1. Introduction
The Cambrian Period is characterized by four positive and six negative excursions in the marine δ13C record that were related to eustatic sea-level changes, perturbations in the oceanic carbon cycle and, as a consequence, extinctions and evolutionary radiations in the Cambrian fauna (Peng et al. Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004; Babcock et al. Reference Babcock, Peng, Geyer and Shergold2005; Zhu, Babcock & Peng, Reference Zhu, Babcock and Peng2006; Peng, Babcock & Cooper, Reference Peng, Babcock, Cooper, Gradstein, Ogg, Schmitz and Ogg2012). One of the prominent positive excursions, the Steptoean positive carbon isotope excursion (SPICE), is situated at the base of the Paibian Stage (Furongian Series; Saltzman, Runnegar & Lohmann, Reference Saltzman, Runnegar and Lohmann1998; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000). It has been identified in carbonate and organic-rich successions of slope and platform environments of Antarctica, Argentina, Australia, England, Kazakhstan, Newfoundland, North and South China, Siberia, Sweden and the USA (e.g. Glumac & Walker, Reference Glumac and Walker1998; Saltzman, Runnegar & Lohmann, Reference Saltzman, Runnegar and Lohmann1998; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000, Reference Saltzman, Young, Kump, Gill, Lyons and Runnegar2011; Peng et al. Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004; Zhu et al. Reference Zhu, Zhang, Li and Yang2004; Lindsay et al. Reference Lindsay, Kruse, Green, Hawkins, Brasier, Cartlidge and Corfield2005; Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011; Kouchinsky et al. Reference Kouchinsky, Bengtson, Gallet, Korovnikov, Pavlov, Runnegar, Shields, Veizer, Young and Ziegler2008; Sial et al. Reference Sial, Peralta, Ferreira, Toselli, Aceñolaza, Parada, Gaucher, Alonso and Pimentel2008, Reference Sial, Peralta, Gaucher, Toselli, Ferreira, Frei, Parada, Pimentel and Pereira2013; Ahlberg et al. Reference Ahlberg, Axheimer, Babcock, Eriksson, Schmitz and Terfelt2009; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Chen et al. Reference Chen, Chough, Han and Lee2011, Reference Chen, Chough, Lee and Han2012; Woods et al. Reference Woods, Wilby, Leng, Rushton and Williams2011; Ng, Yuan & Lin, Reference Ng, Yuan and Lin2014). In carbonate successions the SPICE is characterized by a shift of +4‰ to +6‰ in δ13Ccarb to peak values of about +5‰ (Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Woods et al. Reference Woods, Wilby, Leng, Rushton and Williams2011). A similar positive excursion was observed in organic-rich sequences where δ13Corg values increase by up to 3‰, culminating in a maximum value of about −28‰ (Ahlberg et al. Reference Ahlberg, Axheimer, Babcock, Eriksson, Schmitz and Terfelt2009; Woods et al. Reference Woods, Wilby, Leng, Rushton and Williams2011). Stratigraphically, the SPICE is well calibrated by the first appearance date (FAD) of Glyptagnostus reticulatus (base of the Paibian Stage) below and the FAD of Irvingella species above the δ13Ccarb peak, therefore allowing its global correlation (Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000). Furthermore, in Laurentia its maximum peak broadly coincides with: (1) the Sauk II – Sauk III hiatus (Palmer, Reference Palmer and Taylor1981; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000, Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) as well as (2) a distinct maximum in the biotic diversity in the Pterocephaliid biomere boundary (Rowell & Brady, Reference Rowell and Brady1976; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Peng et al. Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004).
A multitude of interpretations of the SPICE event exists considering increased weathering of terrestrial material or marine 13C-enriched carbonates, leading to an increased burial of 12C-enriched organic carbon under anoxic/euxinic conditions or enhanced sedimentation rates, respectively (Kump et al. Reference Kump, Arthur, Patzkowsky, Gibbs, Pinkus and Sheehan1999; Kump & Arthur, Reference Kump and Arthur1999; Saltzman, Runnegar & Lohmann, Reference Saltzman, Runnegar and Lohmann1998; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000, Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004; Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009). Each of these processes would result in a more positive δ13C signature of ambient seawater. However, the driving mechanisms behind the event are still poorly understood.
Considering the SPICE event as a consequence of weathering and changes in sea-level and climate, further changes in seawater chemistry (other than the perturbation of the global carbon cycle) should have occurred. Indeed, 87Sr-enriched Sr isotope values are typical for the Furongian, indicating increased rates of erosion and continental silicate weathering (Montañez et al. Reference Montañez, Banner, Osleger, Borg and Bosserman1996, Reference Montañez, Osleger, Banner, Mack and Musgrove2000; Denison et al. Reference Denison, Koepnick, Burke and Hetherington1998; Kouchinsky et al. Reference Kouchinsky, Bengtson, Gallet, Korovnikov, Pavlov, Runnegar, Shields, Veizer, Young and Ziegler2008). In addition, the SPICE is often associated with positive excursions in the sulphur isotopic composition of carbonate-associated sulphate (δ34SCAS) and sedimentary pyrite (i.e. chromium-reducible sulphur; δ34SCRS) (Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). The coupling of paired carbon and sulphur isotope trends is interpreted to reflect an elevated rate of organic carbon and pyrite burial during widespread euxinic conditions in Furongian oceans (Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). Carbon and sulphur isotope mass-balance models further indicate that the positive excursions in δ13C and δ34S are associated with a significant rise in the atmospheric O2 level (Saltzman et al. Reference Saltzman, Young, Kump, Gill, Lyons and Runnegar2011).
2. Geological setting
The Karatau Range of southern Kazakhstan represents the north-western prolongation of the Tien Shan Mountains, situated between the Chu-Sarysu Basin in the northeast and the Syrdarya Basin in the southwest (Alexeiev et al. Reference Alexeiev, Cook, Buvtyshkin and Golub2009; Fig. 1a). It is separated into the north-western Bolshoi (Greater) Karatau and the south-eastern Malyi (Lesser) Karatau ranges. Fossiliferous Cambrian – Early Ordovician carbonate deposits of the Malyi Karatau Range were accumulated on the seaward prograding margin of an isolated carbonate platform which belongs to the Aisha-Bibi seamount (width of the mount c. 40 km; Cook et al. Reference Cook, Taylor, Zhemchuzhnikov, Apollonov, Ergaliev, Sargaskaev, Dubinina, Cooper and Stevens1991; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Ergaliev et al. Reference Ergaliev, Zhemchuzhnikov, Ergaliev, Bassett, Popov and Holmer2009). The carbonate succession is exposed in several fault-bounded blocks and shows no or only minor tectonic dislocation (Holmer et al. Reference Holmer, Popov, Koneva and Bassett2001; Alexeiev et al. Reference Alexeiev, Cook, Buvtyshkin and Golub2009).

Figure 1. (a) Simplified map of Kazakhstan. The grey square marks the location of the working area in the surroundings of Zhanatas (southern Kazakhstan). (b) Map of the working area showing the topographic position of Kyrshabakty section (based on Google Earth, December 2014).
The Kyrshabakty section is situated at the Kyrshabakty river, c. 20 km east of the town of Zhanatas (Fig. 1b). The base of the more than 600-m-thick succession is located at 43°32′02″N and 69°57′28″E at an elevation of c. 500 m. Coordinates of the section have been previously misdated by Ergaliev et al. (Reference Ergaliev, Zhemchuzhnikov, Ergaliev, Bassett, Popov and Holmer2009) and Peng et al. (Reference Peng, Babcock, Cooper, Gradstein, Ogg, Schmitz and Ogg2012) as located northeast of Zhanatas at a longitude of 69°51′28″E. The section was investigated under multiple aspects, encompassing detailed analyses on litho- (Cook, et al. Reference Cook, Taylor, Zhemchuzhnikov, Apollonov, Ergaliev, Sargaskaev, Dubinina, Cooper and Stevens1991) and biofacies (Holmer et al. Reference Holmer, Popov, Koneva and Bassett2001; Ergaliev, Reference Ergaliev1980, Reference Ergaliev and Taylor1981, Reference Ergaliev1992; Ergaliev & Ergaliev, Reference Ergaliev, Ergaliev and Daukeev2004, Reference Ergaliev and Ergaliev2008) and carbon isotopes (Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000). Several concepts for a litho- and biostratigraphic subdivision exist for the Kyrshabakty section, which have been significantly modified during the last three decades (for more information see Ergaliev, Reference Ergaliev1980; Holmer et al. Reference Holmer, Popov, Koneva and Bassett2001; Ergaliev & Ergaliev, Reference Ergaliev and Ergaliev2008; Ergaliev et al. Reference Ergaliev, Zhemchuzhnikov, Ergaliev, Bassett, Popov and Holmer2009). Here, we apply the most recent trilobite zonation (Ergaliev et al. Reference Ergaliev, Zhemchuzhnikov, Ergaliev, Bassett, Popov and Holmer2009) and the subdivision of the sedimentary succession into informal lithostratigraphic units (units I–XII) as established by Ergaliev (Reference Ergaliev1980). At Kyrshabakty, only 9 (units IV–XII) of the 12 informal units are exposed. The basal units IV and V (up to 100 m in thickness) are composed of dark laminated argillaceous limestone, limestone and carbonate claystone representing a basin-plain environment (Fig. 2). Upsection, units VI–XI (c. 490 m in thickness) are dominated by dark laminated limestone, nodular limestone and carbonate breccias, interpreted as turbidites of carbonate submarine fan deposits (Fig. 2). Grey to brown dolostone, dolomitic limestone and (partly oolitic) limestone are exposed in unit XII, and reflect a shallow-subtidal lagoonal or tidal-flat environment. According to Cook et al. (Reference Cook, Taylor, Zhemchuzhnikov, Apollonov, Ergaliev, Sargaskaev, Dubinina, Cooper and Stevens1991), Ergaliev & Ergaliev (Reference Ergaliev and Ergaliev2008) and Ergaliev et al. (Reference Ergaliev, Zhemchuzhnikov, Ergaliev, Bassett, Popov and Holmer2009), the sedimentary succession of Kyrshabakty section was accumulated on the north-eastern flank (recent orientation) of the Aisha-Bibi seamount. It covers the stratigraphic interval ranging from the undefined Cambrian Series 3/Stage 5 (uppermost Peronopsis ultimus zone, known as ‘Lenan’ Stage in Kazakh nomenclature; Geyer & Shergold, Reference Geyer and Shergold2000; Ergaliev et al. Reference Ergaliev, Zhemchuzhnikov, Ergaliev, Bassett, Popov and Holmer2009) to the Furongian Series/undefined Stage 10 (Micragnostus mutabilis-Lophosaukia zones, known as Batyrbaian Stage of the Kazakh nomenclature; Ergaliev et al. Reference Ergaliev, Zhemchuzhnikov, Ergaliev, Bassett, Popov and Holmer2009).

Figure 2. Stratigraphic column and geochemical data for the Kyrshabakty section (southern Kazakhstan). Placement of international and regional stratigraphic subdivision is based on Ergaliev & Ergaliev (Reference Ergaliev and Ergaliev2008), Ergaliev et al. (Reference Ergaliev, Zhemchuzhnikov, Ergaliev, Bassett, Popov and Holmer2009) and Geyer & Shergold (Reference Geyer and Shergold2000). A. acutatus – Acutatagnostus acutatus; G. reticulatus – Glyptagnostus reticulatus; I. inexpectans – Innitagnostus inexpectans; I. tropica – Irvingella tropica; I. ivshini – Ivshinagnostus ivshini; P. ultimus – Peronopsis ultimus; P. vastulus – Pseudagnostus vastulus; P. punctuosus – Ptychagnostus punctuosus.
Within the interval covering the Glyptagnostus stolidotos to Pseudagnostus vastulus-Irvingella tropica trilobite zones (informal units VII and VIII), the SPICE event was identified by Saltzman et al. (Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000).
Here, we present the sulphur isotope values of carbonate-associated sulphate (δ34SCAS) and chromium-reducible sulphur (δ34SCRS) of 58 carbonate samples of the Kyrshabakty section, covering the middle Drumian–Jiangshanian stages (Fig. 2). In addition, samples were analysed for their carbonate carbon (δ13Ccarb) and oxygen (δ18Ocarb) isotopes, and their elemental concentrations (Ca, Fe, Mg, Mn and Sr). This extensive dataset provides the unusual opportunity to study the relationships between marine carbon and sulphur cycles in context with environmental, biological and climate changes during Furongian time. From chemostratigraphic studies and an in-depth evaluation of the biostratigraphic constraints, we try to unravel the processes and conditions which forced the parallel excursions in δ13C and δ34S during the SPICE interval.
3. Material and methods
Sample preparation and analytical work were conducted in the laboratories of the universities of Münster and Kiel using standard procedures. In a first step, contaminations such as calcite veins or weathering crusts were removed. Bulk-rock samples were subsequently powdered to <63 μm using a tungsten carbide ring and puck mill, delivering whole-rock material for isotope and element analyses.
δ13Ccarb and δ18Ocarb analyses were performed automatically using a GasBench II connected via a ConFlow-II Interface to a Finnigan MAT DeltaPlusXL. Delta values are reported relative to the V-PDB standard. The analytical reproducibility was generally better than 0.1‰ for δ13Ccarb and 0.2‰ for δ18Ocarb.
Extraction of carbonate-associated sulphate (CAS) from carbonate rocks was performed by applying the rigid extraction protocol of Wotte, Shields-Zhou & Strauss, (Reference Wotte, Shields-Zhou and Strauss2012) and Wotte et al. (Reference Wotte, Shields-Zhou and Strauss2012), known to deliver the most accurate δ34SCAS values. Up to 694 g per sample was used to analyse CAS concentration and isotopic composition (Table 1). In addition to the CAS extraction, the concentration and isotopic composition of chromium-reducible sulphur (CRS or pyrite sulphur) were determined. Analyses of CRS are essential in order to obtain information about alteration of CAS due to sulphate oxidation and/or bacterial sulphate reduction (Wotte, Strauss & Sundberg, Reference Wotte, Strauss and Sundberg2011; Wotte, Shields-Zhou & Strauss, Reference Wotte, Shields-Zhou and Strauss2012; Wotte et al. Reference Wotte, Shields-Zhou and Strauss2012). CRS was extracted from 0.8–37.7 g of dried insoluble residue subsequent to CAS extraction, basically following the extraction procedure described by Canfield et al. (Reference Canfield, Raiswell, Westrich, Reaves and Berner1986). δ34SCAS and δ34SCRS were measured using a Thermo-Finnigan Delta Plus mass spectrometer connected to an Elemental Analyser (EA-IRMS). Results are reported in the usual delta notation as per mil (‰) difference to the V-CDT standard. Reproducibility was better than ±0.3‰.
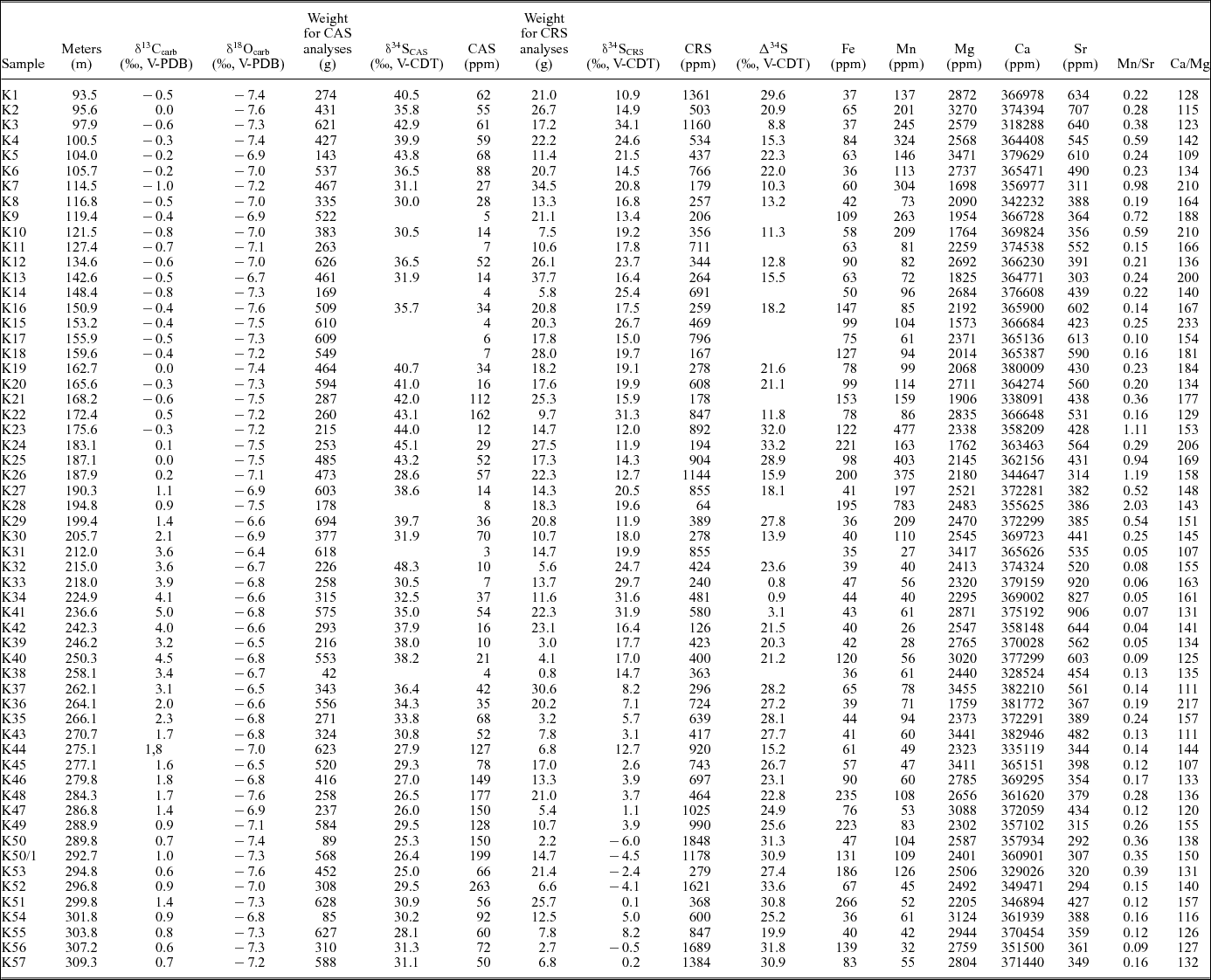
Table 1. Isotopic and elemental composition and initial weights for CAS and CRS analyses of samples from Kyrshabakty section.
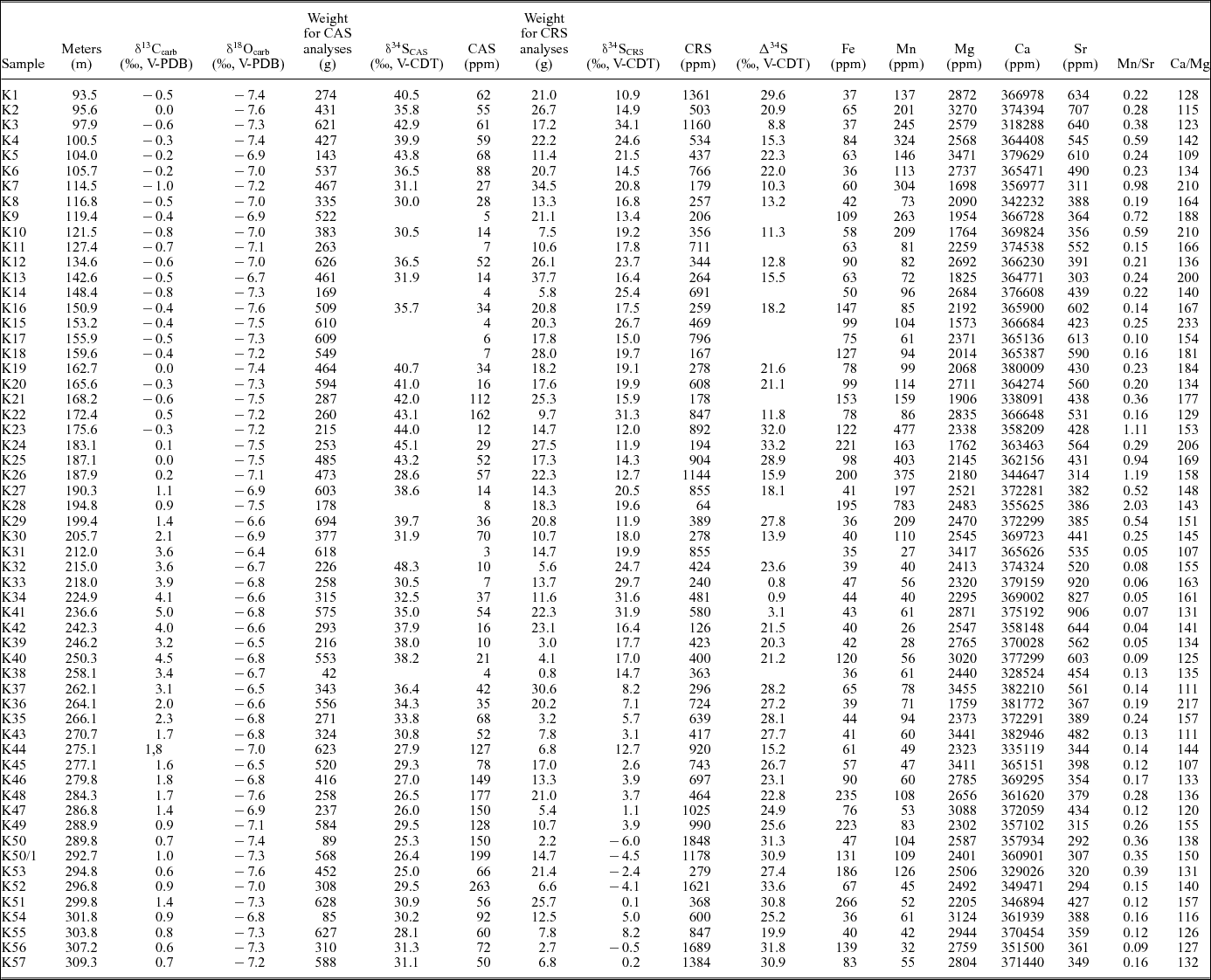
In order to evaluate carbonate diagenesis, elemental concentrations (Ca, Fe, Mg, Mn and Sr) were measured (Wotte, Strauss & Sundberg, Reference Wotte, Strauss and Sundberg2011; Wotte, Shields-Zhou & Strauss, Reference Wotte, Shields-Zhou and Strauss2012; Wotte et al. Reference Wotte, Shields-Zhou and Strauss2012). Subsequent to the reaction of 1 g of sample material with 20 mL of acetic acid (2%), suspension was filtered after 48 hours of reaction time. The filtrate was analysed for their elemental abundances by simultaneous inductively coupled plasma optical emission spectrometry (ICP-OES, SPECTRO CIROS SOP) after calibration with matrix-matched standards. Analytical precision was better than 0.5%. Accuracy was monitored by the analysis of USGS MAG-1 standard and found to be within 1% of recommended values (GeoReM online data base). Elemental concentrations were corrected for the insoluble residue and are reported in ppm.
4. Results
δ13Ccarb data vary between −1.0‰ and +0.5‰ within the informal units VI and VII (Goniagnostus nathorsti – Glyptagnostus stolidotos zones), increase within the Glyptagnostus stolidotos and Innitagnostus inexpectans-Prochuangia biozones (middle–upper part of unit VII) towards +4.1‰, and finally reach a maximum value of +5.0‰ in the lower Acutatagnostus acutatus-Erixanium biozone (upper part of unit VII) (Fig. 2; Table 1). Subsequently, δ13Ccarb values continuously decrease upsection, reaching a minimum of +0.6‰ (sample K56) at the top of the measured section. The establishment of a new baseline after the prominent excursion is discernible with post-excursion δ13Ccarb values of c. 1–3‰ more positive than data prior to the excursion.
An almost identical trend is described by the δ18Ocarb data, increasing from minimum values of −7.6‰ towards a maximum of −6.5‰ within the Glyptagnostus stolidotos to the Acutatagnostus acutatus-Erixanium biozones (Fig. 2; Table 1). The data continuously decrease towards a minimum of −7.6‰ near the top of the section.
CAS concentrations vary between 3 ppm and 263 ppm (Fig. 2; Table 1). Respective δ34SCAS values range from 25.0‰ to 48.3‰ showing two distinctive positive excursions within the lower part of unit VII (Kormagnostus simplex – Glyptagnostus stolidotos biozones; maximum 45.1‰ at K24) and the upper part of unit VII – basal unit VIII (Acutatagnostus acutatus-Erixanium to Pseudagnostus vastulus-Irvingella tropica biozones; maximum 38.2‰ at K40). Both positive trends are separated by a few data points exhibiting a high variability between 28.6‰ (K26) and 48.3‰ (K32). Starting from the second positive excursion (38.2‰ at K40), δ34SCAS values decrease rapidly, culminating in a minimum of 25.0‰ (Fig. 2; Table 1). δ34SCRS data fluctuate between −6.0‰ at the top and 34.1‰ at the base of the succession. δ34SCRS of the lower part of the section (up to the base of the Glyptagnostus reticulatus zone) vary between 10.9‰ and 34.1‰. A subsequent increase in δ34SCRS values culminates in a maximum peak of 31.9‰ (basal Acutatagnostus acutatus-Erixanium biozone) followed by a rapid drop to −6.0‰ (Fig. 2; Table 1). Corresponding CRS concentrations range from 126 ppm to 1848 ppm with no clear stratigraphic trend. Calculated Δ34S values (δ34SCAS – δ34SCRS) range from 0.8‰ to 33.6‰ (Fig. 2; Table 1). The post-excursion δ34SCAS and δ34SCRS baselines are significantly less positive than the sulphur isotope values before the events (Fig. 2). Upsection, both curves show a principle trend in δ34S towards less positive values.
Elemental concentrations show no major variation across the succession with values of 35–266 ppm for Fe, 26–783 ppm for Mn, 1573–3471 ppm for Mg, 292–920 ppm for Sr and 31.8–38.3 wt% for Ca (Table 1).
5. Diagenetic constraints
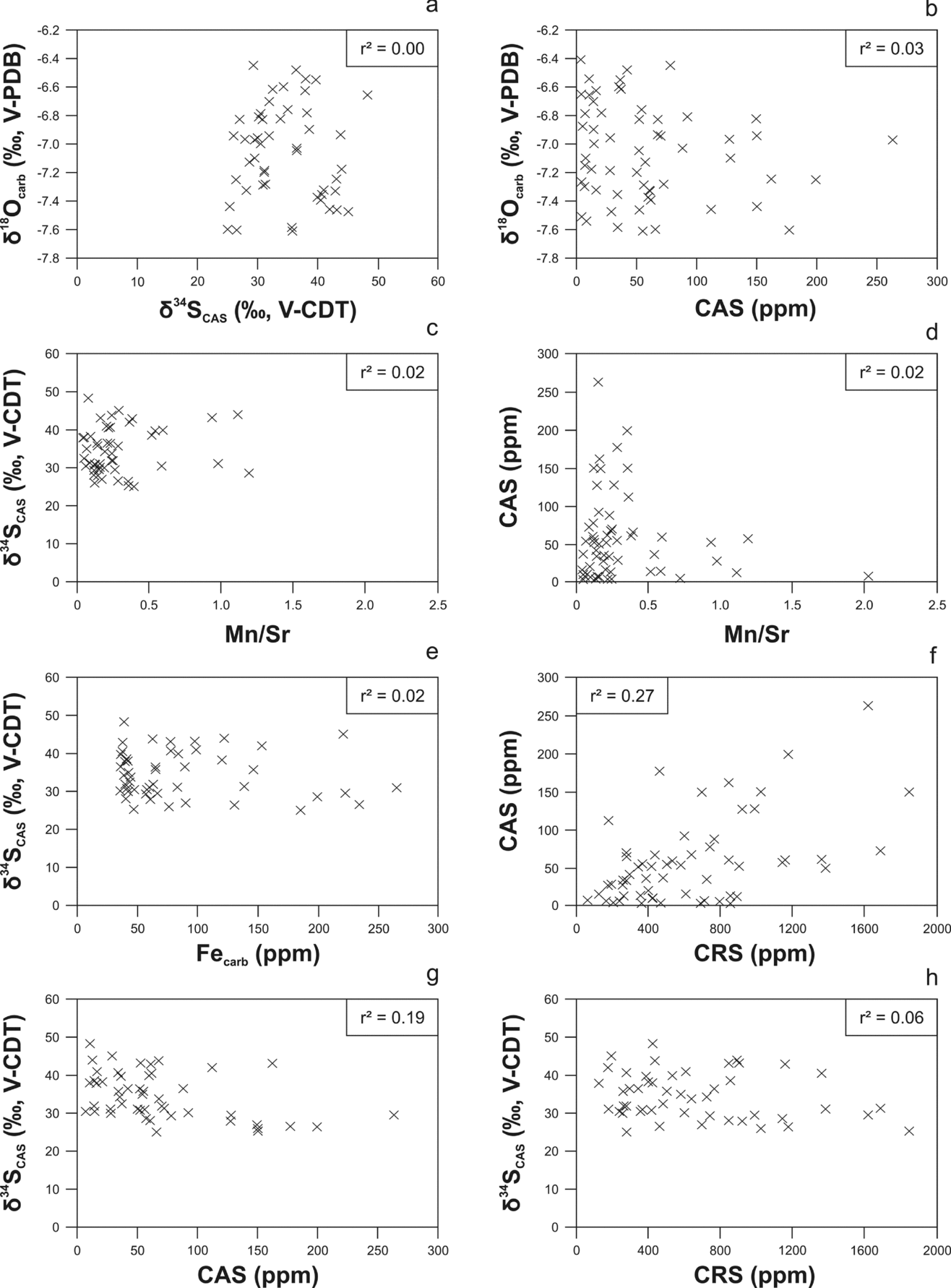
Prior to interpreting the carbon and sulphur isotopes, an evaluation of the degree of post-depositional alteration is essential. Respective proxies include δ18Ocarb values and elemental abundances and ratios (especially of Ca, Fe, Mg, Mn and Sr) of the carbonate matrix (Brand & Veizer, Reference Brand and Veizer1980; Veizer et al. Reference Veizer, Ala, Azmy, Bruckschen, Buhl, Bruhn, Carden, Diener, Ebneth, Godderis, Jasper, Korte, Pawellek, Podlaha and Strauss1999; Wotte, Strauss & Sundberg, Reference Wotte, Strauss and Sundberg2011; Wotte, Shields-Zhou & Strauss, Reference Wotte, Shields-Zhou and Strauss2012; Wotte et al. Reference Wotte, Shields-Zhou and Strauss2012). Samples characterized by δ18Ocarb > −10.0‰ and a Mn/Sr < 10 (or even < 2–3) are frequently considered as representing a diagenetically well-preserved isotopic signal (Kaufman, Jacobsen & Knoll, Reference Kaufman, Jacobsen and Knoll1993; Kaufman & Knoll, Reference Kaufman and Knoll1995). Our carbonate samples display δ18Ocarb values more positive than −10.0‰ and an Mn/Sr of <3 (Figs 3a–d, 4a–b; Table 1). Furthermore, Ca/Mg is regarded as a proxy signal for determining the effect of dolomitization, with Ca/Mg >50 considered as representing carbonate samples with the least-altered δ34SCAS values (Marenco et al. Reference Marenco, Corsetti, Kaufman and Bottjer2008). Our Ca/Mg is generally above 50, indicating no or only minor diagenetic alteration of sampled material (Fig. 3c, e, f; Table 1).

Figure 3. Cross-plots of stable isotope data, CAS and CRS concentrations, and element data from Kyrshabakty section. No covariance between proxies of diagenesis versus stable isotopes indicates the absence of any alteration of sample material and corresponding isotope data. (a) δ18Ocarb (‰, V-PDB) against δ13Ccarb (‰, V-PDB). (b) δ18Ocarb (‰, V-PDB) against Mn/Sr. (c) δ18Ocarb (‰, V-PDB) against Ca/Mg. (d) δ13Ccarb (‰, V-PDB) against Mn/Sr. (e) δ34SCAS (‰, V-CDT) against Ca/Mg. (f) δ13Ccarb (‰, V-PDB) against Ca/Mg. (g) δ34SCAS (‰, V-CDT) and δ34SCRS (‰, V-CDT) against δ34SNaCl (‰, V-CDT). δ34SNaCl values differ from the δ34SCAS and δ34SCRS data, indicating no influence of the non-CAS fraction on the final CAS and CRS isotopic values.

Figure 4. Cross-plots of stable isotope data, CAS and CRS concentrations, and element data from Kyrshabakty sections. The absence of any correlation between δ34SCAS, CAS and proxies for diagenesis suggests that the sulphur isotopic composition of carbonate-associated sulphate was not diagenetically altered. (a) δ18Ocarb (‰, V-PDB) against δ34SCAS (‰, V-CDT). (b) δ18Ocarb (‰, V-PDB) against CAS (ppm). (c) δ34SCAS (‰, V-CDT) against Mn/Sr. (d) CAS (ppm) against Mn/Sr. (e) δ34SCAS (‰, V-CDT) against Fecarb (ppm). (f) CAS (ppm) against CRS (ppm). (g) δ34SCAS (‰, V-CDT) against CAS (ppm). (h) δ34SCAS (‰, V-CDT) against CRS (ppm).
In order to evaluate a post-depositional resetting of the primary marine isotope signal, cross-plots of seawater proxies versus proxies of diagenetic alteration are applied (Banner & Hanson, Reference Banner and Hanson1990; Jacobsen & Kaufman, Reference Jacobsen and Kaufman1999; Wotte et al. Reference Wotte, Álvaro, Shields, Brown, Brasier and Veizer2007; Derry, Reference Derry2010; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). Plots using a combination of δ13Ccarb, δ18Ocarb, Mn/Sr and Ca/Mg show no covariance, indicating the absence of any substantial post-depositional carbonate alteration and the corresponding isotope data (Figs. 3, 4). In addition, no correlations exist between the proxies of diagenesis versus CAS and CRS concentrations and/or their sulphur isotope compositions (Figs 3, 4). We therefore assume that a severe and/or heterogeneous alteration of our carbonate samples is absent.
Caution needs to be applied when considering the quality of isotopic values and concentrations of CAS and CRS. Prior to the final liberation of CAS from the carbonate lattice it is essential to eliminate all leachable non-CAS sulphur-bearing phases (Kampschulte, Bruckschen & Strauss, Reference Kampschulte, Bruckschen and Strauss2001; Kampschulte & Strauss, Reference Kampschulte and Strauss2004; Wotte, Strauss & Sundberg, Reference Wotte, Strauss and Sundberg2011; Wotte, Shields-Zhou & Strauss, Reference Wotte, Shields-Zhou and Strauss2012; Wotte et al. Reference Wotte, Shields-Zhou and Strauss2012). A consecutive leaching procedure with 10% NaCl (aq) is therefore considered as delivering the most pristine CAS and CRS concentrations and isotopic compositions (for more information see Wotte et al. Reference Wotte, Shields-Zhou and Strauss2012). Although no leachable non-CAS was detected for 47 out of the 58 samples after the first NaCl-leaching, double leaching was necessary for 11 samples in order to eliminate all non-CAS compounds (Table 2) from the sample powder. δ34SNaCl values vary between 11.1‰ (K50/1) and 34.9‰ (K26) with concentrations between 2 ppm (K9) and 67 ppm (K6). These values clearly differ from the δ34SCAS and δ34SCRS data, indicating no influence of the non-CAS fraction on the final CAS isotope values (Fig. 3g; Table 2).
Table 2. Comparison of δ34SNaCl, δ34SCAS and δ34SCRS data, and SO4 concentrations generated from NaCl leachings.

Another aspect concerning CAS is that its abundance decreases during progressive diagenetic alteration, but with no influence on the δ34SCAS data (Hurtgen et al. Reference Hurtgen, Halverson, Arthur and Hoffman2006; Gill, Lyons & Frank, Reference Gill, Lyons and Frank2008). It should be considered, however, that the primary δ34SCAS signal can only be affected by sulphate derived from either sulphide oxidation or bacterial sulphate reduction if that sulphate is incorporated into the carbonate (Kampschulte & Strauss, Reference Kampschulte and Strauss2004; Wotte et al. Reference Wotte, Shields-Zhou and Strauss2012). The clearly less positive δ34SCAS value of sample K26 (28.6‰) correlates with enhanced CRS (1144 ppm) and Fe (200 ppm) concentrations, probably indicating an incorporation of sulphate derived from pyrite oxidation (Table 1). Considering the successive leaching with NaCl, oxidation must have happened during the diagenesis rather than represent an analytical artefact (Marenco et al. Reference Marenco, Corsetti, Kaufman and Bottjer2008). However, no covariance between less-positive δ34SCAS data and high CRS and/or Fe concentrations exist (Fig. 4e, h). For samples K30 and K33, separating the maximum δ34SCAS value (48.3‰ at K32) and the two distinctive positive excursions during the Guzhangian–Paibian interval, there is no correlation between the less positive δ34SCAS values (31.9‰ at K30 and 30.5 at K33) and enhanced CRS and/or Fe concentrations (Table 1). Regarding the arguments above, we consider our δ13Ccarb, δ18Ocarb, δ34SCAS and δ34SCRS data as reflecting the isotopic composition of ambient seawater. For further discussions on diagenetic alteration of δ34SCAS values see Wotte, Shields-Zhou & Strauss (Reference Wotte, Shields-Zhou and Strauss2012) and Wotte et al. (Reference Wotte, Shields-Zhou and Strauss2012).
6. Discussion
6.a. δ13Ccarb values: stratigraphic variations and biostratigraphic constraints
Our δ13Ccarb values of the Kyrshabakty section reproduce and extend the dataset of Saltzman et al. (Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000). The δ13Ccarb peak value of +5.0‰ (+4.82‰ in Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000) is situated within the Acutatagnostus acutatus-Erixanium biozone and represents the maximum amplitude of the SPICE event (Fig. 2). The onset of the SPICE is generally considered as coinciding with the FAD of Glyptagnostus reticulatus (base of the Paibian; Peng et al. Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004). At the Kyrshabakty section, the evolution towards more positive δ13Ccarb values starts during the uppermost Guzhangian Stage (Kormagnostus simplex and Glyptagnostus stolidotus biozones; Fig. 2). A similar positive trend prior to the FAD of G. reticulatus is reported for the majority of sections (Australia, China, France, Nevada, Newfoundland, Sweden and UK) at which the SPICE has been identified (Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Álvaro et al. Reference Álvaro, Bauluz, Subías, Pierre and Vincaїno2008; Ahlberg et al. Reference Ahlberg, Axheimer, Babcock, Eriksson, Schmitz and Terfelt2009; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Chen et al. Reference Chen, Chough, Han and Lee2011, Reference Chen, Chough, Lee and Han2012; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011; Woods et al. Reference Woods, Wilby, Leng, Rushton and Williams2011). Even at the Paibian stratotype section, the base of the SPICE does not correspond to the base of the G. reticulatus zone but to the upper G. stolidotus biozone situated below (see Peng et al. Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004). At the Siberian Kulyumbe section, the rise in δ13C seems to start directly at the G. reticulatus zone and culminates in a maximum value of c. +5‰ (Kouchinsky et al. Reference Kouchinsky, Bengtson, Gallet, Korovnikov, Pavlov, Runnegar, Shields, Veizer, Young and Ziegler2008; Woods et al. Reference Woods, Wilby, Leng, Rushton and Williams2011). Within the Wanliangyu section (Shandong Province, North China) the proposed SPICE event reaches a peak value of +3.5‰ and covers the uppermost Neodrepanura, the Chuangia and the lowermost Changshania-Irvingella trilobite zones (Chen et al. Reference Chen, Chough, Han and Lee2011). Consequently, the rise of the δ13C values starts clearly prior to the FAD of G. reticulatus of time-equivalent sections (Geyer & Shergold, Reference Geyer and Shergold2000). The same section is repeatedly published by Chen et al. (Reference Chen, Chough, Lee and Han2012) who shifted the SPICE excursion into a higher stratigraphic position, now starting at the basal Chuangia biozone. However, the general problem of sections of the Shandong Province is the missing Prochuangia biozone that normally occurs between the Neodrepanura and Chuangia biozones (Chen et al. Reference Chen, Chough, Han and Lee2011, Reference Chen, Chough, Lee and Han2012). This absence is probably caused by a non-deposition during subaerial exposure (Chen et al. Reference Chen, Chough, Han and Lee2011). In consequence, a fundamental portion of the δ13C curve is probably absent, hiding information about its original duration, magnitude and shape. The starting point of the SPICE event at the basal Chuangia biozone (as illustrated by Chen et al. Reference Chen, Chough, Lee and Han2012, fig. 10) therefore has to be wrong. Within the Tangwangzhai section, North China, the peak value of the proposed SPICE event reaches +2.2‰ and covers the uppermost Neodrepanura to basal Changshania-Irvingella trilobite zones (Zhu et al. Reference Zhu, Zhang, Li and Yang2004, fig. 4). The Prochuangia biozone is again absent, likely explaining the small magnitude of the SPICE at this location. Similar to the Wanliangyu section of Chen et al. (Reference Chen, Chough, Han and Lee2011), the SPICE at Tangwangzhai starts below the G. reticulatus biozone. The SPICE excursion of the Shuangqiao section (Hubei Province, North China) starts at the uppermost Neodrepanura zone and covers the Prochuangia – lower Changshania-Irvingella zones (Ng, Yuan & Lin, Reference Ng, Yuan and Lin2014). A maximum of +3.2‰ is reached in the lower Chuangia biozone which is followed by a short prominent negative excursion (−1.5‰) and a subsequent rise to almost maximum values (Ng, Yuan & Lin, Reference Ng, Yuan and Lin2014). Again, the δ13C excursion starts prior to the FAD of G. reticulatus.
At the Australian Mount Whelan 1 drillcore δ13Ccarb data start to rise clearly prior to the FAD of G. reticulatus, reaching a peak value of >6‰ in the upper Paibian (upper Idamean according to the Australian nomenclature; Geyer & Shergold, Reference Geyer and Shergold2000; Kruse, Jago & Laurie, Reference Kruse, Jago and Laurie2009; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). Biostratigraphy shows that isotope data cover a late Guzhangian – early Jiangshanian (late Mindyallan – early Iverian) interval (Green & Balfe, Reference Green and Balfe1980; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000) and not only the Guzhangian–Paibian interval (Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). δ13Ccarb data of the Australian Mount Murray section rise from +2.4‰ to a peak value of +4.2‰ some metres above the FAD of Irvingella tropica (Shergold, Reference Shergold1982, Reference Shergold1993) and show a subsequent decrease towards +1.0‰ (Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011; Fig. 5). Data cover a Paibian–Jiangshanian interval (Idamean–Iverian; Geyer & Shergold, Reference Geyer and Shergold2000; Kruse, Jago & Laurie, Reference Kruse, Jago and Laurie2009). According to Shergold (Reference Shergold1982, Reference Shergold1993), I. tropica occurs at 87 m at the Mount Murray section and defines the Iverian base (Geyer & Shergold, Reference Geyer and Shergold2000). This datum was misused by Gill et al. (Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011) who plotted I. tropica at c. 120 m, consequently shifting the Idamean–Iverian boundary into a stratigraphic younger position. However, a correlation with δ13Ccarb data of the nearby Mount Whelan 1 drillcore suggests a position of Mount Murray data at the maximum point to decreasing limb of the SPICE (Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). Considering the low sampling density of the Mount Murray section, it is hard to compare the δ13Ccarb curves of both sections precisely (Fig. 5). At sections of Missouri (TE-1Texas County Core) and Newfoundland (Felix Cove section), the rise in δ13C pre-dates the FAD of Aphelaspis which is equivalent to the G. reticulatus zone (Geyer & Shergold, Reference Geyer and Shergold2000; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). At the Laurentian House Range and Lawson Cove sections (both Utah; Saltzman, Runnegar & Lohmann, Reference Saltzman, Runnegar and Lohmann1998) documentation begins at the basal Aphelaspis zone (equivalent to the G. reticulatus zone; Geyer & Shergold, Reference Geyer and Shergold2000), preventing characterization of the SPICE onset.
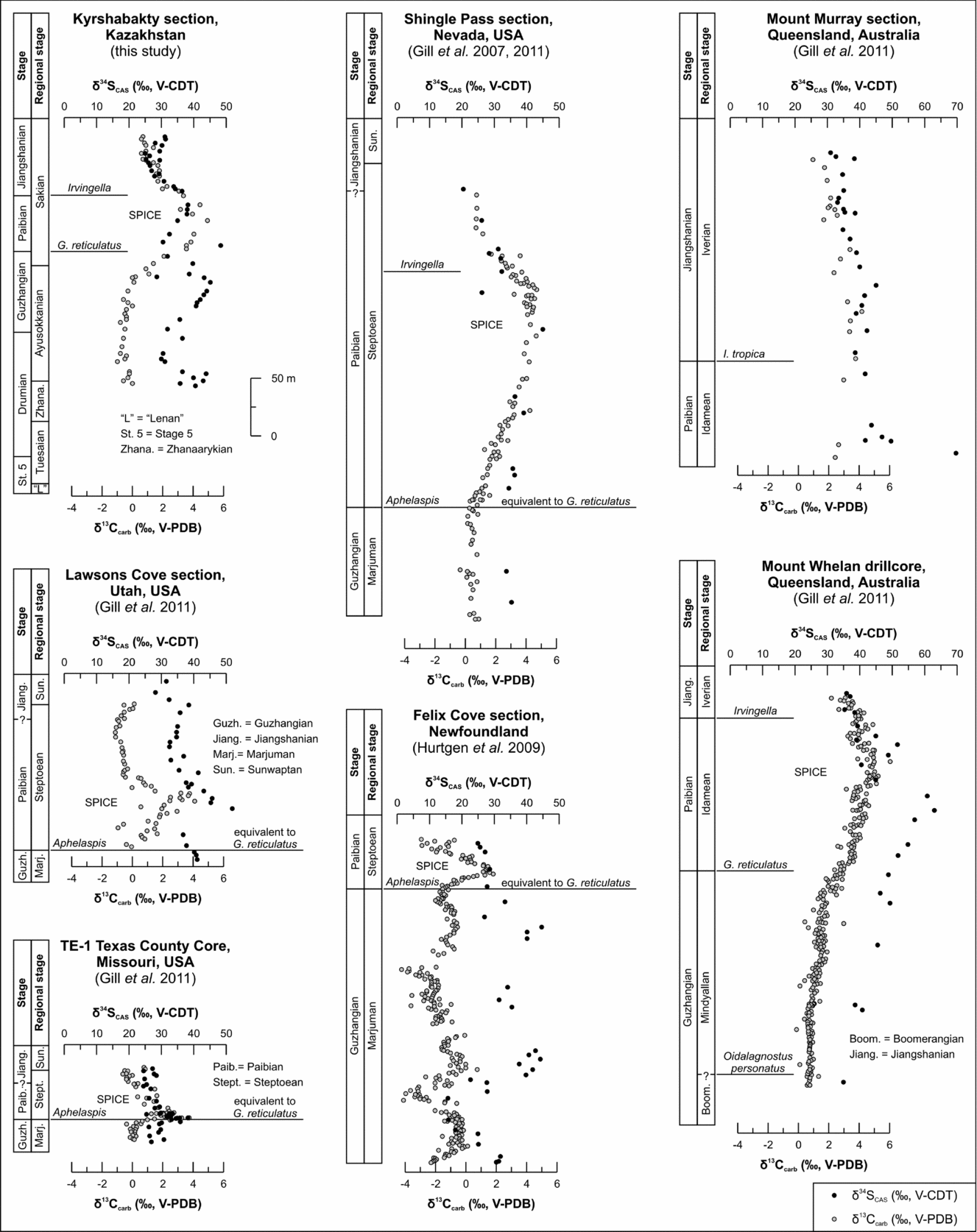
Figure 5. Global comparison of parallel δ13Ccarb and δ34SCAS data during the Cambrian Series 3 to Furongian interval. Data are derived this study, Gill, Lyons & Saltzman (Reference Gill, Lyons and Saltzman2007), Gill et al. (Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011) and Hurtgen et al. (Reference Hurtgen, Pruss and Knoll2009), and are calibrated for uniform vertical and lateral scale. Variations in the onset of local SPICE and SPICE-related δ34SCAS curves, their peak values, amplitudes and absolute values indicate a heterogeneous Furongian seawater reservoir with respect to sulphate and carbon. Supportive biostratigraphic information is derived from Saltzman et al. (Reference Saltzman, Runnegar and Lohmann1998) for Lawsons Cove and Shingle Pass, from Saltzman et al. (Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) and Boyce & Knight (Reference Boyce and Knight2005) for Felix Cove, from Shergold (Reference Shergold1982) for Mount Murray, and from Green and Balfe (Reference Green and Balfe1980) and Geyer & Shergold (Reference Geyer and Shergold2000) for Mount Whelan 1.
It becomes obvious that, despite the general similarities documented for the SPICE in all studied sections, the individual peak values and amplitudes differ (Ahlberg et al. Reference Ahlberg, Axheimer, Babcock, Eriksson, Schmitz and Terfelt2009; Woods et al. Reference Woods, Wilby, Leng, Rushton and Williams2011). The most-negative δ13Ccarb values are recorded from Newfoundland (+1.8‰; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Fig. 5) whereas the most-positive data are reported from Kazakhstan (+5‰, this study; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Figs 2, 5), South China (+5‰; Wa’ergang section; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000) and Australia (>6‰; Mount Whelan 1 core; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011; Fig. 5). For sections of North China, variations in the magnitude of δ13Ccarb values are probably related to the absence of the Prochuangia biozone. Discrepancies in onset, peak values and shape of the SPICE are generally attributed to biostratigraphic uncertainties, different thicknesses of individual successions and variations in sample density (Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Ahlberg et al. Reference Ahlberg, Axheimer, Babcock, Eriksson, Schmitz and Terfelt2009; Woods et al. Reference Woods, Wilby, Leng, Rushton and Williams2011). While differences in the magnitude of isotope excursions are probably related to the chemical gradient of seawater, there is no reason for biostratigraphic discrepancies misdating the onset of the SPICE if a well-established palaeontological database supports the geochemical data. According to the rules for biostratigraphic correlation of the International Commission on Stratigraphy (ICS), the base of a stage or series has to be defined by the FAD of cosmopolitan faunal element. This datum has to be considered as a globally recognizable time-equivalent marker. The FAD of G. reticulatus represents such a synchronous stratigraphic datum which defines the base of the Paibian/Furongian (Peng et al. Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004; Peng, Babcock & Cooper, Reference Peng, Babcock, Cooper, Gradstein, Ogg, Schmitz and Ogg2012). In consequence, variations in the onset of the SPICE should therefore not be an artefact of biostratigraphic discrepancies. However, biostratigraphic information on the onset but also on the duration of the SPICE of some published sections needs re-investigation (e.g. stratigraphic occurrences of G. reticulatus and Irvingella; see Chen et al. Reference Chen, Chough, Han and Lee2011, Reference Chen, Chough, Lee and Han2012; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). Frequently, chemostratigraphic studies across the Furongian section lack a detailed (sample to sample basis) biostratigraphic control and in-depth evaluation of the biostratigraphic constraints of the stratigraphic sections under study. This becomes relevant if sample material is limited, as in the case of drillcores. Keeping all these factors in mind, it still seems most probable that differences in δ13C datasets indicate a Furongian seawater reservoir affected by regional variations. Consequently, the ocean was probably not as homogeneous with respect to δ13C as generally assumed (see also Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009).
6.b. δ34SCAS and δ34SCRS values: stratigraphic variations and biostratigraphic constraints
A parallel evolution of carbon and sulphur isotope excursions has been observed from various sedimentary successions of Australia, Sweden and western US (Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). Our positive excursions in δ34SCAS and δ34SCRS occur almost at the same level as the δ13C excursion, that is, the level of the SPICE (Fig. 2). The δ34SCRS peak value (31.9‰) at Kyrshabakty almost coincides with the carbon isotope maximum. The co-occurrence of these maxima indicates that the amount of organic matter stimulates bacterial sulphate reduction and the formation of pyrite, resulting in the observed 34S enrichment (Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011).
Similar to the δ13Ccarb values, the onset of excursions, peak values, amplitudes and absolute values of δ34SCAS and δ34SCRS vary among the localities (Fig. 5). The most-positive δ34SCAS values are reported from Mount Murray and Mount Whelan 1 (both Queensland, Australia) with 69.4‰ and 62.9‰, respectively (Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). The least-positive value (38.1‰) is documented from Missouri. At Kyrshabakty the δ34SCAS peak value of 48.3‰ occurs in a stratigraphically lower position than the δ13Ccarb maximum (Figs 2, 5). Our δ34SCAS data increase during the Lejopyge armata biozone, culminating in three positive excursions prior to and after the SPICE maximum (Figs 2, 5). This feature is still preserved when considering the least-positive value (28.6‰ at K26) as diagenetically altered (Fig. 2). It therefore indicates a stepwise evolution of the SPICE-related positive δ34SCAS excursion. The large shifts in δ34SCAS are probably indicative of ocean water which was sensitive to, for example, changes in pyrite burial and/or Δ34S (Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009). Similar pre-SPICE variations could be identified from drillcores of Missouri (TE-1 Texas County Core) and Mount Whelan 1 (Queensland, Australia), and probably from the Felix Cove section of Newfoundland (Fig. 5; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). Whether the lack of similar trends in δ34SCAS from sections of Nevada and Utah (Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011) results from limited sampling frequency and/or diagenetic alteration, or reflects the primary isotopic signature of Furongian seawater sulphate sulphur, requires further investigation.
A positive δ34SCAS excursion prior to the SPICE becomes evident (Fig. 5) when calibrating the various datasets on uniform vertical and horizontal scales. The δ34SCAS peak pre-dates the SPICE maximum in almost all sections (except Missouri) at different stratigraphic positions (Fig. 5). According to Gill et al. (Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011), a pre-occurrence of the δ34SCAS excursion probably indicates a sulphate reservoir which was more sensitive to changes than the marine DIC pool. Differences in stratigraphic position of the onset and the maximum δ34SCAS values of the SPICE-related δ34SCAS excursion and variations in their excursion magnitude are indicative of a heterogeneous Furongian seawater sulphate reservoir with low sulphate concentrations (Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). This heterogeneity is not limited to the Upper Cambrian. Comparable differences were also reported by Wotte, Strauss & Sundberg (Reference Wotte, Strauss and Sundberg2011) and Wotte et al. (Reference Wotte, Shields-Zhou and Strauss2012) for the traditional Lower–Middle Cambrian boundary interval of Gondwana, Siberia and Laurentia. However, it should be kept in mind that the various datasets of δ34SCAS and δ34SCRS were generated using different methods of extraction (Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Wotte, Shields-Zhou & Strauss Reference Wotte, Shields-Zhou and Strauss2012). The consequent and complete elimination of non-CAS sulphur is essential in order to exclude the generation of a mixed δ34SCAS signal which does not represent the primary composition of seawater. The elimination of non-CAS sulphur often requires consecutive leaching steps (Wotte, Shields-Zhou & Strauss, Reference Wotte, Shields-Zhou and Strauss2012; Wotte et al. Reference Wotte, Shields-Zhou and Strauss2012). A detailed assessment of prevailing analytical methods revealed that both a consecutive elimination of non-CAS sulphur and a careful evaluation of the degree of diagenetic alteration of the CAS and CRS signals (concentration and isotopic composition) were only rarely realized (Wotte, Shields-Zhou & Strauss, Reference Wotte, Shields-Zhou and Strauss2012). Caution therefore needs to be applied when comparing/interpreting different datasets of δ34SCAS and δ34SCRS.
6.c. Evidence for non-euxinic oceanic conditions during the SPICE
The parallel evolution of positive carbon and sulphur isotope excursions during the Furongian has been interpreted as representing widespread ocean anoxia/euxinia and increased burial of pyrite and organic matter (Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). Additional evidence for anoxic/euxinic conditions derives from decreasing Mo concentrations during the SPICE event (Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011). An increase of anoxia during the SPICE is also indicated by Δ34S values showing an opposite trend parallel to the positive δ13Ccarb, δ34SCAS and δ34SCRS excursions (Fig. 2). The co-occurrence of low Δ34S and very positive δ34SCAS values indicate extensive pyrite burial under euxinic conditions from low and continuously decreasing sulphate concentrations (Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011).
However, is the interpretation of widespread subsurface anoxic or even euxinic oceanic conditions really valid for the time interval associated with the SPICE event? The SPICE interval of almost all sections is characterized by the occurrence of benthic faunal elements, clearly indicating the availability of oxygen in the bottom water of worldwide low- to high-latitude environments. Algal mats, brachiopods, sponge spicules, probable conulariids and camaroids, echinoderms and other pelmetazoan ossicles are reported from, for example, the Kyrshabakty section (Kazakhstan; Holmer et al. Reference Holmer, Popov, Koneva and Bassett2001; Koneva & Ushatinskaya, Reference Koneva and Ushatinskaya2010), the Mount Murray section (Queensland, Australia; Shergold, Reference Shergold1982), the Montagne Noire (France; Álvaro et al. Reference Álvaro, Bauluz, Subías, Pierre and Vincaїno2008), Västergötland (Sweden; Ahlberg et al. Reference Ahlberg, Szaniawski, Clarkson and Bengtson2005), the Shingle Pass section (Nevada; Palmer, Reference Palmer1965; Saltzman, Runnegar & Lohmann, Reference Saltzman, Runnegar and Lohmann1998), the Lawsons Cove section (Utah; R.L. Freeman and J.F. Miller, pers. comm.) and other localities of Arizona, Minnesota, Missouri, Montana, Nevada, Tennessee, Texas, Utah, Wisconsin and Wyoming (e.g. Palmer, Reference Palmer1954; Grant, Reference Grant1965; Kurtz, Reference Kurtz1971, Reference Kurtz1975; Freeman & Sitt, Reference Freeman and Sitt1996; Cowan et al. Reference Cowan, Fox, Runkel and Saltzman2005; Rieboldt, Reference Rieboldt2011; Miller, Evans & Dattilo, Reference Miller, Evans, Dattilo, Derby, Fritz, Longacre, Morgan and Sternbach2012). In addition, the SPICE interval of sections of the Port au Port Peninsula (western Newfoundland) is characterized by the occurrence of stromatolites, brachiopods and ostracods (Boyce & Knight, Reference Boyce and Knight2005), suggesting a similar fauna for the Felix Cove section (published by Saltzman et al. Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009). Proposed widespread euxinic oceanic conditions or even an anoxic event are inconsistent with these data. However, it seems to be most obvious that the deep ocean and sections free of any benthic faunal content reflect regional anoxia (Ahlberg et al. Reference Ahlberg, Axheimer, Babcock, Eriksson, Schmitz and Terfelt2009).
6.d. Processes forcing the SPICE: cooling phase and/or pH decline
According to Saltzman et al. (Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000, Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004), the peak of the SPICE corresponds to a maximum regression on Laurentia marked by the continent-wide Sauk II – Sauk III subsequence boundary. Evidence for a drop in sea-level is given for example by a karst surface within the siliciclastic-dominated Davis Formation of Utah, probably coinciding with the Sauk II – Sauk III boundary (Miller et al. Reference Miller, Evans, Kurtz, Repetski, Mulvany and Thompson2013). Saltzman et al. (Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000, Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) discussed a potential linking between globally enhanced weathering of marine carbonate-platform-deposits and a glacial event promoting the SPICE. However, there is no evidence for ice sheets during that interval (Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000). High 87Sr/86Sr values in seawater, probably also induced by widespread sea-level fall, also indicate increased erosion and continental weathering for the SPICE interval (Montañez et al. Reference Montañez, Banner, Osleger, Borg and Bosserman1996, Reference Montañez, Osleger, Banner, Mack and Musgrove2000; Denison et al. Reference Denison, Koepnick, Burke and Hetherington1998; Kouchinsky et al. Reference Kouchinsky, Bengtson, Gallet, Korovnikov, Pavlov, Runnegar, Shields, Veizer, Young and Ziegler2008). Additional evidence for sea-level fall is given by the exposure of carbonate platforms and the deposition of siliciclastic sequences (e.g. Australia; Lindsay et al. Reference Lindsay, Kruse, Green, Hawkins, Brasier, Cartlidge and Corfield2005) or the collapse of the Aisha-Bibi seamount in the Malyi Karatau Range of Kazakhstan (Cook et al. Reference Cook, Taylor, Zhemchuzhnikov, Apollonov, Ergaliev, Sargaskaev, Dubinina, Cooper and Stevens1991). A drop in sea-level would also contradict the development of widespread euxinic conditions in a variety of marine depositional environments ranging from deeper siliciclastic settings to shelfal carbonate platforms. However, an enhanced continental weathering would also deliver additional sulphate into the seawater, which in turn would be inconsistent with a low seawater sulphate level during the SPICE. This aspect needs further investigation. Indirect indication for a eustatic sea-level fluctuation is given by Elrick et al. (Reference Elrick, Rieboldt, Saltzman and McKay2011), who analysed δ18Ophos of phosphatic brachiopods from central and western US. Decreasing δ18Ophos values (from c. 15‰ to c. 13‰ V-SMOW) parallel to the rising limb of SPICE would indicate a general warming of seawater during the Aphelaspis–Dunderbergia interval (lower Steptoean; Geyer & Shergold, Reference Geyer and Shergold2000). The proposed warming trend is followed by a successive cooling during the upper Dunderbergia and Elvinia biozones (upper Steptoean; Geyer & Shergold, Reference Geyer and Shergold2000). However, such a warming trend is not conformable to a global sea-level fall. Interestingly, our δ18Ocarb values show an opposite trend. δ18Ocarb data evolve in parallel to δ13Ccarb, rising from −7.5‰ V-PDB (K24) to a maximum of −6.4‰ V-PDB (K42), followed by a continuous decrease (Figs 2, 3a). There is no indication that diagenetic processes affected our δ18Ocarb data (Figs 3, 4). Furthermore, data fit very well with the known δ18O record of the entire Cambrian to basal Ordovician section (Jaffrés, Shields & Wallmann, Reference Jaffrés, Shields and Wallmann2007; Prokoph, Shields & Veizer, Reference Prokoph, Shields and Veizer2008; Trotter et al. Reference Trotter, Williams, Barnes, Lécuyer and Nicoll2008). For this reason we consider our δ18Ocarb data as representing the primary oxygen isotope signal. Increasing δ18Ocarb values during the lower part of the SPICE at the Kyrshabakty section indicate a cooling of seawater, whereas the falling limb would represent a continuous warming trend. This trend in δ18Ocarb values is less well developed in Saltzman et al. (Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000). A similar parallel evolution between δ13Ccarb and δ18Ocarb is reported from the Siberian Kulyumbe section with peak δ18Ocarb values of c. −6.0‰ and an amplitude of 3–4‰ (Kouchinsky et al. Reference Kouchinsky, Bengtson, Gallet, Korovnikov, Pavlov, Runnegar, Shields, Veizer, Young and Ziegler2008). In contrast, no sympathetic shifts are documented for paired δ13Ccarb and δ18Ocarb values available from, for example, the Wa’ergang section (North China; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000), the Tangwangzhai section (South China; Zhu et al. Reference Zhu, Zhang, Li and Yang2004) or sections in Nevada, Missouri or Tennessee (Glumac & Walker, Reference Glumac and Walker1998; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011).
Alternatively, the co-varying trend between δ13Ccarb and δ18Ocarb data could also be interpreted with respect to changes in [CO3 2−] and pH of ocean water. McCrea (Reference McCrea1950) and Usdowski & Hoefs (Reference Usdowski and Hoefs1993) demonstrated that the δ18O values of rapidly precipitated CaCO3 essentially depend on the pH of the solution. Experiments with living foraminifera have shown that δ13C and δ18O of calcite shells become isotopically heavier as seawater pH or [CO3 2−] decrease (Spero et al. Reference Spero, Bijma, Lea and Bemis1997). This becomes important when applying oxygen isotopes for reconstructing palaeoclimatic or palaeoenvironmental changes (Spero et al. Reference Spero, Bijma, Lea and Bemis1997; Zeebe, Reference Zeebe2001).
Zeebe (Reference Zeebe1999) demonstrated that an increase of seawater pH by 0.2–0.3 units causes a decrease in δ18O of 0.22–0.33‰, which would usually be interpreted as an increase in seawater temperature. More recently, Uchikawa & Zeebe (Reference Uchikawa and Zeebe2010) calculated the possible effect of the pH on δ13C and δ18O during the Paleocene–Eocene thermal maximum (PETM) based on culture experiments with planktonic foraminifera. The authors determined via modelling that a change in pH by −0.1 to −0.3 for the average surface ocean equates to changes in δ18O and δ13C of up to 0.7‰ and c. 2.1‰, respectively. Changes in carbon input and release time and/or regional ocean geometry would affect these values accordingly (Uchikawa & Zeebe, Reference Uchikawa and Zeebe2010). In consequence, a pH decline could probably explain the +1‰ shift in our δ18Ocarb values during the rising limb of the SPICE. Whether the same correlation between δ18Ocarb values and pH could be transferred to inorganically precipitated carbonates needs further investigation (Kim, Hillaire-Marcel & Micci, Reference Kim, Hillaire-Marcel and Micci2006). Whether a drop in seawater pH could be maintained for a duration of 2–4 Ma (estimated duration of the SPICE; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000, Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004) also has to be investigated. If our interpretation is correct, no euxinic conditions would be required to explain the SPICE event. Such a scenario would then be associated with a sea-level rise, as proposed by Peng et al. (Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004) for the basal Furongian. A transgression probably also resulted in enhanced accumulation of organic matter in shallow-marine shelf environments (Wenger & Baker, Reference Wenger and Baker1986) and was therefore associated with fundamental changes in the ocean sulphate and carbonate chemistry, but also in alkalinity.
7. Conclusions
The Furongian base is often associated with the lower part of a eustatic sea-level rise, but also with the onset of a prominent positive excursion in δ13C (SPICE; Peng et al. Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004; Peng, Babcock & Cooper, Reference Peng, Babcock, Cooper, Gradstein, Ogg, Schmitz and Ogg2012). In multiple sections, the SPICE co-occurs with positive excursions in δ34SCAS and δ34SCRS, interpreted as reflecting an elevated rate of organic carbon and pyrite burial during widespread anoxic or euxinic conditions (Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011).
The Furongian Kyrshabakty section, southern Kazakhstan, preserves positive excursions of oxygen isotopes and sulphur isotopes of CAS and CRS in accordance with the positive δ13Ccarb peak, identified as the SPICE. The Kazakh SPICE excursion spans an interval ranging from the Glyptagnostus stolidotus to the upper Pseudagnostus vastulus-Irvingella tropica biozones. Its onset therefore clearly pre-dates the base of the Paibian Stage/Furongian Series identified by the FAD of Glyptagnostus reticulatus. A similar positive trend prior to the FAD of G. reticulatus is reported for the majority of sections at which the SPICE has been identified (Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000; Gill, Lyons & Saltzman, Reference Gill, Lyons and Saltzman2007; Álvaro et al. Reference Álvaro, Bauluz, Subías, Pierre and Vincaїno2008; Ahlberg et al. Reference Ahlberg, Axheimer, Babcock, Eriksson, Schmitz and Terfelt2009; Hurtgen, Pruss & Knoll, Reference Hurtgen, Pruss and Knoll2009; Chen et al. Reference Chen, Chough, Han and Lee2011, Reference Chen, Chough, Lee and Han2012; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011; Woods et al. Reference Woods, Wilby, Leng, Rushton and Williams2011). δ34SCAS data already increase during the Lejopyge armata biozone, culminating in three positive excursions prior to and after the SPICE maximum. This three-part evolution indicates a stepwise evolution of the SPICE-related positive δ34SCAS excursion, characterized by distinct fluctuations. Differences in onset, magnitude and shape of SPICE and SPICE-related δ34SCAS and δ34SCRS excursions indicate a Furongian seawater reservoir with low sulphate concentration and heterogeneous in its carbonate carbon and sulphate sulphur isotopic composition.
The SPICE interval of almost all sections offers a diverse benthic fauna, strongly suggesting oxygenated conditions at the seafloor. Proposed widespread euxinic oceanic conditions are inconsistent with these data, but regional anoxia and a euxinic deep ocean cannot be excluded.
Our δ18Ocarb values evolve in accordance to the SPICE. The basal rise in δ18Ocarb is interpreted as representing a cooling phase, whereas the upper falling limb indicates a continuous warming of seawater. Such climatic variation would be associated with widespread sea-level fall, increased erosion and continental weathering, the exposure of carbonate platforms and the deposition of siliciclastic sequences (Montañez et al. Reference Montañez, Banner, Osleger, Borg and Bosserman1996, Reference Montañez, Osleger, Banner, Mack and Musgrove2000; Denison et al. Reference Denison, Koepnick, Burke and Hetherington1998; Saltzman et al. Reference Saltzman, Ripperdan, Brasier, Lohmann, Robinson, Chang, Peng, Ergaliev and Runnegar2000, Reference Saltzman, Cowan, Runkel, Runnegar, Stewart and Palmer2004; Lindsay et al. Reference Lindsay, Kruse, Green, Hawkins, Brasier, Cartlidge and Corfield2005; Kouchinsky et al. Reference Kouchinsky, Bengtson, Gallet, Korovnikov, Pavlov, Runnegar, Shields, Veizer, Young and Ziegler2008). An alternative interpretation of the parallel δ13Ccarb and δ18Ocarb trends suggests a close link between seawater pH and [CO3 2−] concentration (McCrea, Reference McCrea1950; Usdowski & Hoefs, Reference Usdowski and Hoefs1993; Spero et al. Reference Spero, Bijma, Lea and Bemis1997; Zeebe, Reference Zeebe1999). Modelled changes in pH ranging from −0.1 to −0.3 for the average surface ocean correspond to a +0.7‰ and c. +2.1‰ shift in δ18O and δ13C, respectively (Uchikawa & Zeebe, Reference Uchikawa and Zeebe2010). A decline in pH could therefore probably also explain the +1‰ shift in our δ18Ocarb values.
Data from our recent work challenge much of the interpretations that have been proposed for the SPICE interval. However, the present multitude of uncertainties remaining requires further (re-)investigations of Furongian successions with respect to their isotopes (δ13C, δ18O and δ34S), elemental concentration and, essentially, their faunal content, allowing a biostratigraphic calibration of sections and strengthening the chemostratigraphic constraints of the SPICE event.
Acknowledgements
We are grateful to Gappar Kh. Ergaliev (Kazakh National Academy of Sciences, Almaty, Kazakhstan), Michael G. Bassett and Leonid E. Popov (both National Museum of Wales, Cardiff, UK) for organizing the 14th International Conference of the Cambrian Subdivision Working Group in 2009, held in southern Kazakhstan, during which fieldwork and sampling was performed. Anja Cording (University of Cologne), Artur Fugmann and Andreas Lutter (both University of Münster) are thanked for their assistance with isotope analyses. We also thank Dieter Garbe-Schönberg, Ulrike Westernströer and Karin Kissling (all University of Kiel) for performing the ICP-OES analyses. Constructive comments by Benjamin C. Gill (Virginia Tech) greatly improved an earlier version of the manuscript. We thank Alan Jay Kaufman (University of Maryland) and an anonymous reviewer for their helpful comments. Philip Leat (University of Leicester) is thanked for his editorial guidance. Generous funding from the German Research Foundation (Grant WO 1215/4) is gratefully acknowledged.
Declaration of interest
None