1. Introduction
Diploporite ‘cystoids’ are a clade of early to middle Palaeozoic (Ordovician–Devonian) blastozoan echinoderms characterized by dipore-based respiratory structures, globular thecae commonly with irregular plating, and a passive suspension-feeding sessile mode of life (Broadhead, Reference Broadhead, Broadhead and Waters1980). Although many diploporites were free-living as adults, lying directly but loosely on the seafloor, others remained permanently affixed to hard substrates throughout life (e.g. Paul, Reference Paul1971, Reference Paul1973; Frest, Mikulic & Paul, Reference Frest, Mikulic and Paul1977; Frest, Strimple & Paul, Reference Frest, Strimple and Paul2011). Consequently, attachment structures representing direct cementation to isolated skeletal substrates (e.g. shells), isolated lithified substrates (e.g. cobbles, exhumed concretions) and hardground surfaces have the potential to provide much insight into diploporite palaeobiology and palaeoecology, particularly in settings where articulated thecae are rare or absent (Thomka & Brett, in press). Indeed, given the propensity for rapid post-mortem disarticulation of diploporite thecae (Brett, Moffat & Taylor, Reference Brett, Moffat, Taylor, Maples and Waters1997) and the relative robustness of many echinoderm attachment structures (Lewis, Reference Lewis and Sprinkle1982), research focused on holdfasts may yield important ecological insights.
Hard substrates are commonly densely encrusted by pelmatozoan holdfasts (see Brett, Reference Brett1988, Reference Brett, Bassett, Lane and Edwards1991; Wilson & Palmer, Reference Wilson and Palmer1992; Taylor & Wilson, Reference Taylor and Wilson2003; McLaughlin, Brett & Wilson, Reference McLaughlin, Brett, Wilson, Pratt and Holmden2008 among many others); however, multiple generations of encrusters can be preserved attached to a single surface. Although this time-averaging (Walker & Bambach, Reference Walker and Bambach1971) complicates ecological interpretations derived from direct observations, taphonomic data can be used to successfully differentiate attachment structures that were subjected to varying degrees of physical or biological alteration prior to burial (Brett & Liddell, Reference Brett and Liddell1978; Paul & Bockelie, Reference Paul and Bockelie1983; Brett, Deline & McLaughlin, Reference Brett, Deline, McLaughlin, Ausich and Webster2008). Hence, assessment of diploporite holdfast preservation can be critical for understanding short-term trends in substrate preferences, biotic interactions and time resolution in select Palaeozoic hard substrate environments.
A recently discovered carbonate hardground in the middle Silurian of southeastern Indiana, United States was encrusted by numerous attachment structures representing a variety of crinozoan and blastozoan echinoderms, including abundant diploporite thecal attachments (Thomka & Brett, in press). This surface presents a rare opportunity not only to document spatial and substrate relationships among diploporites, an area of research in which few studies have been published (see Gil Cid & Domínguez-Alonso, Reference Gil Cid, Domínguez-Alonso and Barker2000 for an important exception), but also to record the signature of taphonomic processes on skeletal modules that have the highest chance of preservation and recognition in a group with an overall poor fossil record. In addition, reconstructing the taphonomic history of diploporite holdfasts has led to documentation of several biotic interactions between diploporites and associated biota that have significance for Silurian benthic palaeosynecology.
2. Locality and stratigraphic setting
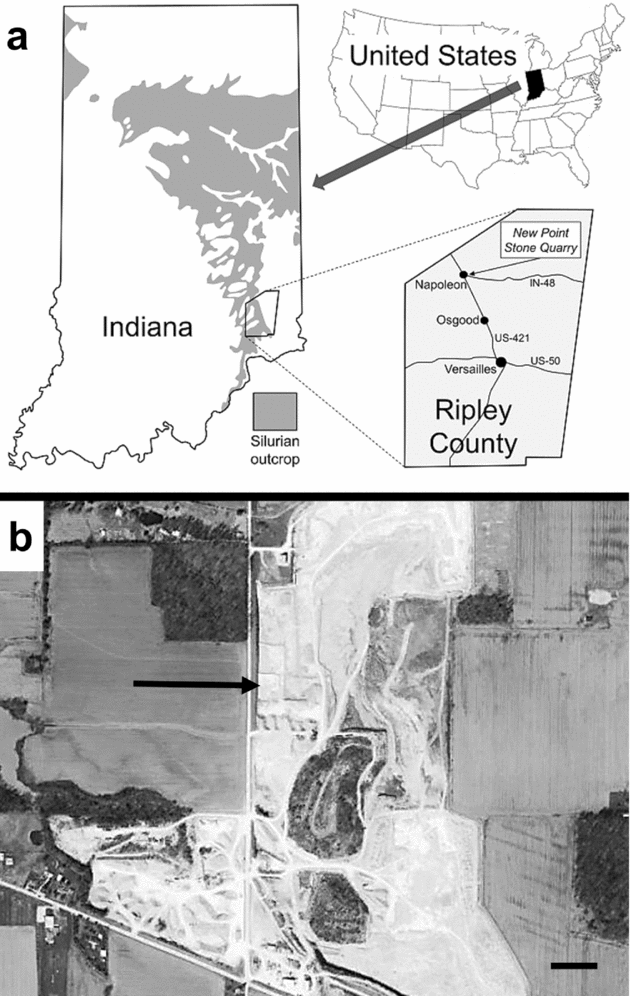
The encrusted hardground serving as the basis of this study is exposed at the northern end of the New Point Stone Company quarry located approximately 1 km east of Napoleon in northern Ripley County, southeastern Indiana, east-central United States (Fig. 1). This is not an active portion of the quarry and has not historically been visited by fossil collectors or geological field trips. The surface was cleared of overburden, presumably by a bulldozer, and has served as a benched-off potential dump pile site. Fortunately, scrapes created by quarrying machinery are readily recognizable and are easily distinguished from taphonomic features produced prior to sediment burial. Further, no evidence for skeletal disarticulation or significant dissolution occur from exposure of the surface to modern weathering.
Figure 1. Study site. (a) Location of study area approximately 1 km east of Napoleon in northern Ripley County, southeastern Indiana, east-central United States. Modified from Thomka & Brett (in press). (b) Aerial photograph of quarry, with holdfast-bearing hardground marked by the arrow. Scale bar is approximately 100 m.
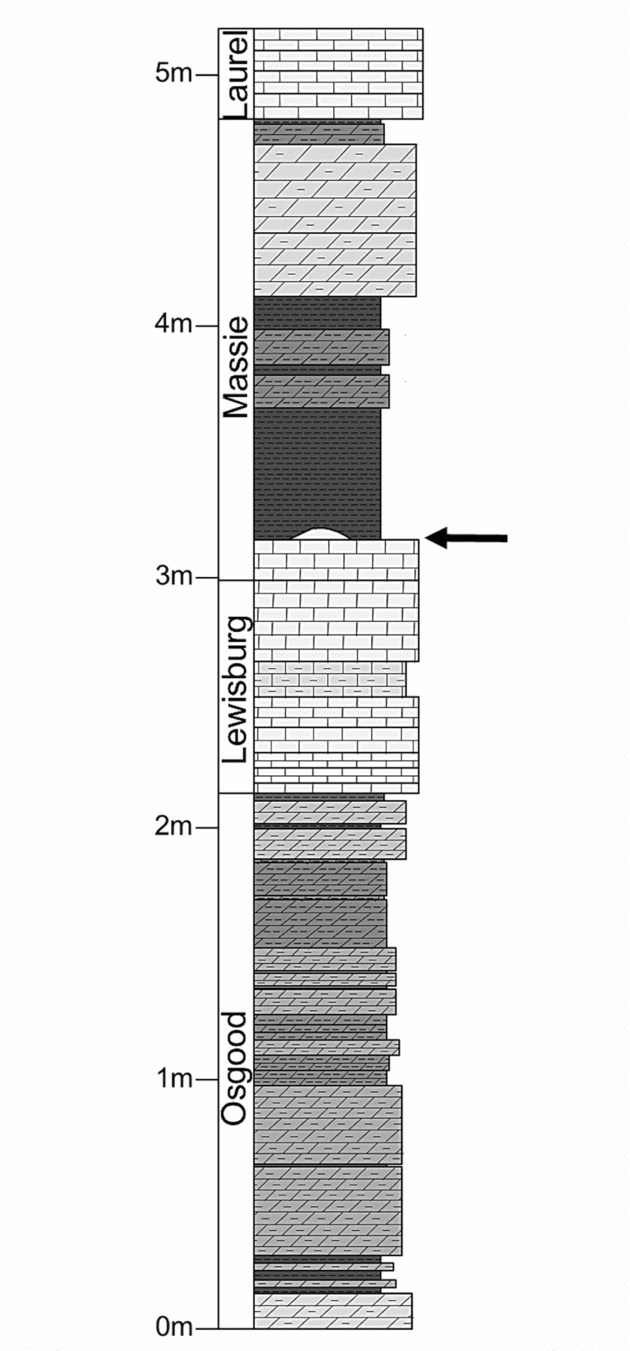
Historically, much of the stratigraphy exposed at the Napoleon quarry was considered part of a single unit, the Osgood Formation (Foerste, Reference Foerste1897) or Osgood Member of the Salamonie Dolomite (Pinsak & Shaver, Reference Pinsak and Shaver1964). The horizon under study is identified in older literature as that separating the ‘middle Osgood carbonate’ from the ‘upper Osgood shale’ (e.g. Frest, Strimple & Paul, Reference Frest, Strimple and Paul2011; Fig. 2). In light of recent regional lithostratigraphic revisions (Kleffner et al. Reference Kleffner, Cramer, Brett, Mikulic, Kluessendorf, Johnson, Sandy and Goldman2012; Brett et al. Reference Brett, Cramer, McLaughlin, Kleffner, Showers and Thomka2012), this surface is located at what is now considered the contact between the basal carbonate and overlying mudstone lithofacies of the Massie Formation (Silurian: Wenlock, Sheinwoodian; Fig. 2).
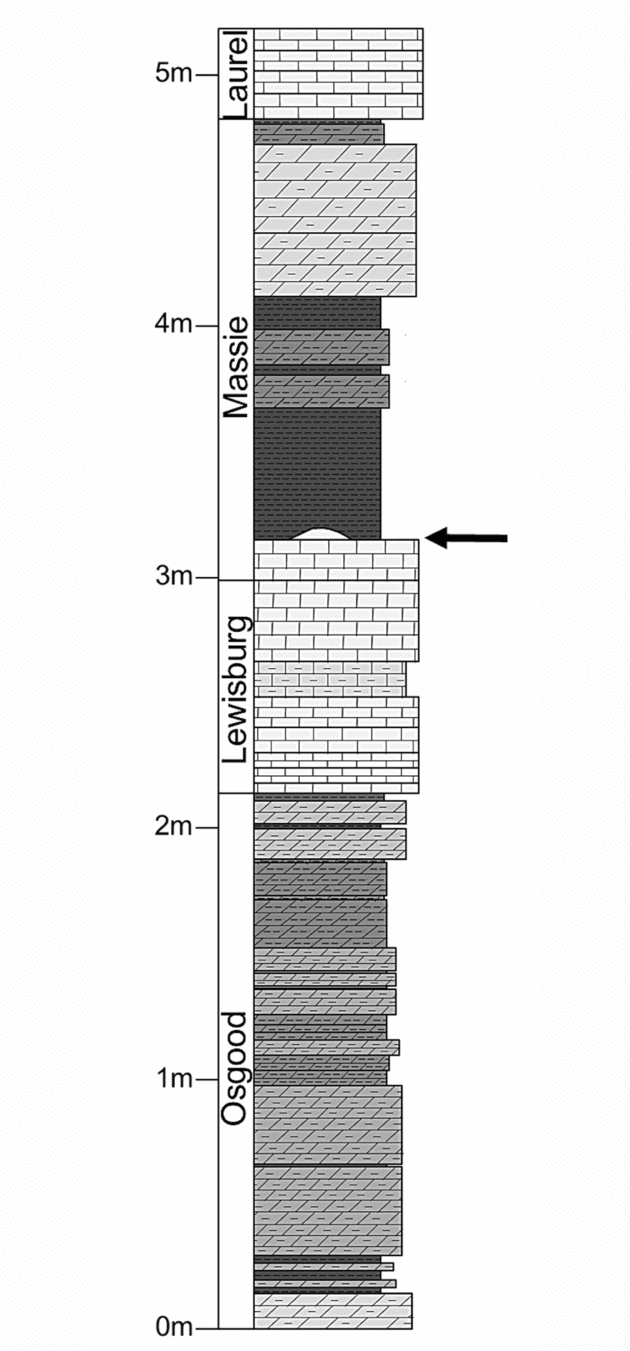
Figure 2. Stratigraphic column of a portion of strata exposed at the Napoleon quarry with formation-scale lithostratigraphic units labelled. The encrusted hardground is marked by the arrow. Note that the Brassfield Formation, underlying the Osgood Formation, is not depicted, nor is the full thickness of the Laurel Formation. Stratigraphic terminology follows Brett et al. (Reference Brett, Cramer, McLaughlin, Kleffner, Showers and Thomka2012). From Thomka & Brett (in press).
This contact represents a major flooding surface developed during an episode of rapid sea-level rise (McLaughlin et al. Reference McLaughlin, Brett, Wilson, Pratt and Holmden2008; Brett et al. Reference Brett, Cramer, McLaughlin, Kleffner, Showers and Thomka2012). The net effects were a shut-off of clastic sedimentation and stasis of redox boundaries in the shallow subsurface, allowing cementation of the seafloor (e.g. Brett, Reference Brett1995). Such flooding surfaces are consistent and predictable throughout the Northern Appalachian Basin (Brett, Goodman & LoDuca, Reference Brett, Goodman and LoDuca1990; Brett & Ray, Reference Brett and Ray2005; McLaughlin, Brett & Wilson, Reference McLaughlin, Cramer, Brett, Kleffner, Maria and Counts2008), and this specific surface is widely traceable and easily recognizable throughout the Cincinnati Arch region and Northern Appalachian Basin, where it is typically associated with small, biogenically produced build-ups (Brett, Goodman & LoDuca, Reference Brett, Goodman and LoDuca1990; McLaughlin et al. Reference McLaughlin, Brett, Wilson, Pratt and Holmden2008; Brett et al. Reference Brett, Cramer, McLaughlin, Kleffner, Showers and Thomka2012). These microbioherms (sensu Archer & Feldman, Reference Archer and Feldman1986) are present at the Napoleon quarry and are encrusted, along with the planar hardground surface from which the microbioherms themselves emanate, by pelmatozoan attachment structures (Fig. 2). Although diploporite encrustation of microbioherms at this horizon has been reported by Frest, Strimple & Paul (Reference Frest, Strimple and Paul2011), no diploporite thecal attachments have been discovered attached to microbioherm masses at the locality in this study (see Section 5.c below).
Lithologically, the encrusted surface is a skeletal packstone to grainstone composed primarily of comminuted echinoderm ossicles. This surface is immediately overlain by, and stands in stark contrast to, medium grey mudstones representing deposition during a sea-level highstand (Brett et al. Reference Brett, Cramer, McLaughlin, Kleffner, Showers and Thomka2012). The hardground is slightly undulatory owing to storm scouring prior to cementation (see also Palmer & Palmer, Reference Palmer and Palmer1977). Slightly elevated crests are characterized by smaller and better sorted bioclastic particles and a total absence of clastic mud, whereas slightly depressed troughs are coarser, moderately to poorly sorted, and contain a minor proportion of mud. Diploporite attachments are cemented only to the crests of the hardground (Thomka & Brett, in press).
3. Methods and materials
Attachment structures were studied in the field for state of completeness, nature and location of mineral overgrowths, relationships to other biogenic or physical structures, and presence and clarity of internal features. Critical specimens were removed from the hardground with a hand-held diamond-tipped saw and reposited at the Cincinnati Museum Center in Cincinnati, Ohio (repository numbers beginning with CMC are listed in appropriate figure captions). Many holdfasts, however, were left in place on the hardground to permit future studies. Holdfasts left in place were nevertheless inspected for taphonomic properties. Nearly all specimens displayed features well enough that they did not require extensive laboratory preparation or cleaning.
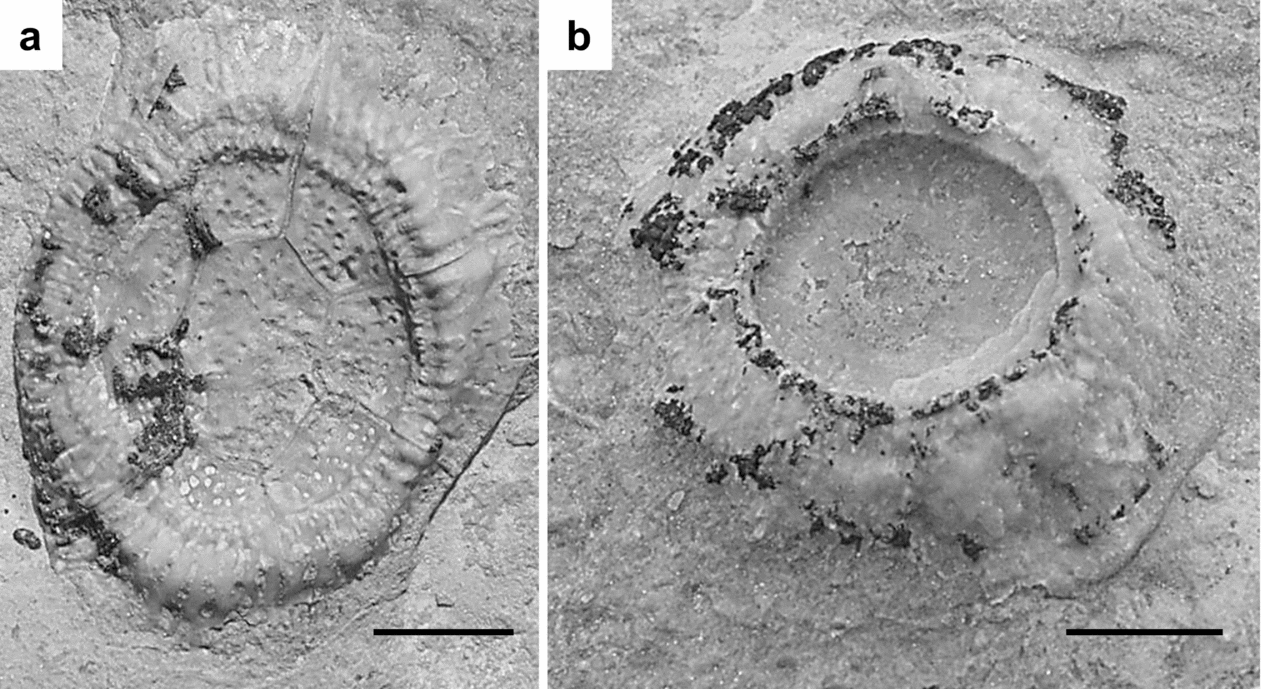
Work by Thomka & Brett (in press) showed that diploporite attachment structures from the Napoleon hardground are of two morphologies. Large, ovoid, relatively thin-walled structures composed of seven to eight pore-bearing plates surrounding a central heptagonal plate (Fig. 3a) have been attributed to the wide-based trematocystinid holocystitid Paulicystis. Smaller, circular, thick-walled structures composed of five to seven plates bearing pores and branching canals (Fig. 3b) were attributed to an undetermined diploporite, likely a pentacystinid or holocystinid holocystitid. Although Paulicystis holdfasts are less common than the smaller discoidal structures, sufficient numbers of specimens were discovered (over 60) to permit documentation of taphonomic patterns for both morphotypes.
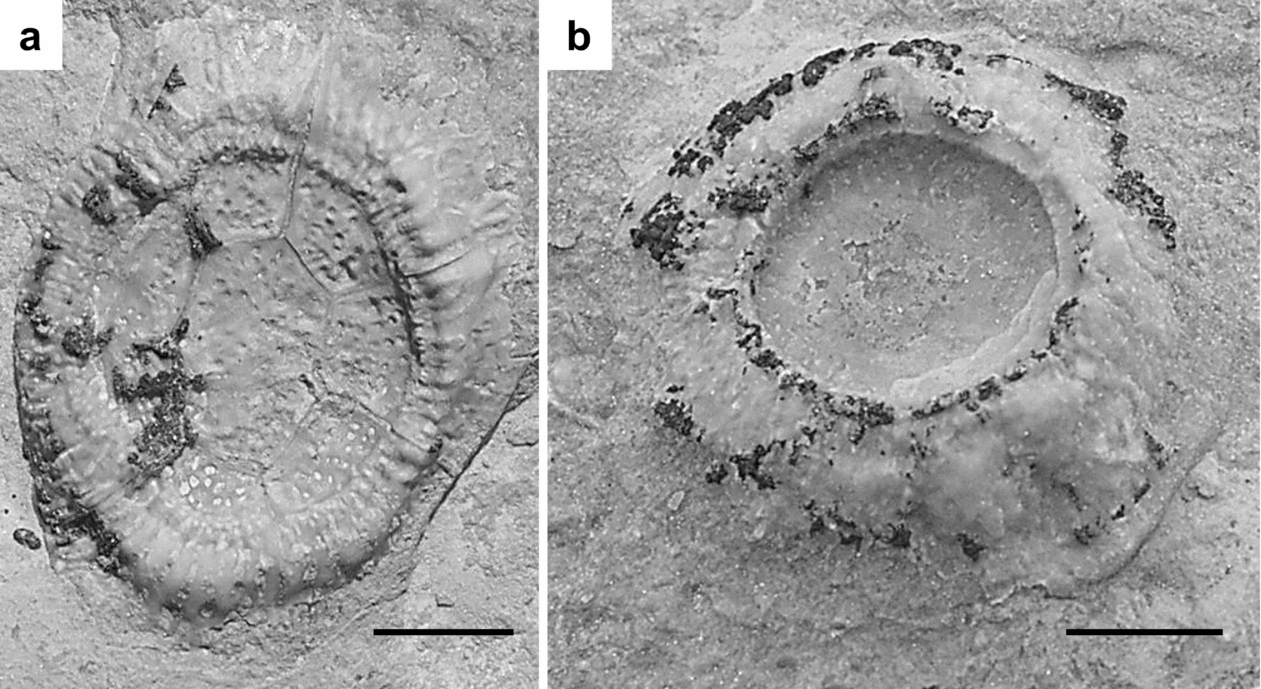
Figure 3. Well-preserved examples of the two diploporite attachment structure morphotypes present on the Napoleon hardground. (a) Thin-walled holdfast composed of seven marginal wall plates surrounding a central heptagonal plate, all pore-bearing (CMC IP 69526). This structure has been attributed to Paulicystis. (b) Thick-walled, multi-plated holdfast with radiating canals, pores and stereomic overgrowth of plate contacts (CMC IP 69527). The generic identity of these structures is currently unknown. Scale bars are 0.5 cm.
4. Results
4.a. General considerations
Documentation of taphonomic features allowed recognition of a spectrum of preservational states that is demonstrably not the result of ontogenetic or ecophenotypic variation. Ecophenotypic variation can largely be excluded because all studied structures are present on the same local substrate conditions on a single hardground surface. Thus, although local substrate-controlled variability has been documented within other co-occurring pelmatozoan holdfasts, these differences reflected morphological responses to local conditions associated with hardground crests, hardground troughs and microbioherms, respectively (Thomka & Brett, Reference Thomka and Brett2013). In contrast, diploporite attachment structures were exclusively on hardground crests (Thomka & Brett, in press). Ontogenetic variability can also be excluded because specimens with the same taphonomic characteristics span a considerable size range and, conversely, specimens of the same size occur in a variety of taphonomic states. Secondary overgrowth of plate sutures in the smaller, ovoid holdfasts is not an ontogenetic signature of mature individuals: clarity of plate contacts is a taphonomic feature and not indicative of an immature individual that did not yet generate secondary stereom to overgrow plate boundaries (Thomka & Brett, in press).
Taphonomic patterns are detailed below for both of the diploporite attachment structure morphotypes. Taphonomic processes are divided into two categories: biostratinomic processes refer to the physical, biological and chemical events that occurred during the interval in between the death of the organisms and final burial (in contrast to the more restricted definition of this term originally established by Seilacher, Reference Seilacher and Ginsburg1973); diagenetic processes refer to all events that occurred in between the final burial of an organism and its discovery as a fossil.
4.b. Biostratinomic disarticulation and degradation
None of the holdfasts on the hardground surface are attached to partial or complete thecae. This indicates that soon after death, the theca underwent rapid disarticulation and became detached from the aboral attachment structure, which was firmly cemented to the substrate. Rare partially articulated thecae were discovered on the hardground, but only in hardground troughs and never on the hardground crests where holdfasts are found; further, most identifiable thecae are attributable to Holocystites scutellatus, which lacked a permanent attachment (Fig. 4). While the theca, composed of numerous small ossicles, underwent rapid and complete skeletal disarticulation, associated attachment structures, despite also comprising multiple ossicles, remained more intact, owing to their cementation to the substrate. Hence, following the rapid detachment of thecae, holdfasts were subjected to in situ degradation.

Figure 4. Partially articulated theca of Holocystites scutellatus in a hardground trough (specimen uncollected, currently on hardground). These diploporites lack permanent attachments and are not affiliated with the holdfasts analysed in this study. The stubby stele is marked by the ‘s’ and the oral area is marked by the ‘o’. Scale bar is 1 cm.
For Paulicystis attachment structures, well-preserved specimens, representing individuals that underwent detachment of the thecae followed shortly by final burial, are characterized by the presence of clearly visible plates in the depressed central area of the holdfast, as shown in Figure 3a. The majority of specimens have evidence of continued degradation during exposure such that only the marginal wall remains identifiable but the interior is indistinct or eroded away completely (Fig. 5). Evidently the thinness of the depressed interior of these attachment structures (i.e. the interior of the stele sensu Brett, Reference Brett1981, Reference Brett, Bassett and Lawson1984) made this portion particularly prone to destruction relative to the thicker and more strongly sutured holdfast margin.

Figure 5. Thecal attachment structures of Paulicystis that experienced differing amounts of weathering prior to final burial. (a) Holdfast (CMC IP 69530) with eight clearly recognizable component plates comprising the margin, but a central cavity obscured by weathering; this represents a holdfast subjected to exposure intermediate between the specimens in Figure 3a and (b). (b) Moderately weathered holdfast (CMC IP 70214) with faint plate contacts, partially eroded margin, particularly on one side, and obscured and partially eroded central cavity; this represents a holdfast subjected to exposure intermediate between the specimens in (a) and (c). (c) Moderately weathered holdfast (CMC IP 70215) with faint plate contacts, strongly eroded margin with a simple ovoid outline and reduced height, and central cavity that was breached to expose the underlying substrate. (d) Severely weathered holdfast (CMC IP 70216) with faint plate contacts, preservation of only the tallest and thickest portions of the margin, and a central cavity that was breached to expose the underlying substrate. All scale bars are 1 cm.
Among the Paulicystis attachment structures that experienced sufficient weathering to remove or obscure holdfast interiors, a spectrum of preservational states can be recognized. Holdfasts subjected to minor weathering are characterized by a well-developed margin with visible plate contacts (Fig. 5a). The plates of holdfasts in this taphonomic state are always in close contact with each other. The thin, pore-bearing, parabolically depressed central area was almost immediately destroyed, being present only on a single specimen (Fig. 3a). In addition, the thin, slightly lobate, substrate-parallel outermost margin may be eroded away, giving the holdfast a simple ovoid outline, as shown in Figure 5a; however, some structures have been discovered that retain at least a partial thin margin and also display enhanced weathering of the central cavity and a thick, vertical marginal wall (Fig. 5b). Continued exposure results in obscured plate boundaries, giving attachment structures a massive appearance; progressive erosion of the elevated marginal wall, decreasing the vertical relief of structures; and erosion of the thin, lobate margins, giving specimens simple, narrower outlines (Fig. 5c). It is possible that erosion of the elevated margin occurs before the lobate margins, but most specimens have erosion at both sites. Specimens with evidence of having been subjected to physical weathering for the greatest duration are characterized by high marginal walls that were altered to produce lobate margins (Fig. 5d), although these reflect degradation of what was previously the highest and thickest portions of holdfasts; the lobate margins of well-preserved holdfasts are composed of thin, delicate skeletal material, whereas the lobate margins of poorly preserved holdfasts are remnants of robust portions of the organism. Interior cavities are always completely eroded away, and the height of marginal walls is decreased (Fig. 5d). In fact, attachment structures in this taphonomic state are only recognizable as low, oblong rings representing the remnants of what were formerly the highest parts of the marginal wall (Fig. 5d).
For specimens attributable to the unknown holocystitids (Fig. 3b), a similar biostratinomic progression can be established. The most well-preserved holdfasts are characterized by plate sutures that are not obvious, pores that are only clearly visible in the interior of depressed central cavities, and radiating canal structures that are not visible. Further, the thin outermost margin, in contact with the substrate, is intact and has a slightly lobate shape (Fig. 3b). Continued exposure results in erosion of the thin outermost margin, creating a slightly ragged or irregular margin that ends sharply at the thick, elevated marginal wall (Fig. 6b, c) and erosion of the thin, pore-bearing material at the bottom of the interior cavity, exposing the underlying substrate (Fig. 6a, b). Many specimens in this taphonomic state also display increased visibility of pores, particularly on the exteriors of holdfasts (Fig. 6a, c). A few specimens maintain a portion of the outermost margin around part of the holdfast, but it is typically dramatically thinned or absent in other portions of the holdfast (Fig. 6c). Moderately weathered holdfasts are characterized by complete erosion of the outermost margin, giving the structure a circular outline (Fig. 6d, f). Plate boundaries may be visible, reflecting degradation of secondary stereom overgrowths (Fig. 6d, e). Pores can become readily visible on the exterior of the structure as well as the interior, if any portion of the bottom of the interior cavity remains (Fig. 6e). The most prominent feature of moderately weathered diploporite attachment structures is the enhancement of branching, radiating canal structures (Fig. 6d, e), interpreted by Thomka & Brett (in press) as serving a sensory function. These structures, which resemble canaliculi (Moore, Jeffords & Miller, Reference Moore, Jeffords and Miller1968; see also Franzén-Bengtson, Reference Franzén-Bengtson1983), can become the dominant macroscopic feature of holdfasts in this taphonomic state (Fig. 6d). Continued degradation obscures all structural details except for occasional pores on the exterior of holdfasts. Attachment structures subjected to this degree of weathering are relatively thick, circular structures with indistinct plate contacts and few surface features (Fig. 6f).

Figure 6. Thecal attachment structures of undetermined holocystitid diploporites that experienced differing amounts of weathering prior to final burial. (a) Relatively well preserved, upward-tapering holdfast (CMC IP 69528) with a circular outline (reflecting removal of slightly lobate marginal material), moderately visible pores, faint plate contacts and canal structures, and a pyritized inner ring, and a deep, sediment-filled central cavity; this represents a holdfast subjected to exposure intermediate between the specimens in Figure 3b and (b). (b) Relatively well-preserved holdfast (CMC IP 70217) with poorly visible pores, faint plate contacts, a visible but un-pyritized inner ring and a formerly circular marginal outline that been slightly degraded; this represents a holdfast subjected to exposure intermediate between the specimens in (a), (c) and (d). (c) Moderately preserved holdfast (CMC IP 70218) with remnants of the lobate margin (heavily degraded and thinned on one side) and interior platform, but poorly visible pores and moderately visible plate contacts and canal structures; this represents a holdfast subjected to exposure intermediate between the specimens in (a) and (d). (d) Moderately to poorly preserved holdfast (CMC IP 70219) with a weathered margin and interior resulting in thinned, circular outlines to both, and prominent canal structures and moderately visible plate contacts and pores; this represents a holdfast subjected to exposure intermediate between the specimens in (b), (c) and (e). (e) Moderately to poorly preserved holdfast (CMC IP 70220) with a circular margin beginning to weather irregularly, no trace of an interior ring, and prominent canal structures, pores and plate contacts; this represents a holdfast subjected to exposure intermediate between the specimens in (d) and (f). (f) Poorly preserved holdfast (CMC IP 70221) with perfectly circular outline, no inner ring, faint canal structures, pores and plate contacts; this represents a holdfast subjected to exposure intermediate between the specimens in (e) and Figure 7a. All scale bars are 1 cm.
Holdfasts that were very strongly weathered prior to burial are difficult to identify as diploporite thecal attachment structures without comparison to more complete specimens. Examples are shown in Figure 7. Some attachment structures have minor separation between the upper portions of component plates. However, the cementation of the basal portions of these structures to the hardground surface was strong enough to prevent complete removal of plates; consequently, holdfasts remained in place even as plates slightly shifted away from each other at sutures (Fig. 7a). Some holdfasts are so worn that they are little more than calcareous ‘pads’ resembling isolated crinoid columnals (Fig. 7b). Others are characterized by greater vertical relief but display radiating canal structures so prominently that other features are obscured (Fig. 7c). Attachment structures in this state resemble the septate basal discs of Cleiocrinus described by Brett & Liddell (Reference Brett1978). Severely degraded diploporite holdfasts on the Napoleon hardground rarely permit distinction between the two holdfast morphotypes; however, the more circular shape of the majority of structures in this taphonomic state suggests that the producers were primarily undetermined holocystitids associated with the holdfast type depicted in Figure 3b. It is possible that the thinness of Paulicystis attachment structures relative to the other morphotype resulted in complete destruction of these holdfasts, whereas the thicker, presumably more robust, morphotype could experience greater degradation without obliteration.

Figure 7. Diploporite attachment structures that were strongly weathered prior to final burial. (a) Multi-plated holdfast (specimen uncollected) showing prominent pores; wide, interior cavity breached to expose underlying substrate; and separation between component plates. Scale bar is 1 cm. (b) Holdfast (CMC IP 70222) that is reduced to a low, calcareous pad with prominent canal structures and moderately visible plate contacts. Scale bar is 0.5 cm. (c) Barely recognizable holdfast (CMC IP 70223) preserved as a heavily pyritized disc. Scale bar is 0.5 cm.
4.c. Overgrowth relationships
Research on modern environments indicates that available space is a critical limiting factor in hard substrate communities (Dayton, Reference Dayton1971; Jackson et al. Reference Jackson, Goreau and Hartman1971; Jackson, Reference Jackson1977, Reference Jackson1979). Increasingly, evidence for competitive biotic interactions on Palaeozoic hardgrounds and encrusted skeletal elements has been documented, supporting the ecological paradigm established for the Recent. The Napoleon hardground provided several examples of biostratinomic processes involving overgrowth of diploporite attachment structures by other organisms and spatial relationships between diploporite attachment structures.
Laminar stenolaemate bryozoans overgrew diploporite holdfasts (Fig. 8a, b). The term ‘overgrew’ is used here in favour of ‘encrusted’ because bryozoan zoaria clearly began astogeny on the hardground crests and expanded onto the outer surface of holdfasts that were cemented to the same surface, rather than passively beginning growth on the holdfasts themselves. It is interesting to note that bryozoan colonies never overgrew the articular area of diploporite attachments and many appear truncated or broken, suggesting that the lower part of the theca remained articulated to the attachment structure and was likely overgrown itself, but subsequently detached through decay-induced thecal disarticulation. In all instances, bryozoans overgrew diploporite holdfasts, and no evidence for biotic response to overgrowth by diploporites has been detected. The consistent overgrowth of solitary organisms by colonial organisms supports relationships documented in Recent (e.g. Jackson, Reference Jackson1977) and Palaeozoic (Sprinkle & Rodgers, Reference Sprinkle and Rodgers2010) environments, wherein an adaptive advantage is bestowed on colonial taxa presumably because they have a faster growth rate.

Figure 8. Relationships between diploporite attachment structures and other encrusters on the Napoleon hardground. (a, b) Overgrowth of holdfasts by laminar stenolaemate bryozoan colonies (CMC IP 70224 and CMC IP 70225, respectively). (c, d) Diploporite holdfast overgrown by attachment structure of Caryocrinites (Rhombifera). Note that these Caryocrinites attachment structures are more conical and morphologically simple than the branching dendritic radices common to this genus, and that diploporite attachments are poorly preserved, comparable to the specimen in Figure 7b (CMC IP 70228 and CMC IP 70229, respectively). (e) Attachment structure encrusted by a small (juvenile crinoid?) holdfast with a round lumen (CMC IP 70226; at the approximate ‘3 o'clock’ position). (f) Two diploporite attachment structures in direct contact with each other. Note that the specimen on the left is better preserved than that on the right, and that the interaction between these two resulted in modification of the shape of both specimens from a strongly circular outline to a more polygonal outline (CMC IP 70227). All scale bars are 1 cm.
Diploporite holdfasts on hardground crests are also rarely overgrown by attachment structures of the rhombiferan Caryocrinites (Fig. 8c, d). The overgrowing holdfasts can be identified as those of Caryocrinites because of the presence of a distinctively large, trilobate lumen (Brett, Reference Brett1978 b). In all instances, the encrusted diploporite attachment structures represent the morphotype attributable to the undetermined holocystitids. Further, diploporite attachments encrusted by rhombiferan holdfasts are always quite poorly preserved. The encrusted holdfast in Figure 8c is little more than a septate pad (comparable to the taphonomic state of the specimen in Fig. 7b), and that in Figure 8d is a topographically irregular disc with an appearance dominated by canal structures. Interestingly, dendritic radices belonging to Caryocrinites as well as crinoid taxa with similar holdfast morphologies are otherwise absent on the hardground crests occupied by diploporites; evidently, Caryocrinites did not directly encrust the crests of the Napoleon hardground itself but, instead, preferentially overgrew select diploporite holdfasts (Thomka & Brett, Reference Thomka and Brett2013). Why this relationship existed, and why only degraded diploporite attachments that were exposed and weathered for a relatively long time were utilized as substrates remains enigmatic. One potential explanation is that some poorly preserved diploporite attachment structures served as emergent ‘islands’ of hard substrata protruding above a thin veneer of sediment deposited during the initial phases of burial of the hardground.
One specimen (CMC IP 70226) has encrustation of a fairly well-preserved diploporite attachment structure by a small holdfast of unknown affinities (Fig. 8e). The overgrowing structure is circular, contains a minute, centrally located, circular lumen, and does not display obvious radicles. This may represent overgrowth by a juvenile crinoid, possibly a small cladid or disparid, or a coronoid (Brett et al. Reference Brett, Frest, Sprinkle and Clement1983). Another slab (CMC IP 70227) recovered from the Napoleon hardground contains two diploporite attachment structures in direct contact with each other (Fig. 8f). This pair of holdfasts is significant because it may preserve evidence for spatial competition: the typically circular outlines of attachment structures were modified to more polygonal shapes along the hemisphere where the specimens meet. Such outline modifications have been documented in edrioasteroids (Meyer, Reference Meyer1990; Sumrall, Sprinkle & Bonem, Reference Sumrall, Sprinkle and Bonem2006) and holdfasts of the crinoid Anomalocrinus (Brett, Deline & McLaughlin, Reference Brett, Deline, McLaughlin, Ausich and Webster2008) that grew in crowded conditions. This is a common phenomenon among sessile non-colonial invertebrates. It is interesting to note that one of the specimens is better preserved than the other and that the better preserved holdfast displays the most pronounced outline flattening (Fig. 8f, specimen on left). It is also possible that, rather than reflecting competition, the more degraded specimen may actually represent the isolated holdfast of a dead diploporite that affected the growth of a living individual, represented by the less degraded specimen. Further support for this scenario comes from the slightly lobate margins of the less degraded specimen, which overgrow the more degraded holdfast to a minor extent (Fig. 8f). This relationship might represent a form of gregarious behaviour similar to that proposed for clustered crinoid attachment structures described by Donovan, Harper & Håkansson (Reference Donovan, Harper and Håkansson2007) and Donovan & Harper (Reference Donovan and Lewis2010).
Although several types of overgrowth interactions can be documented on the Napoleon hardground and its diploporite attachment structure population, it is not possible to construct a competitive hierarchy of encrusters, as is common in modern ecological studies (e.g. Jackson & Buss, Reference Jackson and Buss1975; Connell, Reference Connell and Mackie1976; Jackson, Reference Jackson1979; see also McKinney, Reference McKinney1995). This is owing to the low number of observed interactions and the paucity of observations on overgrowth relationships between the various organisms that encrusted diploporite holdfasts. It does appear, however, that diploporite attachment structures formed the bottom of any competitive network, as they did not overgrow other organisms and might not have even been able to completely overgrow isolated holdfasts representing dead individuals.
4.d. Diagenetic processes
Nearly all specimens have pyrite crystals on the external surface of the holdfast margin. Pyrite mineralization is a common feature throughout the Appalachian Foreland Basin during this interval, in contrast to slightly older deposits, which tend to preserve iron in different phases (McLaughlin, Emsbo & Brett, Reference McLaughlin, Emsbo and Brett2012). In some cases, pyrite is amorphous, lacking obvious crystal habit; in others, the pyrite is macrocrystalline, in a cubic form. Morphologic variability in diagenetic pyrite is common in deposits containing abundant pyrite, and the distribution of pyrite crystal morphologies has been documented in detail by several case studies (e.g. Hudson, Reference Hudson1982; Canfield & Raiswell, Reference Canfield, Raiswell, Allison and Briggs1991).
Pyrite can be distributed seemingly randomly over holdfasts, but certain specimens have preferential pyritization of the slightly depressed innermost marginal wall (Fig. 6a) to form an ‘interior ring’ of pyrite (Thomka & Brett, in press). The restriction of pyrite to interior rings suggests early diagenetic processes associated with a chemically reactive microenvironment (sensu lato; see McNamara et al. Reference McNamara, Orr, Kearns, Alcalá, Anadón and Mollá2009) generated by decay of soft tissues. This assertion is supported by pyritization of canal structures in radicles of the dendritic radicular attachment structures attributable to co-occurring rhombiferans and crinoids.
Interestingly, the large size of many pyrite crystals suggests a late diagenetic origin that, at first glance, appears to contradict the evidence for early diagenesis discussed above. This most likely reflects late diagenetic pyrite growth that was seeded by early diagenetic crystallites, which developed in decay-induced microenvironments (see also Canfield & Raiswell, Reference Canfield, Raiswell, Allison and Briggs1991; Thomka & Lewis, in press). Because much diagenetic mineralization is surface reaction-controlled, pyrite that formed very rapidly after death, even if amorphous or microcrystalline, can serve as a locus for later mineral growth as local redox reactions liberated ions that were rapidly incorporated into minerals that then nucleated on the antecedent crystallites. Hence, even though macrocrystalline pyrite reflects late diagenetic processes, its distribution provides insight into sites of early diagenesis which, in turn, likely indicate locations of reducing microenvironments.
A large body of literature exists on early diagenetic minerals and their relationship to preservation of unmineralized tissues (e.g. Allison, Reference Allison1988; Allison & Briggs, Reference Allison, Briggs, Allison and Briggs1991; Briggs, Reference Briggs2003), and it is beyond the scope of this study to provide an extensive review of this subject. Nevertheless, it is well documented that decay of connective tissues can lead to mineralization, essentially preserving a guide to the distribution of material that was lost relatively early in the taphonomic history of specimens. Preservation of soft tissues in Palaeozoic echinoderms has been previously documented by Glass & Blake (Reference Glass and Blake2004), Glass (Reference Glass2006), Kammer & Ausich (Reference Kammer and Ausich2007) and Ausich, Bartels & Kammer (Reference Ausich, Bartels and Kammer2013), among others. To be clear, we do not suggest that soft tissues are preserved on the Napoleon hardground fauna, but the non-random distribution of pyrite on the diploporite holdfasts studied here matches the location of where connective tissue would be expected, and the decay of these presumed tissues could easily have led to mineral growth. It is unclear at this point as to whether these connective tissues were ligamentary or muscular in composition, but it seems more likely that ligaments were used to simply connect plates in the aboral portion of the theca to plates comprising holdfasts.
5. Discussion: palaeoecologic implications
5.a. Taphonomy of attachment structures
Although much published data are available on echinoderm taphonomy (see summaries by Lewis, Reference Lewis, Broadhead and Waters1980; Donovan, Reference Donovan and Donovan1991; Brett, Moffat & Taylor, Reference Brett, Moffat, Taylor, Maples and Waters1997; Ausich, Reference Ausich, Jangoux and Lawrence2001; Nebelsick, Reference Nebelsick, Heinzeller and Nebelsick2004), few studies to date have addressed, much less focused on, taphonomic patterns among pelmatozoan attachment structures. Notable exceptions include studies of Ordovician holdfasts by Brett & Liddell (Reference Brett and Liddell1978) and Brett, Deline & McLaughlin (Reference Brett, Deline, McLaughlin, Ausich and Webster2008). Both of these studies documented morphologic features that became visible as a result of weathering, in strong agreement with the results of the current study. A network of radiating, branching canals nearly identical to those in diploporite holdfasts from the Napoleon hardground was brought out by weathering in the terminal holdfasts of the Ordovician crinoids Anomalocrinus (Brett, Deline & McLaughlin, Reference Brett, Deline, McLaughlin, Ausich and Webster2008) and ?Carabocrinus (Lewis, Reference Lewis and Sprinkle1982). In addition, weathering brings out pore structures in the holdfast of the Ordovician crinoid Cleiocrinus (Brett & Liddell, Reference Brett and Liddell1978) and the holdfast morphogenus Podolithus (Conkin, Reference Conkin2003), as well as unidentified discoidal attachments from the Silurian of Gotland, Sweden (Franzén, Reference Franzén1977). The multi-plated nature of the integument of Cleiocrinus was also revealed through weathering (Brett & Liddell, Reference Brett and Liddell1978).
Whereas it may be somewhat counterintuitive that pristine, unweathered specimens are often less capable of clearly showing morphologic features than are imperfectly preserved specimens, it is logical when considering pelmatozoan echinoderm attachment structures. The final configuration of attachment structures is dictated by three factors: (1) relationship to column or stele (i.e. is the attachment simply a modified portion of the pelma or is it a specialized, unique terminal ossicle or multi-plated structure?); (2) capacity to overgrow primary skeletal structures with secondary stereom (i.e. does the ultimate attachment closely correspond to distal skeletal morphology or is it concealed by amorphous skeletal overgrowth?); and (3) substrate consistency (i.e. is the substrate stable, permitting direct attachment by a simple structure or is it unstable, requiring increased surface area for greater purchase?). More detailed considerations of the relationships between pelmatozoans and substrate are discussed by Brett (Reference Brett, Bassett and Lawson1984, Reference Brett, Bassett, Lane and Edwards1991) and Seilacher & MacClintock (Reference Seilacher and MacClintock2005); but in the context of this discussion, it is apparent that certain palaeoenvironmental conditions can trigger minor to substantial modification via secondary stereom secretion. Hence, originally multi-plated or otherwise skeletally complex structures may appear simple or undifferentiated while the organism was alive or shortly after death. Nevertheless, minor differences in density between stereomic overgrowths, underlying solid primary skeletal structures, and underlying multi-plated primary skeletal structures are highlighted by weathering during exposure. Future research into pelmatozoan autecology is likely to benefit significantly from detailed analysis of attachment structures in various states of completeness and displaying various extents of degradation. Pristine specimens may prove most useful for initial taxonomic identification, but taphonomic evaluation will highlight palaeoecologically significant structural frameworks and skeletal modifications.
The variation in taphonomic states within this assemblage of attachment structures reflects within-habitat time-averaging (Kidwell, Reference Kidwell, Kidwell and Behrensmeyer1993), as expected given the inferred slow sedimentation rate. The consequent ‘time-richness’ of this fauna makes such assemblages ideal for providing insights into intrinsic factors that influence the taphonomy of a given taxon, as an entire preservational spectrum can be documented from a single surface with few interpretive complications arising from preservation in different taphofacies (see Brett & Baird, Reference Brett and Baird1986). Documentation of taphonomic properties of specimens at low taxonomic levels among an assemblage recovered from the same general taphofacies is critical to understanding the magnitude of taphonomic variability that can potentially be expressed by echinoderm skeletal modules (e.g. holdfasts, thecae, columns; Allison, Reference Allison1990; Thomka et al. Reference Thomka, Lewis, Mosher, Pabian and Holterhoff2011, Reference Thomka, Mosher, Lewis and Pabian2012). Such knowledge is important in recognition and identification of specimens in settings where ideal or complete preservation is uncommon.
5.b. Significance of diagenetic fossil colour
A number of recent publications have documented colour variation among fossil echinoderms, resulting in recognition of original, biologically meaningful organic molecules (Wolkenstein et al. Reference Wolkenstein, Gross, Heinz and Schöler2006, Reference Wolkenstein, Głuchowski, Gross and Marynowski2008; O'Malley, Ausich & Chin, Reference O'Malley, Ausich, Chin, Ausich and Webster2008, Reference O'Malley, Ausich and Chin2013; Niedźwiedzki, Salamon & Wolkenstein, Reference Niedźwiedzki, Salamon and Wolkenstein2011). Preservation of biomarkers in ancient organisms requires exceptional preservation, and isolated attachment structures showing evidence of varying extents of exposure and degradation are unlikely to record primary organic signatures. The holdfasts studied here display colour variations, but rather than reflecting the presence of taxon-specific organic molecules, this phenomenon represents diagenetic colouration. Recently, Kolbe et al. (Reference Kolbe, Zambito, Brett, Wise and Wilson2011) studied Palaeozoic brachiopod shells and documented consistent relationships between diagenetic fossil colour and taphonomic state. Macrofossils on the Napoleon hardground do not display the variety of colouration evident within the brachiopod dataset of Kolbe et al. (Reference Kolbe, Zambito, Brett, Wise and Wilson2011), but consistent relationships, corroborated by other taphonomic criteria, can be recognized in the holdfast population studied here.
Well-preserved attachment structures are nearly always whitish to light grey, and poorly preserved specimens become increasingly dark (e.g. Fig. 6a–e). This relationship is consistently true for diploporites, but is also well illustrated by the colour difference between the well-preserved Caryocrinites holdfasts that encrusted remnants of diploporite attachment structures in Figures 8c, d. Interestingly, the correlation between increasing darkness and degradation persists only when considering moderately preserved specimens: holdfasts that are particularly incomplete display relatively light colours (Figs 5d, 8f).
Results on diploporite holdfast colouration, although preliminary and not subjected to quantitative analysis, agree with the results of Kolbe et al. (Reference Kolbe, Zambito, Brett, Wise and Wilson2011), who found a similar darkening of brachiopod shells in association with evidence for prolonged exposure. This pattern was interpreted as being related to increased sulfide mineralization, an explanation that also seems appropriate for the attachment structures studied here. Although documentation of colour variation was not the primary objective of this study, colour nevertheless provided an additional line of evidence for assessing degree of degradation. The fact that darkening is consistent with physical signs of increased exposure corroborates interpretations made in Section 4.b. Finally, it is hoped that inclusion of data on diagenetic fossil colour here, although only in the form of relatively simple observations, will encourage incorporation of diagenetic colour data along with other taphonomic indicators in future analyses of ancient deposits.
5.c. Encrustation of microbioherms
Although diploporite holdfasts are common on the Napoleon microbiohermal hardground, the fistuliporoid bryozoan-dominated microbioherms themselves are not encrusted by diploporites. Instead, microbioherms served as substrates for long-stemmed pelmatozoans: attachment structures attributable to the monobathrid camerate crinoid Eucalyptocrinites and the hemicosmitid rhombiferan Caryocrinites are quite abundant (Thomka & Brett, Reference Thomka and Brett2013). Interestingly, these same taxa, as well as other long-stemmed crinoids such as Periechocrinus and Dimerocrinites (Brett, Reference Brett, Bassett and Lawson1984), occur in hardground troughs. The occurrence of pelmatozoans with the longest columns, and therefore the greatest capability to elevate thecae well above the substrate, on top of microbioherms is counterintuitive, as these taxa could easily access higher currents while occupying relatively low areas (such as the hardground troughs where they are also common). Diploporites lack a true column and are incapable of elevating themselves more than a few centimetres above the substrate: one would logically expect the positive topographic relief of microbioherms to be the ideal substrate for diploporite encrustation. Instead, they are restricted only to hardground crests, which are elevated relative to hardground troughs, but are nevertheless considerably lower than microbioherms.
The reason for this segregation of attachment structures is currently unclear, but Thomka & Brett (Reference Thomka and Brett2013) discussed some potential explanations. It is possible that diploporites have particularly poor larval dispersal abilities and, consequently, that larvae simply could not become established atop microbioherms. This seems unlikely, however, as diploporite thecal attachment structures are occasionally present on hardground crests immediately adjacent to microbioherms. Another unlikely explanation for the absence of diploporites on Napoleon microbioherms and the extreme secondary thickening of other holdfasts (Fig. 9) is protection against anoxic, corrosive or otherwise toxic pore waters, as described by Seilacher & MacClintock (Reference Seilacher and MacClintock2005). The diverse benthic fauna and lack of evidence for increased organic matter concentration at the microbiohermal surface supports the interpretation of an oxygenated hardground.

Figure 9. Holdfast of Eucalyptocrinites (Crinoidea) on microbioherm, displaying extreme swelling by growth of secondary stereom. L – distorted lumen; S – secondary stereomic overgrowth. The exact reasons for this skeletal modification, which is also observed on the attachment structures of the rhombiferan Caryocrinites, are unknown. Scale bar is 1 cm
A potential cause that is more plausible, but hard to test, involves antagonistic interactions between encrusting pelmatozoans and components (microbes?) of the microbioherm. Because diploporites are thecally attached (Brett, Reference Brett, Bassett and Lawson1984), their steles, filled with viscera, would have been close to the substrate, and any adverse effects resulting from interactions with microbioherm biota or sediment would have been potentially dangerous to diploporites. In contrast, crinoid and rhombiferan holdfasts were far removed from thecae, and the narrow axial canals of distal columns belonging to encrusting taxa would have contained only a minimal amount of viscera. Further, radicles of Caryocrinites are composed of solid, undifferentiated stereom (Brett, Reference Brett1978 b); and those of Eucalyptocrinites, although composed of true cirral ossicles, are overgrown and thickened by rigid secondary stereom. This would have been quite effective in protecting viscera within columns or dististelar radicles from underlying or surrounding sediment.
An additional potential explanation for the absence of diploporites on microbioherms is that encrustation of the outer surfaces of microbioherms by long-stemmed pelmatozoans occurred after the hardground surface was buried by a thin layer of clastic sediment. If true, then the diploporites surrounding the topographically positive microbioherms would have been adapted for soft substrata, lacking a permanent attachment structure. In this scenario, diploporite encrustation would have occurred earlier in the growth history of microbioherms. Therefore, future work will focus on searching for diploporite attachment structures on interior laminae of cut microbioherms from the Napoleon hardground.
5.d. Bioerosion and embedment structures
Borings preserved in hard substrates have a long geologic record, and the Ordovician–Devonian is considered an interval of intensified bioerosion, both in terms of diversity of bioeroding organisms and variety of bored substrates (Warme, Reference Warme and Frey1975; Radtke, Hofmann & Golubic, Reference Radtke, Hofmann and Golubic1997; Taylor & Wilson, Reference Taylor and Wilson2003). Therefore, it is interesting to note the complete absence of borings into the Napoleon hardground. The undulating hardground is not bored, but is instead characterized by abundant firmground Thalassinoides burrows (see Bromley, Reference Bromley and Frey1975) presumably produced by trilobites (Cherns, Wheeley & Caris, Reference Cherns, Wheeley and Caris2006). These firmground burrows pre-date encrustation by diploporites, as evidenced by attachment structures that conform to burrow microtopography as well as an absence of Thalassinoides that cross-cut holdfasts. Hence, the surface must have been cemented enough to allow encrustation and oxygenated enough to support a diverse epibenthic fauna, but somehow it lacked organisms that bored into the hardground to produce domiciles.
One might suggest that the Napoleon surface lacked borings because it was never fully cemented, making it a firmground. Encrustation of firmgrounds by discoidal echinoderm holdfasts is known (Zamora et al. Reference Zamora, Clausen, Álvaro and Smith2010), but the surface studied here is densely encrusted – not only by pelmatozoans, but by laminar bryozoans and corals as well – and is heavily mineralized. These are characteristics of hardgrounds – commonly with borings – in the lower Palaeozoic of the Appalachian Foreland Basin and are not typical in firmgrounds (McLaughlin, Brett & Wilson, Reference McLaughlin, Brett, Wilson, Pratt and Holmden2008). It is far more likely that the surface under study here is a true hardground. Alternatively, it might be suggested that a veneer of some sort, possibly a microbial mat, might have developed on top of the hardground surface, preventing borings (e.g. Cornell, Brett & Sumrall, Reference Cornell, Brett and Sumrall2003). This also seems unlikely given the microbioherms that encrust the same horizon, the absence of diagenetic evidence for organic coatings on the hardground (i.e. pyritic or carbonaceous films), and the local substrate exposed beneath diploporite holdfasts subjected to weathering, which is identical to the surrounding, exposed hardground surface. The absence of borings into hardground troughs is likely owing to thin burial of these topographic lows by sediment washed off hardground crests and microbioherms. Ongoing research may reveal why much of the Napoleon hardground was not a suitable substrate for domichnial borings, but it may simply be a peculiarity without a detectable cause.
In contrast to the hardground surface, bioerosion structures are present on skeletal material associated with the hardground and microbioherms. The embedment structure Tremichnus (sensu Brett, Reference Brett1985; but see Pickerill & Donovan, Reference Pickerill and Donovan1998; Donovan & Pickerill, Reference Donovan and Pickerill2002) is common on crinoids and diploporites from this locality (Fig. 10a, e), and often associated with severe swelling (Fig. 10e), indicating that the structures were produced while the echinoderm host was still alive. Interestingly, no holdfasts have been discovered with swollen borings, suggesting that attachment structures were not sites of parasitism, whereas thecae were infested. The palaeoecology of these infestations is relatively well-studied and their occurrence is known to be host-specific (Frest, Mikulic & Paul, Reference Frest, Mikulic and Paul1977; Brett, Reference Brett1978 a, Reference Brett1985; Eckert, Reference Eckert1988; Widdison, Reference Widdison and Barker2001; Donovan & Lewis, Reference Donovan and Lewis2010; Frest, Strimple & Paul, Reference Frest, Strimple and Paul2011); therefore, it is possible that the diploporites affiliated with these attachment structures were not hosts.

Figure 10. Bioerosion and embedment structures on skeletal material from the Napoleon site. (a) Cirriferous crinoid column, slightly swollen in response to the embedment structure Tremichnus. Scale bar is 1 cm. (b) Large pluricolumnal (Periechocrinus?) with numerous small, unswollen Trypanites borings. Scale bar is 0.25 cm. (c) Small, vertically oriented column with several very small, unswollen Trypanites borings. Scale bar is 0.25 cm. (d) Calyx of Eucalyptocrinites displaying numerous Trypanites borings. Scale bar is 1 cm. (e) Articulated diploporite theca (Holocystites sp. cf. H. clavus) with numerous examples of the swollen embedment structure Tremichnus cystoidiphilus. Scale bar is 1 cm. Specimens in (d) and (e) are in the University of Cincinnati Geology Department Teaching Collection.
The simple boring Trypanites is moderately common, occurring almost exclusively on crinoid columns (Fig. 10b, c) and, less commonly, calyxes (Fig. 10d). No Trypanites have been discovered in isolated diploporite thecal plates, partially articulated thecae or attachment structures. The preferential excavation of domichnial borings in crinoidal material over diploporite material does not have an obvious explanation; perhaps the somewhat thinner, porous cystoid plates were unsuitable, whereas the thick, less porous and particularly abundant crinoid columns were substrates capable of being bored. In studies of Palaeozoic borings produced by acrothoracic barnacles, Rodda & Fisher (Reference Rodda and Fisher1962) and Ettensohn (Reference Ettensohn1978) documented that borings were preferentially, if not exclusively, present in the thickest skeletal material available, logically concluding that boring organisms favour thick skeletal substrates for production of domichnial bioerosion structures. Although the producer of Trypanites is most likely a sipunculid worm and certainly not a barnacle, the ecological relationship inferred above is not taxon-specific. Biostratinomic evidence described in Section 4.b above demonstrated that at least some attachment structures remained exposed on the hardground for extended periods of time; this would have made such thick, calcareous masses ideal substrates for boring even if the hardground surface itself was an unsuitable substrate for some reason. Holdfasts were overgrown in some cases by bryozoans and other pelmatozoans (Fig. 8), indicating that these structures were suitable substrates for some organisms, but apparently not the producers of Trypanites.
6. Discussion: stratigraphic implications
Hardground surfaces are distributed non-randomly throughout the stratigraphic record (Brett, Reference Brett1988, Reference Brett1995; Wilson & Palmer, Reference Wilson and Palmer1992; for more specific examples from the Appalachian Basin see McLaughlin, Brett & Wilson, Reference McLaughlin, Cramer, Brett, Kleffner, Maria and Counts2008; McLaughlin et al. Reference McLaughlin, Brett, Wilson, Pratt and Holmden2008). A number of independent lines of evidence contribute to the conclusion that the encrusted hardground studied here is genetically linked to a rapid change in base level that led to ecologic, environmental and geochemical conditions favouring incipient sea-floor cementation in low turbidity bottom water (see discussion in Section 2 above). This unique combination of physical parameters made possible the development of a distinctive biofacies and taphofacies reflecting sediment starvation. Because of the distinctive sedimentary conditions established during colonization of the Napoleon hardground, this surface can be used as an important stratigraphic marker for regional correlation (Brett, Goodman & LoDuca, Reference Brett, Goodman and LoDuca1990; Brett & Ray, Reference Brett and Ray2005; Brett et al. Reference Brett, Cramer, McLaughlin, Kleffner, Showers and Thomka2012) and, more importantly, the occurrence of similar surfaces and biotas can be predicted at analogous positions within other third-order (and potentially smaller scale) sequences in the Silurian of North America and elsewhere.
The potential for prediction of comparable faunas at major flooding/starvation surfaces may lead to significant new discoveries of middle Silurian diploporite taxa, although preservation as articulated thecae is unlikely given inferred palaeoenvironmental parameters (e.g. Brett & Baird, Reference Brett and Baird1986; Brett, Moffat & Taylor, Reference Brett, Moffat, Taylor, Maples and Waters1997). Holocystitid diploporites are known from coeval facies in the Romeo Member of the Joliet Formation in northeastern Illinois and southeastern Wisconsin (Frest, Mikulic & Paul, Reference Brett, Moffat, Taylor, Maples and Waters1977; Frest, Brett & Witzke, Reference Frest, Brett, Witzke, Boucot and Lawson1999; Frest, Strimple & Paul, Reference Frest, Strimple and Paul2011; D. G. Mikulic, pers. comm. 2012), considerably upramp from the deposits studied here and quite disparate lithologically. In such settings, diploporites and associated pelmatozoans are typically abundant only in direct association with microbiohermal build-ups (e.g. Frest, Brett & Witzke, Reference Frest, Brett, Witzke, Boucot and Lawson1999). Correlations between strata of the Cincinnati Arch region and the rim of the Michigan Basin (i.e. Illinois and Wisconsin) have historically been complicated, but the occurrence of diploporites and their encrusting attachment structures on hard substrata appears to have at least regional biostratigraphic significance (see also Frest, Mikulic & Paul, Reference Frest, Mikulic and Paul1977; Frest, Brett & Witzke, Reference Frest, Brett, Witzke, Boucot and Lawson1999; Frest, Strimple & Paul, Reference Frest, Strimple and Paul2011).
Microbioherms and a relatively diverse pelmatozoan fauna are also well documented at the contact between the Laurel Limestone and Waldron Shale (Wenlock: Homerian) in the Cincinnati Arch region (Foerste, Reference Foerste1898; Kindle & Barnett, Reference Kindle and Barnett1909; Halleck, Reference Halleck1973; Frest, Reference Frest1975; Ausich, Reference Ausich1975; Archer & Feldman, Reference Archer and Feldman1986; Frest, Brett & Witzke, Reference Frest, Brett, Witzke, Boucot and Lawson1999), a surface that is known to represent a hardground in nearly all localities where it is exposed (Halleck, Reference Halleck1973; McLaughlin et al. Reference McLaughlin, Brett, Wilson, Pratt and Holmden2008). The consistent presence of microbioherms at a major flooding/starvation surface that is slightly younger than, but at an exactly analogous position to, the hardground in the lower Massie Formation lends credence to the stratigraphic interpretation of the Napoleon quarry and the conditions responsible for genesis of hardgrounds in both sequences. In both the Sheinwoodian and the Homerian, rapid base level rise resulted in sediment starvation and the dramatic transition from carbonate to mudrock lithologies, separated by a hardground that serves as a substrate for a unique assemblage of encrusting and build-up-producing taxa. This contact, in spite of obfuscation of surface details by dolomitization (e.g. McLaughlin et al. Reference McLaughlin, Brett, Wilson, Pratt and Holmden2008), represents an excellent prospect for future discoveries of rare Silurian echinoderms, including diploporites, which become significantly rarer faunal elements above the Massie Formation. By late Sheinwoodian–Homerian time, holocystitid diploporites are thought to be restricted in distribution to shallow-water deposits in western Ohio and around the southern rim of the Michigan Basin (Frest, Mikulic & Paul, Reference Frest, Mikulic and Paul1977; Frest, Strimple & Paul, Reference Frest, Strimple and Paul2011); concentrated collection at the Laurel–Waldron contact, however, may very well reveal their persistence in deeper settings in the Cincinnati Arch region.
7. Conclusions
Thecal attachment structures attributable to diploporite echinoderms are fairly common on a hardground, representing sediment starvation during a major sea-level highstand, within the Silurian (Wenlock) Massie Formation in Ripley County, southeastern Indiana, United States. Observation reveals that these attachment structures do not display uniform preservation with regard to visibility of plate contacts and internal morphologic features, sites of early diagenetic mineralization and relationships to encrusting organisms.
Well-preserved specimens appear solid, with poorly visible plate contacts, pores and canals, as well as a pore-bearing platform flooring the depressed interior of the structure. Pre-burial weathering resulted in the enhancement of pores, canals and plate contacts; increased weathering resulted in removal of the platform floor to expose underlying sediment. Particularly poorly preserved specimens display separation of component plates, though plates remain in association and cemented to the hardground.
Few attachment structures are encrusted by other organisms, but laminar bryozoans occasionally overgrew the upper surface of holdfasts following detachment of the thecae. Degraded holdfasts were encrusted by the attachments of Caryocrinites. No diploporite holdfasts are bored; the exact reasons for this are unclear.
Nearly all specimens have macrocrystalline pyrite on their upper and lateral surfaces. Some specimens have preferentially pyritized ‘inner rings’, and co-occurring dendritic attachment structures contain pyritized lumens. Although the large size of pyrite crystals suggests a late diagenetic origin, the distribution of pyrite may indicate late diagenetic growth of pyrite on early diagenetic precursor crystallites that formed through decay of ligamentary tissue.
In a larger sense, taphonomic analysis of pelmatozoan attachment structures has great potential for providing insights into sedimentary dynamics, palaeoautecology and biotic interactions and simultaneously serving as a tool for comparative taphonomic analysis and stratigraphic correlation. The occurrence of time-averaged holdfast populations on major flooding/starvation surfaces can be predicted using an approach that integrates sequence stratigraphy and taphofacies analysis. Such surfaces can be prospected for rare taxa and faunal occurrences that are palaeoecologically and stratigraphically significant.
Acknowledgements
This research was funded by a Dry Dredgers Paul Sanders Award to JRT. The owners and management of the New Point Stone quarry near Napoleon, Indiana graciously allowed access to the study site. Tom Bantel (Dry Dredgers) introduced the hardground surface to the authors and Don Bissett (Dry Dredgers) collected the reposited specimens. Gary Motz (University of Cincinnati) assisted with photography. Earlier versions of this article were improved by the constructive reviews provided by two anonymous reviewers. This work is a contribution to the International Geoscience Programme (IGCP) Project No. 591 – The Early to Middle Paleozoic Revolution.