Management Implications
Ventenata dubia (ventenata) is a nonnative species that is quickly spreading into the Great Plains ecoregion. Along with it, negative impacts such as lower forage availability for cattle and wildlife and reduced biodiversity can be expected. These negative effects have been documented in the Great Basin and Pacific Northwest. This research was conducted to answer several questions surrounding conservation efficacy. We concluded that removal of V. dubia results in a positive response from perennial grasses and that this perennial forage base is of higher quality. Further, whereas annual grasses such as V. dubia grow quickly and senesce early in the growing season, perennial grasses continue growing later into the growing season before senescing. This provides a longer green grazable forage window, giving landowners and land managers greater control and reliability of their forage resources. Additionally, V. dubia is unpalatable to livestock and therefore has little forage value. Both studies presented here took place 1 yr after control of V. dubia. In that time, species richness and biodiversity did not change as a result of V. dubia removal. Whether this is due to failure of native species to reestablish or that they were simply not reduced in the first place is not clear. We also found that total biomass and nutrient mass did not increase, even with increases in more nutritious perennial grasses. This is likely due to replacement of annual grass biomass with an equal amount of perennial grass biomass because of limited resources. However, available forage and nutrient mass from palatable perennial grasses increased. Therefore, preventing invasion by V. dubia is preferable. With this in mind, removal of V. dubia resulted in many benefits to ecosystem goods and services and is worth pursuing where invasion occurs.
Introduction
Western North American rangelands have been invaded by nonnative species that have caused considerable damage to ecosystem services (DiTomaso et al. Reference DiTomaso, Monaco, James and Firn2017), such as providing forage for cattle and wildlife. Several of these invasive species are cool-season annual grasses such as the well-known European species cheatgrass [Bromus tectorum L.]. Other cool-season annual grass invaders like ventenata [Ventenata dubia (Leers) Coss.] and medusahead [Taeniatherum caput-medusae (L.) Nevski] have also become problematic in rangelands of the western United States, particularly in the Great Basin and grasslands of the Pacific Northwest (Hironaka Reference Hironaka1992; Scheinost et al. Reference Scheinost, Stannard and Prather2008a, Reference Scheinost, Stannard, Prather and Yenish2008b; Wallace et al. Reference Wallace, Pavek and Prather2015; Wallace and Prather Reference Wallace and Prather2016; Washington State Noxious Weed Control Board 2016; Young Reference Young1992). In high densities, these grasses tend to displace native species and desirable introduced grasses, due in part to their life histories (D’Antonio and Vitousek Reference D’Antonio and Vitousek1992). In some cases, substantive ecological changes occur, such as conversion to an annual grass–dominated system.
Ventenata dubia was introduced into North America no later than 1952 in Spokane County, WA (Barkworth et al. Reference Barkworth, Capels and Long1993). Since that time, it has spread to other western states and Canada (Fryer Reference Fryer2017). In the Pacific Northwest, V. dubia was spreading at an estimated rate of 1.2 million ha yr−1 in 2001 (Fryer Reference Fryer2017). Although the first documentation of V. dubia in Wyoming was in 1997, this documentation went unnoticed until V. dubia was confirmed in northeastern Wyoming with self-sustaining populations in 2016 before control efforts began (Garner and Lakes Reference Garner and Lakes2019). This recently invaded area is within the Great Plains ecoregion, where V. dubia threatens to spread rapidly as it has done farther west.
Winter annual grasses such as V. dubia usually germinate in the fall and take advantage of early spring moisture, although some annual grasses are able to produce seed early in the summer even if they fail to germinate until spring (Young Reference Young1992). This early use of spring moisture by annual grasses depletes soil moisture that would normally be available to desirable perennial grasses, causing water stress for perennials earlier in the summer than would be expected without invasive grasses (D’Antonio and Vitousek Reference D’Antonio and Vitousek1992; Evans et al. Reference Evans, Holbo, Eckert and Young1970). This depletion of resources, in addition to disturbances, such as fire, that reduce the abundance of perennial species (Anderson and Inouye Reference Anderson and Inouye2001; Melgoza et al. Reference Melgoza, Nowak and Tausch1990), is a possible mechanism that has allowed cool-season annual species to undermine the dominance of perennial grasses.
Reductions in perennial forage grasses cause other issues as well. Ventenata dubia has little value as a forage species early in the growing season (Pavek et al. Reference Pavek, Wallace and Prather2011). Any small value is short-lived, as V. dubia becomes unpalatable once it becomes dry (Fryer Reference Fryer2017; but see work by McCurdy et al. [Reference McCurdy, Watts, Chibisa, Prather and Laarman2017] on pelletizing V. dubia). Although this invasive annual grass has been present in North America for nearly 70 yr, it has only recently been documented in the Great Plains, where its impacts on ecosystem goods and services, such as forage production and biodiversity, are still being discovered. Reduced forage production has been observed in areas infested by annual grasses such as V. dubia (Jones et al. Reference Jones, Norton and Prather2018). The forage quality of cool-season perennial grasses, often used to improve forage resources on former cropland, have been tracked through a growing season in a planted, weed-free setting (Jenson et al. Reference Jensen, Robins, Rigby and Waldron2016). However, to our knowledge, no studies have documented how forage quality and quantity of an invaded rangeland change through a growing season.
Ventenata dubia can reduce biodiversity of areas it invades (Jones et al. Reference Jones, Norton and Prather2018, Reference Jones, Davis and Prather2020) and has negative impacts on wildlife and its habitat, namely birds, through trophic cascades (Mackey Reference Mackey2014). Similar to other invasive species, species richness and diversity were negatively correlated with increasing V. dubia cover (Jones et al. Reference Jones, Norton and Prather2018, Reference Jones, Davis and Prather2020). In some areas, this species has been observed replacing B. tectorum and T. caput-medusae—two infamously difficult species to control over the long term—as dominants (Barkworth et al. Reference Barkworth, Capels and Long1993).
Details on the effects of conservation and restoration on rangelands affected by invasive species are lacking (Sheley et al. Reference Sheley, James, Rinella, Blumenthal, DiTomaso and Briske2011). Historically, restoration efforts have often attempted to return ecosystems to a previous condition (Choi Reference Choi2007; Choi et al. Reference Choi, Temperton, Allen, Grootjans, Halassy, Hobbs, Naeth and Torok2008; Hobbs Reference Hobbs2007). Large restoration projects with goals such as this have raised concerns about whether such a goal is feasible. Critics argue such goals are too idealistic to apply in real-world situations with economic, social, and political interests in addition to the large ecological changes that have occurred, which prevent a return to previous ecological states (Choi et al. Reference Choi, Temperton, Allen, Grootjans, Halassy, Hobbs, Naeth and Torok2008; Hobbs Reference Hobbs2007).
With these concerns and research needs in mind, we studied two conditions: a Northern Great Plains rangeland invaded by V. dubia and a rangeland previously invaded by V. dubia that was subsequently controlled using herbicides. By comparing these two conditions, we evaluate whether chemical control of V. dubia improves attributes of rangeland condition. This overarching goal can be broken down into two questions: (1) Does V. dubia control improve forage quality and quantity? (2) Does V. dubia control increase plant species richness and diversity? Understanding the effects of V. dubia control on forage and plant community characteristics is a critical step in making informed recommendations to weed managers and restoration practitioners in this new region of spread.
Materials and Methods
Study Area
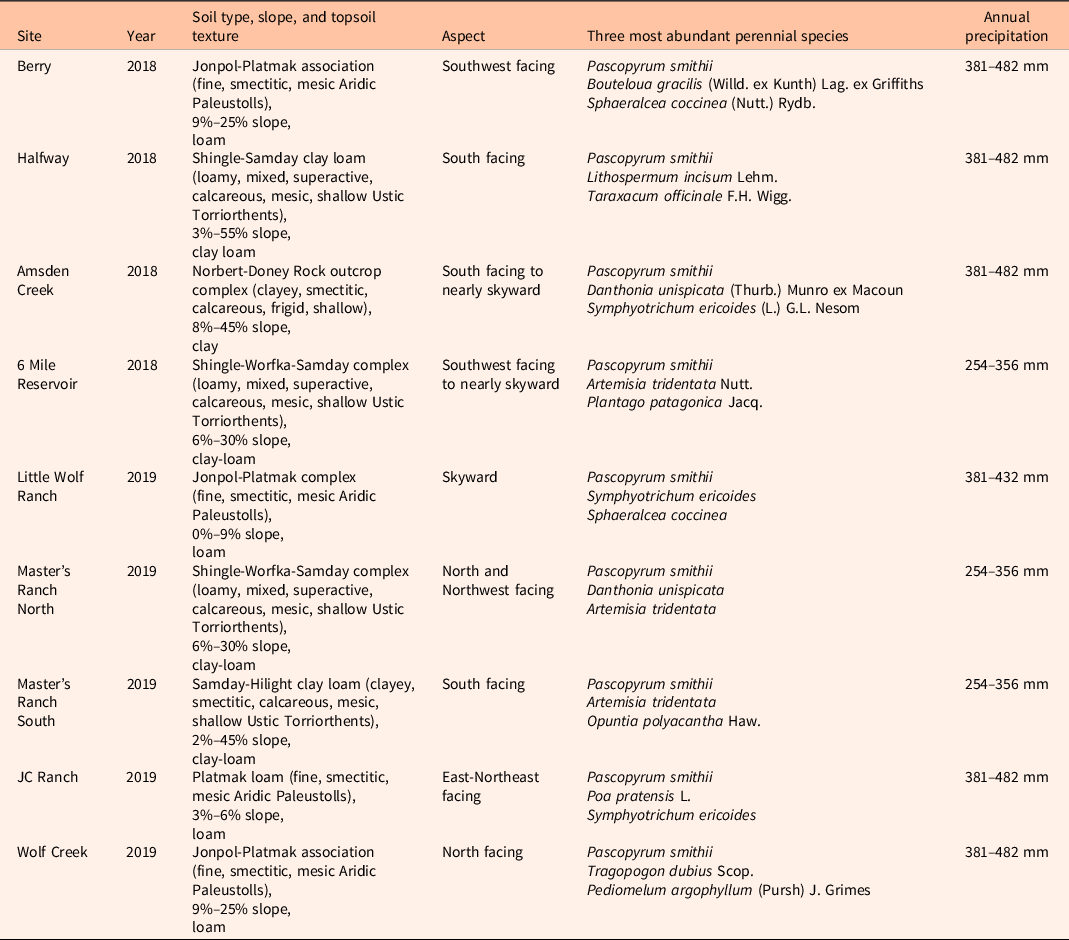
We conducted two single-year studies separated in space and time in conjunction with a cooperative, landscape-scale program focused on managing invasive annual grasses. Both studies took place in a region that represents a transition between northern mixed prairie and sagebrush grasslands. As part of an early detection–rapid response program for T. caput-medusae and V. dubia, the Northeast Wyoming Invasive Grasses Working Group (NEWIGWG) applied herbicide aerially to multiple sites in 2017 and 2018 to control annual grasses on sites where V. dubia was the dominant invasive grass (Supplementary Table S1). NEWIGWG treated approximately 1,200 ha in 2017, in numerous patches ranging from 20 to 400 ha per patch, which resulted in multiple instances of treated/nontreated paired sites across the landscape. Herbicide treatments consisted of fall treatments of 123g ae ha−1 of aminopyralid plus 123 g ai ha−1 of imazapic in 2017. In 2018, NEWIGWG treated another 2,800 ha in the same vicinity as before using 73 g ai ha−1 of indaziflam in a total solution of 46.8 L ha−1. Sites for both years varied in slope, aspect, and plant community and accounted for a wide range of environmental conditions within the locale (Table 1). Mean annual precipitation ranges from 254 to 482 mm and topsoils are clay, loam, clay-loam, or very fine sandy loam (USDA-NRCS 2019).
Table 1. Site information for the 2018 and 2019 studies of Ventenata dubia removal in Sheridan County, WY. a

a Treatment was 123 g ai ha−1 of imazapic and 123 g ae ha−1 of aminopyralid for the 2018 study and 73 g ai ha−1 of indaziflam for the 2019 study.
Data Collection
We collected vegetation data from four sites in 2018 and five sites in 2019 for treatments that occurred in 2017 and 2018, respectively, using slightly different approaches in each year (Figure 1). For our 2018 data collection, we sampled canopy cover by species along three 15.24-m transects placed in treated and nontreated portions of each site. Transects within a treatment area were separated by 30 to 60 m and between areas (treated/nontreated) were separated by at least 15 m. We sampled three 0.25-m2 quadrats spaced evenly along each transect (Figure 1). In each quadrat, we recorded canopy cover by species as a cover class: trace = 0% to 1%; class 1 = 1% to 5%; class 2 = 5% to 25%; class 3 = 25% to 50%; class 4 = 50% to 75%; class 5 = 75% to 95%; class 6 = 95% to 100%. We then clipped all aboveground herbaceous biomass near ground level, sorted it into functional groups (perennial grasses, perennial forbs, annual forbs, and annual grasses), dried the samples at 60 C for 48 h, and weighed them.

Figure 1. Field vegetation sampling scheme for Ventenata dubia treatment sites in 2018 and 2019 in northeast Wyoming. Plant cover and biomass data were collected in July from three 0.25-m2 quadrats along three transects per treatment per site at four sites in 2018. Plant biomass data were collected each month from two separate 0.25-m2 quadrats randomly placed within four 30-m2 sampling “plots” per treatment per site at five sites in 2019. Plant species cover data were collected only in July 2019. Vegetation data were pooled to the treatment × site level for statistical analysis (each site was treated as a replicate).
Sampling in 2019 was adjusted from the 2018 study. Instead of sampling once in July, as in the 2018 study, we sampled monthly from May to October, excluding September, at all five locations. At each site, we sampled four paired 30-m2 treated and nontreated plots along a treatment boundary (Figure 1). The size of treated and nontreated areas varied (see above), but all plots for each site were placed within 100 m of one another. We haphazardly placed two 0.25-m2 quadrats in each plot at each biomass collection date—avoiding previously clipped quadrats to prevent sampling of regrowth. We collected all aboveground herbaceous biomass in these quadrats, separated it into the same functional groups described earlier, and pooled each functional group for each plot before drying and weighing. In July, we collected additional data on species richness and canopy cover by species in each quadrat before clipping biomass. We collected these data using the same method as in 2018, but with adjusted cover classes for increased resolution of data as follows: trace = 1% to 3%, class 1 = 3% to 10%, class 2 = 11% to 20% class 3 = 21% to 30%, class 4 = 31% to 40%, class 5 = 41% to 50%, class 6 = 51% to 60%, class 7 = 61% to 70%, class 8 = 71% to 80%, class 9 = 81% to 90%, and class 10 = 91% to 100%.
Forage Quality
For the 2018 study, after we recorded the weight for each functional group, we pooled the biomass from a subset of sites into perennial and annual grasses for further nutritional content analysis. We ground this pooled biomass and analyzed the samples for crude protein and total digestible nutrients (TDN) using the combustion method (Padmore Reference Padmore1990) and acid detergent fiber method (ANKOM Technology Reference Technology2017). Crude protein and TDN are used because these are available and commonly used measures of nutrition.
For 2019 forage analysis, after biomass collection, we randomly selected plots from each site to provide separate samples: annual grass, perennial grass, and composite samples including all functional groups to provide a measure of total nutrients on the site. Randomization was done once and applied to both treatments and all sites for consistency of the samples. As previously, the primary metrics of forage quality were crude protein and TDN.
Data Analysis
For each sampling year, we pooled data among transects (2018) or plots (2019) by treatment (treated, nontreated) within a site and considered each site a replicate (Figure 1). In 2018, we used paired t-tests to compare biomass, cover, species richness, species diversity, crude protein, and TDN between herbicide treatments for the single sampling time in July. For the 2019 data, we used lack-of-fit tests and the Akaike information criterion to sequentially compare full and reduced regression models to evaluate effects of sampling time (month), herbicide treatment, and model shape (linear vs. curvilinear). We evaluated total biomass, biomass of separate functional groups, nutrient composition (%), and nutrient mass (kg ha−1). We calculated the Shannon-Wiener diversity index (Shannon Reference Shannon1948) using the midpoints of each canopy cover class as a measure of abundance for native and all species at each site. We analyzed species richness and diversity at the site level in treated and nontreated areas using paired t-tests, as we did in 2018. We analyzed and visualized all data using R v. 3.6.3 (R Core Team 2016) using base R and the ggplot2 and dplyr packages.
Results and Discussion
2018 Study
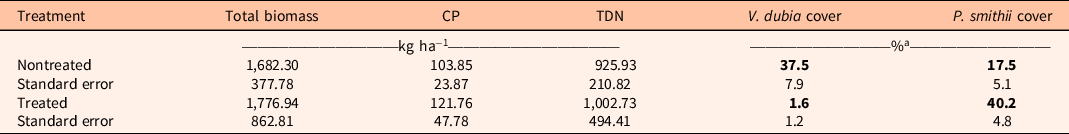
The imazapic and aminopyralid herbicide mix reduced total annual grass biomass, the most dominant of which was V. dubia (P = 0.002) and cover of V. dubia (P < 0.001; Table 2; Supplementary Table S1) effectively in 2018, 1 yr after treatment. However, we did not see an increase in perennial grass biomass with treatment. Canopy cover of western wheatgrass [Pascopyrum smithii (Rydb.) Á. Löve], a species either dominant or codominant at all sites, increased (P = 0.04; Table 2). Because perennial grass cover appeared to respond positively to V. dubia removal, the lack of a statistically significant biomass response was likely high variability in biomass production across sites.
Table 2. Mean biomass, crude protein (CP), total digestible nutrients (TDN), and cover of Ventenata dubia and Pascopyrum smithii before and after treatment with 123 g ai ha−1 of imazapic and 123 g ae ha−1 of aminopyralid in northeast Wyoming, 2018.

a Bold values are statistically significant (P < 0.05).
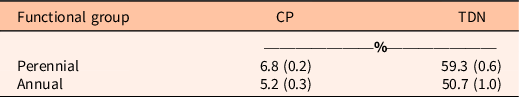
Annual grasses had lower crude protein and TDN than perennial grasses (P < 0.001; Table 3) in 2018. This is likely because annual grasses had senesced by July, when the samples were taken. Regardless, it is an indication of the loss of forage value due to V. dubia invasion. Loss of perennial grasses and replacement with annual grasses that become unpalatable early in the growing season result in a loss of nutrients available to cattle and wildlife (Fryer Reference Fryer2017; McCurdy et al. Reference McCurdy, Watts, Chibisa, Prather and Laarman2017).
Table 3. Crude protein (CP) and total digestible nutrients (TDN) of nontreated samples of perennial and annual grasses collected in July 2018 from four sites in Sheridan County, WY. a

a Standard error is in parentheses. Means between annual and perennial grasses differed for both crude protein and TDN (P < 0.001). All findings are significant (P < 0.001).
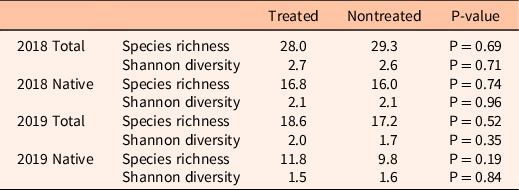
Control of V. dubia did not affect species richness 1 yr after treatment in 2018 (Table 4). Nontreated sites had 29.5 (±4.1) total species present, 17.3 (±2.29) of which were native. Treated sites had 27 (±5.7) total species with an average of 16.8 (±4.17) being native. Shannon diversity of total species and native species was similar between treatments (Table 4).
Table 4. Total and native species richness and Shannon diversity for 2018 and 2019 with associated P-values of treated and nontreated plots in Sheridan County, WY. a

a Treatment was 123 g ai ha−1 of imazapic and 123 g ae ha−1 of aminopyralid for the 2018 study and 73 g ai ha−1 of indaziflam for the 2019 study. Species richness and Shannon diversity did not change as a result of Ventenata dubia control.
2019 Study
NEWIGWG used indaziflam in 2018 under a Section 18 registration for Wyoming. Indaziflam is a PRE root growth–inhibiting herbicide (Sebastian et al. Reference Sebastian, Scott and Juliana2016a, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017) thought to remain close to the soil surface for several years, thus targeting annual species, which must reproduce by seed each year. Indaziflam is an effective herbicide for several winter annual species, including V. dubia (Sebastian et al. Reference Sebastian, Scott and Juliana2016a). Similar to the herbicides used in the 2018 study, indaziflam reduced annual grass biomass to nearly zero the year after treatment (2019), which was sustained through the growing season (P < 0.001; Figure 2).

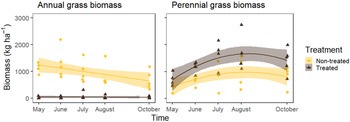
Figure 2. Annual and perennial grass biomass through the 2019 growing season with and without treatment with 73 g ai ha−1 indaziflam in Sheridan County, WY. Annual grass biomass was reduced to nearly zero after treatment (P < 0.001), while perennial grass biomass increased (P < 0.001). Differences in life histories are also apparent. Equations are as follows: nontreated annual grass biomass: y = 1239.12 − 117.62x [1]; treated annual grass biomass: y = 49.58 − 2.16x [2]; nontreated perennial grass biomass: y = 433 + 337.94x − 52.72x 2 [3]; treated perennial grass biomass: y = 693.75 + 582x − 87.58x 2 [4].
Because V. dubia was controlled effectively in both studies despite using different herbicides, we can be confident similar results between the two studies are due to removal of V. dubia and not simply direct effects of the herbicides themselves. Annual and perennial biomass curves differed between treatments (P < 0.001; Figure 2). In nontreated plots, annual grass biomass had a linear downward trend through the growing season as annual grasses quickly senesced and began to deteriorate after reproducing. In contrast, perennial grass biomass increased from the start of the growing season to peak biomass in August before senescing. This trend was true whether treated or nontreated; however, V. dubia control resulted in an overall increase in perennial grass biomass the season after treatment (P < 0.001; Figure 2). This difference in patterns between annual and perennial biomass is important, because it affects when forage is palatable and nutritious to cattle and wildlife. Biomass data indicate that perennial grasses have a longer, more stable, and more predictable green, grazable forage window than annual grasses. In this way, V. dubia control has given the land manger more grazing flexibility and reliability of the forage resource.
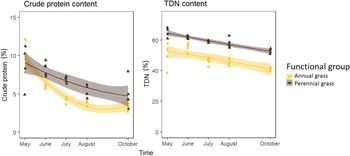
Regression of annual and perennial grass nutrient content in 2019 corroborated results from 2018. Perennial grasses had higher content of crude protein and TDN overall (P < 0.001; Figure 3). However, early in the growing season, when V. dubia is still green and growing, perennial and annual grasses had similar crude protein content. It would be tempting to view this as a positive. In this way, V. dubia is similar to B. tectorum, in that it could be valuable as an early forage (Young and Allen Reference Young and Allen1997) if it were palatable. However, this small benefit would need to be weighed properly against the many negatives.

Figure 3. Crude protein and total digestible nutrients (%) of perennial and annual grasses through the 2019 growing season in Sheridan County, WY. Perennial grasses were higher in both crude protein and TDN (P < 0.001). Equations are as follows: perennial grass crude protein (%): y = 9.08 − 1.40x + 0.11x 2 [5]; annual grass crude protein (%): y = 9.55 − 3.34x + 0.43x 2 [6]; perennial grass TDN (%): y = 64.42 − 2.44x [7]; annual grass TDN (%): y = 55.91 − 2.99x [8].
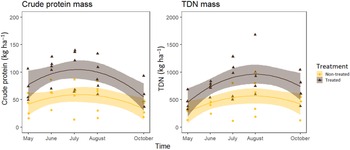
Given our findings that perennial grasses are higher in nutrient content and more abundant 1 yr after treatment, one would expect to see more nutrients available on the landscape. Treatment resulted in a higher percentage of crude protein and TDN (P < 0.001) of composite samples. However, even though perennial grasses and composite samples have higher nutritive content, converting to mass (kg ha−1) of crude protein or TDN did not reflect this. Total crude protein (P = 0.79) and TDN mass (P = 0.28) did not differ between treated and nontreated areas. This is likely due to the loss of V. dubia biomass confounding with the mass of these nutrients. In other words, the mass of crude protein and TDN lost from annual grass biomass removal was replaced by that gained from increases in perennial grasses, but not exceeded. Here it is important to keep in mind that even though V. dubia does have comparable nutrient mass, it is reportedly unpalatable to cattle unless it has been processed by pelletizing (McCurdy et al. Reference McCurdy, Watts, Chibisa, Prather and Laarman2017). Nutrients are not available to cattle or wildlife if they are not ingested. For this reason, we also analyzed nutrition in 2019 using only perennial grass biomass. Treatment increased mass of available crude protein and TDN (P < 0.001; Figure 4). Because perennial grass biomass increased as a result of V. dubia removal, we saw a corresponding increase in the mass of crude protein and TDN available to livestock and wildlife. This increase of available nutrients via annual grass control and restored perennial grasses on the landscape has direct benefits to grazing animals (Haferkamp et al. Reference Haferkamp, Grings, Heitschmidt, Macneil and Karl2001) and is one of the main benefits of V. dubia removal.

Figure 4. Mass of available crude protein and total digestible nutrients (TDN) found in perennial grasses through the growing season of 2019 for treated (73 g ai ha−1 indaziflam) and nontreated sites in Sheridan County, WY. Crude protein and TDN mass (P < 0.001) both increased as a result of treatment. Equations are as follows: nontreated available crude protein mass: y = 41.93 + 15.37x − 3.41x 2 [9]; treated available crude protein mass: y = 77.27 + 25.72x − 6.04x 2 [10]; nontreated available TDN mass: y = 278.51 + 197.06x − 33.66x 2 [11]; treated available TDN mass: y = 454.15 + 337.19x − 56.02x 2 [12].
Because V. dubia invasion has been documented lowering species diversity (Jones et al. Reference Jones, Norton and Prather2018, Reference Jones, Davis and Prather2020; Novak et al. Reference Novak, Cristofaro, Maguire and Sforza2015), we hypothesized some increase of species diversity with V. dubia control. Although removal of V. dubia resulted in large changes in cover for some species, such as P. smithii, we did not find a change in species richness 1 yr after treatment as a result of V. dubia removal in either study (P > 0.18; Table 4). In 2019, nontreated plots had 17.2 (±2.94) total species and 9.8 (±1.83) native species per site, while treated plots had 18.6 (±0.98) total species and 11.8 (±0.86) native species per site. Species diversity was also unaffected by treatment. This was true whether analyzing diversity for total species (P = 0.35) or native species only (P = 0.84). Shannon diversity was 1.69 (±0.27) and 1.58 (±0.15) for total and native species diversity, respectively, on nontreated plots. Treated plots had Shannon diversity indexes of 2.04 (±0.15) and 1.51 (±0.23) for total and native species, respectively.
Other than the possibility that species richness is unaffected by V. dubia on our study sites, which could be because they may already be negatively impacted by other nonnative species, the lack of species richness response could be due to a lack of recovery time after treatment. It may be that V. dubia has not been present at the sites long enough to extirpate any species as it has done farther west, where it has been present for longer. Indaziflam can remain in the soil for three or more years (Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016b, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017) while these sites were sampled 1 yr posttreatment. Previous work indicates that indaziflam does not negatively impact species richness or abundance 1 yr after treatment and may increase richness in the second year after treatment in rangelands of Colorado (Clark et al. Reference Clark, Sebastian, Nissen and Sebastian2019; Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016b, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017). Species richness and diversity data from 2020, the second year after indaziflam application (data not presented), did not display an increase in species richness or diversity in this transitional ecoregion of northeast Wyoming. However, our data clearly support that species richness is not further negatively impacted by indaziflam in this ecosystem. Indaziflam is therefore a potential tool for weed control where decreasing species richness is a concern. However, it should be noted that confident conclusions could not be made for annual forbs, as they constituted a very small (<1 g m−2) proportion of biomass on these sites, making trends difficult to measure effectively.
Species diversity is perhaps a more mathematical issue. The lack of response here is surprising, because we observed apparent visual differences between treated and nontreated areas. Our observations show that the reduction in V. dubia was replaced with a similarly dominant abundance of P. smithii, making the calculated diversity similar (Table 2; Supplementary Table S1). Pascopyrum smithii is a rhizomatous perennial grass and was the dominant native grass species at all sites. In this case, additional management may be required to affect richness and diversity.
These results are important and powerful when taken together in context. Ventenata dubia removal was a successful restoration effort that improved aspects of rangeland condition. The benefits of V. dubia control are not limited to simply increasing perennial forage biomass and available nutrition, often an explicitly stated goal of conservation, but having a more predictable, steady forage base through the growing season due to the demonstrated differences in biomass production between annual and perennial grasses. Our nutrition analyses would suggest that V. dubia could serve as an early forage, as its crude protein content is similar to that of perennial grasses early in the season and treatment did not result in higher total nutrient mass. It may be tempting to consider this high nutrition from annual grasses a benefit if more palatable annual grasses such as B. tectorum were dominant. Although B. tectorum was at one point thought of as beneficial in some ways (Young and Allen Reference Young and Allen1997), we believe this would be unwise. From our regression analyses, we demonstrated that the benefits would only be realized for a short period early in the year and would not offset the many negatives of annual grass invasion.
This study shows the impacts of annual grass control on sites where V. dubia was the dominant invader rather than the direct impacts of V. dubia itself on northern mixed prairie plant communities, because we did not include noninvaded sites in the analysis. However, this research documents improved forage quality and quantity where V. dubia and other, less-dominant, annual grasses were controlled—particularly with indaziflam. Landowners and managers in the Northern Great Plains concerned with maintaining grazing productivity and flexibility may benefit from proactive monitoring and management of this species in the region where it is rapidly expanding.
Acknowledgments
We thank the Natural Resource Conservation Service and the Sustainable Rangelands Roundtable for primary funding of this research. The Nature Conservancy provided partial support for this work through the Nebraska chapter’s J.E. Weaver Competitive Grants Program. This research was supported in part by the intramural research program of the U.S. Department of Agriculture, National Institute of Food and Agriculture, Hatch accession number 1013280. We also thank Master’s Ranch, Little Wolf Ranch, JC Ranch, and Wolf Creek Ranch for allowing our research to take place on their properties. Finally, we thank Wyoming Game and Fish, Wyoming State Lands, Wyoming Agricultural Experiment Station, Beth Fowers, Jordan Skovgard, Jaycie Arndt, Jodie Crose, Tyler Jones, Shawna LaCoy, Nancy Webb, Heidi Schueler, Steve Paisley, Kelsey Crane, and Kerry White for their contributions to this research in the lab and field. No conflicts of interest have been declared.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2021.20