallett, Prati and Klironomos2006).
Effects of invasion by nonnative plants on soil chemistry have been documented for a large number of plant types and habitats, including nonnative grasses in Hawaiian woodlands and California shrublands (Mack et al. Reference Mack, D’Antonio and Ley2001; Yelenik and D’Antonio Reference Yelenik and D’Antonio2013; Yelenik and Levine Reference Yelenik and Levine2010); nonnative shrub and grass species in deciduous forests in the eastern United States (Ehrenfeld et al. Reference Ehrenfeld, Kourtev and Huang2001); and nonnative herbs in Chinese forests (Niu et al. Reference Niu, Liu, Wan and Liu2007). While these studies linked invasion to change in a wide range of soil chemistry variables, nonnative plant invasion appears to have particularly impacted nitrogen (N) dynamics within ecosystems (Ehrenfeld Reference Ehrenfeld2010; Liao et al. Reference Liao, Peng, Luo, Zhou, Wu, Fang, Chen and Li2008), which may stem partly from many studies focusing on invasion of N-fixing species into habitats without a history of N-fixers (Corbin and D’Antonio Reference Corbin and D’Antonio2011). A meta-analysis of effects of both N-fixing and non–N-fixing invasive plants on N and carbon (C) cycling in forest, grassland, and wetland ecosystems using data from 94 experimental studies and 19 soil variables found that soil ammonium (NH4+) and nitrate (NO3−) pools climbed by 30% and 17%, respectively, following invasion, while mineralization and nitrification rates jumped 52% and 53%, respectively (Liao et al. Reference Liao, Peng, Luo, Zhou, Wu, Fang, Chen and Li2008). Conversely, invasion appeared to be accompanied by less dramatic changes in soil C pools (∼7%; Liao et al. Reference Liao, Peng, Luo, Zhou, Wu, Fang, Chen and Li2008).
Coastal dunes are one ecosystem in which soil chemistry changes following invasion have been well studied, at least for certain species. Iceplant [Carpobrotus edulis (L.) L. Bolus], a common invader in many of the world’s dune systems, exerts a well-documented physiochemical influence on invaded areas by reducing soil pH and available water; increasing organic matter (OM); and in certain instances, increasing total N, NO3−, NH4+, phosphorus (P), organic C, and sodium (Na+) levels (Badalamenti et al. Reference Badalamenti, Gristino, Laudicina, Novara, Pasta and La Mantia2016; Conser and Connor Reference Conser and Connor2009; D’Antonio and Mahall, unpublished data in Conser and Connor Reference Conser and Connor2009; D’Antonio and Mahall Reference D’Antonio and Mahall1991; Novoa et al. Reference Novoa, González, Moravcová and Pyšek2013, Reference Novoa, Rodríguez, Richardson and González2014; Vilà et al. Reference Vilà, Tessier, Suehs, Brundu, Carta, Galanidis, Lambdon, Manca, Médail, Moragues, Traveset, Troumbis and Hulme2006). Carpobrotus edulis leaves have high levels of tannin and antibacterial compounds (Novoa et al. Reference Novoa, Rodríguez, Richardson and González2014; van der Watt and Pretorius Reference van der Watt and Pretorius2001) that may reduce litter decomposition rates (Vilà et al. Reference Vilà, Tessier, Suehs, Brundu, Carta, Galanidis, Lambdon, Manca, Médail, Moragues, Traveset, Troumbis and Hulme2006) and acidify soils, thereby reducing nitrification rates and increasing leaching of calcium (Ca2+) and Mg2+ (D’Antonio and Mahall, unpublished data in Conser and Connor Reference Conser and Connor2009). All these soil chemistry changes create a positive feedback effect for C. edulis, favoring it over native dune plant species (Novoa and González Reference Novoa and González2014). N-fixing shrubs such as bush lupine (Lupinus arboreus Sims) have also proven to be an issue in New Zealand dunes and northern California prairies, almost doubling N soil concentrations and increasing mineralization rates within normally N-limited systems (Hetherington and Wilson Reference Hetherington and Wilson2019; Maron and Jefferies Reference Maron and Jefferies1999; Pickart et al. Reference Pickart and Sawyer1998). Continued N deposition through L. arboreus death or leaf drop promotes habitat conversion by encouraging establishment of weedy annuals such as ripgut grass (Bromus diandrus Roth) (Maron and Connors Reference Maron and Connors1996; Maron and Jefferies Reference Maron and Jefferies1999).
Another common invader in coastal dune systems worldwide is European beachgrass [Ammophila arenaria (L.) Link], a C3 grass native to northern Europe (van der Putten et al. Reference van der Putten, Yeates, Duyts, Schreck Reis and Karssen2005; Weber Reference Weber2003). Many studies have evaluated how A. arenaria has been able to proliferate in introduced portions of its range, with most focusing on how differences in the soil microbial community between native and introduced ranges might enable this species to flourish, often at the expense of native plants and wildlife (Dangremond et al. Reference Dangremond, Pardini and Knight2010; Muir and Colwell Reference Muir and Colwell2010). Little, if any research, however, has focused on changes in soil chemistry following invasion by A. arenaria, even though, given its predilection to form dense monocultures in invaded portions of its range, this species would seemingly have a strong potential to exert an effect. A recent study at Point Reyes in northern California demonstrated that A. arenaria invasion is accompanied by distinct changes in the soil microbial community; specifically, lower abundances of putatively identified nitrifiers, fermentative bacteria, fungal parasites, and fungal dung saprotrophs and a higher abundances of cellulolytic bacteria and a class of arbuscular mycorrhizal fungi (Archaeosporomycetes; Parsons et al. Reference Parsons, Sayre, Ender, Rodrigues and Barberán2020b). Reductions in nitrifiers and changes in the proportions of cellulolytic and fermentative bacteria would strongly point to the potential for changes in cycling of both N and C in A. arenaria–invaded dunes.
Changes in soil chemistry—and microbial communities—with invasion are concerning, because many of these invasion-associated soil changes do not dissipate with restoration, creating “legacy effects” that ultimately may hamper, or even preclude, the success of restoration efforts (Corbin and D’Antonio Reference Corbin and D’Antonio2011; Reinhart and Callaway Reference Reinhart and Callaway2006; Yelenik and D’Antonio Reference Yelenik and D’Antonio2013). Nutrient concentrations and native plant cover at intermediate (13 mo) and long-term (5 yr) post-restoration intervals in New Zealand and California sites where L. arboreus was removed largely returned to levels similar to or even slightly higher, respectively, than those of untreated areas (Hetherington and Wilson Reference Hetherington and Wilson2019; Maron and Jefferies Reference Maron and Jefferies2001). A number of C. edulis removal studies have found that, while some soil chemical legacy effects reverted within a year to pre-restoration conditions (salinity, Na+, Mg2+), others did not (pH, OM, NH4+, NO3−/NO2−), and other variables were potentially intermediate between uninvaded and invaded conditions (P, soil moisture; D’Antonio and Mahall, unpublished data in Conser and Connor Reference Conser and Connor2009; Novoa et al. Reference Novoa, González, Moravcová and Pyšek2013; Santoro et al. Reference Santoro, Jucker, Carranza and Acosta2011, Reference Santoro, Jucker, Carboni and Acosta2012). These chemical legacy effects promote new invasion by ruderal nitrophilous species within C. edulis removal areas (Novoa et al. Reference Novoa, González, Moravcová and Pyšek2013; Santoro et al. Reference Santoro, Jucker, Carranza and Acosta2011, Reference Santoro, Jucker, Carboni and Acosta2012), which reduces establishment of native dune species and overall species diversity (Badalamenti et al. Reference Badalamenti, Gristino, Laudicina, Novara, Pasta and La Mantia2016; Novoa et al. Reference Novoa, González, Moravcová and Pyšek2013).
Starting in 2001, Point Reyes National Seashore (PRNS), a unit of the U.S. National Park Service (NPS) located along the northern California coast, embarked on an extensive program of coastal dune restoration by removing A. arenaria and C. edulis. These species were likely planted in the early to mid-1900s by ranchers and other residents trying to prevent coastal dune sands from migrating into adjacent pastures, ranch facilities, and roads. By 2009, more than 60% of the park’s roughly 890 ha of coastal dune, bluff, and scrub habitat was estimated to be dominated by A. arenaria and C. edulis (NPS 2009). Between 2001 and 2018, approximately 110 ha of primarily A. arenaria and, to a much lesser degree, C. edulis had been removed from 200 ha of coastal dunes using a combination of manual removal (i.e., hand pulling or small hand tools), mechanical removal (i.e., using heavy equipment), and chemical treatment.
These restoration efforts have met with mixed success, with mechanical removal areas remaining largely unvegetated for many years except for recruitment by a very rare plant species (Tidestrom’s lupine [Lupinus tidestromii Greene]) and a disturbance-adapted weed (European searocket [Cakile maritima Scop.]). Herbicide treatment areas in the backdunes, which are older dune features further from the beach that typically support later successional vegetation communities, have largely been dominated by persistent standing dead A. arenaria and its thick litter (“thatch”) for 5 to 8 yr posttreatment, essentially creating a physical legacy effect (Parsons et al. Reference Parsons, Sayre, Ender, Rodrigues and Barberán2020b). With the exception of herbicide-treated foredunes (younger dune features directly adjacent to the beach), both types of restoration approaches at least initially failed to promote extensive or diverse recruitment by native dune plant species despite an abundant seedbank and were instead subject to invasion by new weeds or “secondary” invaders (Parsons et al. Reference Parsons, Ender and Ryan2020a). In recent years, colonization by natives within mechanical removal areas has improved substantially, but issues with native recruitment and secondary invader establishment persist within herbicide-treated backdunes (Parsons et al. Reference Parsons, Ender and Ryan2020a). As cover of primary invaders (A. arenaria and C. edulis) within restored areas remains low due to vigilant retreatment efforts, other factors appear to be constraining native dune habitat evolution and development. The recent microbial study at PRNS suggested that some of the changes in the microbial soil community brought about by invasion of A. arenaria, such as reduced abundances of nitrifiers, fermentative bacteria, fungal parasites, and fungal dung saprotrophs did not dissipate even 5 yr after restoration by herbicide treatment (Parsons et al. Reference Parsons, Sayre, Ender, Rodrigues and Barberán2020b).
We hypothesize that long-term colonization by dense stands of A. arenaria changes the chemistry of dune soils at PRNS relative to uninvaded areas (invasion effects) and that some of these changes may persist following invasives’ removal, creating legacy effects that may ultimately decelerate or even derail restoration efforts. In addition, physical legacy effects such as delayed decomposition of A. arenaria litter or thatch in herbicide-treated backdunes may at least partially stem from soil chemistry alterations. We further hypothesize that, even if chemical legacy effects do not persist, soils in restored areas may not necessarily revert to conditions more characteristic of comparable native dune habitats due to restoration itself having an effect that may differ depending on the approach used (treatment effect). The presence or intensity of invasion, legacy, and treatment effects may vary depending on secondary factors such as habitat successional stage; year or interannual variation in climatic conditions such as precipitation; distance from the ocean; or elevation. In this study, we conducted 2 yr of soil sampling within native dune (dune mat, dune scrub) and highly A. arenaria–invaded habitats, as well as former highly A. arenaria–invaded habitats that have been restored either mechanically or through herbicide treatment to evaluate OM, C, N, P, other macronutrients, pH, soil moisture, and exchangeable cations between habitats. We also assessed whether relationships existed between soil chemistry and vegetation-related measures of habitat evolution such as percent cover of native and nonnative plants, standing dead A. arenaria biomass, thatch, and bare ground.
Materials and Methods
Study Area
PRNS is located approximately 48 km north of San Francisco, CA, USA (Figure 1). Most of the lands at PRNS fall on a peninsula that juts out from California’s coastline into the Pacific Ocean and is perched along the western edge of the San Andreas Fault. Dunes at PRNS are largely composed of a modern largely continuous foredune ridge established by current and past A. arenaria–dominated vegetation and a complex series of undulating backdunes and depressions associated with an older, mostly stabilized parabolic dune field, including narrow deflation plains (slacks). In northern portions of a property formerly owned by the telecommunications company AT&T (AT&T North/South; Figure 1), climbing dunes overlay a high-elevation sandstone feature that reaches 61 m at its highest point.

Figure 1. Map of the study areas in Point Reyes National Seashore (PRNS), showing three sites (Abbotts Lagoon, AT&T North, and AT&T South) and location of PRNS along the northern California coast.
PRNS dunes represent a range of successional stages from early to midsuccessional communities such as foredunes and native dune mat, so-called yellow dunes, to later successional ones such as backdunes and native dune scrub. The latter two are often referred to as “gray” and “brown” dunes and are usually situated further inland (>300 m from the ocean) or, in the case of dune scrub, sometimes at higher elevations along the immediate coast (Parsons et al. Reference Parsons, Ender and Ryan2020a). Both uninvaded foredunes and dune mat are sparsely vegetated, largely open sand habitats supporting herbaceous communities, including those in the following alliances: Leymus mollis (Trin.) Pilg.; Abronia latifolia Eschsch.–Ambrosia chamissonis (Less.) Greene; and Eriophyllum stoechadifolium Lag.–Erigeron glaucus Ker Gawl.–Eriogonum latifolium Sm. (Buck-Diaz et al. Reference Buck-Diaz, Sikes and Evens2021). Dune scrub is a dense, shrub-dominated habitat represented by alliances such as Lupinus chamissonis Eschsch.–Ericameria ericoides (Less.) Jeps. (Buck-Diaz et al. Reference Buck-Diaz, Sikes and Evens2021). A range of invasion conditions exist within PRNS dunes from sparsely to highly invaded, with the highly clonal nature of A. arenaria and C. edulis favoring eventual establishment of dense monocultures exceeding 98% cover (Parsons et al. Reference Parsons, Ender and Ryan2020a). Some invaded systems retain vestiges of native dune mat or dune scrub that have either not been invaded or only very sparsely invaded by C. edulis and a few other nonnatives despite being, in some instances, almost completely surrounded by A. arenaria or C. edulis for decades (Parsons et al. Reference Parsons, Ender and Ryan2020a). The largest native dune mat habitat in PRNS is found at Abbotts Lagoon (Abbotts; ∼20.8 ha), although there are also smaller dune mat areas at AT&T (∼1 ha). Native dune scrub is prevalent at both Abbotts and AT&T, although the most intact stands in terms of low number and percent cover of nonnatives occur at AT&T North.
PRNS dunes provide habitat for 11 species listed by the U.S. Fish and Wildlife Service as endangered or threatened. Therefore, dune restoration efforts have focused on areas that should have the greatest benefit for critical rare species. To date, most of the park’s restoration efforts have focused on removing A. arenaria using a combination of manual removal, mechanical removal, and chemical treatment. Mechanical removal involves use of excavators and bulldozers to invert or “flip” the surface 1 to 1.5 m of rhizome-contaminated soils with less-contaminated soils 2 to 3 m below the soil surface. Initial chemical treatment is performed using herbicide backpacks with a single nozzle wand and directed spraying of a mixture of 1% imazapyr (Habitat®, isopropylamine (IPA), 27.77% ai, SePRO, Carmel, IN, USA); 2% glyphosate (Roundup Custom®, IPA, 53.8%, Monsanto, St Louis, MO, USA); 1.5% of a modified vegetable oil surfactant (Competitor®, 98.0%, Wilbur-Ellis, Fresno, CA, USA); and 1% blue dye (various manufacturers) onto target invasive plants. PRNS elected—and received approval—to use aquatic-label herbicides (e.g., Habitat®) to minimize toxicity within sensitive dune habitats, even though they are not aquatic.
Soil types in PRNS dunes consist of hummock mounds and hills of loose sand with no profile development (Soil Conservation Service 1985). For this reason, coastal dune sands tend to be strongly nutrient limited, especially for the essential nutrients N and P and, to a lesser extent, K+, while Ca2+ and Mg2+ levels are typically adequate for plant growth (Barbour et al. Reference Barbour, De Jong and Pavlik1985; Koske and Polson Reference Koske and Polson1984; Willis and Yemm Reference Willis and Yemm1961; Woodhouse and Hanes Reference Woodhouse and Hanes1967). Nutrient cycles in California dunes have not been studied except at Point Reyes (Holton et al. Reference Holton, Barbour and Martens1991) and Humboldt Bay (Nature Conservancy, unpublished data in Pickart and Sawyer Reference Pickart and Sawyer1998; Rose Reference Rose1988). Holton et al. (Reference Holton, Barbour and Martens1991) found that total N in earlier successional habitats (foredune, native dune mat) at Point Reyes averaged only 0.004%. In contrast, total N in later successional habitats such as dune scrub ranged from 0.5% to 6.0%, while OM remained low (0.004% to 0.2%; Holton and Johnson Reference Holton and Johnson1979).
The climate in Point Reyes is Mediterranean, with hot/dry summers and cool/wet winters. Coastal areas in California are generally cooler and have less intra-annual variability in temperature than inland regions, with fog, winds, and salt spray being important climatic influences. Annual temperature is estimated to average 12.1 C, and precipitation near the project area is estimated to average 54 cm yr−1 (2006 to 2020; WRCC 2020). Total rainfall can vary dramatically between years, as is evident for total precipitation numbers for 2019 (89.5 cm) and 2020 (32 cm; WRCC 2020). Fog blankets the Point Reyes coast about 30% of the time between June and October (Goodman Reference Goodman1982 in Ingraham and Matthews Reference Ingraham and Matthews1995), generating anywhere from 1.0 to 5.2 L site−1 yr−1 of additional moisture during what is otherwise California’s driest period (June to October; Ingraham and Matthews Reference Ingraham and Matthews1995). Another climatic influence is windblown salt spray, which occurs during both winter and summer seasons (Clayton Reference Clayton1972). Salt spray studies conducted at Point Reyes estimated deposition values at coastal sites ranging from 3 to 18 mg (dm2)−1 d−1 (Barbour Reference Barbour1978; Holton and Johnson Reference Holton and Johnson1979).
Selection of Sites for Soil Sampling
Samples were collected at 51 sites in 2019 and 2020 at Abbotts Lagoon (38.110076°N, 122.959294°W) and AT&T (38.09643°N, 122.964371°W; Figure 1). The Abbotts area falls south of a large coastal lagoon, with AT&T directly bordering Abbotts to the south (Figure 1). To evaluate whether A. arenaria invasion changed soil chemistry relative to conditions in uninvaded dune habitat, we focused soil sampling in (1) uninvaded or sparsely invaded (<5% cover of invasives) native dune habitats (dune mat, dune scrub; Figure 2A and B) and (2) highly invaded A. arenaria (>90% cover) dunes (Figure 2C). We also assessed the potential for legacy or treatment effects by sampling formerly highly invaded A. arenaria dunes that had been restored either (3) mechanically (Figure 2D) or (4) using herbicide.

Figure 2. Representative photographs of sampled habitats, including (A) sparsely invaded native dune mat; (B) sparsely invaded native dune scrub; (C) highly invaded Ammophila arenaria monoculture; and (D) former highly invaded areas being restored mechanically.
We used a random stratified sampling approach to either select sampling locations from existing 2 by 1 m vegetation monitoring plots or to create plots in invaded areas, as well in native dune mat, dune scrub, and herbicide-treated foredunes. We generally interspersed sampling within habitats among sites (Abbotts, AT&T North, AT&T South); however, this was not always possible (Table 1). Chemical treatment of A. arenaria has been performed at all three sites, but mechanical removal was only conducted at Abbotts in 2011 due to the high restoration cost per acre and indirect impacts from subsequent sand remobilization on adjacent native habitats and ranchlands (Table 1). Subsequently, PRNS switched its restoration approach primarily to herbicide treatment, with treatment projects occurring in a spatially sequential manner starting in the north with Abbotts (2011 to 2015) and then moving south to AT&T North (2016) and AT&T South (2017 to 2018; Table 1; Figure 1). Most of the remaining heavily invaded A. arenaria areas primarily occur at the very southern portion of AT&T, south of current restoration efforts, although a few isolated stands of backdune A. arenaria–invaded areas do persist within restoration areas where nontreatment buffers to organic pastures were established (Table 1). Sampling of native dune mat and dune scrub was also more intensive at Abbotts and AT&T North, respectively, due to higher proportions of these habitats at those sites (Table 1).
Table 1. Number of soil samples analyzed from restored and unrestored dunes in Point Reyes National Seashore (PRNS). a
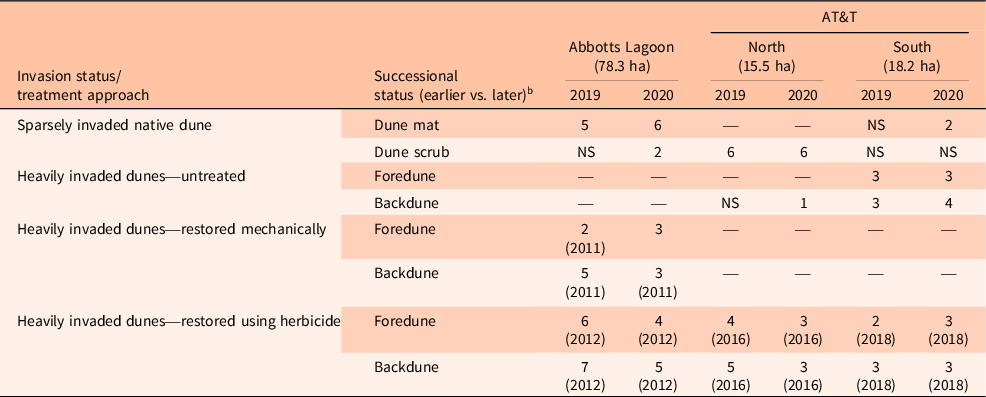
a For restored areas, initial treatment year is shown in parentheses below sample number. Dash connotes that specified habitat was not available at study site. NS, present, but not sampled.
b Successional status column shows sites in earlier successional habitats in darker shading and later successional habitats with lighter shading.
Within areas of varying invasion status (native, invaded, mechanically restored, herbicide treated), we also ensured approximately equal sampling effort between earlier successional habitats (native dune mat, foredunes) and later successional habitats (backdunes, native dune scrub), as soil texture and chemistry would be expected to differ based on successional stage (Holton and Johnson Reference Holton and Johnson1979), and successional status may influence invasion, legacy, and treatment effects (Table 1). In 2020, a few sampling sites were dropped and new ones added to improve interspersion of habitats among sites and uniformity of sampling effort among successional habitats (Table 1).
Soil Sampling Methods
We conducted soil sampling in late May (20, 22, 29 to 30) in 2019 and early June (1 to 3) in 2020, the peak growing season for most dune vegetation communities. Within plots, surface soils were collected using a 30-cm slotted mini-soil probe to a depth of 30 cm, with multiple cores taken using a cross-diagonal sampling pattern. Sampling location varied in proximity to plants, although sampling avoided collection of dense roots. We separated soils into several plastic bags marked for different analytical laboratories and transported samples in our backpacks in contractor bags with multiple ice packs until samples could be transferred to coolers in our vehicles.
Vegetation Sampling Methods
To evaluate whether soil chemistry appeared linked to changes in vegetation-related variables in herbicide-treated backdunes, we conducted vegetation monitoring using previously and newly established 2 by 1 m plots in 2020. We assessed cover of vegetation and other functional groups using a PVC quadrat separated into 10 by 10 cm grids for cover estimation by the point-intercept method, with a maximum of 171 sampling points. At each sampling point, plant species present (both alive and dead) were recorded once regardless of the number of “hits,” along with functional groups such as bare ground, thatch (intact dead stems >10 cm), and detritus (finer OM ≤10 cm). In addition to grouping species into native, nonnative, and primary and secondary invader species, we also grouped native dune species according to successional stage, as certain species appear strongly associated with different successional stages of dune habitat (early, mid, late, and dune scrub) related to their ability to tolerate disturbance from wind, wave, salt spray, and sand burial (Barbour et al. Reference Barbour, De Jong and Johnson1976; Hesp Reference Hesp1991). Native and nonnative species richness was calculated from the total number of native and nonnative plant species present in each plot regardless of whether “hit” by intercept sampling or not.
Laboratory Analyses—Soils
We used commercial laboratories for soils analyses. In 2019, soils were only sent to Western A&L Agricultural Laboratory (Western A&L, Modesto, CA, USA). In 2020, soils were shipped to both Western A&L and University of California, Davis, Analytical Laboratory (UC Davis, Davis, CA, USA). Soils for Western A&L were stored in coolers in a basement at PRNS until all sampling was completed during 2019 and 2020 and then shipped with ice packs. Samples for UC Davis were shipped priority overnight with ice packs after every sampling day for analysis of NH4+ (2.0 N KCL extract; ppm), NO3− (2.0 N KCL extract; ppm), total Kjeldahl nitrogen (TKN; wet method; %), PO4− (weak Bray; ppm), and total organic carbon (TOC; combustion; %).
The following additional analyses were performed using standard laboratory methods: OM (loss-on-ignition at 360 C; %); PO4− (Olsen; ppm); sulfate (SO42−; 1.0 N ammonium acetate at pH 7; ppm); K+ (saturated paste–inductively coupled plasma atomic emission spectrometry [ICP AES]; ppm); Mg2+ (saturated paste–ICP AES; ppm); Ca2+ (saturated paste–ICP AES; ppm); Na+ (saturated paste–ICP AES; ppm); pH (saturated paste); soil moisture ((wet wt.–dry wt./dry wt.); %); and soluble salts (saturated paste–conductivity; mmhos cm−1). Percent exchangeable (Exc) cations (K+, Mg2+, Ca2+, Na+, H+) and CEC (meq 100 g−1) were calculated using the procedure of 1.0 N ammonium acetate extraction at pH 7. Soluble salts were not assessed in 2020. In 2020, soil texture (Na-hexametaphosphate hydrometer; % sand/silt/clay) was assessed for 12 sampling sites. We calculated the C:N ratio of soils for June 2020 samples using the ratio of %TOC divided by %total N, which was calculated by adding %TKN to the very small percentage of inorganic N (NO3−, NH4+).
Statistical Analyses
Hypothesis Testing
Invasion effects were evaluated by comparing soil chemistry of sparsely invaded native dune mat or dune scrub sampling sites with that of highly invaded A. arenaria ones. For variables demonstrating an invasion effect, legacy effects were explored by comparing results of highly invaded A. arenaria sites with those of herbicide-treated and mechanically restored areas to determine whether restoration resulted in a change. As restoration may affect soil chemistry, even if invasion does not, variables without legacy effects that differed between restored and native dune areas following restoration were classified as treatment effects. Secondary factors that may indirectly influence invasion, legacy, and treatment effects were also evaluated. These included successional status (“earlier” successional or foredune/dune mat, “later” successional or backdune/dune scrub), year (2019, 2020), site (Abbotts, AT&T North, AT&T South), distance from the ocean, and elevation.
Permutational Multivariate Analysis Of Variance
We used a permutational multivariate analysis of variance (PERMANOVA; Oksanen et al. Reference Oksanen, Blanchet, Friendly, Kindt, Legendre, McGlinn, Minchin, O’Hara, Simpson, Solymos, Henry, Stevens, Szoecs and Wagner2019) to assess competing hypotheses explaining overall dune soil chemistry values. Sixteen candidate models consisting of additive and interaction terms of successional status, invasion status, year, site, distance from ocean, and elevation were ranked using a custom script to calculate Akaike’s information criterion (AIC) for small sample sizes. Based on our analyses, the best model incorporated a direct effect of year and an interaction between invasion status and successional status.
Principal Components Analysis
Before analysis was conducted, three sample sites were removed using Mahalanobis distance to detect outliers in multivariate space (distances > 6.58). We then conducted unconstrained ordination on scaled soil chemistry variables from 2019 and 2020 using principal components analysis (PCA) with rotated axes (Oksanen et al. Reference Oksanen, Blanchet, Friendly, Kindt, Legendre, McGlinn, Minchin, O’Hara, Simpson, Solymos, Henry, Stevens, Szoecs and Wagner2019). We linked the PCA to PERMANOVA results. Variables in PERMANOVA models with the lowest AIC were used to plot visual groupings on the PCA results. We used the scree plot technique to determine the number of principal components that should be retained for interpretation.
GLMM
We used separate generalized linear mixed models (GLMM; Bates et al. Reference Bates, Maechler, Bolker and Walker2015) to explain patterns for each individual soil chemistry variable that showed strong factor loadings in PCA. (Soil moisture and soluble salts were not analyzed.) Based on AIC rankings in PERMANOVA, models were built with an interaction between invasion status and successional status, a direct effect of year, and both fixed and random intercepts for site. We did not assess competing models for each variable; rather, we followed the form of the best (lowest AIC) model found for the PERMANOVA using all the soil chemistry values. Traditional transformations of dependent variable distributions were unsuccessful in providing reasonable GLMM residual plots, and we therefore normalized data via ordered quantile normalization (Peterson Reference Peterson2019). We corrected for multiple testing using the false discovery rate method (Benjamini and Hochberg Reference Benjamini and Hochberg1995).
Non-metric Multidimensional Scaling
To evaluate temporal changes in vegetation-related and soil chemistry variables in backdunes based on time since herbicide treatment, we conducted unconstrained ordination with non-metric multidimensional scaling (NMDS; Oksanen et al. Reference Oksanen, Blanchet, Friendly, Kindt, Legendre, McGlinn, Minchin, O’Hara, Simpson, Solymos, Henry, Stevens, Szoecs and Wagner2019) on data from backdune areas where herbicide was applied in 2012, 2016, or 2018. We then fit environmental vectors selected based on permutation tests onto the NMDS to determine soil chemistry variables that best explained changes in vegetation-related variables over time.
All analyses were done in R v. 4.0.2 (R Core Team 2020).
Results and Discussion
Invasion Status
Invasion by A. arenaria exerted a dramatic effect on soil chemistry relative to that of sparsely invaded native dune habitats, influencing 74% of 19 variables assessed, with the exception of C:N, NH4+, PO4−, Ca2+, and Exc-Na+. Table 2 presents results of statistical analyses, while Table 3 summarizes which variables showed invasion effects (i.e., which variables in invaded habitats were not statistically equivalent with those in native ones) and, for variables exhibiting invasion effects, whether means were generally higher or lower than those in native habitats (up/down arrows) and whether differences were restricted to just one successional stage (“early” or “late”). Other than NH4+ and PO4, almost every organic (OM, TKN, TOC) and inorganic (NO3−, SO42−, K+, Mg2+, Na+) source of macronutrients and exchangeable cations (H+, K+, Mg2+, Ca2+) appeared altered by invasion, along with pH and CEC (Tables 2 and 3). Invasion effects were more pronounced in certain successional habitats for half of the variables (Tables 2 and 3). This is evident in Figure 3, which graphically displays standardized means for earlier and later successional stages in both native and invaded habitats (as well as restored habitats). Only TKN, NH4+, C:N, PO4−, Exc-K+, and Exc-Ca2+ variables showed no interaction between invasion and successional status (Tables 2 and 3). Supplementary Table 1 provides summary statistics for all soil chemistry variables analyzed for different invasion and successional status habitats in 2019 and 2020.
Table 2. Statistical results for GLMM analyses. a

a Bold values represent significant t-test differences for slope estimates of variables relative to InvStatus [Native], SuccStatus [Early], Year [2019], and InvStatus [Native]*SuccStatus [Early]. Information on random effects related to site include variance (σ2), random intercept variance (τ00), and intraclass correlation coefficient (ICC). Marginal and conditional R2 of the model are also presented. We corrected for multiple testing using false discovery rate (FDR) method (Benjamini and Hochberg Reference Benjamini and Hochberg1995). Exc, exchangeable; OM, organic matter; TOC, total organic carbon; TKN, total Kjeldahl nitrogen.
*P < 0.05.
**P < 0.01.
***P < 0.001.
Table 3. Effects of invasion by Ammophila arenaria and habitat restoration on soil variables from 2019 to 2020. a

a Invasion effects are those in which there are significant differences between sparsely invaded native and highly invaded areas. Effects that persist following restoration of invaded areas are characterized as legacy effects. Variables without legacy effects that differ between restored and native areas following restoration are classified as treatment effects. Variables with significant statistical interaction between invasion and successional status are noted in the last column. Legacy and treatment effects are separated on the basis of restoration approach (mechanical removal, herbicide treatment). Arrows indicate direction of difference in specified habitat (invaded or restored) relative to native habitats (↑, higher; ↓, lower; ↕, both higher and lower depending on successional status). Italicized references to Early or Late refer to effects present only in specific type of successional status habitat (earlier, later). Exc, exchangeable; OM, organic matter; TOC, total organic carbon; TKN, total Kjeldahl nitrogen.

Figure 3. Individual GLMM effects plots for invasion status (x axis) by successional status (earlier/later) for selected soil chemistry variables. Plots present standardized means with 95% confidence intervals, with dashed line representing overall means for each variable standardized to 0.
Multivariate ordination results point to successional status factors perhaps being more critical to soil chemistry than invasion status. On the PCA’s first axis (PC1; 44.06% of variance), earlier successional habitats generally grouped on the left side of the biplot regardless of invasion or restoration status with a few exceptions (Figure 4). Mechanically restored backdune plots (later successional) overlapped strongly with earlier successional native dune mat plots, and invaded foredune plots fell in the center of the biplot, straddling earlier and later successional habitat groupings (Figure 4). Interpretation of PC2—which accounted for less than a third of the variance (16.92%) explained by PC1—was more complicated, but it largely represented the separation between restored and unrestored habitats, with sparsely invaded native dune mat (Figure 4, bottom left) grouping separately from mechanically restored and herbicide-treated foredunes (Figure 4, upper left). Sparsely invaded native dune scrub (Figure 4, upper right) grouped separately from invaded backdunes (Figure 4, bottom right), with herbicide-treated backdunes juxtaposed between them. Both PC2 and PC1 showed a statistically significant relationship between the interaction of invasion and successional status, as well as generally with invasion status, but only PC1 differed between sparsely and highly invaded habitats (Table 3).
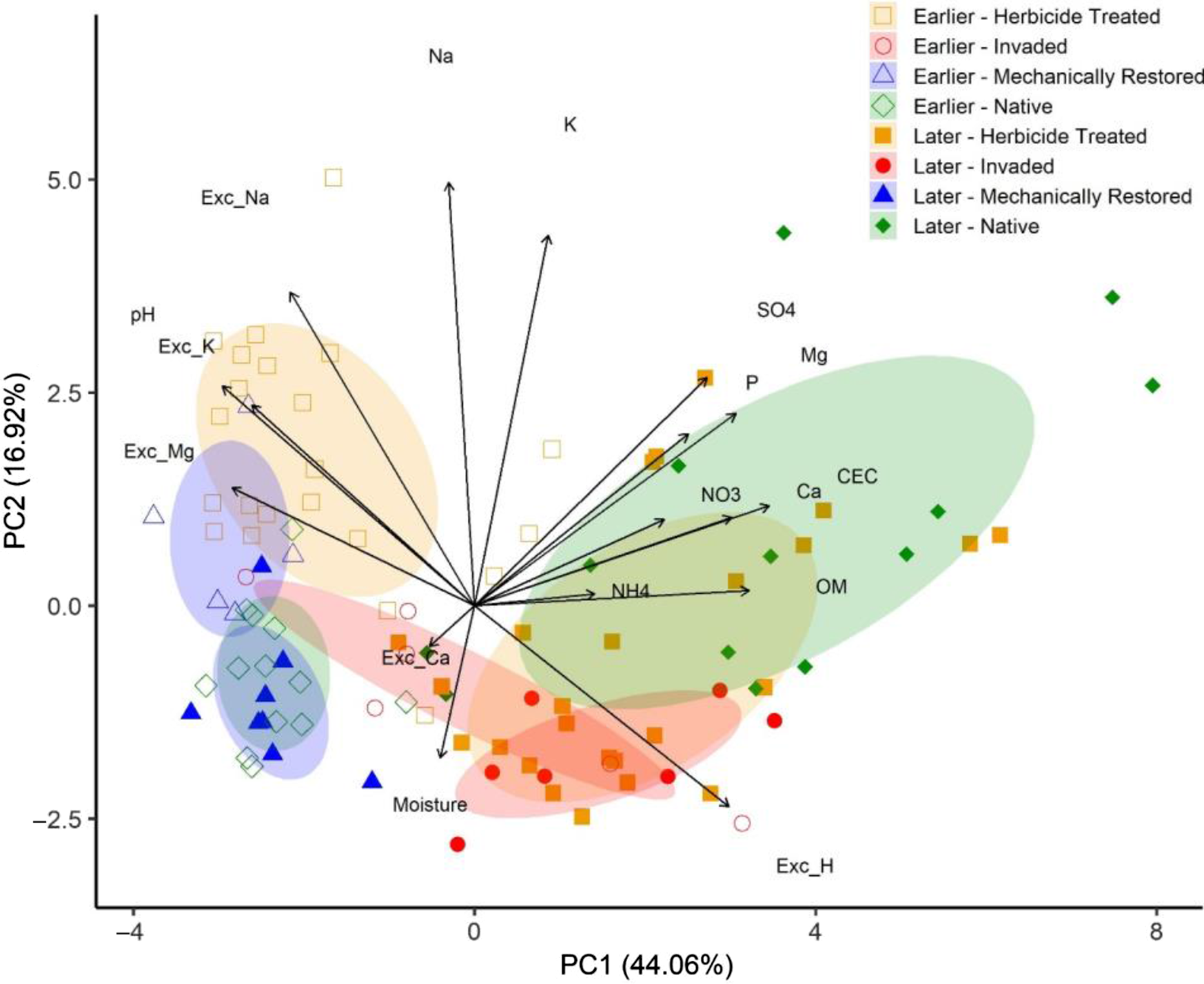
Figure 4. Principal components analysis (PCA) biplot depicting relationship of habitats of varying invasion status and successional stage and quantitative soil chemistry variables for 2019 and 2020. Scree plots indicate that two components captured most of the variance. The arrows point in the direction of maximum change of that variable, and their length is proportional to their correlation with the ordination axis and thus their relation to pattern of habitat variation. Ellipses represent 50% confidence intervals. Exc, exchangeable; OM, organic matter.
Macronutrient dynamics within coastal dunes are a balancing act between geologic and climatic forces. Unlike terrestrial ecosystems where weathering of soils and rocks generates some of the K+, Mg2+, Ca2+, and P needed by plants, sandy soils in dunes are already highly weathered and mostly unable to supply these macronutrients (Art et al. Reference Art, Bormann, Voigt and Woodwell1974; Etherington Reference Etherington1967; Smith et al. Reference Smith, Allen and Daly1985). Most inputs of N, oxygen (O), and SO42−, as well as base cations such as K+, Mg2+, Ca2+, and Na+, into coastal dunes come from meteorological sources such as rain, salt spray, and fog (Clayton Reference Clayton1972; Etherington Reference Etherington1967; Holton et al. Reference Holton, Barbour and Martens1991; Jacob et al. Reference Jacob, Waldman, Monger and Hoffmann1985; van der Valk Reference van der Valk1974). Due to the very low abundance of clay, sandy soils also do not have a strong capacity for binding cations that enter the system from elsewhere and are prone to considerable leaching (Art et al. Reference Art, Bormann, Voigt and Woodwell1974; van der Valk Reference van der Valk1974). OM provides virtually all the exchange surface capable of binding cations (Art et al. Reference Art, Bormann, Voigt and Woodwell1974; Etherington Reference Etherington1967), and OM adsorption is strongly dependent on pH, with CEC typically lower at lower pH values (Smith et al. Reference Smith, Allen and Daly1985).
Our results corroborate the well-documented pH gradient between early and late successional coastal dune habitats (Figure 3A), with lower pH values (<5.0 to 6.5) prevalent in late successional or “gray/brown” dunes (except for those deriving from calcareous geologic substrates such as limestone) and higher pH values (>6.5) common in early successional “yellow” dunes (Espejel Reference Espejel1992; Kooijman et al. Reference Kooijman and Morriën2020). Higher alkalinity of earlier successional soils may be reinforced by higher levels of calcium carbonate, which is often leached from later successional soils (Randall Reference Randall1973; Smith et al. Reference Smith, Allen and Daly1985), as well as higher cation influx due to proximity to the ocean. Within our study area, pH values ranged from 6.24 to 7.27 in native dune mat and mechanically restored foredunes (“yellow” dunes) to 5.10 to 5.90 in backdunes and native dune scrub (“gray” dunes; Figure 3A). The percentage of OM increased with successional age consistent with the fact that percent vegetation and shrub cover is higher in native dune scrub than in foredunes or native dune mat (Barbour et al. Reference Barbour, De Jong and Pavlik1985; Kooijman et al. Reference Kooijman and Morriën2020; Figure 3B). Higher OM concentrations in later successional soils were associated with higher levels of organic nutrients such as TOC and TKN compared with those in earlier successional ones (Figure 3C and D). Interestingly, CEC, which is typically reduced in soils with lower pH, was much higher in later successional habitats with lower pH than that of earlier successional ones with higher pH (Figure 3L).
OM and TOC levels in highly invaded soils exceeded those in native dunes, but differences are particularly distinct between invaded foredunes and native dune mat (Table 3; Figure 3B and C). These differential invasion effects on foredunes may account for invaded foredunes being the only earlier successional habitat that did not group clearly on the left of PC1, but rather was shifted toward later successional habitats on the right (Figure 4), suggesting that invasion successionally ages foredunes. Accumulation of high amounts of OM increases CEC in highly invaded foredunes relative to that present in sparsely invaded ones (Figure 3L). However, OM decomposition generates acidic H+ cations in both invaded foredunes and backdunes seemingly in excess of atmospheric contributions of base cations from fog, rainfall, and salt spray, leading to reduced pH, higher levels of acidic cations (Exc-H+), and lower levels of exchangeable base cations such as K+, Mg2+, and Ca2+ (Table 3; Figure 3M–P). At lower pH values, acidic cations (H+) preferentially bind to OM, displacing base cations such as K+, Mg2+, and Na+ (Brady and Weil Reference Brady and Weil1999; Figure 3M–O). Exchangeable base cations start becoming more important at a pH of 5 and increase considerably at pH 6, at which point Exc-H+ (and Exc-Al3+) concentrations taper off considerably (Brady and Weil Reference Brady and Weil1999). Of the exchangeable base cations, only Exc-Na+ did not appear affected by invasion (Tables 2 and 3).
While pH was moderately acidic in both invaded and native later successional habitats, NO3− levels remained much lower in the former than the latter (Tables 2 and 3; Figure 3F). Recent microbial studies at PRNS documented that invaded dunes supported significantly fewer nitrifying bacteria than sparsely invaded ones (Parsons et al. Reference Parsons, Sayre, Ender, Rodrigues and Barberán2020b). Higher concentrations of NO3− in acidic later successional habitats (1.47 to 5.53 ppm) relative to circumneutral earlier successional ones (0.39 to 2.77 ppm) suggests that lower pH values are not constraining nitrification: even in low pH situations, nitrification can still proceed if enough Ca2+ and Mg2+ are present (Brady and Weil Reference Brady and Weil1999). Anions such as NO3− and SO42− would be expected to quickly leach out of sandy systems, as they do not bind to OM, but primarily to clay particles and oxides (Brady and Weil Reference Brady and Weil1999). Dune soils at PRNS were uniformly low in clay (2%) and silt (3%; n = 12). Sulfate concentrations were higher in invaded foredunes than native earlier successional habitats, which may result from a combination of higher rates of production from OM decomposition and potentially lower rates of leaching than in other less densely vegetated sandy soils (Tables 2 and 3; Figure 3H).
P is typically a major limiting nutrient in dune and other terrestrial ecosystems, but as noted earlier, invasion did not appear to affect inorganic PO4−, despite it affecting related factors such as OM. While other studies have shown lower-pH dune habitats to have reduced concentrations of inorganic PO4− relative to higher-pH habitats (Kooijman et al. Reference Kooijman and Morriën2020), inorganic PO4− levels in later successional habitats at PRNS were substantially higher (7.3 to 25.7 ppm) than in circumneutral earlier successional ones (2.1 to 9.4 ppm; Figure 3G). In northern Europe, inorganic phosphate levels were lower in moderately acidic dunes than in higher pH calcareous ones, but levels of phosphates more weakly sorbed to OM and organic P—rather than to ferric oxides—increased, which rendered more PO4− available for plant uptake in lower-pH dunes (Kooijman et al. Reference Kooijman and Morriën2020).
While Exc-Mg2+ was lower in invaded soils, Mg2+ in soil solution was higher, at least in invaded foredunes (Tables 2 and 3; Figure 3J). As noted earlier, cations such as Mg2+ are probably largely supplied by meteorological sources. While appreciable differences in nutrient deposition would not be expected between early and late successional habitats with respect to fog or rainfall, a Point Reyes study found that sites 500 m inland received 20% to 50% of the salt spray volume received by seaward cliff-edge sites (Barbour Reference Barbour1978; Holton and Johnson Reference Holton and Johnson1979). Deposition rates of Ca2+, Na+, and Mg2+ from winter salt spray in central California coastal areas exceeded those of inland sites (2.2 km from coast) by as much as a factor of 4, while K+ influx was roughly equivalent (Clayton Reference Clayton1972). Large amounts of these cations are taken up by plants (van der Valk Reference van der Valk1974), with plant tissue concentrations roughly similar between inland and coastal sites despite differential deposition rates (Clayton Reference Clayton1972). Ionic patterns within soils reflect steady-state cycling of K+, Mg2+, and Ca2+ in California coastal systems, with root exudation and litterfall release increasing concentrations in surface soils, while sandy soils reduce concentrations at greater depths (1 to 2 m) through leaching (Clayton Reference Clayton1972). While deposition rates of cations may be higher in earlier successional habitats, denser, taller vegetation in later successional areas may better trap cations from meteorological inputs through foliar absorption and eventual turnover through root exudation or litterfall release, leading to higher levels of cations in these soils. Later successional native dunes in PRNS supported higher concentrations of K+, Ca2+, Mg2+, and Na+ in soils (Figure 3I–K).
Invaded foredunes may have deviated from this successional trend of higher base cations in later successional soils, because dense monocultures of A. arenaria in invaded foredunes enhance the potential for canopy interception, foliar absorption, and translocation of Mg2+ into roots or litter and eventually into soils. Soil Mg2+ levels in native dune scrub, which supports relatively dense stands of intermixed shrubs and forbs, appeared comparable to those of invaded backdunes (Figure 3J). Similar trends might have been expected with K+, Ca2+, and Na+, but instead showed slightly different patterns. Invaded foredunes did have higher Na+—if not K+—concentrations than native dune mat, but invaded backdunes had much lower Na+ and K+ levels than native dune scrub (Tables 2 and ; Figure 3I–K). Ca2+ showed no effect with regard to invasion status (Tables 2 and 3).
Legacy Effects
Legacy effects were interpreted as invasion effects that did not dissipate following restoration, with soil conditions remaining comparable to those in earlier or later successional invaded habitats or both (Table 3). Restoration of formerly highly invaded A. arenaria habitats through either mechanical removal or herbicide treatment appeared to reverse only a few invasion effects. Of the 14 variables affected by A. arenaria invasion, roughly 60% (8) exhibited some type of legacy effect, with persistent differences in pH, CEC, NO3−, SO42−, Mg2+, Na+, Exc-H+, and Exc-Mg2+ (Tables 2 and 3). In half of the cases, differences persisted in only one type of successional habitat, even if conditions under invaded scenarios differed from native habitats for both types (Table 3).
Herbicide-treated areas were subject to 50% more chemical legacy effects than mechanical restoration ones, specifically for pH, SO42−, Mg2+, Na+, CEC, and Exc-H+, with effects split equally between both types of successional habitats or just later successional ones (Table 3; Figure 3). Legacy effects in soil chemistry following restoration are likely to be highly influenced by other invasion legacies, namely physical ones such as delayed decomposition of A. arenaria in herbicide-treated backdunes (Parsons et al. Reference Parsons, Sayre, Ender, Rodrigues and Barberán2020b). At PRNS, approximately 50% of herbicide-treated A. arenaria in backdunes remained as undecomposed standing dead biomass 2 to 3 yr posttreatment (Figure 5A), with standing dead cover in the oldest site, Abbotts, not dropping to single-digit percentages until 6 yr after restoration (Parsons et al. Reference Parsons, Ender and Ryan2020a; Figure 5B). Litter cover also stayed elevated after 6 yr (70%; Parsons et al. Reference Parsons, Ender and Ryan2020a; Figure 5C), which may limit plant establishment by suppressing germination (Xiong and Nilsson Reference Xiong and Nilsson1999).

Figure 5. Representative photographs of (A) standing dead Ammophila arenaria biomass in 3-yr-old plot at AT&T South with sparse native colonization and A. arenaria regrowth; (B) remaining standing dead A. arenaria biomass in 7-yr-old herbicide-treated plot at Abbotts with increased colonization by native dune plants and secondary invaders such as nonnative annual grasses; and (C) A. arenaria “thatch” or large litter.
Some researchers have postulated that invasives can exert effects on soil chemistry by decomposing very quickly, which, for N-rich weeds, can create a pulse of inorganic N that attracts persistent nitrophilous weeds (Liao et al. Reference Liao, Peng, Luo, Zhou, Wu, Fang, Chen and Li2008) such as those observed in the herbicide-treated backdunes (e.g., B. diandrus). Others have countered that decomposition rates of invasives are not necessarily faster than those of natives or, even if they are, do not necessarily influence nutrient cycling (Ehrenfeld Reference Ehrenfeld2003; Jo et al. Reference Jo, Fridley and Frank2016; Kueffer et al. Reference Kueffer, Klingler, Zirfass, Schumacher, Edwards and Güsewell2008; Prescott and Zukswert Reference Prescott and Zukswert2016). In some instances, decomposition of invasives’ litter may actually proceed more slowly than that of natives, particularly when lignin content of invasives is comparatively higher (Godoy et al. Reference Godoy, Castro-Díez, Van Logtestijn, Cornelissen and Valladares2010; Windham Reference Windham2001), with delayed decomposition often leading initially to immobilization and depletion of N from soils by microbes.
While no published information could be found on lignin content or other plant chemistry characteristics of A. arenaria, preliminary data from plant material collected in PRNS suggest that these factors cannot entirely account for the substantial decomposition delays (Parsons et al. Reference Parsons, Ender and Ryan2020a). With the exception of C:N (70.8:1), values for cellulose, hemicellulose, lignin, total phenols, and silicon fell at the lower or, at most, intermediate range of those reported for other grass species (Parsons et al. Reference Parsons, Ender and Ryan2020a). Annual grasses occurring in coastal prairie at Bodega, directly north of Point Reyes, had C:N ratios closer to 47.36:1, very similar to L. arboreus (47.74:1), with highest N mineralization occurring in Year 1 for grasses and Year 3 for L. arboreus, probably due to the higher lignin content of L. arboreus (14.7%; Bastow et al. Reference Bastow, Preisser and Strong2008). A study on U.S. East Coast marshes found that both standing dead culms and litter of invasive common reed [Phragmites australis (Cav.) Trin. ex Steud.], which had a C:N ratio of 63:1, decomposed extremely slowly, with senescent material projected to take 9 yr to reach 10% of its original dry weight (Windham Reference Windham2001). Based on our plant chemistry data, litter processing rates would be expected to decelerate, as microbes would need to scavenge N from soils to bring C:N ratios of A. arenaria—which otherwise contains low amounts of recalcitrant compounds—down to ratios more favorable for bacterial activity. Given that the N content of coastal dune soils is naturally low, at least moderate delays in decomposition should be anticipated due to N limitation, as has been documented in other nutrient-poor ecosystems (Godoy et al. Reference Godoy, Castro-Díez, Van Logtestijn, Cornelissen and Valladares2010; Kueffer et al. Reference Kueffer, Klingler, Zirfass, Schumacher, Edwards and Güsewell2008). Soil C:N ranged from 15.8 to 17.8 in herbicide-treated backdunes (Supplementary Table 1) compared with medians of 12:1 in most arable soils (Brady and Weil Reference Brady and Weil1999), which supports the premise of reduced N and slower breakdown and accumulation of OM.
Soil chemistry—and physical—legacy effects appeared to be reversed much more quickly in herbicide-treated foredunes, probably due to frequent deposition of sand washed up onto adjacent beaches (Parsons et al. Reference Parsons, Ender and Ryan2020a). Sand influxes thickly blanketed A. arenaria necromass within 4 yr (<10% standing dead cover, 18% litter; Parsons et al. Reference Parsons, Ender and Ryan2020a), creating a new soil substrate largely freed from invasion effects (Table 3). Slow decomposition of buried A. arenaria litter may have continued, however, to fuel elevated SO42− and Mg2+ concentrations in herbicide-treated foredunes and to maintain higher OM in soils that contributed to elevated CEC more consistent with invaded foredunes (Figure 3B, H, J, and L).
Without countering influences from frequent sand deposition, standing dead A. arenaria biomass and thatch in herbicide-treated backdunes are considerably more persistent, and slowly decomposing litter may have a more pervasive influence on soil chemistry. Gradual breakdown of persistent OM in herbicide-treated backdunes may continue to generate H+ in excess of what could be countered by base ion influx, leading to higher levels of Exc-H+ and continued reductions in pH (Table 3; Figure 3A and M). However, decomposition in herbicide-treated backdunes seemingly did not much alter N levels (TKN, NH4+) post-restoration (Table 3; Figure 3D and E). While recent microbial research at PRNS suggested that abundance of nitrifying bacteria remained low after herbicide treatment (Parsons et al. Reference Parsons, Sayre, Ender, Rodrigues and Barberán2020b), NO3− concentrations actually increased in treated backdunes relative to invaded backdunes, becoming more comparable with levels in native dune scrub (Table 3; Figure 3F).
Based on NMS multivariate ordination of vegetation-related variables in herbicide-treated backdune plots, strong overlap existed between different treatment years (i.e., 2012, 2016, 2018), but plots with higher standing dead A. arenaria biomass and thatch cover characterized more recently treated plots (2018, some 2016; Figure 6). Standing dead biomass/thatch cover dropped in the oldest treated plots (2012), which were distinguished by higher cover of native species, including those representative of native dune scrub communities, as well as cover of nonnative and secondary invader species (Figure 6). Plots of intermediate treatment age had higher diversity of native and nonnative plant species, but lower total cover and possibly higher levels of detritus than older plots (Figure 6). Statistically significant higher levels of soil OM and N (TKN, NH4+) occurred in older plots, as shown in Figure 6 (permutation, P < 0.05), which may reflect eventual physical breakdown of standing biomass into surface litter and then into soil OM. It may also capture a shift in N cycling from immobilization—and possible depletion of N in more recently treated plots—to mineralization and increases in N in older treatment plot soils, as the C:N ratio of remaining litter slowly drops over time, and litter N is mobilized.

Figure 6. Non-metric multidimensional scaling (NMDS) biplot depicting relationship of vegetation-related community variables in herbicide-treated backdune areas of varying treatment age (2012, 2016, 2018) and statistically significant soil chemistry variables in 2019 and 2020. Statistically significant soil chemistry variables were identified through permutation tests. Ellipses represent 80% confidence intervals
Fewer chemical (n = 3) and physical legacy effects persisted following mechanical removal (Table 3). By flipping surface soils with subsurface soils, mechanical removal in the foredunes exposed subsoils that may have been higher in SO42− due to past leaching (Figure 3H). Inversion of topsoil and subsoil layers may also account for continued depletion relative to native habitats of NO3− in mechanically restored backdunes and of Exc-Mg2+ in mechanically restored foredunes (Figure 3F and O), possibly because most cations such as Mg2+ are leached out of subsoil layers, unlike Na+ (Clayton Reference Clayton1972; Etherington Reference Etherington1967). As Figure 4 indicates, with the exception of Ca2+, exchangeable cations such as Mg2+, K+, and Na+ were generally higher in earlier successional habitats at PRNS despite the highest soil solution concentrations being in later successional soils (Figure 3N and O).
Treatment Effects
Ultimately, the goal of restoration is to shift systems altered by invasion toward conditions—and processes—that more closely mimic those of target native habitats. This goal can certainly be derailed by lingering legacy effects of invasion, but restoration can change certain aspects of soil chemistry even if invasion did not. These treatment effects may, or may not, bring restored systems closer to re-creating desired conditions. At PRNS, native dune mat and dune scrub generally represent the target habitats for earlier and later successional areas, respectively.
Based on GLMM analyses (Table 3), both mechanically restored and herbicide-treated areas did not have soil chemistry similar to native habitats, regardless of time since restoration. Of the 19 soil chemistry variables assessed, 79% (n = 15) showed some evidence of a treatment effect in which soil conditions within restored areas not impacted by legacy effects were still dissimilar to those in native habitats (Table 3). While herbicide treatment areas have had to contend more with legacy effects, particularly in backdunes, mechanical restoration through “horizon flipping” has—not surprisingly—upended soil chemistry evolutionary trajectories in foredunes and even more so in backdunes relative to those of target successional communities. Almost 87% of treatment effects occurred in mechanical restoration areas. Our PCA ordination points to mechanically restored backdunes converging more with native dune mat than dune scrub, with PC1 and PC2 axes both showing strong overlap between these groups (Figure 4). Mechanically restored foredunes, meanwhile, are slightly offset from these two groups, positioning closer to herbicide-treated foredunes along PC2 (Figure 4). Mechanical removal essentially reset the successional clock, at least in the backdunes. It is possible that, over time, mechanically restored backdunes will evolve from a soil chemistry perspective, incrementally moving closer to conditions in native dune scrub communities, but the data are certainly not suggestive of any convergence at 8 to 9 yr post-restoration. In comparison, in herbicide-treated dunes, only 32% of variables evinced evidence of treatment effects, with most of these concentrated in foredunes (Table 3; Figure 3). Herbicide-treated foredunes had higher pH, NO3−, K+, Na+, and Exc-Na+ than native dune mat, while herbicide-treated backdunes had lower K+ and Ca2+ than native dune scrub (Table 3; Figure 3).
Year
Sampling year was identified as another important factor in the GLMM analysis, although it did not show any interaction with invasion or successional status. A number of variables showed a statistically significant relationship with sampling year, including OM, N (TKN, NH4+, NO3−), PO4−, K+, Mg2+, Na+, and some exchangeable cations (K+, Na+; Table 2). In general, most variables exhibited higher concentrations in 2019 (OM, N, PO4−, K+, Mg2+, and Exc-K+), while only Na+ and Exc-Na+ displayed higher levels in 2020.
The years 2019 and 2020 bracketed two extremes in California climatic variability, with 2019 having considerably above-average rainfall and 2020 having considerably below-average rainfall. Levels of organic and inorganic N and inorganic P (PO4−) seemingly flourished under the wetter climatic regime. Earlier research at PRNS indicated that rainfall could funnel as much as 1.6 kg N ha−1 yr−1 into coastal dune systems, with NH4+ contributions slightly higher (0.9 kg ha−1) than NO3− (0.7 kg ha−1; Holton et al. Reference Holton, Barbour and Martens1991). In addition to higher rainfall supplying more nutrients, higher soil moisture during wet years may also accelerate breakdown of OM, increasing organic N (TKN) and NH4+ and NO3− in years such as 2019. As discussed earlier, turnover of OM may play one of the largest roles in N and P cycling within coastal dunes. Holton et. al. (Reference Holton, Barbour and Martens1991) estimated that, of the approximately 390 kg ha−1 total N budgeted for PRNS dunes, 78% of that was organic N in soils, with another 18% tied up in plant biomass, and the remaining 4% being inorganic.
Higher litter decomposition rates in wet years may also trigger pulses of K+ and Mg2+ into soils, as K+ concentrations within coyote brush (Baccharis pilularis DC.) sampled at both coastal and inland sites in central California were among the highest of any of cations, with Mg2+ third highest behind Ca2+ (Clayton Reference Clayton1972). During wet years, higher levels of K+ and Mg2+ influx into soils from moisture-associated increases in decomposition would also be subsidized by increased deposition from rainfall and a higher number and intensity of storm-driven salt spray events (Clayton Reference Clayton1972; van der Valk Reference van der Valk1974). While plant uptake of Na+ is reduced compared with other cations, Clayton (Reference Clayton1972) suggested that, in coastal California soils, it is less subject to leaching, at least at deeper depths. Less rainfall in 2020 may have resulted in less translocation of Na+ to lower soil depths in PRNS dunes, while salt spray and fog deposition continued to maintain atmospheric inputs to the system.
Summer fog drip might be perceived as a boon for California’s coastal systems during low-rainfall years such as 2020. At PRNS, N input from fog was almost 3 times higher (4.2 kg N ha−1 yr−1) than that from rainfall (Holton et al. Reference Holton, Barbour and Martens1991). In contrast to trends for rainfall, concentrations of NO3− were much higher in fog than NH4+ (Holton et al. Reference Holton, Barbour and Martens1991; Jacob et al. Reference Jacob, Waldman, Monger and Hoffmann1985), resulting in contribution of approximately 1.4 kg ha−1 NO3− and 1.1 kg ha−1 NH4+ during the summer sampling period (Holton et al. Reference Holton, Barbour and Martens1991).
Another potential non–precipitation-related atmospheric source of N to coastal systems could be N fixation, which has been documented as a bigger contributor to N budgets than atmospheric deposition in Welsh dunes (Jones et al. Reference Jones, Sowerby, Williams and Jones2008). N-fixing taxa in coastal dunes include not only nodulated legumes, but cyanobacteria in biological soil crusts (Jones et al. Reference Jones, Sowerby, Williams and Jones2008) and A. arenaria and other dune grasses (L. mollis) that support N-fixing bacteria that are either endophytic (Oregon; Dalton et al. Reference Dalton, Kramer, Azios, Fusaro, Cahill and Kennedy2004) or free-living in the rhizosphere (e.g., Wales and northwest Europe; Abdel Wahab and Wareing Reference Abdel Wahab and Wareing1980; Ruppel Reference Ruppel1989). Based on Holton’s work, N fixation contributed negligible N to PRNS dunes due to the relatively sparse abundance of N-fixing species such as lupines (Lupinus spp.) and beach pea [Lathyrus littoralis (Nutt.) Endl.], and there was no evidence of either N-fixing algae or endophytic or free-living bacteria in soil and roots of A. arenaria at PRNS (Holton et al. Reference Holton, Barbour and Martens1991). In systems such as those in Europe and Oregon, where A. arenaria is dominant and known to support N-fixing bacteria, N fixation could potentially represent a much larger contributor to the N budget than it may at PRNS.
Given that coastal dunes are heavily reliant on meteorological contributions from rain, fog, and salt spray for nutrient budgets, this system would seem particularly vulnerable to potential climate change effects. While models for Mediterranean regions generally predict a drier and warmer climate in the future, some models suggest that certain portions of northern and central California could become wetter (Maloney et al. Reference Maloney, Camargo, Chang, Colle, Fu, Geil, Hu, Jiang, Johnson, Karnauskas, Kinter, Kirtman, Kumar, Langenbrunner and Lombardo2014; Polade et al. Reference Polade, Gershunov, Cayan, Dettinger and Pierce2017), with coastal areas turning windier, cooler, and either less or more foggy during warmer months (Lebassi et al. Reference Lebassi, González, Fabris, Maurer, Miller, Milesi, Switzer and Bornstein2009; O’Brien Reference O’Brien2011; Pozo Buil et al. Reference Pozo Buil, Jacox, Fiechter, Alexander, Bograd, Curchitser, Edwards, Rykaczewski and Stock2021; Snyder et al. Reference Snyder, Sloan, Diffenbaugh and Bell2003; Sydeman et al. Reference Sydeman, García-Reyes, Schoeman, Rykaczewski, Thompson, Black and Bograd2014). These disparities in modeling results create a high degree of uncertainty regarding future impacts of climate change on coastal dune systems. Regardless, some of these changes could alter and potentially exacerbate the impact that invasion of A arenaria, as well as C. edulis, have on soil chemistry.
In conclusion, invasion by A. arenaria strongly influences soil chemistry, altering conditions relative to native habitats for 74% (14) of the measured variables. Specifically, invasion appeared to successionally “age” invaded foredunes, edging them closer to later successional habitats in terms of chemistry. Almost 60% (8) of the invasion effects persisted following restoration, with legacy effects particularly prevalent in herbicide-treated backdunes where active sand deposition could not counter legacy effects of invasion as it did in herbicide-treated foredunes through burial of standing dead A. arenaria biomass, litter, and invasion-altered soils with new sand. Mechanical removal resulted in fewer legacy effects, but more treatment effects that left mechanically restored areas definitively dissimilar to native habitat counterparts. Only 5 of the 19 assessed variables in mechanically restored areas showed convergence with conditions in comparable native habitats. Removal of A. arenaria through inversion of surface and subsoil layers essentially restarted the successional timeline in mechanically restored backdunes, creating soil conditions more comparable with earlier successional native habitats. Soil chemistry appears to play only a minor role in delayed decomposition of A. arenaria, although litter breakdown may have been limited to some extent by low N in soils. Ultimately, N-mediated delays in microbial decomposition of the high C:N litter may have acted to ecologically “amortize,” or incrementally stagger over time, the considerable impacts to nutrient cycling that would have otherwise been expected from more immediate breakdown of such a large volume of invasive biomass.
Invasive plant removal alone may not be sufficient to restore native ecosystems due to lingering physical, chemical, and biological legacy effects inherited from invaders (Corbin and D’Antonio Reference Corbin and D’Antonio2012; Kardol and Wardle Reference Kardol and Wardle2010; Suding et al. Reference Suding, Gross and Houseman2004). As we and others have observed, the type of legacy—and treatment—effects may be strongly linked to eradication technique, with effects differing depending on the type of invaded habitat being restored (Hetherington and Wilson Reference Hetherington and Wilson2019). At PRNS, herbicide treatment has yielded so far the best results for foredunes, and mechanical removal, perhaps the best for backdunes, but cost and indirect impacts associated with mechanical removal make future use of this method infeasible. Regardless of technique, however, additional restoration measures—or “interventions that mitigate biogeochemical and physical effects” such as soil amelioration or removal of biomass (Kardol and Wardle Reference Kardol and Wardle2010; Suding et al. Reference Suding, Gross and Houseman2004; Wolfe and Klironomos Reference Wolfe and Klironomos2005)—may be required to successfully restore PRNS dunes, particularly herbicide-treated backdunes. Ultimately, the primary constraint on decomposition within backdunes at PRNS is probably microbial, not chemical. Previous microbial research at PRNS has shown that the abundance of cellulolytic bacteria—or microbes involved in breakdown of cellulose in plant litter—dropped dramatically following restoration using herbicide treatment (Parsons et al. Reference Parsons, Sayre, Ender, Rodrigues and Barberán2020b). Large reductions in celluloytic bacteria could stymie microbial efforts to process the extremely large volume of litter in treated A. arenaria monocultures, thereby helping to foster this physical legacy effect.
Future studies will attempt to further elucidate disconnects in the microbial decomposition process in herbicide-treated backdunes that may be hindering efforts to restore coastal dunes at PRNS. This information would then be used to design an adaptive restoration project. Decomposition could potentially be accelerated through introduction of whole-soil inoculum from native communities with higher expected decomposer abundance (e.g., dune scrub; Wolfe and Klironomos Reference Wolfe and Klironomos2005.) Alternatively, demand on decomposers could be reduced through pre- or post-restoration controlled burns. Until restoration approaches can be developed that integrate methods for resolving legacy effects, PRNS might best be advised to focus efforts on areas in which restoration yields more immediate benefits, such as herbicide treatment of foredune areas, where frequent sand deposition from adjacent beaches can almost instantaneously bury legacy effects. In the interim, vigilant, long-term retreatment of both primary and secondary invaders will be critical to restoration success.
Acknowledgments
We thank Michael Spaeth, Savannah Fuqua, and Jacob Longmeyer for assistance with field sampling. We thank P. Baye for information on dune morphology and J. Callaway for information on soils and for reviewing the draft article. Funding came from the U.S. National Park Service, as well as grants from nonprofit organizations (Point Reyes National Seashore Association, Point Reyes Station, CA, USA; Hanford Fund, Sonoma, CA, USA). Any use of trade names is for descriptive purposes only and does not imply endorsement by the U.S. Government. No conflicts of interest have been declared.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2021.16











