Management Implications
Rangeland weeds cause severe ecological impacts, including depleting soil moisture and nutrients, reducing plant diversity and community productivity, altering fire frequency, and reducing recreational land values. Several herbicides approved for use in natural areas and rangeland can negatively affect native species, while the duration of weed control can be highly variable. Long-term weed control is critical in allowing sufficient time for native species recovery; therefore, herbicide options are needed that provide multiyear control without impacting the native plant community. Indaziflam, a newer herbicide option for preemergent invasive winter annual grass management, can provide control for 3 or more years, although there has been limited research on its effects on native species. A field study was conducted to evaluate changes in the native plant community composition from two annual grass herbicides, imazapic and indaziflam, as well as changes from two broadleaf herbicides, picloram and aminocyclopyrachlor, in diverse native perennial grass and forb communities. The study evaluated species richness and species abundance in the plant community for 2 yr. Picloram decreased native species abundance throughout the duration of the study across both sites, while aminocyclopyrachlor decreased species abundance at one site. Imazapic and indaziflam did not decrease species abundance or richness at either site over 2 yr. The results presented here suggest that indaziflam is an option for land managers to control winter annual grasses without negatively impacting existing native perennial species. In sites with a remnant native plant community, the multiyear winter annual grass control provided by indaziflam may allow enough time to achieve native species recovery.
Introduction
Downy brome (Bromus tectorum L.), an exotic winter annual grass, has emerged as one of the most invasive and problematic weeds in western rangeland and natural areas, with an estimated 14% annual spread rate (DiTomaso et al. Reference DiTomaso, Masters and Peterson2010; Duncan et al. Reference Duncan, Jachetta, Brown, Carrithers, Clark, DiTomaso, Lym, McDaniel, Renz and Rice2004). Although B. tectorum typically germinates in the fall after cool, wet weather, plants are opportunistic and can germinate anytime the growing conditions are favorable (Beck Reference Beck2009). This variable germination cycle has allowed B. tectorum to thrive in arid and semiarid western climates by rapidly growing and depleting available soil moisture and nutrients before most native species break dormancy in the spring (Knapp Reference Knapp1996; Mack and Pyke Reference Mack and Pyke1983). Invasions in natural ecosystems can cause severe negative impacts by reducing native plant diversity and lowering community productivity, increasing fire frequency, and displacing native vegetation that is critical wildlife and pollinator habitat (Abatzoglou and Kolden Reference Abatzoglou and Kolden2011; Beck Reference Beck2009; Billings Reference Billings1994; DiTomaso et al. Reference DiTomaso, Masters and Peterson2010; Knapp Reference Knapp1996; Monaco et al. Reference Monaco, Mangold, Mealor, Mealor and Brown2017; Whisenant Reference Whisenant1990).
By the 1930s researchers had begun to recognize B. tectorum invasions as a serious issue in rangeland (Mack Reference Mack1981; Price et al. 1948; Warg Reference Warg1938). Since then, extensive research has been conducted on mechanical, cultural, biological, and chemical control of this exotic grass. Thus far, herbicides have been the most effective and widely used weed management strategy for B. tectorum on rangeland and natural areas (Diamond et al. Reference Diamond, Call and Devoe2012; Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, Davis and Menalled2013; Monaco et al. 2017). Since its release in 1996, imazapic has been the primary herbicide used to control B. tectorum on rangeland, because it has both PRE and POST activity and is selective at relatively low use rates (Anonymous 1996; Kyser et al. Reference Kyser, Wilson, Zhang and DiTomaso2013; Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, Davis and Menalled2013). Several other herbicides, including glyphosate, sulfometuron, and rimsulfuron, have traditionally been used for B. tectorum control in non-crop sites (Kyser et al. Reference Kyser, Wilson, Zhang and DiTomaso2013; Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016). Although adequate control is often achieved with these herbicide options the first year of application, control can be inconsistent or short term, and injury to desirable species can occur (Kelley et al. Reference Kelley, Fernandez-Gimenez and Brown2013; Kyser et al. Reference Kyser, Wilson, Zhang and DiTomaso2013; Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, Davis and Menalled2013; Morris et al. Reference Morris, Monaco and Rigby2009; Thacker Reference Thacker, Ralphs and Monaco2009; Whitson and Koch Reference Whitson and Koch1998; Whitson et al. Reference Whitson, Majerus, Hall and Jenkins1997). In many invasive situations, short-term control (<2 yr) does not allow the remnant native plant community enough time to reestablish/recover and become competitive (Chambers et al. Reference Chambers, Pyke, Maestas, Pellant, Boyd, Campbell, Espinosa, Havlina, Mayer and Wuenschel2014; Elseroad and Rudd Reference Elseroad and Rudd2011). As B. tectorum infestations continue to spread, shifting native perennial grass systems to ecosystems dominated by winter annual grasses, land managers need strategies that provide long-term control of this weed.
Indaziflam is a broad-spectrum, PRE herbicide first released in 2011 for use in several perennial cropping systems and later used for weed control in turfgrass, ornamentals, forestry, and non-crop industrial sites (Anonymous 2011a, 2011b; Brabham et al. Reference Brabham, Lei, Gu, Stork, Barrett and DeBolt2014). In 2016 a supplemental label for indaziflam was approved for the release or restoration of desirable vegetation in natural areas, open spaces, wildlife management areas, and fire rehabilitation areas, specifically targeting invasive winter annual grass control in these sites (Anonymous 2016).
Indaziflam is a cellulose biosynthesis inhibitor, representing a unique site of action with no reported cases of resistance in the field (Brabham et al. Reference Brabham, Lei, Gu, Stork, Barrett and DeBolt2014; Tateno et al. Reference Tateno, Brabham and DeBolt2016). Indaziflam has a longer soil residual than other herbicides commonly used for B. tectorum management, providing 3 or more years of control (Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a). In most rangeland situations, this length of control allows enough time for release of the remnant native perennial community (Chambers et al. Reference Chambers, Pyke, Maestas, Pellant, Boyd, Campbell, Espinosa, Havlina, Mayer and Wuenschel2014; Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a). Sebastian et al. (Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a) found that indaziflam will selectively control B. tectorum without impacting perennial grass and forb biomass, even leading to significant increases in biomass due to reductions in B. tectorum (Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a). This research suggests native perennial species are tolerant to indaziflam, although studies assessing impacts to community composition and native species abundance following indaziflam applications have not been conducted.
The objective of this study was to evaluate tolerance of several native species to indaziflam applications and compare tolerance with other commonly used grass and broadleaf rangeland herbicides. We hypothesized that indaziflam would significantly reduce B. tectorum cover without decreasing native species abundance compared with the other herbicides evaluated.
Materials and Methods
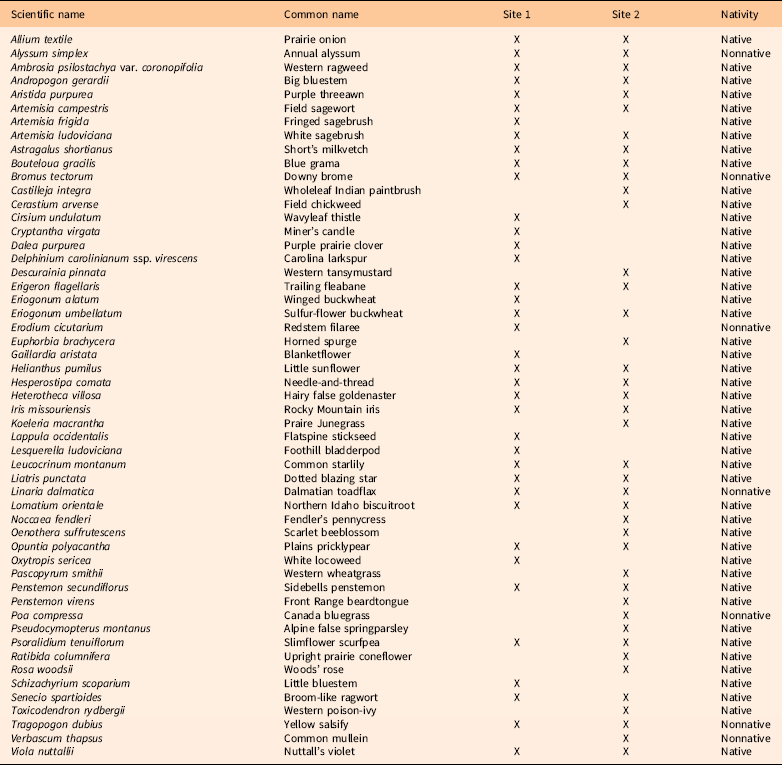
Site Description
The experiments were established in 2015 at two sites in Jefferson County, CO, containing B. tectorum with a co-occurring native grass, forb, and shrub community. Site 1 (39.760°N, 105.239°W) was located on Mount Galbraith Open Space, and Site 2 (39.894°N, 105.271°W) was located on El Dorado Mountain Open Space. Sites were approximately 15 km apart in the Western High Plains region of the Great Plains ecoregion. In June 2015, before herbicide application, we conducted an initial inventory of the plant species by recording all plant species present at each site within the boundaries of the plots. A visual estimate of B. tectorum canopy cover (%) was also done at both sites. Site 1 was categorized as ~30% to 40% B. tectorum cover with 33 native species and 5 co-occurring nonnative species (Table 1). By the following year (2016), B. tectorum cover at Site 1 decreased to ~5% and maintained a similar cover level throughout the course of the study. Site 2 had ~60% to 70% B. tectorum cover with 35 native species and 6 co-occurring nonnative species (Table 1).
Table 1 List of species occurring at Site 1 and Site 2 with their nativity status.

The soil at Site 1 was Ratake rocky loam (loamy-skeletal, micaceous, frigid, shallow Typic Haplustolls), with 2.3% organic matter and 6.0 pH in the top 20 cm (USDA-NRCS 2014). Site 1 was located on a 30° rocky slope, and the average elevation was 1,839 m (6,035 ft). The soil at Site 2 was Flatirons stony sandy loam (clayey-skeletal, smectitic, mesic Aridic Paleustolls), with 4.9% organic matter and 6.6 pH in the top 20 cm (USDA-NRCS 2014). Site 2 was located on a 25° rocky slope, and the average elevation was 1,995 m (6,544 ft). Mean annual precipitation based on the 30-yr average (1981 to 2010) was 468 mm at Site 1 and Site 2 based on the closest weather station (approximately 8 km from each site) (Western Regional Climate Center 2018).
Both sites received an additional 252 mm of precipitation above their 30-yr average in 2015. A statewide drought occurred in 2016, and total precipitation for the sites decreased to 148 mm below the 30-yr average (Western Regional Climate Center 2018). In 2017 the sites received precipitation similar to the 30-yr average. The 30-yr mean annual temperature for both sites was 10.2 C, and the average temperature for 2015 was close to the 30-yr average. The average temperatures for 2016 and 2017 were 1.8 C and 2.2 C warmer, respectively (Western Regional Climate Center 2018).
Experimental Design
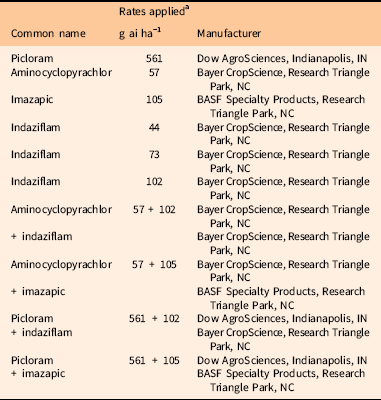
Herbicide applications were made June 2, 2015 and timed in accordance with label recommendations for Dalmatian toadflax [Linaria dalmatica (L.) Mill.] control, one of the co-occurring nonnative species (Anonymous 2018). The herbicides targeting B. tectorum were applied at an early PRE application timing. Bromus tectorum was in the ripening stage and actively setting seed. Native forb growth stage ranged from postflowering, early flowering to preflowering. Ten herbicide treatments and one nontreated control were established in 3 by 6 m plots arranged in a randomized complete block design with six replications (Table 2). All treatments were applied with a CO2-pressurized custom-built backpack sprayer using 11002LP flat-fan nozzles (TeeJet® Spraying Systems, P.O. Box 7900, Wheaton, IL 60187) delivering 187 L ha−1 at 207 kPa.
Table 2 Herbicides and rates applied in evaluating Bromus tectorum control and native species tolerance.

a All treatments included 0.25% v/v nonionic surfactant.
Treatment Evaluations and Data Analysis
To account for variability across the study area, native forb and shrub species were counted individually throughout the entire area of each plot from May to August (2016 and 2017) to determine species richness (total number of species) and abundance (number of individuals per species). Counts were conducted biweekly, targeting different species each time to account for varying life cycles, and individual species were counted only once per growing season. For rhizomatous or clonal plants, each clumping patch or grouping of stems was counted as one individual. To determine B. tectorum and native grass canopy cover, percent cover estimates of all grass species were determined by conducting visual evaluations across each entire plot (18-m2 plot area) in August 2016 and 2017. Species richness was defined as the total number of different species occurring by plot (18-m2 plot area), while species abundance was defined as total number of individuals per species per plot (18-m2 plot area). Native grass cover was collected as percent cover per species; however, due to variability across the sites, species were combined into cool-season (C3) and warm-season (C4) cover categories. Cover data for B. tectorum and perennial grasses were arcsine square-root transformed to meet the assumptions of normality.
To test treatment effects on B. tectorum cover, a repeated-measures linear mixed-effects model was created using the ‘lme4’ package in R v. 3.4.3, testing for treatment effects at α=0.05 (R Core Team 2017). The fixed factors included in the model were treatment, year, and interactions, with year as the repeated measure; block was included as a random factor. Further analysis of the treatment and year effect was performed using the ‘lsmeans’ package in R (R Core Team 2017) to obtain comparisons between all pairs of least-squares means by year with a Tukey adjustment (P<0.05). For grass cover data, the same analysis was performed for C3 grass cover and C4 grass cover. After rejection of the null hypothesis of equal variance for Sites 1 and 2, grass cover was analyzed separately by site.
Species richness was calculated by determining the number of native species in each plot. After failure to reject the null hypothesis of equal variance (P=0.3401), sites were combined. A generalized linear mixed model was used to analyze count data, with treatment and year as fixed factors and block as the random factor. Count data for species richness were assumed to follow a Poisson distribution after failure to reject the null hypothesis that sample frequencies differed significantly from the expected frequencies under a Poisson distribution (P=0.1113). Significant pairwise differences between richness were determined post hoc using a least-squares means test with a Tukey adjustment (‘stats’ and ‘lsmeans’ packages; R Core Team 2017).
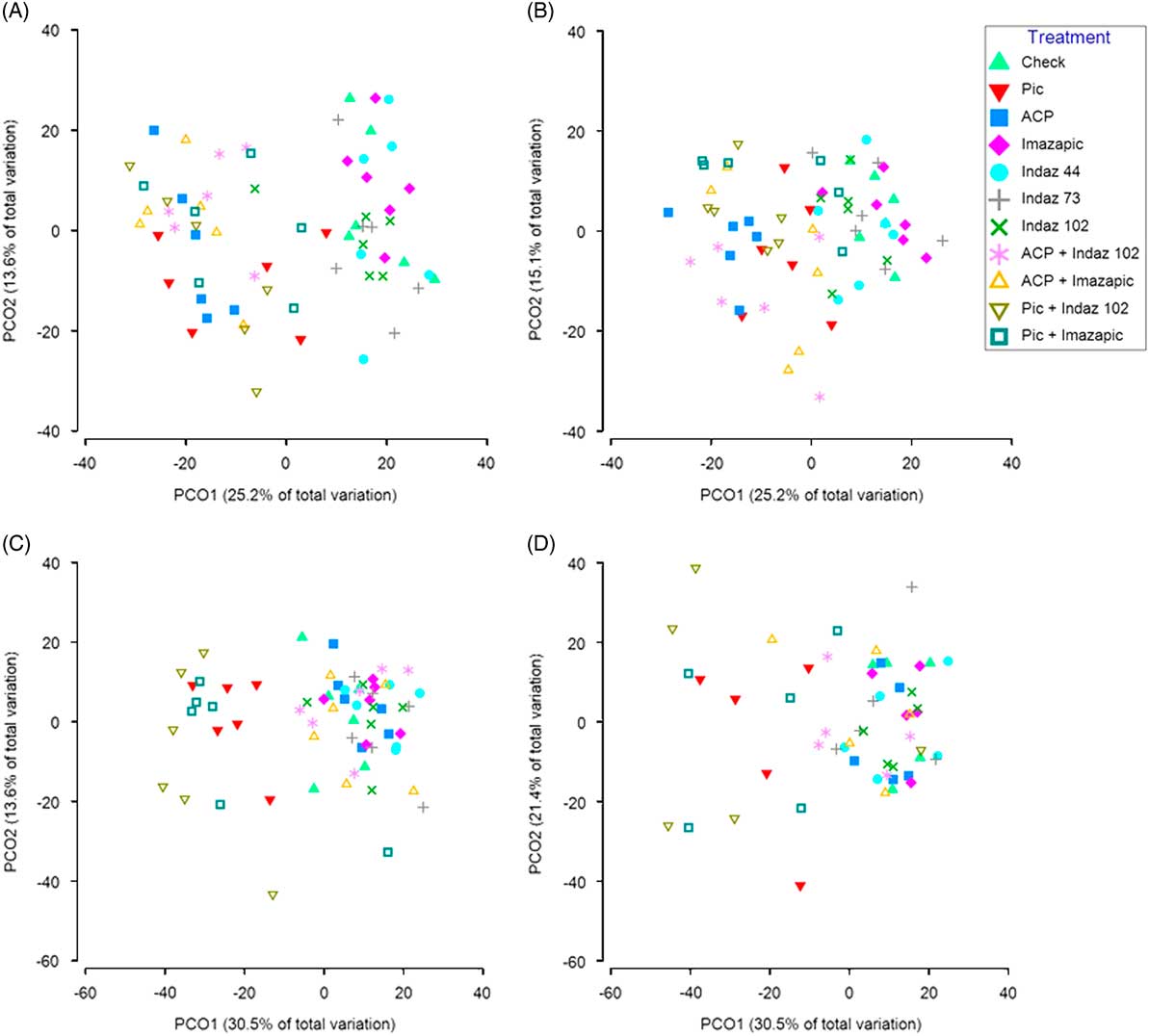
To test treatment effects on overall native forb and shrub community abundance, dissimilarity matrices were generated on the collected abundance data using the Bray-Curtis method in Primer v. 7 (Bray and Curtis Reference Bray and Curtis1957; Clarke and Gorley Reference Clarke and Gorley2015). Due to varying species amounts and types occurring at each site, sites and years were analyzed separately. Count data for each species were square-root transformed before creation of a resemblance matrix for each site and year by using Bray-Curtis dissimilarity measures. Homogeneity of variance (or dispersion) at each site by year was tested using permutational analysis of multivariate dispersions and was significant for Site 2 in 2017 (Site 1, 2016 P=0.109; Site 2, 2016 P=0.257; Site 1, 2017 P=0.055; Site 2, 2017 P=0.002). There was a dispersion effect at Site 2 in 2017. Permutational multivariate analysis of variance (PERMANOVA) is largely unaffected by heterogeneity for balanced designs and is more powerful than other tests in detecting actual changes in community structure (Anderson and Walsh Reference Anderson and Walsh2013). Therefore, the resulting resemblance matrices were used to generate principal coordinate analyses (PCoA) to visualize differences among treatments. PERMANOVA was conducted to test treatment effects on native forb and shrub community composition (Anderson Reference Anderson2001; Anderson et al. Reference Anderson, Gorley and Clarke2008). PERMANOVA can be used as a nonparametric alternative to MANOVA and allows analysis of multiple variables (i.e., species counts) when data do not meet the assumptions of MANOVA. PERMANOVA were conducted using partial sums of squares on 999 permutations of residuals under a reduced model. Factors considered in the model were treatment as a fixed factor and block as a random factor. All multivariate analyses were conducted using PRIMER v. 7 and PERMANOVA+ (Primer-E, Plymouth, UK). Pairwise tests were performed by treatment levels using PERMANOVA+ (Primer-E, Plymouth, UK). Similarity percentage analysis (SIMPER) (Primer v. 7) for significant treatments was then used to identify specific species accounting for greater than 60% of the dissimilarity in community composition compared with the nontreated control. The analysis revealed whether the dissimilarity was primarily due to increases or decreases in species abundance.
Results and Discussion
Bromus Tectorum Control
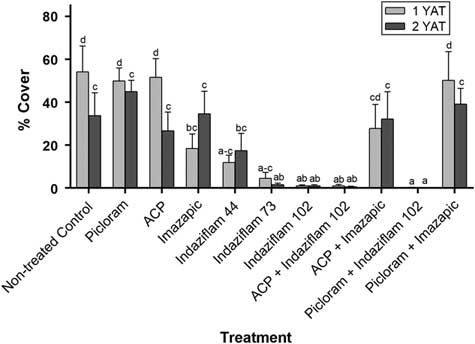
Bromus tectorum cover decreased significantly at Site 1 during 2016 and 2017; therefore, only Site 2 was analyzed for treatment impacts to B. tectorum cover. Treatment was the only significant factor impacting B. tectorum cover (P<0.001) (Supplementary Table S1). Compared with the nontreated control, all treatments containing indaziflam had less B. tectorum cover at 1 yr after treatment (YAT) (0% to 22% cover). Indaziflam at the highest rate (102 g ai ha−1) alone or tank mixed with aminocyclopyrachlor or picloram resulted in only 0.7±0.3% (mean ± SE) B. tectorum cover at 1 YAT. The only other treatment to reduce B. tectorum cover at 1 YAT was imazapic applied alone (21.6±0.6%) (Figure 1). Indaziflam at the highest rates (73 and 102 g ai ha−1) alone or tank mixed with aminocyclopyrachlor or picloram continued to reduce B. tectorum cover at 2 YAT (0.8±0.3%) (Figure 1). Although our data only represent one site, they were consistent with past studies showing multiyear B. tectorum control with indaziflam treatments compared with short-term (<1 yr) control with imazapic treatments (Kyser et al. Reference Kyser, DiTomaso, Doran, Orloff, Wilson, Lancaster, Lile and Porath2007; Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, Davis and Menalled2013; Morris et al. Reference Morris, Monaco and Rigby2009; Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a).

Figure 1 Percentage Bromus tectorum cover at Site 2 at 1 yr after treatment (YAT) (2016) and 2 YAT (2017). Letters indicate significant differences among herbicide treatments across years, using least-squares means (P<0.05). Herbicide treatment rates are as follows: picloram (561 g ai ha−1), aminocyclopyrachlor (ACP; 57 g ai ha−1), imazapic (105 g ai ha−1), indaziflam (44, 73, and 102 g ai ha−1), and nontreated control.
Impacts to Native Grasses
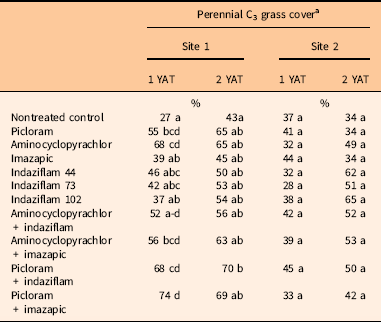
All native grasses occurring across the two sites were perennial grasses. Site 1 had significant treatment (P<0.001) and year (P=0.02) effects for C3 grasses; however, the interaction of year by treatment was not significant (Supplementary Table S1). Comparisons made to the nontreated control showed increases in C3 grass cover at 1 YAT for treatments containing picloram and aminocyclopyrachlor (average of 27% C3 grass cover in nontreated control plots compared with 52% to 74% cover in the picloram- and aminocyclopyrachlor-treated plots). By 2 YAT, the only significant difference in C3 grass cover at Site 1 was between the nontreated control and picloram plus indaziflam treatment (Table 3). There was no difference in warm-season grass cover at 1 or 2 YAT at Site 1 (Supplementary Tables S1 and S2). With little competition from B. tectorum at Site 1, differences in perennial grass cover were likely due to the forb reduction from the broadleaf herbicides. At Site 2 there was no treatment effect on C3 grass cover (P=0.6324) (Supplementary Table S1). For C4 grass cover, a post hoc Tukey test revealed no significant pairwise differences between individual means separated by treatment (Supplementary Tables S1 and S2).
Table 3 Mean percentage cover of perennial cool season (C3) grasses at both sites 1 and 2 yr after treatment (YAT).

a Means followed by the same letter do not differ significantly within year at P<0.05.
Impacts to Species Richness
The treatment by year interaction was not significant (P=0.4609) for species richness although there was a treatment effect (P<0.001). The only treatment to impact species richness was picloram combined with indaziflam 1 YAT, which reduced species richness compared with the nontreated control. The picloram plus indaziflam treatment had an average richness of 7.4±0.66 compared with the nontreated control with an average species richness of 12.8±0.59. By 2 YAT no impacts to species richness were observed (Supplementary Figure S1).
Impacts to Community Composition
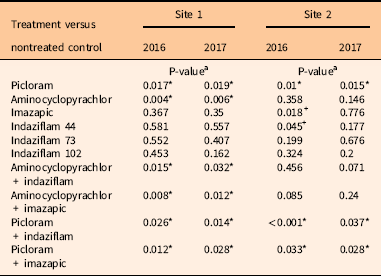
Visualization of the PCoA plots suggested changes in community composition due to herbicide treatments at both sites, so a PERMANOVA analysis was performed to determine any treatment differences (Figure 2). For Site 1, PERMANOVA analysis showed impacts to the community composition of native species from herbicide treatments at 1 and 2 YAT (P<0.001). All treatments containing picloram and aminocyclopyrachlor impacted species abundance compared with nontreated control plots at both 1 and 2 YAT (Table 4). Further analysis with SIMPER revealed that both broadleaf herbicide treatments decreased the abundance of most native forbs and shrubs included in the analysis (Supplementary Data File S1). Hairy false goldenaster [Heterotheca villosa (Pursh) Shinners var. villosa] and western ragweed [Ambrosia psilostachya DC. var. coronopifolia (Torr. & A. Gray) Farw.] were most impacted, contributing to more than 20% of the dissimilarity to the nontreated control at both 1 and 2 YAT (Supplementary Data File S1). No treatments resulted in increased species abundance at Site 1 (Table 4; Figure 2). At Site 2, the PERMANOVA also showed treatment effects to community composition of native species in both years (P<0.001). At 1 YAT, treatments containing picloram reduced the abundance of most species, while treatments of imazapic alone and indaziflam at 44 g ai ha−1 increased species abundance compared with the nontreated control (Table 4). In year 2, treatments containing picloram still reduced species abundance; however, no treatments increased species abundance compared with the nontreated control (Table 4; Figure 2). Picloram had the greatest impacts to H. villosa and trailing fleabane (Erigeron flagellaris A. Gray), accounting for more than 30% and 40% of the dissimilarity to the nontreated control in 1 and 2 YAT, respectively (Supplementary Data File S1). In the imazapic-alone treatment, the greatest increases to abundance were to A. psilostachya, western poison-ivy [Toxicodendron rydbergii (Small ex Rydb.) Greene], and H. villosa, accounting for almost 40% of the dissimilarity to the nontreated control (Supplementary Data File S1). Indaziflam (44 g ai ha−1) had the greatest increases to A. psilostachya, horned spurge (Euphorbia brachycera Engelm.), Nuttall’s violet (Viola nuttallii Pursh), and sidebells penstemon (Penstemon secundiflorus Benth.), accounting for almost 45% of the dissimilarity (Supplementary Data File S1). Additional species contributing to the dissimilarity to the nontreated control can be viewed in Supplementary Data File S1. Reducing B. tectorum abundance can lead to increases in perennial grass and forb abundance as the competition for resources is removed (Monaco et al. Reference Monaco, Mangold, Mealor, Mealor and Brown2017; Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a; Thill et al. Reference Thill, Beck and Callihan1984; Whitson and Koch Reference Whitson and Koch1998); therefore, the increases in species abundance in Site 2 in the indaziflam and imazapic treatments are likely due to the reduction in B. tectorum cover.

Figure 2 Principal coordinate analysis (PCoA) of native forb and shrub species abundance separated by treatment. Treatments farther away from the nontreated (check, represented by the green triangle) had more dissimilarities in community composition. The analysis was based on the Bray-Curtis dissimilarity matrix constructed using the square-root-transformed species counts from Site 1 in 2016 (A) and 2017 (B) and Site 2 in 2016 (C) and 2017 (D). The percent of variation explained is given in brackets on the x- and y-axes. ACP, aminocyclopyrachlor; Indaz, indaziflam; PIC, picloram.
Table 4 Pairwise comparisons of herbicide treatments versus nontreated control for native forb abundance from permutational multivariate analysis of variance analysis.

a P-values marked with asterisks (*) are considered significant abundance reductions at the <0.05 level. P-values marked with plus signs (+) are considered significant abundance increases at the <0.05 level.
The impacts to the native plant community differed between the two sites, although some responses to treatments were the same. At Site 1, no treatments resulted in increased native forb or shrub abundance compared with the nontreated control, while broadleaf herbicides increased C3 grass cover. At the same site, both broadleaf herbicides (picloram and aminocyclopyrachlor) reduced native forb and shrub abundance, while the annual grass herbicides (indaziflam and imazapic) had no impact on the overall community composition. The shift to a more C3 grass–dominated community in the plots treated with broadleaf herbicides is likely due to the reduction in forb and shrub abundance (Arnold and Santelmann Reference Arnold and Santelmann1966; Greet et al. Reference Greet, Mealor and Kniss2016). At Site 2, which was characterized by 60% to 70% B. tectorum cover, only picloram decreased species abundance, while increases in species abundance at 1 YAT were observed among a few treatments that reduced B. tectorum cover. Decreases in species richness were also observed from one picloram treatment at this site as well. These findings support extensive research showing decreases in native forb abundance from picloram applications and more recent work showing transient forb decreases from aminocyclopyrachlor applications (Arnold and Santelmann Reference Arnold and Santelmann1966; Carter and Lym Reference Carter and Lym2018; Greet et al. Reference Greet, Mealor and Kniss2016; Ortega and Pearson Reference Ortega and Pearson2011; Thilmony and Lym Reference Thilmony and Lym2017; Wagner and Nelson Reference Wagner and Nelson2014).
Much of the research examining herbicide impacts on native species abundance is compounded by noxious weed competition at the site (Arnold and Santelmann Reference Arnold and Santelmann1966; Beran et al. Reference Beran, Masters and Gaussoin1999; Carter and Lym Reference Carter and Lym2018; Davies and Sheley Reference Davies and Sheley2011; Elseroad and Rudd Reference Elseroad and Rudd2011). This can make separating herbicide impacts from invasion impacts difficult. In sites dominated by invasive weeds, especially long-term invasions, the diversity of the native plant community has already been compromised, while in noninvaded, intact plant communities, there is a higher potential for loss as native species have not been impacted by nonnative invaders (Davies and Sheley Reference Davies and Sheley2011; Duncan et al. Reference Duncan, Jachetta, Brown, Carrithers, Clark, DiTomaso, Lym, McDaniel, Renz and Rice2004). In a study conducted by Ortega and Pearson (Reference Ortega and Pearson2011), the authors presented an impact gradient for picloram that coincided with spotted knapweed (Centaurea stoebe L.) invasion levels. Their study found that native forb cover declined >20% in treated plots versus control plots at noninvaded sites, while impacts to forb cover were minimal in sites with moderate to high C. stoebe invasion levels. The authors concluded that differences in picloram effects on native species was due to the strength of release effects from the invasive species, as the increased diversity in sites void of C. stoebe had more loss potential than sites already suffering the effects of invasion. This offers a possible explanation for why decreased native species abundances were observed from aminocyclopyrachlor treatments in the site with a more intact native plant community versus the site dominated by B. tectorum.
To date, the two published field studies showing indaziflam treatments resulting in long-term B. tectorum control reported no observable negative impacts to native species (Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a). Bromus tectorum control with indaziflam at 58 g ai ha−1 lasted 3 yr with no injury to crested wheatgrass [Agropyron cristatum (L.) Gaertn.] and western wheatgrass [Pascopyrum smithii (Rydb.) Á. Löve] or impacts to forb species richness (Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016). Another study by Sebastian et al. (Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a) reported 2 yr of B. tectorum control from indaziflam (44, 73, and 102 g ai ha−1), with increased perennial grass and forb biomass and no impact to forb species richness. Our data corroborate previous findings of native species tolerance to indaziflam applications, while also showing that the community composition and abundance of native species is not impacted. The literature on impacts to perennial species abundance with imazapic applications is more diverse, and past findings have been variable, showing no impact to species abundance or impacts to specific perennial species, especially in areas with low annual precipitation or during periods of drought (Beran et al. Reference Beran, Masters and Gaussoin1999; Kyser et al. Reference Kyser, DiTomaso, Doran, Orloff, Wilson, Lancaster, Lile and Porath2007; Monaco et al. Reference Monaco, Osmond and Dewey2005; Morris et al. Reference Morris, Monaco and Rigby2009; Shinn and Thill Reference Shinn and Thill2002). Our study found no evidence of decreases in species abundance with imazapic applications. Our findings from Site 2 are consistent with previous research showing multiyear B. tectorum control with indaziflam applications (Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a) and variability in control with imazapic applications (Davies and Sheley Reference Davies and Sheley2011; Davison and Smith Reference Davison and Smith2007; Elseroad and Rudd Reference Elseroad and Rudd2011).
One important aspect land managers must take into account when considering the results from this study and developing large-scale weed management plans is interannual variability in plant community composition. Although B. tectorum was initially a target species for control in this study, the cover at one site decreased to a negligible level (<5% cover) the year after treatments were applied. Bromus tectorum invasion levels can decrease during periods of drought and return with increased fall and winter moisture (Mack and Pyke Reference Mack and Pyke1983). Climatic variation can also impact weed control and injury to native species from herbicide treatments (Evans et al. Reference Evans, Eckert, Burgess and Young1969; Sebastian et al. Reference Sebastian, Nissen, Westra, Shaner and Butters2017c).
Indaziflam is an effective tool for multiyear B. tectorum control (Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a), and our results suggest that this herbicide can be used in non-crop sites without impact to native perennial species. Land managers should consider impacts to the plant community when using broadleaf herbicides in these sites, as there is a potential to decrease forb and shrub abundance and shift to a more grass-dominated ecosystem. Integrating indaziflam into current management programs could provide the length of B. tectorum control needed to deplete the invasive annual grass seedbank and release the remnant plant community (Chambers et al. Reference Chambers, Pyke, Maestas, Pellant, Boyd, Campbell, Espinosa, Havlina, Mayer and Wuenschel2014; Elseroad and Rudd Reference Elseroad and Rudd2011; Sebastian et al. Reference Sebastian, Nissen, Sebastian and Beck2017b). Reestablishing the dominant native perennial plant community further increases the resistance and resilience of that ecosystem to future B. tectorum invasions and decreases fine fuels from invasive annual grass that are associated with wildfires (Chambers et al. Reference Chambers, Pyke, Maestas, Pellant, Boyd, Campbell, Espinosa, Havlina, Mayer and Wuenschel2014). Future studies should evaluate the length of B. tectorum control and native species tolerance across varying climates and soil types, specifically in more arid regions such as the Great Basin.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2019.4
Acknowledgments
We would like to thank Jefferson County Parks and Open Space Small Grants Program for funding this project, as well as Jerry Bader from the Jefferson County Parks and Open Space for his assistance in locating the research sites. We would also like to thank the two anonymous reviewers who provided excellent comments for the revision of the article. No conflicts of interest have been declared.