1. Introduction
Pachyvaranus crassispondylus Arambourg, Reference Arambourg1952 is a problematic squamate erected on the basis of some isolated vertebrae, which are frequent fossil material in the Maastrichtian phosphates of Morocco. Only four of these vertebrae were figured (Arambourg, Reference Arambourg1952, pl. XLI, figs 4–7). Osteoderms were also referred to this species (Arambourg, Reference Arambourg1952, pl. XLI, figs 2, 3) but their assignment to Squamata is hereafter questioned. Arambourg (Reference Arambourg1952) only shortly described the specimens and provided a brief diagnosis: ‘aquatic lacertilian with shortened vertebrae and centra triangular, short, almost as wide as long, and remarkable by their general pachyostosis’ (translated from French). It is worth pointing out that the osteoderms were not taken into account in the diagnosis.
Subsequent studies dealing with Pachyvaranus are rare and recent. Bardet et al. (Reference Bardet, Cappetta, Pereda Suberbiola, Moutys, Al Maleh, Ahmad, Khrata and Gannoum2000) described vertebrae from the Early Maastrichtian phosphates of Syria that were found to conform well with those of P. crassispondylus, although they were referred to Varanoidea gen. et sp. indet. These vertebrae are here referred to P. crassispondylus. Buffrénil et al. (Reference Buffrénil, Bardet, Pereda Suberbiola and Bouya2008) studied the inner structure of dorsal vertebrae of Pachyvaranus from the Maastrichtian phosphates of Morocco, and from this have drawn palaeoenvironmental and functional inferences.
The affinities of Pachyvaranus within Squamata are not clear and have been very rarely discussed in the literature. Arambourg's (Reference Arambourg1952) opinion on Pachyvaranus relationships remains unclear. He considered it as an aquatic lacertilian and referred it to ‘Aigialosauridae’, although he acknowledged that none of the aigialosaurids have vertebrae similar to those of Pachyvaranus. Also, Arambourg (Reference Arambourg1952) noted similarities with vertebrae of Varanidae, except for their pachyostosis s.s. (pachyostosis sensu stricto corresponds to an increase of cortical deposits leading to an alteration of the bone morphology by causing an increase in bone volume: Buffrénil & Rage, Reference Buffrénil and Rage1993), a character lacking in all extant varanids, and lizards in general. Lee, Caldwell & Scanlon (Reference Lee, Caldwell and Scanlon1999) briefly mentioned that Pachyvaranus might be a pachyophiid snake, but without giving any argument in favour of this opinion. On the basis of the lack of a zygosphene–zygantrum system in Pachyvaranus, Lee & Scanlon (Reference Lee and Scanlon2002) discarded affinities with dolichosaurs and aigialosaurs, but did not suggest relationships for this taxon. Molnar (Reference Molnar2004) regarded Pachyvaranus as a platynotan, without further details. Finally, Rage & Néraudeau (Reference Rage and Néraudeau2004) considered it as a Varanoidea incertae sedis and pointed out that, despite its general resemblance to ‘aigialosaurs’, it probably does not belong to mosasauroids because it lacks a zygosphene–zygantrum system.
New specimens of Pachyvaranus, including numerous vertebrae and an articulated incomplete specimen (see Section 3.a), have been discovered recently in the Maastrichtian phosphates of Morocco, in the framework of an active French–Moroccan collaboration (see Acknowledgements).
On the basis of both Arambourg's and the new collected specimens, this work aims to re-describe in detail the vertebral anatomy of Pachyvaranus crassispondylus and to re-evaluate its affinities within squamates.
Institutional abbreviations: BSP – Bayerische Staatssammlung für Paläontologie und historische Geologie (München, Germany); MNHN – Muséum National d'Histoire Naturelle (Paris, France); OCP – Office Chérifien des Phosphates (Khouribga, Morocco); YPM – Yale Peabody Museum of Natural History (New Haven, Connecticut, USA).
2. Geographical and geological frameworks
During Late Cretaceous and Early Palaeogene times, platforms bordering the southern margin of the Mediterranean Tethys were characterized by phosphatic sedimentation in warm and shallow-marine environments (Lucas & Prévôt-Lucas, Reference Lucas, Prévôt-Lucas, Nairn, Ricou, Vrielynck and Dercourt1996). Phosphatic deposits, forming part of this so-called ‘Mediterranean Tethyan phosphogenic Province’, currently crop out from North Africa to the Middle East, where they are exploited as a valuable economic resource.
The phosphatic deposits of Morocco (Fig. 1a) crop out in several basins, the main ones being the Oulad Abdoun (Fig. 1b) and the Ganntour (Fig. 1c) basins, located in the central Moroccan Meseta. Stratigraphically, they range from the latest Cretaceous (basal Maastrichtian) to the Middle Eocene (basal Lutetian), spanning one of the longest time intervals of all Tethyan phosphates (Lucas & Prévôt-Lucas, Reference Lucas, Prévôt-Lucas, Nairn, Ricou, Vrielynck and Dercourt1996). The Maastrichtian phosphatic series in the Daoui area (NE Oulad Abdoun Basin) is very condensed, being only about 2–5 m thick. It includes a basal bone-bed (BBB) overlain by two main phosphatic layers named ‘Couche III inférieure’ (or Lower CIII level – LCIII) and ‘Couche III supérieure’ (or Upper CIII level – UCIII) (Fig. 1d). It is regarded as Late (but not latest) Maastrichtian in age on the basis of selachian associations (Cappetta, Reference Cappetta and Schultze1987). The Maastrichtian series near Ben Guerir (central Ganntour Basin), has undergone greater subsidence, being normally about 20 m thick but occasionally reaching a thickness of up to 45 m (Fig. 1e). It is composed of several phosphatic levels numbered from basement to top ‘Couche 6’ to ‘Couche 2’ (C6–C2), overlain by a last level called ‘Sillon X’ (SX). On selachian tooth evidence, the series ranges from the Early (C6) to the latest (SX) Maastrichtian (H. Cappetta, pers. comm.). Isolated vertebrae of Pachyvaranus are abundant in both basins. In the Daoui area (Oulad Abdoun), they are found in both lower and upper CIII levels (Fig. 1d). In the Ben Guerir area (Ganntour), most phosphatic levels contain vertebrae of Pachyvaranus, which are more abundant in the upper part of the Maastrichtian series (C3 and C2 levels) (Fig. 1e).

Figure 1. Location map and stratigraphy of the phosphatic basins of Morocco. (a) Position of the main phosphatic basins of Morocco. (b) Details of the Oulad Abdoun Basin with location of the phosphate zones currently exploited (Daoui, Meraa, Sidi Chennane). (c) Details of the Ganntour Basin with location of phosphate zones currently exploited (Ben Guerir, Youssoufia). (d) Stratigraphical log of the Maastrichtian phosphatic series in the Oulad Abdoun Basin near Oued Zem. (e) Stratigraphical log of the Maastrichtian phosphatic series in the Ganntour Basin near Ben Guerir. Phosphatic levels numbered from basement to top ‘Couche 6’ to ‘Couche 2’ (C6–C2), overlain by ‘Sillon X’ (SX). Abbreviations: BBB – Basal bone-bed; Fl – flints; LCIII – Lower CIII level; Li – limestones; Ma – marls; Ph – phosphates; SX – ‘Sillon X’; UCIII – Upper CIII level. The black stars indicate the stratigraphical levels where material of Pachyvaranus has been found.
The phosphatic deposits of Syria crop out mainly in the Palmyrides Chain, where they are relatively widespread, ranging from Santonian to Late Eocene (see Bardet et al. Reference Bardet, Cappetta, Pereda Suberbiola, Moutys, Al Maleh, Ahmad, Khrata and Gannoum2000 for details). Phosphatic deposits constitute the basal member of the Sawwaneh Formation, of Late Cretaceous age. The vertebrate evidence, especially the selachians, indicates an Early Maastrichtian age for most of these phosphatic outcrops, notably those where few isolated vertebrae of Pachyvaranus have been recovered (Bardet et al. Reference Bardet, Cappetta, Pereda Suberbiola, Moutys, Al Maleh, Ahmad, Khrata and Gannoum2000).
According to the occurrence of Pachyvaranus material in the different phosphatic basins of both Morocco and Syria, the stratigraphical range of Pachyvaranus is thus the whole Maastrichtian stage.
3. Material and methods
3.a. Material
Remains of Pachyvaranus consist of vertebrae and ribs only, in total about 49 specimens (Table 1). The available material includes: (1) the original vertebrae collected by Arambourg (Reference Arambourg1952), some having been figured but most remaining unfigured; (2) new specimens, including an articulated portion of vertebral column with ribs, coming from roughly the same stratigraphical levels (Maastrichtian) and from the same geographical areas (Ganntour and Oulad Abdoun basins) as the original specimens; (3) one vertebra from the Early Maastrichtian of Syria. Moreover, two other isolated vertebrae assigned to Pachyvaranus are kept in the collections of the YPM (Connecticut, USA) and BSP (Germany), respectively (Smith, Bhullar & Holroyd, Reference Smith, Bhullar and Holroyd2008; Smith, pers. comm. 2009).
Table 1. List of the material of Pachyvaranus crassispondylus Arambourg, Reference Arambourg1952 with corresponding anatomical, geographical, stratigraphical and reference data

Abbreviations: M –Morocco; S – Syria. For vertebrae four measurements are provided: WPr – width across the prezygapophyses; WPt – width across the postzygapophyses; WPa – maximal width across the paradiapophyses; L – length between the cotylar rim and the condylar tip. X – not available or non-applicable; italicized numbers – estimated from one half of the vertebra.
3.b. Methods
The position of Pachyvaranus isolated vertebrae within the vertebral column was inferred after comparisons with living lizards, more specifically Varanus griseus (MNHN uncatalogued specimen), Varanus rudicollis (MNHN AC 1983–6), Varanus varius (MNHN AC 1910–12) and, to a lesser extent, with several mosasaurid genera, using Russell's (Reference Russell1967) general descriptions.
In all these taxa, the vertebral series bear the following general characters: (1) cervicals with paradiapophyses and hypapophyseal peduncles; (2) dorsals with paradiapophyses only; (3) sacrals with strong sacral processes and centrum lacking any ventral projection; (4) caudals with transverse processes and/or haemal peduncles for chevrons.
Moreover, although sub-regions are not clearly defined, gradual variation, as described hereafter, allows determination of the approximate position of each isolated vertebra within regions: (1) in cervicals, length and size of the hypapophyseal peduncle decreases as one proceeds anteroposteriorly so that it is reduced to a small knob in posteriormost cervicals; (2) in dorsals, the ventral border of the paradiapophyses becomes progressively more dorsal as one proceeds anteroposteriorly; (3) in caudals, zygapophyses and transverse processes progressively become smaller and finally disappear posteriorly, whereas haemal arches appear posteriorly; the caudal region is divided into anterior caudals (‘pygals’) with transverse processes only, median caudals with transverse processes and chevrons, and posterior caudals with chevrons only. Although there are generally numerous pygal vertebrae in mosasaurids (see Russell, Reference Russell1967), most Anguimorpha bear only one pygal (Hoffstetter & Gasc, Reference Hoffstetter, Gasc and Gans1969).
Based on this general description, four vertebral regions are recognized in Pachyvaranus: cervical, dorsal, sacral and caudal.
When applicable, the following measurements were taken for each vertebra: width across the prezygapophyses (WPr) and across the postzygapophyses (WPt), maximal width across the paradiapophyses (WPa), and length between the cotylar rim and the condylar tip (L) (Table 1).
4. Systematic palaeontology
SQUAMATA Oppel, Reference Oppel1811
varanoidea Camp, Reference Camp1923
Pachyvaranidae clade nov.
Definition. All varanoids closer to Pachyvaranus than to Varanus and any other varanoid (stem-based taxon).
Pachyvaranus Arambourg, Reference Arambourg1952
Diagnosis. As for the type and only known species.
Pachyvaranus crassispondylus Arambourg, Reference Arambourg1952
Holotype. MNHN PMC 1, a mid-dorsal vertebra (Arambourg, Reference Arambourg1952, pl. XLI, fig. 5).
Type locality and horizon. ‘Tranchée du chemin de fer, côte 313’, Youssoufia (formerly Louis Gentil), West Ganntour Basin, Morocco; Phosphates, imprecise Maastrichtian level (Arambourg, Reference Arambourg1952, pp. 27–8, fig. 7).
Referred specimens (see Table 1 for details). MNHN PMC 2–4 (Arambourg, Reference Arambourg1952), MNHN PMC 20–21 (Buffrénil et al. Reference Buffrénil, Bardet, Pereda Suberbiola and Bouya2008), MNHN PMC 35–40 (Arambourg's unpub. material), OCP DEK/GE 461–496 (new collected specimens), MNHN CHA 3 (Bardet et al. Reference Bardet, Cappetta, Pereda Suberbiola, Moutys, Al Maleh, Ahmad, Khrata and Gannoum2000).
Provenance (see Table 1 for details). Oulad Abdoun Basin: basal bone-bed of Casbah Tadla and ‘Couche III supérieure’ of Daoui; Ganntour Basin: ‘Phosphates’ of Youssoufia, ‘Couche 6’ to ‘Couche 2’ of Ben Guerir; Palmyrean Basin: Phosphates of Charquieh A mine and Soukkari outcrop.
Revised diagnosis. Varanoid differing from all squamates in the dorsally notched neural spine (autapomorphy); from all non-pythonomorph varanoids in its intense pachyostosis s.s.; from Pythonomorpha in the absence of zygosphene and zygantrum. Moreover, differs from Helodermatidae in the presence of peduncles for the articulation of chevrons; from Necrosauridae in the small size of the neural canal and the subrectangular shape of the neural spine; from Varanidae in the absence of precondylar constriction; and from Lanthanotidae in the triangular shape of the centrum.
Pachyvaranus is characterized by the combination of the following characters: pachyostotic s.s. vertebrae (except anteriormost cervicals) and ribs; procoelous vertebrae; hypapophysis of central origin; absence of zygosphene–zygantrum system; vertebrae wider anteriorly than posteriorly; posterior shift of the maximum depth of interzygapophyseal constriction; subtriangular shape of the centrum in ventral view; cotyle and condyle dorso-ventrally depressed; absence of precondylar constriction; relatively small neural canal; transversely notched neural spine; zygapophyses markedly inclined on the horizontal and lying clearly above the floor of the neural canal; absence of any trace of prezygapophyseal processes; presence of large paracotylar foramina and absence of parazygosphenal foramina; peduncles for the articulation of caudal chevrons.
5. Description
5.a. General morphology
For anatomical nomenclature, see Fig. 2. Almost all vertebrae display pachyostosis s.s., a distinctive character of this taxon. In the centrum, the presence of pachyostosis s.s. is inferred from the convexity of the lateral borders in ventral view (Fig. 3f, i). When it also affects the neural arch and the neural spine, pachyostosis s.s. renders these parts of the vertebrae swollen (Fig. 4e). Where pachyostosis s.s. is very intense, vertebrae become very massive and display a very bloated aspect (Fig. 3g).

Figure 2. Pachyvaranus crassispondylus Arambourg, Reference Arambourg1952. Ben Guerir Area. Ganntour Basin. Maastrichtian. Pictures illustrating morphological features described in the text. (a, b) Anterior cervical vertebra OCP DEK/GE 461; (c, d) mid-dorsal vertebra OCP DEK/GE 479; in (a), (c) anterior and (b, d) left lateral views. Scale bar equals 5 mm. Abbreviations: Co – condyle; Ct – cotyle; Dr – anterodorsal diapophyseal ridge; Er – epidiapophyseal ridge; Hp – hypapophyseal peduncle; Pcf – paracotylar foramen; Pd – paradiapophysis; Pl – ventral parapophyseal lamina; Prz – prezygapophysis; Ptz – postzygapophysis; Sr – subcentral ridge.

Figure 3. Pachyvaranus crassispondylus Arambourg, Reference Arambourg1952. Ben Guerir Area. Ganntour Basin. Maastrichtian. (a–e) anterior cervical vertebra OCP DEK/GE 461; (f–l) posterior cervical vertebrae; (f) OCP DEK/GE 463; (g, h) OCP DEK/GE 464; (i–l) OCP DEK/GE 465; in (a, g, k) anterior; (b, h) posterior; (c, j) dorsal; (d, f, i) ventral; and (e, l) left lateral views. Scale bars equal 5 mm.

Figure 4. Pachyvaranus crassispondylus Arambourg, Reference Arambourg1952. (a–j) Ben Guerir Area. Ganntour Basin; (k) Grand Daoui. Oulad Abdoun Basin. Maastrichtian. (a, b) anterior dorsal vertebra OCP DEK/GE 466; (c, d) mid dorsal vertebra OCP DEK/GE 479; (e–g) posterior dorsal vertebra OCP DEK/GE 482; (h–j) posteriormost dorsal vertebra OCP DEK/GE 491; (k) posteriormost dorsal vertebra OCP DEK/GE 490; in (a, c, e) anterior; (b, h, k) ventral; (d) right lateral; (f, i) dorsal; and (g, j) left lateral views. Scale bars equal 5 mm.
Vertebrae from the different regions of the column present several common features. In dorsal view, vertebrae are much larger anteriorly than posteriorly, resulting from the important lateral projection of the paradiapophyses that extend laterally beyond the level of the prezygapophyses. This aspect is stressed by the fact that vertebrae are even wider across the prezygapophyses (paradiapophyses not taken into account) than across the postzygapophyses (Fig. 4f), which is apparently a feature unique to aquatic varanoids from the Cretaceous (J.C.R., pers. obs.). All centra are procoelous. The cotyle and condyle are dorso-ventrally depressed and there is no precondylar constriction. The neural spine is well-developed. The neural canal is relatively small. The zygapophyses are markedly inclined on the horizontal and they lie at a high level, clearly above the floor of the neural canal. Prezygapophyses lack any trace of prezygapophyseal processes, but more or less similar salients occur in a few vertebrae (see below). There is no zygosphene–zygantrum system. The presence, size and number of paracotylar, subcentral and lateral foramina is very variable from one vertebra to another and does not reflect any particular position along the vertebral column. However, all vertebrae lack parazygosphenal foramina.
5.b. Cervical vertebrae
Cervical vertebrae are comparatively more elongate than anterior dorsals. They bear a ventral hypapophyseal peduncle, continuous with the centrum, which is very posteriorly located and projects postero-ventrally (Fig. 3e). The hypapophyses are not fused to the centra. Other characteristics of the cervical vertebrae are the anteroventral projection of the paradiapophyses beyond the ventral border of the cotyle (Fig. 3g), as well as the horizontal direction of the axes of the cotyle and condyle.
Anterior cervical vertebrae (OCP DEK/GE 461, 462). These vertebrae are particularly elongated. They do not display pachyostosis s.s. in their centrum but their neural arch is perhaps slightly pachyostotic s.s. The hypapophyseal peduncle is relatively long and ends as an ovoid facet whose main axis is directed anteroposteriorly (Fig. 3d, e); the facet is concave and faces posteroventrally. On this facet articulates the distal element of the hypapophysis (hypocentrum sensu Hoffstetter & Gasc, Reference Hoffstetter and Gasc1967) that is not preserved in the available specimens. It must be specified that, as opposed to Hoffstetter & Gasc (Reference Hoffstetter and Gasc1967), Caldwell (Reference Caldwell2000) and Caldwell & Calvo (Reference Caldwell and Calvo2008) considered the hypapophyseal peduncle to be the entire hypapophysis, whereas the distal element was labelled intercentrum (which is the other name of the hypocentrum). The paradiapophyseal articular facets are very posteriorly located at about half the length of the centrum, and even beyond in the anteriormost vertebra OCP DEK/GE 461 (Fig. 3e). The facets are small and ovoid in OCP DEK/GE 461 (Fig. 3e) but dorso-ventrally more extended in OCP DEK/GE 462. There is no epidiapophyseal ridge (sensu Rage & Néraudeau, Reference Rage and Néraudeau2004), but a marked ridge connects the dorsal border of the paradiapophysis to the prezygapophysis; this ridge is hereafter referred to as anterodorsal diapophyseal ridge (Fig. 2b). Each paradiapophysis is prolonged anteroventrally by a salient lamina, hereafter referred to as ventral parapophyseal lamina (Fig. 2b), which ends below the ventral limit of the cotyle where it forms a ventral projection. These projections entail the embayment of the ventral rim of the cotyle in both anterior and ventral aspects (Fig. 3d). It must be specified that the ventral projection of the parapophyseal lamina is similar, but likely not homologous, to the parapophyseal processes that occur in vertebrae of advanced snakes (Hoffstetter, Reference Hoffstetter1939). The posterior part of the centrum is narrow, but the anterior part is highly modified by the paradiapophyses and the ventral lamina (Fig. 3d). The neural spine runs from the level of the posterior limit of the prezygapophyses to the posterior border of the neural arch (Fig. 3c). The anterior border is inclined posteriorly (Fig. 3e). As the neural arch slopes markedly anteriorly, the anterior border is clearly longer than the posterior one. In dorsal view, the neural spine widens posteriorly and ends as a tubercle (eroded; Fig. 3c). The neural spine is particularly short. The anterior edge of the neural arch forms an anterior projection similar to that of the varanoid Saniwa (Fig. 3c; Rieppel & Grande, Reference Rieppel and Grande2007); there are no articular facets, hence it is neither a zygosphene nor a pseudozygosphene. The neural canal is as high as the cotyle. The latter is strongly enlarged transversely and its dorsal and ventral borders are almost straight and parallel. Prezygapophyses are located much higher than the paradiapophyses (Fig. 3e). The best preserved, although damaged, prezygapophysis of vertebra OCP DEK/GE 462 suggests that prezygapophyses are very elongated and protrude beyond the anterior border of the centrum.
Posterior cervical vertebrae (OCP DEK/GE 463–465). In these vertebrae, pachyostosis s.s. is strong in the centrum, whose lateral borders are slightly convex (Fig. 3f, i) and slightly less intense in the neural arch. The hypapophyseal peduncle is very short and reduced to a small knob (Fig. 3f). In OCP DEK/GE 463–464, the distal concave facet is preserved (doubtfully in OCP DEK/GE 463). However, in OCP DEK/GE 465 the peduncle is shorter than in the two other specimens, does not display any articular facet, and barely projects from the centrum (Fig. 3i, l). OCP DEK/GE 465 thus likely occupied a more posterior position in the neck than OCP DEK/GE 464 and OCP DEK/GE 463. Paradiapophyses are located more anteriorly than in anterior cervicals, at the level of the posterior limit of the prezygapophyses (Fig. 3l). They are very dorso-ventrally elongated, dorsally almost reaching the level of the prezygapophyseal facet (Fig. 3k, l). A short epidiapophyseal ridge, which anteriorly originates from the dorsal, recurved part of the paradiapophysis, is present, as well as the anterodorsal diapophyseal ridge. The epidiapophyseal ridge and the subcentral ridge, the latter of which limits the ventral face of the centrum laterally, define a marked triangular and concave area posterior to the paradiapophysis; the concavity of this area may be reduced by pachyostosis s.s. Paradiapophyseal articular facets extend dorsoventrally and their dorsal part bends posteriorly. They are peculiarly globular in their median part and protrude into the triangular concave area posterior to the paradiapophysis (Fig. 3l), a characteristic up to now not reported from other taxa. The ventral parapophyseal lamina is present but, as a result of the anterior position of the articular facet, it is clearly shorter and less oblique than in anterior cervicals. Moreover, it appears to be thicker (Fig. 3f). As in anterior cervicals, the extremity of the lamina projects ventrally beyond the cotyle rim (Fig. 3f). The cotyle is less enlarged transversely. The ventral face of the centrum assumes a subtriangular shape and is wider anteriorly than long. OCP DEK/GE 464 and OCP DEK/GE 465 preserve the neural spine. This neural spine is higher and thicker than in the anterior cervicals (Fig. 3k) and occupies almost the whole length of the neural arch. The anterior and posterior borders are vertical or subvertical (Fig. 3l). It is worth noting that a vertical notch transversally indents the dorsal border of the neural spine close to its posterior limit (Fig. 3j). The dorsal border is flat and clearly thickened in OCP DEK/GE 465 (Fig. 3k). This vertebra is the only cervical one in which a prezygapophyseal facet is well preserved; the facet is relatively elongate and antero-laterally directed (Fig. 3j). This prezygapophysis is strongly inclined on the horizontal (Fig. 3k).
5.c. Dorsal vertebrae and ribs
Dorsal vertebrae lack any trace of salient, either a hypapophyseal peduncle or a haemal keel, on the ventral face of the centrum. The anterodorsal diapophyseal ridge is more marked than in the cervicals and its anterior extremity even looks like a small anterolateral process, which must not be mistaken for a prezygapophyseal process, in some vertebrae (OCP DEK/GE 469, 476, 484). The paradiapophyses are positioned as anteriorly but more dorsally than in the cervicals. Except for their dorsoventral extent, paradiapophyseal articular facets are similar to those of the posterior cervicals. The epidiapophyseal ridge is salient and often sharp; in dorsal view, it is either slightly concave (Fig. 4i) or convex posterolaterally (Fig. 4f) depending on the intensity of pachyostosis s.s. Whereas the axes of the cotyles and condyles are horizontal in cervicals, they are inclined in dorsals, the cotyle facing anteroventrally and the condyle being oriented posterodorsally. The degree of pachyostosis s.s. varies relatively significantly among specimens.
For some partially preserved broken vertebrae (OCP DEK/GE 495, MNHN PMC 20, 36, 39), the dorsal subregion could not be estimated.
Anterior dorsal vertebrae (OCP DEK/GE 466–475, 496, MNHN PMC 3, 35). The overall morphology of these vertebrae does not differ markedly from that of posterior cervicals, except for the absence of hypapophyseal peduncles. However, they are more depressed and wider. Unfortunately, the neural spine is broken away in all the specimens. The ventral face of the centrum is triangular or subtriangular, nearly flat to slightly convex (Fig. 4b). Paradiapophyseal articular facets are slightly more inclined posteriorly than in the cervicals. Except perhaps in the anteriormost dorsal vertebrae (OCP DEK/GE 466 and 467), the ventral parapophyseal laminae are less clearly distinct and do not project ventrally beyond the ventral rim of the cotyle. The ventral border of the paradiapophyses, which is level with the ventral border of the cotyle in the anteriormost dorsal vertebrae (Fig. 4a), is located above this level in the others.
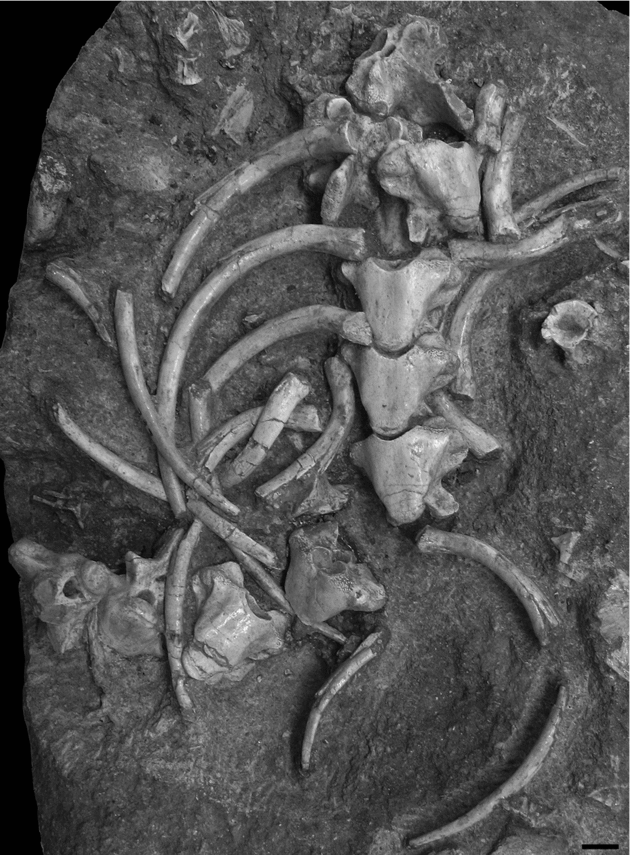
Specimen OCP DEK/GE 496 corresponds to nine associated vertebrae and ribs preserved in a block of matrix (Fig. 5). Only three vertebrae are articulated but they are all preserved in their position of articulation, despite some changes in orientation. Most vertebrae and ribs are preserved in ventral view. From the morphology of the vertebrae, notably from the position of the paradiapophyses observable on the sixth vertebra as preserved, and by comparison with the material described above, these vertebrae are considered as anterior dorsals. Like vertebrae, ribs are heavily pachyostotic s.s. Most are randomly distributed and oriented. However, some of them are preserved in association with the corresponding vertebrae but without being closely articulated to them. Ribs appear holocephalous, with a single concave ovoid articular surface, and deprived of tuber costae. They project lateroventrally and then remain regularly curved, which suggests the absence of lateral compression of the body.

Figure 5. Pachyvaranus crassispondylus Arambourg, Reference Arambourg1952. Grand Daoui. Oulad Abdoun Basin. Maastrichtian. OCP DEK/GE 496. Associated vertebrae and ribs preserved in a block of matrix. Scale bar equals 1 cm.
Mid-dorsal vertebrae (OCP 476–481, MNHN PMC 1–2, MNHN CHA 3). Mid-dorsals have been defined as all dorsal vertebrae exhibiting an intermediate pattern between anterior and posterior dorsal ones.
Posterior dorsal vertebrae (OCP DEK/GE 482–491, MNHN PMC 4, 21, 37, 38). Vertebrae from the posterior dorsal region, except posteriormost ones (see below), differ little from anterior dorsals. The ventral border of the paradiapophyses is located more dorsally than in anterior dorsals, lying clearly above the level of the cotylar ventral rim (Fig. 4e). As a result, the paradiapophyseal facet is shorter dorsoventrally; it appears also to be slightly more inclined posteriorly (Fig. 4g). In connection with this more dorsal position of the paradiapophyses, the lateral parts of the ventral face of the centrum are deflected dorsolaterally, so that only a more or less wide central part remains flat (Fig. 4e). Vertebra OCP DEK/GE 482 preserves the entire neural spine; it is similar in shape to that of the posterior cervical OCP DEK/GE 465, including the presence of the notch in the dorsal border, but it is not as high and not so straight anteriorly (Fig. 4g).
Posteriormost dorsal vertebrae are more elongate and, apart from the width across paradiapophyses, narrower than other dorsals. More specifically, their centrum is only slightly affected by pachyostosis s.s.; it appears to be clearly more cylindrical than triangular (Fig. 4h, k). The dorsoventral extent of the paradiapophyseal facets is short and the ventral border of the paradiapophyses is close to the level of the dorsal limit of the cotyle (Fig. 4j). The neural spine of OCP DEK/GE 490 is partly preserved. It was lower than that of more anterior vertebrae, which is a usual variation among squamates, and has a posterior notch in its dorsal border. Whereas pachyostosis s.s. is intense in OCP DEK/GE 489, it is very weak in the two posteriormost dorsal vertebrae.
5.d. Sacral vertebrae
Two sacral vertebrae are available (OCP DEK/GE 492, MNHN PMC 40). They are characterized by a particularly short and flat centrum (Fig. 6b). Unfortunately, the sacral processes are broken away and only their bases are preserved. They originate from the ventral part of the centrum and appear to have projected postero-laterally (Fig. 6b, c). As in some dorsal vertebrae, the anterodorsal diapophyseal ridge in OCP DEK/GE 492 looks like a small anterolateral process. No neural spine is preserved. The axes of the cotyle–condyle system are oblique. Pachyostosis s.s. is relatively intense in the neural arch of OCP DEK/GE 492. No trace of fusion of one sacral vertebra with another is observable.

Figure 6. Pachyvaranus crassispondylus Arambourg, Reference Arambourg1952. Ben Guerir Area. Ganntour Basin. Maastrichtian. (a–c) sacral vertebra OCP DEK/GE 492; (d–h) pygal vertebra OCP DEK/GE 493; (i–k) mid-caudal vertebra OCP DEK/GE 494; in (a, d, i) anterior; (b, g) ventral; (c, h) left lateral; (e, j) posterior; (f) dorsal; and (k) right lateral views. Scale bars equal 5 mm.
5.e. Caudal vertebrae
Two caudal vertebrae are preserved, one being a pygal, the other a caudal from the mid-portion of the tail.
Pygal (OCP DEK/GE 493). One sub-complete vertebra (Fig. 6d–h) is a pygal one, that is, an anterior caudal with transverse processes but without haemal arches. Its general morphology is very similar to that of posteriormost dorsals. The centrum is sub-cylindrical and slightly pachyostotic s.s., like the rest of the vertebra. A very low and thin sagittal ridge runs on its ventral surface. Transverse processes, located above the level of the cotylar ventral rim, beyond the posterior level of the prezygapophyses, are unfortunately broken, so that their shape and orientation can not been inferred.
Mid-caudal (OCP DEK/GE 494). One incomplete caudal vertebra is available (Fig. 6i–k). It is short and, apart from the transverse processes, relatively narrow. Its neural arch is pachyostotic s.s. The neural spine is inclined posteriorly (Fig. 6k). It is as high as in posterior dorsals and higher than in posteriormost dorsals. The dorsal border is damaged, but the shape of the upper section of the neural spine shows that a posterior notch was present. The transverse processes were present but are broken off. They originate from the dorsal part of the centrum. In ventral view, the centrum is approximately cylindrical; it does not appear to be pachyostotic s.s. Two ventral peduncles for articulation with chevron bones are located posteriorly, against the condyle (Fig. 6k). They are prolonged anteriorly by keels that define a sagittal groove (Fig. 6i, j). The axes of the cotyle–condyle are oblique.
6. Discussion
6.a. Taxonomic determination
The material assigned to Pachyvaranus crassispondylus does not provide enough character data to perform a phylogenetic analysis. Nevertheless, the material is sufficient to provide a more detailed diagnosis of this previously poorly known taxon. Vertebrae are procoelous, as in other squamates. Within squamates, the development of a hypapophysis of central origin with individualized hypocentral elements, the obliquity of the axis of the cotyle–condyle system, the posterior position of the narrowest part of the interzygapophyseal constriction observed in dorsal view, and the presence of articulated caudal chevron intercentra, are characteristics of Varanoidea (Hoffstetter & Gasc, Reference Hoffstetter and Gasc1967; Lee, Reference Lee1997). The absence of a zygosphene–zygantrum system excludes Pachyvaranus from Pythonomorpha (Lee, Reference Lee1997). Within non-pythonomorph Varanoidea, it differs from the Helodermatidae in the presence of peduncles for the articulation of chevron bones (Rieppel, Reference Rieppel1980), from Necrosauridae in the small size of the neural canal and the sub-rectangular shape of the neural spine (A.H. & J.C.R., pers. obs.), from Varanidae in the absence of precondylar constriction and from Lanthanotidae in the triangular shape of the centrum (Rieppel, Reference Rieppel1980). More generally, it differs from all non-pythonomorph varanoids in the intense pachyostosis s.s. of its vertebrae and ribs. It also displays a notched neural spine, which appears to be unmatched within squamates. Moreover, the peculiar globular aspect of the middle part of the paradiapophyseal articular facets in posterior cervicals (although it may be linked to pachyostosis s.s.) has not been reported in another taxon. For these reasons, this taxon should be placed in a new unranked clade of non-pythonomorph Varanoidea. We therefore define the Pachyvaranidae as ‘all varanoids closer to Pachyvaranus than to Varanus and any other varanoid’ (stem-based taxon).
6.b. Palaeoecological implications
Pachyvaranus is characterized by a very intense pachyostosis s.s. in vertebrae and ribs. Moreover, the histological study realized on two dorsal vertebrae by Buffrénil et al. (Reference Buffrénil, Bardet, Pereda Suberbiola and Bouya2008) revealed the presence of osteosclerosis, that is, an increase of inner bone compactness, which is also suggested by the observation of broken vertebrae. Within squamates, pachyostosis s.s. is encountered only in fossil Pythonomorpha from the Cretaceous (review in Houssaye, Reference Houssaye2009). In these taxa, pachyostosis s.s., when present, is always associated with osteosclerosis (A.H., pers. obs.). The presence of pachyostosis s.s. from cervicals to caudals in Pachyvaranus suggests that pachyosteosclerosis (pachyostosis s.s. + osteosclerosis) was probably displayed all along its axial skeleton. In Pachyvaranus, pachyostosis s.s. appears very intense and maximal from the posteriormost cervicals to the posterior dorsals (except the posteriormost ones).
Pachyosteosclerosis is encountered in slow swimmers adapted to shallow water environments, in which the increased skeletal mass is considered to play the role of ballast for hydrostatic regulation of buoyancy and of body trim (Taylor, Reference Taylor2000; Houssaye, Reference Houssaye2009). As highly mineralized bones are brittle (Watkins, Reference Watkins and Watkins1999), osteosclerosis favours propagation of fractures and appears incompatible with a terrestrial habitat (Taylor, Reference Taylor2000; Buffrénil et al. Reference Buffrénil, Bardet, Pereda Suberbiola and Bouya2008). As a consequence, taxa displaying pachyosteosclerosis should not be considered semi-aquatic but fully aquatic forms (Houssaye, Reference Houssaye2009). These conclusions are in accordance with the sedimentary context corresponding to warm and nearshore shallow-marine environments (see Section 2). Pachyvaranus displays regularly curved ribs, which indicates the absence of lateral compression of the body. However, it appears to display high neural spines in the caudal region, which suggests a high, perhaps laterally compressed tail, a typical feature of highly aquatic squamates. The presence of pachyosteosclerosis and of a laterally compressed tail evokes a slow axial undulatory swimmer.
6.c. Palaeobiogeographical implications
It has been suggested that the marine vertebrate faunas, especially the mosasaurid squamates, of the Maastrichtian phosphatic deposits of North Africa and Middle East significantly differ from those of the northern margin (Bardet & Pereda Suberbiola, Reference Bardet and Pereda Suberbiola1996; Bardet, in press). For example, the mosasaurids Halisaurus arambourgi, Globidens phosphaticus, Prognathodon currii and Mosasaurus beaugei are considered as characteristic of the southern margin of the Mediterranean Tethys (Bardet et al. Reference Bardet, Cappetta, Pereda Suberbiola, Moutys, Al Maleh, Ahmad, Khrata and Gannoum2000, Reference Bardet, Pereda Suberbiola, Bouyahyaoui, Iarochene, Bouya and Amaghzaz2004, Reference Bardet, Pereda Suberbiola, Iarochene, Amalik and Bouya2005a,Reference Bardet, Pereda Suberbiola, Iarochene, Bouya and Amaghzazb). The invertebrate faunas found in these North Africa and Middle East latest Cretaceous deposits exhibit similar palaeobiogeographical affinities as the marine vertebrate ones. For example, the oyster Ambigostrea villei is characteristic of the low latitudes of the Mediterranean Tethyian province, having been found in North Africa, the Middle East and southern Europe, while it is absent from northern Europe (Dhondt et al. Reference Dhondt, Malchus, Boumaza and Jaillard1999). Similarly, the ammonite Libycoceras ismaeli is exclusively known at the low latitudes of the southern Tethyan margin (Zaborski & Morris, Reference Zaborski and Morris1999). As shown here, this southern Tethys palaeobiogeographical distribution appears also to apply to Pachyvaranus, only found in the Maastrichtian phosphatic deposits of both Morocco and Syria. These faunal differences between the southern and northern margins of the Mediterranean Tethys could be related to palaeolatitudinal gradients (Bardet & Pereda Suberbiola, Reference Bardet and Pereda Suberbiola1996; Bardet, in press). Indeed, the marine vertebrate assemblages from the Middle East and North Africa, as well as those from West Africa and eastern South America, which show close similarities, are all included in a palaeolatitudinal belt located between 20° N and 20° S.
6.d. The question of the osteoderms
Arambourg (Reference Arambourg1952) tentatively referred fragmentary ‘osteoderms’ to Pachyvaranus crassispondylus with the argument that they were sometimes associated to its vertebrae in the phosphatic outcrops of Morocco. Two of them (MNHN PMC 5, 6), coming from the ‘Couche III’ (Maastrichtian) of Oued Zem and Mohamed Chleuh (Oulad Abdoun Basin), respectively, were only briefly described and figured (Arambourg, Reference Arambourg1952, p. 289, pl. XLI, figs 2, 3).
The fragments are about 2–3 mm thick and despite their size (up to 25 mm) they are flat, without any indication of curvature. One surface bears tubercles, whereas the other is smooth. The tubercles are closely spaced and arranged in more or less parallel rows. In the largest fragment (MNHN PMC 6), the tuberculated ridges seem to radiate from an unpreserved centre of origin. None of the natural margins are preserved so that the extension of these bony elements cannot be estimated. The internal structure (available on the broken margins of the specimens) shows a clearly stratified bony structure made of lamellar bone overlain by dentine tubercles and a shiny, possibly enameloid layer (P. Janvier, pers. comm. 2009).
To our knowledge, this type of ornamentation does not resemble that of osteoderms of any known lizards, as previously pointed out by Rage & Néraudeau (Reference Rage and Néraudeau2004). Due to their large size, it is also unlikely that they could belong to an animal like Pachyvaranus, whose total body length is estimated at up to 1.50 m. Moreover, the internal bone structure does not look like that of reptiles in general and is rather reminiscent of the dermal bone of actinopterygian fishes, in particular teleosts (P. Janvier, pers. comm. 2009).
Finally, these ornamented fragments, which are relatively frequent in the Maastrichtian phosphates of Morocco, closely resemble some cranial bones of the dercetid teleost Stratodus apicalis Cope, Reference Cope1872 (N.B., pers. obs.), especially those from the Maastrichtian phosphates of Morocco and Negev (see Arambourg, Reference Arambourg1952, pl. XXXIV, fig. 11; Chalifa & Lewy, Reference Chalifa and Lewy1991, pl. 2, figs 4, 5). It is particularly surprising that Arambourg did not notice this resemblance.
Morphological and macro-histological observations are thus congruent and argue in favour of an assignment to fish for these remains. For all these reasons, we suggest that these bony elements are not lizard ‘osteoderms’ but rather fragments of fish cranial bones, which probably belong to the dercetid teleost Stratodus apicalis, a large open-sea predator actinopterygian known from the Campanian–Maastrichtian of North America, northwestern Africa and Middle East (see references in Bardet et al. Reference Bardet, Cappetta, Pereda Suberbiola, Moutys, Al Maleh, Ahmad, Khrata and Gannoum2000).
Acknowledgements
This work has benefited from the scientific program of collaboration between the Office Chérifien des Phosphates (OCP, Casablanca), the Ministère de l'Energie, des Mines, de l'Eau et de l'Environnement (MEMEE, Rabat), the Muséum National d'Histoire Naturelle (MNHN, Paris), and the Universities Cadi Ayyad (Marrakech) and Chouaîb Doukkali (El Jadida). We are grateful to staff members of the OCP mining centres of Khouribga and Ben Guerir for hospitality and logistic support, and to the MEMEE for providing administrative facilities and permits. Many thanks to H. Cappetta (CNRS, Montpellier), S. Adnet (Montpellier) and A. Ben Abdallah (MEMEE, Rabat) for their help in collecting material of Pachyvaranus in Ben Guerir. Many thanks to P. Janvier (CNRS, MNHN, Paris) for advice and rereading concerning the part of the manuscript dealing with the attribution of the osteoderms. Thanks also to B. LeDimet, L. Cazes (MNHN, Paris) and C. Bouillet for mechanical preparation of some specimens, and to P. Loubry (CNRS, MNHN, Paris) for the photographs. We also thank two anonymous reviewers for useful comments that improved the manuscript. This research was supported by funds from the National Geographic Society (Grant no. 6627–99) and BQR ‘Vertébrés fossiles des Phosphates du Maroc’ (MNHN, Paris) for N. Bardet; and from Ministerio de Ciencia e Innovación (projects CGL2004-02338/BTE and CGL2007-64061/BTE), Universidad del País Vasco/EHU (9/UPV 00121.310-15303/2003) and Gobierno Vasco/EJ (research group GIC07/14-361) for X. Pereda Suberbiola.