



1. Introduction
The Global Boundary Stratotype Section and Point (GSSP) for the base of the Cenomanian Stage of the Upper Cretaceous was defined on the southern side of Mont Risou, east of Rosans, Haute-Alpes in France (Fig. 1). According to the definition of the stratotype (Kennedy et al. Reference Kennedy, Gale, Lees and Caron2004), the base of the Cenomanian Stage is a level at 36 m below the top of the Marnes Bleues and coincides with the appearance level of the planktonic foraminifera Thalmanninella globotruncanoides ( = Rotalipora globotruncanoides Sigal, Reference Sigal1948).
Figure 1. Palaeogeographic reconstruction for the late Albian (101.0 Ma) according to Hay et al. (Reference Hay, DeConto, Wold, Wilson, Voigt, Schulz, Rossby-Wold, Dullo, Ronov, Balukhovsky, Soeding, Barrera and Johnson1999) showing the position of the GSSP for the base of the Cenomanian at Mont Risou (Hautes-Alps, France) and the Blake Nose Plateau (NW Atlantic Ocean).
The stratigraphic interval from the middle Albian to the upper Cenomanian records the evolution and diversification of the polyphyletic Rotalipora group (Wonders, Reference Wonders1978; Robaszynski et al. Reference Robaszynski, Caron, Amédro, Dupuis, Hardenbol, Gonzalez-Donoso, Linares and Gartner1994, Reference Robaszynski, Amédro, Gonzalez-Donoso and Linares2008; Kennedy et al. Reference Kennedy, Gale, Lees and Caron2004; Gonzalez-Donoso, Linares & Robaszynski, Reference Gonzalez-Donoso, Linares and Robaszynski2007; Gale et al. Reference Gale, Bown, Caron, Crampton, Crowhurst, Kennedy, Petrizzo and Wray2011). The distinctive rotaliporid assemblage that occurs at the base of the Cenomanian is composed of trochospiral, single-keeled species with supplementary apertures on the umbilical side that have been assigned to two different genera: (1) Pseudothalmanninella (ticinensis, tehamaensis), and (2) Thalmanninella (appenninica, balernaensis, gandolfii, brotzeni, globotruncanoides) (see Wonders, Reference Wonders1978, Reference Wonders1980 and discussion in Gonzalez-Donoso, Linares & Robaszynski, Reference Gonzalez-Donoso, Linares and Robaszynski2007 and Lipson-Benitah, Reference Lipson-Benitah2008). The genera assignment is based on the identification of distinctive lineages that originated in late Albian time, as follows: (1) Ticinella praeticinensis – Pseudothalmanninella subticinensis – Pseudothalmanninella ticinensis – Pseudothalmanninella tehamaensis lineage; (2) Ticinella raynaudi group: Thalmanninella praebalernaensis – Thalmanninella balernaensis – Thalmanninella appenninica – Thalmanninella brotzeni – Thalmanninella globotruncanoides lineage.
The genus Thalmanninella is used to identify spiroconvex and umbiliconvex forms that seem, however, to belong to different lineages. For this reason Lipson-Benitah (Reference Lipson-Benitah2008) introduced the genus Parathalmanninella to accommodate the species (praebalernaensis, balernaensis and appenninica) that evolved from the Ticinella raynaudi group. The species brotzeni and globotruncanoides were regarded by Lipson-Benitah (Reference Lipson-Benitah2008) as belonging to a new phyletic lineage descendent from Pseudothalmanninella tehamaensis and, thus, were included in the emended genus Thalmanninella. To date we adopt the taxonomy of Wonders (Reference Wonders1978) and Gonzalez-Donoso, Linares & Robaszynski (Reference Gonzalez-Donoso, Linares and Robaszynski2007) because Pseudothalmanninella tehamaensis can hardly be regarded as ancestral to Th. brotzeni and Th. globotruncanoides according to the data presented in this paper. However, as we suspect that the genus Thalmanninella may be polyphyletic, further investigations are in progress to identify the ancestral species of brotzeni and globotruncanoides and, hence, to clarify their phylogeny and taxonomic position.
Pseudothalmanninella tehamaensis has been rarely documented to occur in latest Albian to early Cenomanian assemblages (Marianos & Zingula, Reference Marianos and Zingula1966). Thalmanninella globotruncanoides has been reported to be very rare at the beginning of its stratigraphic range (Bellier & Moullade, Reference Bellier and Moullade2002; Petrizzo & Huber, Reference Petrizzo and Huber2006a ; Robaszynski et al. Reference Robaszynski, Amédro, Gonzalez-Donoso and Linares2008), its first occurrence being difficult to detect due to the presence in the same assemblage of either transitional specimens close to Th. globotruncanoides (e.g. Rotalipora aff. globotruncanoides, see Bellier & Moullade, Reference Bellier and Moullade2002) or specimens closely resembling Th. brotzeni or other intermediate forms belonging to the Thalmanninella lineage (Robaszynski et al. Reference Robaszynski, Amédro, Gonzalez-Donoso and Linares2008).
The species brotzeni, included in the genus Rotalipora by Sigal (Reference Sigal1958) and Robaszynski & Caron (Reference Robaszynski and Caron1979), was regarded as the marker species of the homonymous zone extending from near the Albian/Cenomanian boundary through the lower Cenomanian (Sigal, Reference Sigal1977; Robaszynski & Caron, Reference Robaszynski and Caron1995; Premoli Silva & Sliter, Reference Premoli Silva and Sliter1995). Wonders (Reference Wonders1978, Reference Wonders1980) considered the species brotzeni to be a junior synonym of Th. greenhornensis, and, in agreement with van Hinte (Reference van Hinte1976), he equated the top of the Albian Stage with the extinction level of Planomalina buxtorfi. Gonzalez-Donoso & Linares in Robaszynski et al. (Reference Robaszynski, Caron, Amédro, Dupuis, Hardenbol, Gonzalez-Donoso, Linares and Gartner1994) re-interpreted R. brotzeni as a junior synonym of the species globotruncanoides and replaced the lower Cenomanian Rotalipora brotzeni Zone with the Rotalipora globotruncanoides Zone (Robaszynski & Caron, Reference Robaszynski and Caron1995; Gale et al. Reference Gale, Kennedy, Burnett, Caron and Kidd1996). Recently, Gonzalez-Donoso, Linares & Robaszynski (Reference Gonzalez-Donoso, Linares and Robaszynski2007) and Robaszynski et al. (Reference Robaszynski, Amédro, Gonzalez-Donoso and Linares2008) reiterated that the two species are synonyms and included the species globotruncanoides in the genus Thalmanninella.
Therefore, owing to the difficulty in reliably distinguishing between the two species described by Sigal (Reference Sigal1948), Thalmanninella brotzeni and Rotalipora globotruncanoides, reconsideration of the species concepts adopted by the scientists was needed. To verify the taxonomic status of Sigal's species, Caron & Premoli Silva (Reference Caron and Premoli Silva2007) examined the type specimens deposited at the Musée National d’Histoire Naturelle (MNHN) in Paris. According to Caron & Premoli Silva's (Reference Caron and Premoli Silva2007, p. 529) re-description and observation of the holotypes of Thalmanninella brotzeni and Rotalipora globotruncanoides, the two species ‘are not conspecific and must be considered two discrete taxa. From a stratigraphic point of view Th. brotzeni first occurs slightly earlier than Th. globotruncanoides’.
Consequently, in this study we aim to clarify the taxonomy of the index species Th. globotruncanoides, Th. brotzeni as well as Ps. tehamaensis and to document their stratigraphic distribution across the Albian/Cenomanian boundary in the stratotype section at Mont Risou (Kennedy et al. Reference Kennedy, Gale, Lees and Caron2004). In addition, the assemblages at Mont Risou are compared with the equivalent and well-preserved planktonic foraminifera associations recorded at Blake Nose Plateau (Ocean Drilling Program, ODP Leg 171B, North Western Atlantic Ocean; Petrizzo & Huber, Reference Petrizzo and Huber2006a ) in order to elucidate the reliability and the stratigraphic distribution of the marker taxa outside the Mediterranean Tethys (Fig. 1). Sigal's type specimens are discussed and for the first time illustrated through stereoscopic images taken by Annachiara Bartolini, curator at the Musée National d’Histoire Naturelle in Paris.
This study is part of a long-term project of the Mesozoic Planktonic Foraminifera Working Group focusing on the taxonomy and phylogeny of Jurassic–Cretaceous planktonic foraminifera by studying and re-illustrating primary type specimens and by documenting stratophenetic changes in wall microstructure and shell morphology. The working group has published results in the peer-reviewed literature (e.g. Petrizzo & Huber, Reference Petrizzo and Huber2006a ,Reference Petrizzo and Huber b ; Ando & Huber, Reference Ando and Huber2007; Caron & Premoli Silva, Reference Caron and Premoli Silva2007; Gonzalez-Donoso, Linares & Robaszynski, Reference Gonzalez-Donoso, Linares and Robaszynski2007; Georgescu & Abramovich, Reference Georgescu and Abramovich2008; Lipson-Benitah, Reference Lipson-Benitah2008; Spezzaferri & Caron, Reference Spezzaferri and Caron2008; Georgescu & Huber, Reference Georgescu and Huber2009; Georgescu, Saupe & Huber, Reference Georgescu, Saupe and Huber2009; Premoli Silva et al. Reference Premoli Silva, Caron, Leckie, Petrizzo, Soldan and Verga2009; Falzoni & Petrizzo, Reference Falzoni and Petrizzo2011; Huber & Leckie, Reference Huber and Leckie2011; Petrizzo, Falzoni & Premoli Silva, Reference Petrizzo, Falzoni and Premoli Silva2011; Petrizzo et al. Reference Petrizzo, Huber, Gale, Barchetta and Jenkyns2012, Reference Petrizzo, Huber, Gale, Barchetta and Jenkyns2013; Ando, Huber & Premoli Silva, Reference Ando, Huber and Premoli Silva2013; Falzoni et al. Reference Falzoni, Petrizzo, Huber and Macleod2014; Huber & Petrizzo, Reference Huber and Petrizzo2014; Haynes, Huber & MacLeod, in press, among others) and has built an online taxonomic database posted on the CHRONOS website (http://www.chronos.org).
2. Considerations on the primary and secondary types of Thalmanninella brotzeni and Thalmanninella globotruncanoides
The stereoscopic images of the primary and secondary types of Th. brotzeni and Th. globotruncanoides show that the specimens are recrystallized and infilled; however, they retain the most important taxonomic characters. The collection includes the holotype (Fig. 2a–c) and one paratype (Fig. 2d–g) of Th. brotzeni, and the holotype (Fig. 2h–j) and three paratypes (Fig. 2k–u) of Th. globotruncanoides. Sigal (Reference Sigal1948) erected each species based on holotypes from Sidi Aissa in Algeria, whereas the paratype(s) are from Morocco. It is worth mentioning that the designation of primary types from different localities was common in earlier foraminiferal classification and, in agreement with Ando & Huber (Reference Ando and Huber2007), this practice is no longer acceptable because it leaves the status of the paratypes questionable, especially if not supported by an accurate record of their stratigraphic ranges.
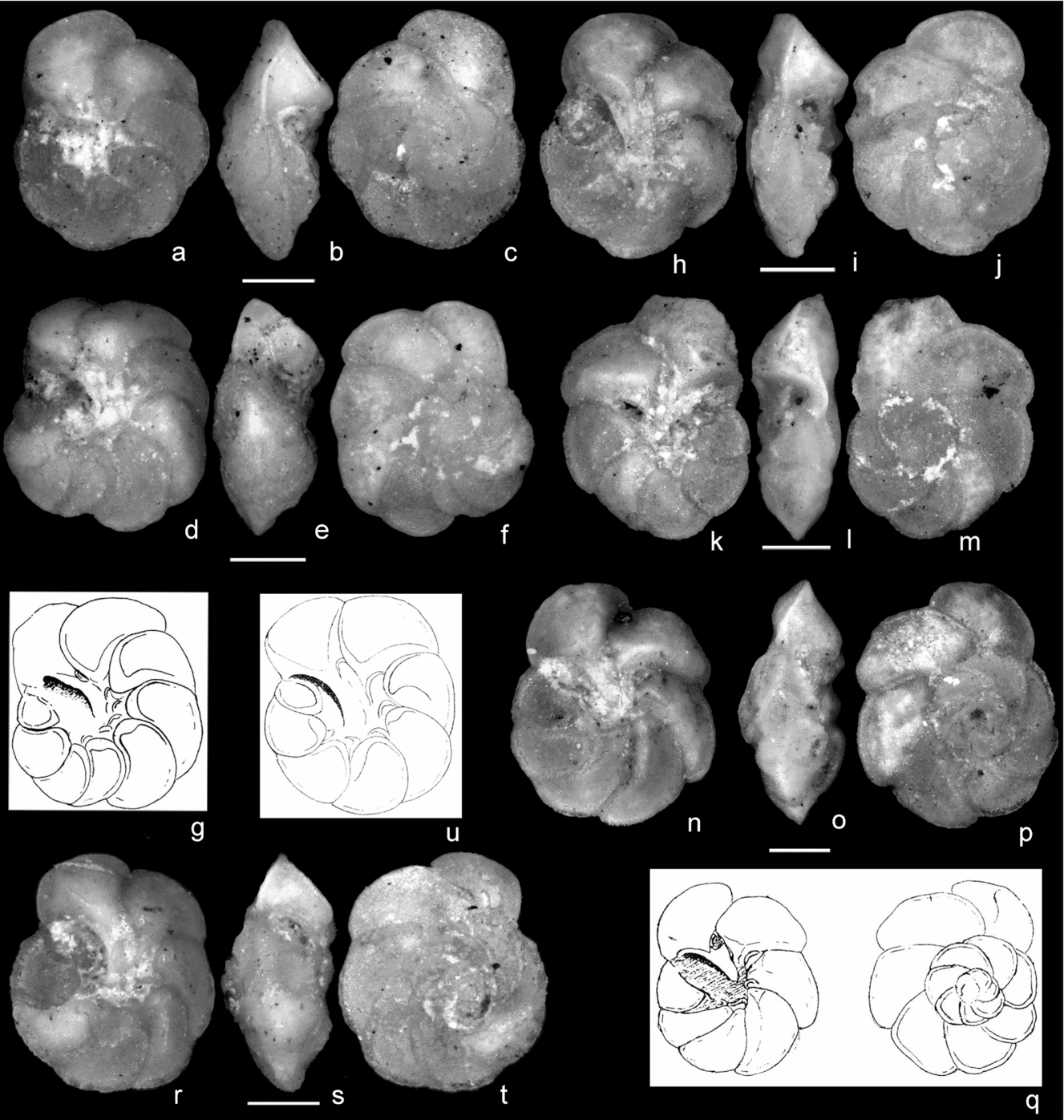
Figure 2. Sigal's type specimens deposited at the Musée National d’Histoire Naturelle in Paris. (a–c) Thalmanninella brotzeni, holotype F6 0843, Sigal, Reference Sigal1948, pl. I, fig. 5a–c, refigured in Sigal, Reference Sigal1952, fig. 25, from the type-level of Sidi Aissa, Algeria, middle Cenomanian, in (a) umbilical, (b) edge and (c) spiral view. (d–g) Thalmanninella brotzeni, paratype F6 0843, Sigal, Reference Sigal1948, pl. II, fig. 7 (original drawing reproduced in (g)), from the middle Cenomanian of Morocco, in (d) umbilical, (e) edge and (f) spiral view. The specimen is here interpreted as a transitional form to Th. greenhornensis (see text). (h–j) Thalmanninella globotruncanoides, holotype F6 0840, Sigal, Reference Sigal1948, pl. 1, fig. 4a–c, refigured in Sigal, Reference Sigal1952, fig. 24, from the type-level of Sidi Aissa, Algeria, middle Cenomanian, in (h) umbilical, (i) edge and (j) spiral view. (k–m) Thalmanninella globotruncanoides, paratype 4, Sigal, Reference Sigal1948, pl. II, fig. 4a, b, from the middle Cenomanian of Morocco, in (k) umbilical, (l) edge and (m) spiral view. (n–q) Thalmanninella globotruncanoides paratype 3, Sigal, Reference Sigal1948, pl II, fig. 3a, b (original drawings reproduced in (q)), from the middle Cenomanian of Morocco, in (n) umbilical, (o) edge and (p) spiral view. The specimen is here interpreted as a primitive form transitional to Th. greenhornensis (see text). (r–u) Thalmanninella globotruncanoides, paratype 5, Sigal, Reference Sigal1948, pl. II, fig. 5 (original drawing reproduced in (u)), from the middle Cenomanian of Morocco, in (r) umbilical, (s) edge and (t) spiral view. The specimen is possibly as an intermediate form between globotruncanoides and greenhornensis (see text). Scale bars = 100 μm
Caron & Premoli Silva (Reference Caron and Premoli Silva2007), after examination of the type materials of Th. globotruncanoides and Th. brotzeni deposited at the MNHN in Paris, considered the two species to be separate taxa based on the differences in growth rate, chamber morphology and position of the supplementary apertures. Because of the museum's curatorial policy at that time, Sigal's type materials could not be photographed and Caron & Premoli Silva (Reference Caron and Premoli Silva2007) re-described and redrew the holotypes of both Th. brotzeni and Th. globotruncanoides (p. 527, fig. 2), and illustrated by scanning electron microscope (SEM) only topotype material (p. 258, plate 1, figs 1, 2). According to Caron & Premoli Silva (Reference Caron and Premoli Silva2007), Th. globotruncanoides differs from Th. brotzeni by being larger in size, in having an almost plano-convex profile instead of symmetrically biconvex, more petaloid chambers on the spiral side and a faster growth rate, a shorter inner spire with the spiral suture lying in a marked depression, a larger and shallower umbilicus, a higher arched primary aperture and supplementary apertures migrating along the sutures out of the umbilical area in the last chambers of the last whorl.
These observations are slightly different from those by Ando & Huber (Reference Ando and Huber2007, p. 173), who examined the topotypes (metatypes) of Th. globotruncanoides and Th. brotzeni deposited in the Cushman Collection at the Smithsonian NMNH. In their opinion Th. globotruncanoides ‘diagnostically exhibits fish-hook ornamentation on the umbilical chamber surfaces in the later part of the last whorl, where it is illustrated by a combination of keeled periphery, raised umbilical suture, periumbilical ridge, and adumbilical ( = toward umbilicus) ridge parallel to the posterior chamber margin’. Ando & Huber's (Reference Ando and Huber2007) species concept fits with the diagnosis given by Robaszynski et al. (Reference Robaszynski, Amédro, Gonzalez-Donoso and Linares2008, p. 127) that stated ‘globotruncanoides has curved and raised sutures between all chambers’. Moreover, Ando & Huber (Reference Ando and Huber2007) did not exclude the possibility that Th. brotzeni may be a pre-adult given the size difference with Th. globotruncanoides (Caron & Premoli Silva, Reference Caron and Premoli Silva2007, p. 525).
Our re-examination of Sigal's holotypes of Th. globotruncanoides and Th. brotzeni, based on the new stereoscopic images, confirms the emended description by Caron & Premoli Silva (Reference Caron and Premoli Silva2007) and demonstrates that both Th. globotruncanoides and Th. brotzeni possess on the umbilical side slightly raised sutures on the first chambers of the last whorl becoming smoother and depressed in the last two chambers (Fig. 2a–c, h–j). Therefore, beside the morphological differences mentioned by Caron & Premoli Silva (Reference Caron and Premoli Silva2007), the most important distinguishing feature for a consistent species assignment is the position of the supplementary apertures, which are completely umbilical in Th. brotzeni, whereas in Th. globotruncanoides the supplementary apertures progressively migrate out of the umbilical area and become sutural on the last chamber.
The paratypes of Th. brotzeni and Th. globotruncanoides (Fig. 2d–g, k–m, n–p, r–u) were collected by Sigal (Reference Sigal1948) from an unclearly specified middle Cenomanian level in Morocco. The paratype of brotzeni (Fig. 2d–g) was illustrated by Sigal (Reference Sigal1948, plate II, fig. 7) only in umbilical view (here reproduced in Fig. 2g). In our opinion, this specimen cannot be attributed to brotzeni as it possesses nine chambers in the last whorl, which are banana-shaped and separated by fully depressed, strongly backwards-curved sutures in umbilical view. It possesses umbilical supplementary apertures, a short inner spire that is depressed with respect to the outer whorl, chambers on the spiral side from subpetaloid in the first three (possibly four) separated by slightly keeled sutures, to radially elongate and separated by keeled sutures, and an acute peripheral margin, keeled and asymmetrically biconvex with the spiral side close to flat and the umbilical side convex. Based on these features, this specimen looks to be transitional to Thalmanninella greenhornensis and does not belong either to brotzeni or globotruncanoides, as already suggested by Ando & Huber (Reference Ando and Huber2007, p. 172). A second specimen identified as a paratype of brotzeni and illustrated only in spiral and umbilical views by Sigal (Reference Sigal1948, plate II, fig. 6a, b) is not present on the microslide deposited in the repository.
Based on our observations of the paratypes of globotruncanoides, we consider paratype 4 (Fig. 2k–m) illustrated by Sigal (Reference Sigal1948, plate II, fig. 4a, b) to be similar to the holotype. On the contrary, globotruncanoides paratype 3 (Fig. 2n–q) illustrated by Sigal (Reference Sigal1948, plate II, fig. 3a, b, here reproduced in Fig. 2q) does not look like a typical globotruncanoides. In edge view and on the umbilical side, the first chambers of the last whorl are narrower and more elongate and separated by more depressed and very curved sutures. These features suggest paratype 3 is a primitive form transitional to Th. greenhornensis (Morrow, Reference Morrow1934). Paratype 5 (Fig. 2r–u) illustrated by Sigal (Reference Sigal1948, plate II, fig. 5), shows a symmetrically biconvex profile, an inner spire flush with the test and supplementary apertures only within the umbilicus (Fig. 2u). The shape of the first chambers in umbilical view as well as the growth rate and the petaloid shape of the last chambers in spiral view recall the features of globotruncanoides. It may represent an intermediate form between globotruncanoides and greenhornensis; however, this assumption is pending in the absence of a precise documentation of the stratigraphic distribution of the paratypes.
Emended descriptions of Th. brotzeni and Th. globotruncanoides are provided to stabilize the species concepts. In addition, the main distinguishing morphological features of the two species are summarized in Figure 3.

Figure 3. Distinguishing morphological features of Th. brotzeni and Th. globotruncanoides. Specimens are from sample 1050C-26R-3, 15–17 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean. See text for further explanation. Scale bar = 100 μm
Genus Thalmanninella Sigal, Reference Sigal1948, emended Gonzalez-Donoso, Linares & Robaszynski, Reference Gonzalez-Donoso, Linares and Robaszynski2007
Type species. Thalmanninella brotzeni Sigal, Reference Sigal1948
Thalmanninella brotzeni Sigal, Reference Sigal1948, emended Figure 2a–c (holotype MNHN F6 0843, Sidi Aissa, Algeria); Figure 4g–i (Mont Risou, France); Figure 4s–u (ODP 1050, Blake Nose Plateau, NW Atlantic Ocean).
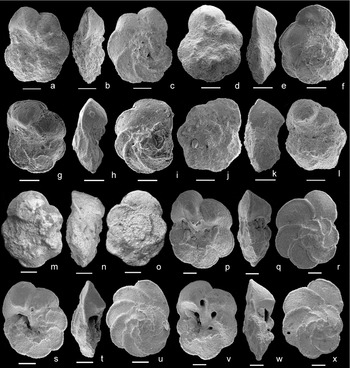
Figure 4. (a–c) Thalmanninella globotruncanoides, sample −36 m, Mont Risou, Hautes-Alpes, France, in (a) umbilical, (b) edge and (c) spiral view. (d–f) Thalmanninella globotruncanoides, sample −30 m, Mont Risou, Hautes-Alpes, France, in (d) umbilical, (e) edge and (f) spiral view. (g–i) Thalmaninnella brotzeni, sample −27 m, Mont Risou, Hautes-Alpes, France, in (g) umbilical, (h) edge and (i) spiral view. (j–l) Thalmanninella gandolfii, sample −48 m, Mont Risou, Hautes-Alpes, France, in (j) umbilical, (k) edge and (l) spiral view. (m–o) Thalmanninella gandolfii, holotype NHM Basel C 25557, level 55, Breggia section, Ticino, Switzerland, Luterbacher & Premoli Silva (Reference Luterbacher and Premoli Silva1962), in (m) umbilical, (n) edge and (o) spiral view. SEM image available on the online Mesozoic Planktonic Foraminiferal Taxonomic Dictionary at http://portal.chronos.org/gridsphere?cid1/4res_taxondb. (p–r) Thalmanninella gandolfii, sample 1050C-26R-5, 18–20 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (p) umbilical, (q) edge and (r) spiral view. (s–u) Thalmanninella brotzeni, sample 1050C-26R-3, 15–17 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (s) umbilical, (t) edge and (u) spiral view. (v–x) Thalmanninella globotruncanoides, sample 1050C-26R-3, 15–17 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (v) umbilical, (w) edge and (x) spiral view. Scale bars = 100 μm.
1948 Thalmanninella brotzeni Sigal, pl. I, fig. 5a–c, refigured in Sigal, Reference Sigal1952, fig. 25 (Sidi Aissa, Algeria).
2007 Thalmanninella brotzeni Sigal; Ando & Huber, pl. A1, fig. 2a–c (metatype USNM P3930, Sidi Aissa, Algeria).
2007 Thalmanninella brotzeni Sigal; Caron & Premoli Silva, pl. 1, fig. 1 (topotype, Sidi Aissa, Algeria).
Emended description. Test coiled in a low to moderate trochospire, almost equally biconvex; equatorial outline subcircular to ovoid, initially compact passing to very slightly lobate; in edge view peripheral margin acute and marked by a keel in all chambers of the last whorl; on spiral side 2.5–3 whorls, with 6–8 chambers in the last whorl, increasing gradually in size as added, crescentic in shape at least twice longer than wide except the last two chambers that become more petaloid; intercameral sutures curved backwards marked by slightly raised, finely beaded keel; on umbilical side chambers subtriangular to subtrapezoidal, initially slowly increasing in size, the last two chambers slightly larger and inflated; intercameral sutures fairly curved backwards marked by a very slightly raised keel to flush with test surface except the last one lying within a gentle depression that may mask the raised suture; intercameral keels extend to the umbilical edge forming a more raised periumbilical ridge, except on the last chamber that exhibits only an adumbilical inflation; umbilicus rather small and deep; primary aperture umbilical-extraumbilical as a medium high arch; a few supplementary apertures, bordered by a thin lip, are visible within the umbilical area; wall macroperforate and smooth.
Distinguishing features. Thalmanninella brotzeni differs from Th. globotruncanoides by having an almost equally biconvex profile, a rather compact less lobate outline, more crescentic chambers separated by strongly backwards-curved intercameral sutures in spiral view growing slowly in size, less prominent periumbilical ridges, a smaller and deeper umbilicus, and supplementary apertures only umbilical in position.
Remarks. This species has been regarded for a long time as a junior synonym of Th. globotruncanoides (Wonders, Reference Wonders1978, Reference Wonders1980; Robaszynski et al. Reference Robaszynski, Caron, Amédro, Dupuis, Hardenbol, Gonzalez-Donoso, Linares and Gartner1994, Reference Robaszynski, Amédro, Gonzalez-Donoso and Linares2008; Robaszynski & Caron, Reference Robaszynski and Caron1995; Gale et al. Reference Gale, Kennedy, Burnett, Caron and Kidd1996; Gonzalez-Donoso, Linares & Robaszynski, Reference Gonzalez-Donoso, Linares and Robaszynski2007).
Stratigraphic distribution. Uppermost part of the Th. appenninica Zone to the lower upper Cenomanian.
Geographical distribution. Algeria, Tunisia, Morocco, France, Central Italy, Israel and the western Atlantic Ocean.
Thalmanninella globotruncanoides (Sigal, Reference Sigal1948), emended
Figure 2h–j (holotype MNHN F6 0840, Sidi Aissa, Algeria); Figure 2k–m (paratype, Morocco); Figure 4a–f (Mont Risou, France); Figure 4v–x (ODP 1050, Blake Nose Plateau, NW Atlantic Ocean)
1948 Rotalipora globotruncanoides Sigal, pl. I, fig. 4a–c, refigured in Sigal, Reference Sigal1952, fig. 24 (Sidi Aissa, Algeria).
1993 Rotalipora (Thalmanninella) globotruncanoides (Sigal); Gonzalez-Donoso & Linares in Robaszynski et al., pl. XIX, figs 2, 4, 5, 9 (Tunisia).
2002 Rotalipora globotruncanoides Sigal; Bellier & Moullade, pl. 2, fig. 4–7 (Blake Nose Plateau, NW Atlantic Ocean).
2004 Rotalipora globotruncanoides Sigal; Kennedy et al., fig. 8, parts 13–15 (Mont Risou, France).
2007 Thalmanninella globotruncanoides (Sigal); Ando & Huber, pl. A1, fig. 1a–c (metatype USNM 387929, Sidi Aissa, Algeria).
2007 Thalmanninella globotruncanoides (Sigal); Caron & Premoli Silva, pl. 1, fig. 2a–c (topotype, Sidi Aissa, Algeria).
Emended description. Test coiled in a low trochospire, umbilico-convex; equatorial outline subcircular to ovoid, spiral side almost plane with inner spire very slightly elevated, peripheral margin acute in edge view and keeled on all chambers of the last whorl; on spiral side 2.5–3 whorls, with 6–7 chambers on the last whorl, initially crescentic then they become slightly petaloid, enlarging rather rapidly in size; surface of the last two chambers slightly concave in the central part giving an almost plane aspect to the trochospire; intercameral sutures curved and marked by a beaded keel slightly raised on the first chambers of the last whorl and becoming smoother in the last two chambers; spiral suture of the first whorls, possibly keeled, located in a gentle depression; on umbilical side chambers enlarging gradually but rather rapidly in size, subtrapezoidal with flat surface in the first three to four, the last three chambers becoming gradually more inflated; intercameral sutures initially gently curved and oblique with respect to the equatorial margin, then increasingly more oblique and markedly depressed; sutural beaded keels slightly visible and flush with test surface along the first four sutures, each extending to form a periumbilical ridge; sutural keels not visible or absent between the last two chambers; the last two more inflated chambers exhibit a prominent smooth fold of the chamber surfaces, located in the inner part of each chamber, that bifurcates from the umbilicus half way towards the periphery; primary aperture umbilical-extraumbilical as a medium high arch; umbilicus rather large and shallow, partially covered by a triangular, long porticus extending from the last chamber; supplementary apertures, bordered by a thin lip, are umbilical in the early outer chambers, then they progressively migrate out of the umbilicus along the inner part of the suture in the last three chambers, the last one being sutural; wall macroperforate and smooth.
Distinguishing features. Thalmanninella globotruncanoides differs from Th. brotzeni in having a more lobate outline and a faster growth rate, a folded surface in the last two inflated and more petaloid chambers, a larger and shallower umbilicus, and supplementary apertures migrating along the sutures and out of the umbilical area through ontogeny. It differs from Th. greenhornensis in having fewer chambers in the last whorl, an umbilico-convex profile, raised umbilical sutures only in the first chambers of the last whorl, and supplementary apertures migrating progressively out of the umbilical area, the last one being sutural. Th. greenhornensis is well differentiated by its numerous elongated chambers in the last whorl that curve backwards both in the spiral and umbilical sides and by having a symmetrically biconvex profile as discussed by Ando & Huber (Reference Ando and Huber2007).
Remarks. Kennedy et al. (Reference Kennedy, Gale, Lees and Caron2004) illustrated two specimens as Rotalipora globotruncanoides from Mont Risou (fig. 8, parts 8–12 and 13–15): the specimen in figure 8, parts 13–15, although distorted in spiral view, is close to the type material of Th. globotruncanoides (see also Ando & Huber, Reference Ando and Huber2007), while the second specimen illustrated in figure 8, parts 10–12, being also distorted, is less convincing.
Stratigraphic distribution. Base of the Cenomanian to the lower upper Cenomanian.
Geographical distribution. Algeria, Tunisia, Morocco, France, Central Italy, Spain and the western Atlantic Ocean.
3. Equivocal species concept of Pseudothalmanninella tehamaensis
The appearance level of Pseudothalmanninella tehamaensis ( = Rotalipora tehamaensis Marianos & Zingula, Reference Marianos and Zingula1966) is a secondary criterion for placing the Albian/Cenomanian boundary (Kennedy et al. Reference Kennedy, Gale, Lees and Caron2004). Pseudothalmanninella tehamaensis was described by Marianos & Zingula (Reference Marianos and Zingula1966) from Dry Creek in California from a single Albian level yielding Planomalina buxtorfi and Pseudothalmanninella ticinensis. In our opinion the primary and secondary types (Fig. 5a–f), although poorly preserved, fit well with Marianos & Zingula's original drawing and description of the species: ‘Test trochoid, biconvex, nearly circular, periphery only weakly lobate, single-keeled; chambers compressed, thickest at the umbilicus, early chambers of last whorl pustulose, later ones smooth, 6–8 in last whorl, increasing slowly in size, normally overlapping dorsally; spiral sutures curved, raised and thickened, curved back at the periphery forming a keel; umbilical sutures radial, depressed near outer edge, becoming raised and thickened near umbilicus, and in some specimens having a thickened ridge around the umbilicus connecting adjacent sutures; umbilicus narrow, deep; primary aperture a high umbilical-extraumbilical arch showing a lip; one secondary aperture, having a lip, per suture in the umbilicus. Dimensions: Greatest diameter of holotype 0.34 mm; least diameter 0.30 mm; thickness 0.16 mm’ (Marianos & Zingula, Reference Marianos and Zingula1966, p. 339).
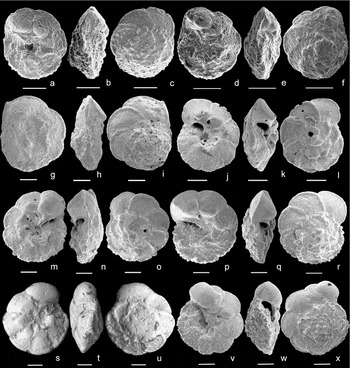
Figure 5. (a–c) Pseudothalmanninella tehamaensis, holotype USNM 641551, Marianos & Zingula, Reference Marianos and Zingula1966, Tehama County, California, Albian, in (a) umbilical, (b) edge and (c) spiral view. (d–f) Pseudothalmanninella tehamaensis, paratype 1 USNM 641552, Marianos & Zingula, Reference Marianos and Zingula1966, Tehama County, California, Albian, in (d) umbilical, (e) edge and (f) spiral view. SEM images of Ps. tehamaensis type material available on the online Mesozoic Planktonic Foraminiferal Taxonomic Dictionary at http://portal.chronos.org/gridsphere?cid1/4res_taxondb. (g–i) Pseudothalmanninella tehamaensis, sample −27 m, Mont Risou, Hautes-Alpes, France, in (g) umbilical, (h) edge and (i) spiral view. (j–l) Pseudothalmanninella tehamaensis, sample 1050C-27R-3, 8–10 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (j) umbilical, (k) edge and (l) spiral view. (m–o) Pseudothalmanninella tehamaensis, sample 1050C-27R-2, 103–105 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (m) umbilical, (n) edge and (o) spiral view. (p–r) Pseudothalmanninella tehamaensis, sample 1050C-27R-2, 103–105 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (p) umbilical, (q) edge and (r) spiral view. (s–u) Pseudothalmanninella ticinensis, in (s) umbilical, (t) edge and (u) spiral view; specimen illustrated by Gandolfi (Reference Gandolfi1942), pl. 4, fig. 11, bed 30, Breggia section, Ticino, Switzerland and deposited at the NHM in Basel. SEM image courtesy of B. T. Huber (Smithsonian Institution). (v–x) Pseudothalmanninella ticinensis, sample 1050C-27R-1, 73–76 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (v) umbilical, (w) edge and (x) spiral view. Scale bars = 100 μm.
Our identification of Ps. tehamaensis across the Albian/Cenomanian boundary interval at Mont Risou (Fig. 5g–i), and in the equivalent and well-preserved planktonic foraminiferal associations recorded at the Blake Nose Plateau (Fig. 5j–r) strictly follow the original description.
Remarkably, over the years the species concept of Ps. tehamaensis has changed and been erroneously interpreted probably because of its similarity with companion species and in the absence of SEM images of the type material (now available at the CHRONOS website, www.chronos.org). In fact, Ps. tehamaensis was either included in the range of variability of Rotalipora greenhornensis (Douglas, Reference Douglas1969; Masters, Reference Masters and Ramsay1977) or of Ps. ticinensis (Barr, Reference Barr1972) or of Rotalipora evoluta (Pessagno, Reference Pessagno1967). Consequently, the species was not considered valid by the European Working Group on Planktonic Foraminifera in the preparation of the Atlas (Robaszynski & Caron, Reference Robaszynski and Caron1979).
Afterwards, Gonzalez-Donoso & Linares in Robaszynski et al. (Reference Robaszynski, Caron, Amédro, Dupuis, Hardenbol, Gonzalez-Donoso, Linares and Gartner1994) called Ps. tehamaensis high-spired forms occurring at Kalaat Senan (Tunisia), in a short stratigraphic interval immediately above the appearance level of Th. globotruncanoides. These authors questioned the validity of the species, it being either very similar to Ps. ticinensis (Fig. 5s–x) or a possible intrapopulation morphotype within the Th. globotruncanoides species-group. The specimen illustrated as Ps. tehamaensis by Gonzalez-Donoso & Linares in Robaszynski et al. (Reference Robaszynski, Caron, Amédro, Dupuis, Hardenbol, Gonzalez-Donoso, Linares and Gartner1994, plate 20, fig. 1) is characterized by having straight and raised umbilical sutures, except on the ultimate chambers, and by possessing a distinctive periumbilical ridge and a slightly elevated trochospire. The validity of Ps. tehamaensis was reaffirmed by Caron in Kennedy et al. (Reference Kennedy, Gale, Lees and Caron2004, p. 30) and its lowest occurrence was used as a secondary marker for the identification of the base of the Cenomanian Stage at the Mont Risou GSSP. The specimen illustrated as Ps. tehamaensis (Kennedy et al. Reference Kennedy, Gale, Lees and Caron2004, fig. 8, parts 7–9) is only in partial agreement with the Ps. tehamaensis species concept adopted by Gonzalez-Donoso & Linares in Robaszynski et al. (Reference Robaszynski, Caron, Amédro, Dupuis, Hardenbol, Gonzalez-Donoso, Linares and Gartner1994), as it shows raised umbilical sutures only on the first two chambers of the ultimate whorl, a slightly developed periumbilical ridge and a high trochospire asymmetrically biconvex. However, since Ps. tehamaensis is a biconvex form and does not possess a high trochospire, its identification at Mont Risou is here revised. Moreover, to stabilize the species concept of Ps. tehamaensis we provide an emended description as follows.
Genus Pseudothalmanninella Wonders, Reference Wonders1978, emended Gonzalez-Donoso, Linares & Robaszynski, Reference Gonzalez-Donoso, Linares and Robaszynski2007
Type species. Globotruncana ticinensis forma tipica Gandolfi, Reference Gandolfi1942
Pseudothalmanninella tehamaensis (Marianos & Zingula, Reference Marianos and Zingula1966), emended
Figure 5a–c (holotype USMN 641551, Dry Creek, Tehama County, California); Figure 5d–f (paratype 1, USMN 641552, Dry Creek, Tehama County, California); Figure 5g–i (Mont Risou, France); Figure 5j–r (ODP 1050, Blake Nose Plateau, NW Atlantic Ocean)
1966 Rotalipora tehamaensis Marianos & Zingula, pl. 38, fig. 4a–c (Dry Creek, Tehama County, California).
not 1994 Pseudothalmanninella tehamaensis (Marianos & Zingula); Gonzalez-Donoso & Linares in Robaszynski et al., pl. 20, fig. 1 (Kalaat Senan, Tunisia).
not 2004 Rotalipora tehamaensis Marianos & Zingula; Caron in Kennedy et al., fig. 8, parts 7–9 (Mont Risou, France).
Emended description. Test trochospiral with a moderately elevated spire; equatorial outline circular, very weakly lobate, peripheral margin acute throughout with one keel; 2.5–3 whorls with 7–9 chambers in final whorl, uniformly and slowly increasing in size, trapezoidal on the umbilical side, petaloid to slightly crescentic in the last ones on the spiral side; in edge view peripheral margin initially close to spiroconvex passing to symmetrically biconvex in adult; on the umbilical side the first chambers of the last whorl are compressed and display a flat or even depressed surface, whereas the last two or three become progressively thicker and characteristically triangular in shape towards the umbilicus; straight and raised umbilical sutures on most chambers except between the last two where raised sutures, if present, may be lying within a gentle depression (raised sutures possibly masked and not visible); raised sutures on the spiral side curved and bending sharply backwards at the peripheral margin; umbilicus small and deep bordered by a distinctive periumbilical ridge; primary aperture umbilical-extraumbilical as a medium arch, supplementary apertures within the umbilical area; wall macroperforate and smooth.
Distinguishing features. Pseudothalmanninella tehamaensis differs from Ps. ticinensis (Fig. 5s–x) by having raised umbilical sutures and a distinctive periumbilical ridge and by possessing flattened instead of inflated chambers on the umbilical side. It differs from Pseudothalmanninella klausi (Lehmann, Reference Lehmann1966; see emendation by Spezzaferri & Caron, Reference Spezzaferri and Caron2008) in having a more compact equatorial outline, a lower trochospire, and raised keeled umbilical sutures in the early chambers of the last whorl, tending to form an extended periumbilical ridge instead of depressed sutures throughout and a faint periumbilical ridge confined to the early chambers.
Remarks. The specimen illustrated as Ps. tehamaensis by Gonzalez-Donoso & Linares in Robaszynski et al. (Reference Robaszynski, Caron, Amédro, Dupuis, Hardenbol, Gonzalez-Donoso, Linares and Gartner1994) shares most of the characters with the holotype and mainly conforms to the emended description; however, it possesses a last chamber that is too inflated and apparently not keeled. It may be interpreted either as an ancestral or an aberrant form of Ps. tehamaensis. Concerning the specimen illustrated by Caron in Kennedy et al. (Reference Kennedy, Gale, Lees and Caron2004), it is keeled throughout but in edge view it is more compressed than the type material, exhibits raised umbilical sutures only in the first two chambers and the periumbilical ridge is less prominent. On the basis of these observations, it may be interpreted as an ancestral form to Ps. tehamaensis or a juvenile specimen.
Phylogenetic relationships. Pseudothalmanninella tehamaensis is inferred to represent the final member of the Pseudothalmanninella lineage (Gonzalez-Donoso, Linares & Robaszynski, Reference Gonzalez-Donoso, Linares and Robaszynski2007). It is interpreted to have evolved from Ps. ticinensis (Fig. 5s–x) by acquiring raised umbilical sutures, developing a distinctive periumbilical ridge and by flattening the chambers on the umbilical side.
Stratigraphic distribution. Uppermost part of the Th. appenninica Zone to the lowermost part of the Th. globotruncanoides Zone. According to Marianos & Zingula (Reference Marianos and Zingula1966), the holotype co-occurs with Planomalina buxtorfi in a single level dated as Albian.
Geographic distribution. California, the western Atlantic Ocean, Tunisia and France.
4. Planktonic foraminifera index species at Mont Risou and Blake Nose Plateau
The planktonic foraminiferal assemblages across the GSSP boundary interval at Mont Risou have been restudied to document the stratigraphic distribution of Th. globotruncanoides, Th. brotzeni and Ps. tehamaensis using the species concepts revised in this study. We confirm the first appearance level of Th. globotruncanoides (Fig. 4a–f) at −36 m as in Kennedy et al. (Reference Kennedy, Gale, Lees and Caron2004), and document the first appearance of Th. brotzeni and Th. gandolfii (Luterbacher & Premoli Silva, Reference Luterbacher and Premoli Silva1962) (Fig. 4g–l) at sample −64 m. However, we have to remark that Th. brotzeni occurs discontinuously up-section and Th. globotruncanoides is rare in sample −36 m and, in agreement with previous data (Kennedy et al. Reference Kennedy, Gale, Lees and Caron2004), it becomes more common from sample −27 m up-section.
Planomalina buxtorfi and Pseudothalmanninella ticinensis disappear at −116 m and −60 m, respectively. Thalmanninella appenninica and Th. balernaensis occur discontinuously from the base to the top of the section studied. Pseudothalmanninella tehamaensis is very rare and shows a very short stratigraphic range (from sample −27 m to −19 m), whereas common transitional morphotypes between Ps. ticinensis and Ps. tehamaensis occur from sample −116 m to −27 m (Fig. 6).
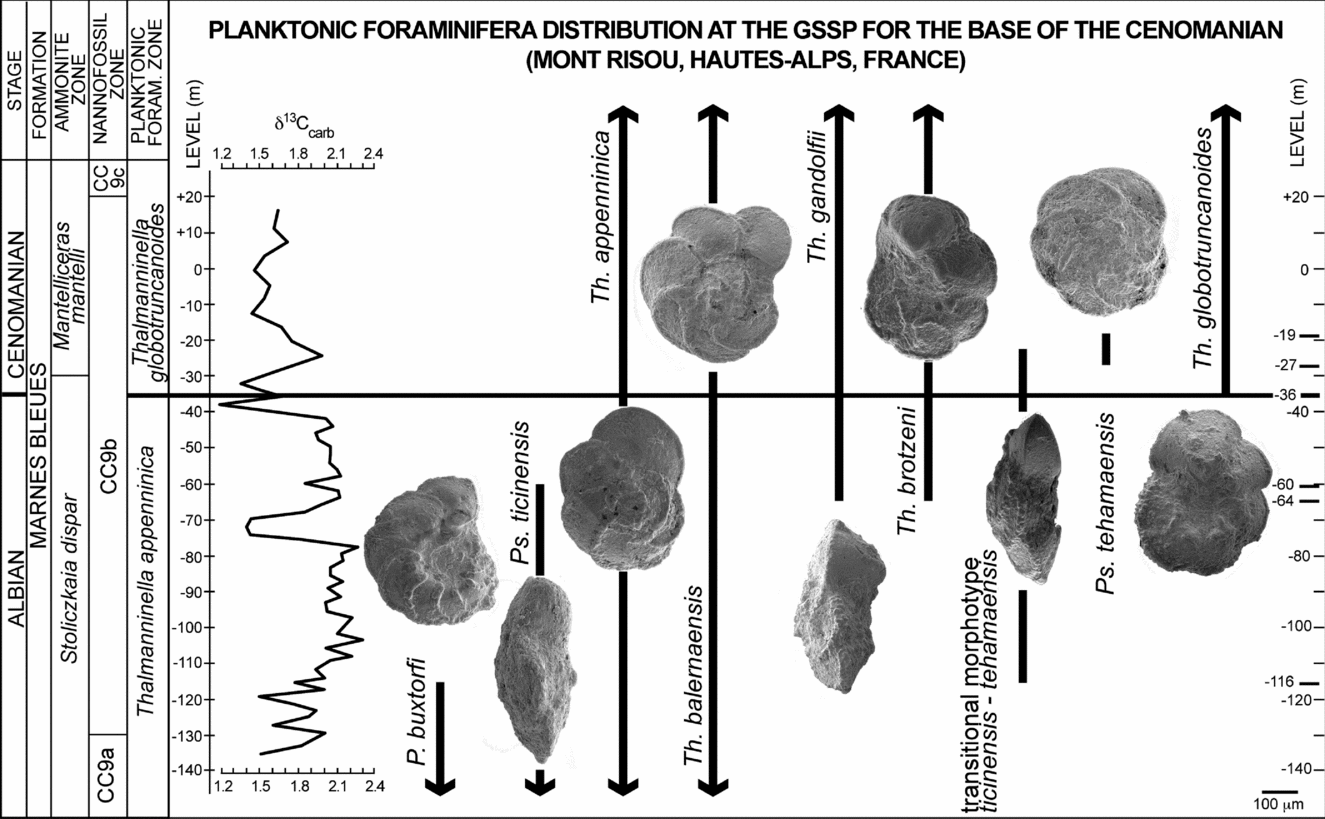
Figure 6. Distribution of selected planktonic foraminifera at the GSSP for the base of the Cenomanian Stage at Mont Risou (Hautes-Alps, France). Ammonite and nannofossils zonations according to Kennedy et al. (Reference Kennedy, Gale, Lees and Caron2004). Carbon isotope curve from Gale et al. (Reference Gale, Kennedy, Burnett, Caron and Kidd1996).
To test the reproducibility of the data, the sequence of bioevents observed across the Albian/Cenomanian boundary at Mont Risou is compared with that found throughout the equivalent stratigraphic interval at Blake Nose Plateau (ODP Leg 171B, North Western Atlantic Ocean) where the planktonic foraminiferal associations are better preserved (Petrizzo & Huber, Reference Petrizzo and Huber2006a ). In general, at Blake Nose the transitional morphotypes from Ps. ticinensis to Ps. tehamaensis, Th. gandolfii (Fig. 4p–r), Th. brotzeni (Fig. 4s–u) and Th. globotruncanoides (Fig. 4v–x) appear in the same stratigraphic order (Fig. 7) documented at Mont Risou. An exception is represented by Ps. tehamaensis, which is first recorded below the lowest occurrence of Th. globotruncanoides. This discrepancy is likely assignable to the rarity and poor preservation of specimens of Th. tehamaensis at Mont Risou preventing the accurate identification of its lowest appearance.

Figure 7. Distribution of selected planktonic foraminifera across the Albian/Cenomanian boundary interval at Hole 1050C Blake Nose Plateau (NW Atlantic Ocean). Nannofossil zonation from Watkins & Bergen (Reference Watkins and Bergen2003). Carbon isotope data according to Petrizzo et al. (Reference Petrizzo, Huber, Wilson and MacLeod2008).
However, the most relevant difference between the two localities is the different stratigraphic range shown by P. buxtorfi: at Blake Nose all the relevant planktonic foraminiferal appearances except for Th. globotruncanoides fall in stratigraphic levels still containing specimens of P. buxtorfi, whereas at Mont Risou P. buxtorfi shows an earlier disappearance and only co-occurs with the transitional Ps. ticinensis – Ps. tehamaensis morphotypes (Figs 6, 7).
Significantly, this finding highlights the diachroneity of the disappearance level of P. buxtorfi and confirms its unreliability for regional and global correlations. In fact, the extinction of P. buxtorfi was originally reported to fall above the appearance levels of Th. brotzeni and Th. globotruncanoides by Sigal (Reference Sigal1977) in his biozonal scheme of the Mediterranean area and by Collignon, Sigal & Grekoff (Reference Collignon, Sigal and Grekoff1979) from North Madagascar. These observations were later confirmed by Robaszynski et al. (Reference Robaszynski, Amédro, Gonzalez-Donoso and Linares2008) in their study of the planktonic foraminiferal assemblages across the upper Albian – lower Cenomanian stratigraphic interval in Central Tunisia.
Specimens similar to the paratype of Th. brotzeni (Fig. 2d–g) and paratype 3 of Th. globotruncanoides (Fig. 2n–q) here interpreted as related to Th. greenhornensis, have not been observed in the assemblages across the Albian/Cenomanian boundary at Mont Risou and Blake Nose.
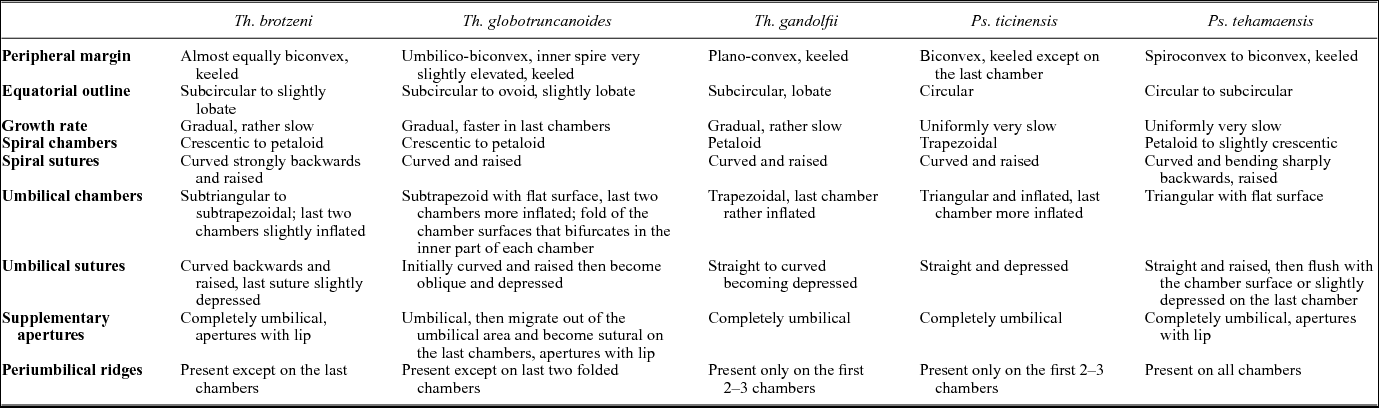
A summary of the main morphological features of Th. brotzeni, Th. globotruncanoides, Th. gandolfii, P. ticinensis and P. tehamaensis is presented in Table 1 to clarify the species concepts adopted in this study.
Table 1. Summary of the main morphological features of the single-keeled species occurring across the Albian/Cenomanian boundary interval.
The development of a continuous umbilical ridge and raised umbilical sutures is the basis for distinguishing Ps. tehamaensis from Ps. ticinensis; however, a consistent distinction between the two species is not always easy at Mont Risou, especially because Ps. tehamaensis shows a short stratigraphic range and is rare, whereas the transitional forms have a longer stratigraphic distribution and dominate in the assemblage (Fig. 6). The evolutionary transition from Ps. ticinensis to Ps. tehamaensis observed at Mont Risou is better documented at Blake Nose (Fig. 7). The transitional specimens from Ps. ticinensis to Ps. tehamaensis are characterized by showing the periumbilical ridge only on the first chambers of the last whorl and/or having the last two chambers slightly inflated so that the sutures seem depressed (Fig. 8a–r). Since the specimen illustrated as Ps. tehamaensis in Kennedy et al. (Reference Kennedy, Gale, Lees and Caron2004, fig. 8, parts 7–9) shows raised umbilical sutures only on the first two chambers of the ultimate whorl and a slightly developed periumbilical ridge, it could likely represent a transitional form in between Ps. ticinensis and Ps. tehamaensis similar to the specimen observed at Blake Nose (Fig. 8d–f).
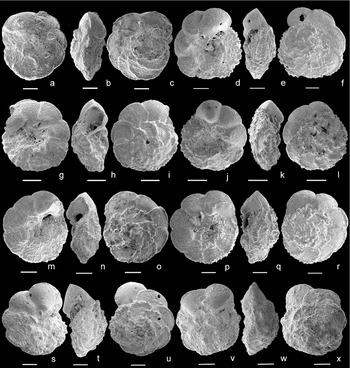
Figure 8. Transitional morphotypes between Ps. ticinensis and Ps. tehamaensis. Specimens with the periumbilical ridge developed only on the first chambers of the last whorl and slightly inflated chambers: (a–c) sample −27 m, Mont Risou, Hautes-Alpes, France, in (a) umbilical, (b) edge and (c) spiral view; (d–i) sample 1050C-27R-2, 103–105 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (d, g) umbilical, (e, h) edge and (f, i) spiral view; (j–l) sample 1050C-27R-3, 105–107 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (j) umbilical, (k) edge and (l) spiral view; (m–o) sample 1050C-27R-5, 4.5–6.5 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (m) umbilical, (n) edge and (o) spiral view; (p–r) sample 1050C-26R-5, 18–20 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (p) umbilical, (q) edge and (r) spiral view. High-spired specimens included in the variability of Ps. ticinensis: (s–u) sample 1050C-27R-3, 8–10 cm, Leg 171B Blake Nose Plateau, NW Atlantic Ocean, in (s) umbilical, (t) edge and (u) spiral view; (v–x) sample −116 m, Mont Risou, Hautes-Alpes, France, in (v) umbilical, (w) edge and (x) spiral view. Scale bars = 100 μm.
Distinctive high-spired morphotypes (Fig. 8s–x) have been observed close to the disappearance level of P. buxtorfi at Mont Risou (sample −116 m) and associated with common P. buxtorfi at Blake Nose (sample ODP 1050C-27R-3, 8–10 cm). These morphotypes are similar to the high-spired Ps. klausi described by Lehmann (Reference Lehmann1966) and recently revised and illustrated by SEM by Spezzaferri & Caron (Reference Spezzaferri and Caron2008), but differ in the absence of the periumbilical ridges. Moreover, Ps. klausi, tentatively considered as an intermediate form between P. ticinensis and P. tehamaensis (see Gonzalez-Donoso, Linares & Robaszynski, Reference Gonzalez-Donoso, Linares and Robaszynski2007), has not been observed in the studied assemblages. We believe that the high-spired morphotypes observed at Mont Risou and Blake Nose strongly resemble Ps. ticinensis, being characterized by having depressed umbilical sutures throughout. To date, we include the high-spired specimens within the Ps. ticinensis population as they are rare and do not show a significant distribution, being unrelated to precise stratigraphic levels.
5. Conclusions
Re-examination of the planktonic foraminiferal assemblages across the GSSP of the base of the Cenomanian Stage at Mont Risou allows revision of the sequence of appearances of the planktonic foraminifera index taxa, as follows (Fig. 6):
-
(1) The evolutionary transition from Ps. ticinensis to Ps. tehamaensis is very distinctive and is documented from the disappearance of P. buxtorfi to the lower range of Th. globotruncanoides;
-
(2) Th. brotzeni appears 28 m below the appearance level of Th. globotruncanoides; Th. brotzeni and Th. globotruncanoides are consistently identified as separate species, but the latter is very rare at the beginning of its stratigraphic range.
The species identifications and their stratigraphic distributions at Mont Risou are in good agreement with the data collected from Blake Nose Plateau in the North Western Atlantic Ocean. An exception is the disappearance level of P. buxtorfi that falls in a stratigraphic level already containing the transitional morphotype Ps. ticinensis – Ps. tehamaensis, Th. gandolfii and Th. brotzeni (Fig. 7) at Blake Nose. This observation causes additional uncertainty regarding the reliability of the last occurrence level of P. buxtorfi for regional and global correlations. A second exception is the lowest appearance of Ps. tehamaensis: at Blake Nose it precedes the lowest occurrence of Th. globotruncanoides, whereas at Mont Risou it appears 9 m above the appearance of Th. globotruncanoides and shows a very short stratigraphic distribution. This discrepancy, probably related to the rarity and poor preservation of Th. tehamaensis specimens at Mont Risou, confirms its unreliability for correlations. Therefore, Th. tehamaensis should not be used anymore as a secondary criterion for placing the Albian/Cenomanian boundary.
Finally, the outstanding question regarding the accurate identification of the index species Th. globotruncanoides is here solved. However, we have to remark on the difficulties related to the identification of the first representative of a new species when it is very rare and shows morphological characters clearly visible only in well-preserved specimens.
Acknowledgements
We wish to thank Annachiara Bartolini, curator at the Musée National d’Histoire Naturelle in Paris, who kindly provided us with the images of Sigal's type material deposited at the museum. Our thanks go to Jim Kennedy (University of Oxford) and Eliza Howlett (collections manager of the Earth Collections at Oxford University Museum of Natural History) for making available the original rock samples of the Mont Risou stratotype section. Careful reviews by Eduardo Koutsoukos and an anonymous reviewer helped to improve the quality of the manuscript. Thanks to Agostino Rizzi (IDPA-CNR, Milano) for operating the SEM and to Dario Soldan and Francesca Falzoni (University of Milan) for processing the samples. Financial support for this research was provided by PUR 2008 (University of Milan). MRP was partially funded through MIUR (Italian Ministry of University and Research) PRIN grant 2010–2011 (2010X3PP8J_001).









