


1. Introduction
During the Jurassic period, the North American plate drifted northward, such that by middle Callovian to middle Oxfordian time, central Wyoming was approximately 23–25°N (Kocurek & Dott, Reference Kocurek, Dott, Reynolds and Dolly1983). The Middle and Upper Jurassic strata of the western interior of the US record a series of marine cycles of transgression and regression (Brenner & Peterson, Reference Brenner, Peterson, Caputo, Peterson and Franczyk1994; Kvale et al. Reference Kvale, Johnson, Mickelson, Keller, Furer and Archer2001). The Sundance Sea was the southern portion of a boreal ‘pocket’ sea, with an outlet to the north. It extended from central South Dakota to western Utah, and from northern New Mexico to the Arctic (Fig. 1; Brenner & Davies, Reference Brenner and Davies1974). On the eastern and southern shorelines, the low relief land shed sediments into the basin. The western shoreline was along highlands that became more tectonically active throughout Oxfordian time. With increasing tectonic activity, coarser sediments were deposited and eventually the basin was filled (Brenner & Davies, Reference Brenner and Davies1974).
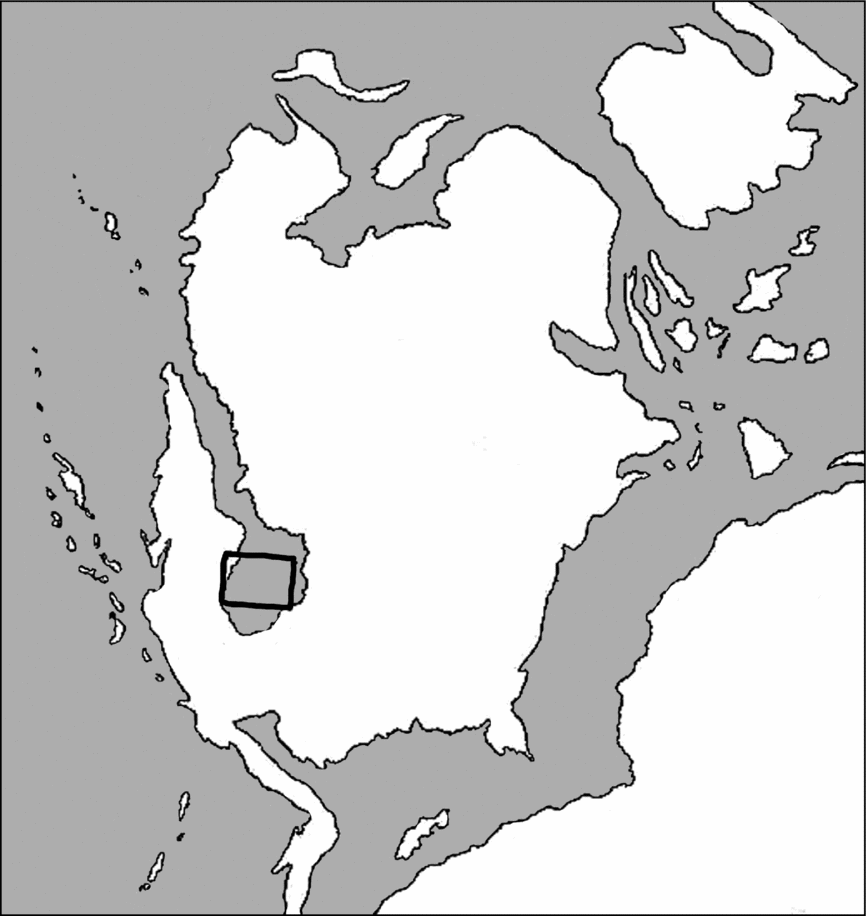
Figure 1. Palaeogeography of North America during the Upper Jurassic, showing the Sundance Sea. Continental areas in white; marine areas in grey. Rectangular outline shows the location of Wyoming. Modified from Rocky Mountain Association of Geologists (1972, p. 184) and online palaeogeographic map by Ronald Blakey (http://cpgeosystems.com/namJ150.jpg).
The upper Sundance Formation records the last incursion of the sea during Oxfordian time, the fourth Jurassic marine cycle in North America. It is bounded by the J4 and J5 unconformities (Pipiringos & O'Sullivan, Reference Pipiringos and O'Sullivan1978; Brenner & Peterson, Reference Brenner, Peterson, Caputo, Peterson and Franczyk1994). Ammonites, ostracodes, and foraminifera suggest that this interval lasted approximately 8 million years (Brenner & Peterson, Reference Peterson, Caputo, Peterson and Franczyk1994), but Kvale et al. (Reference Kvale, Johnson, Mickelson, Keller, Furer and Archer2001, figure 5) estimated about 4 million years. The sediments, which now make up the Redwater Shale Member of the Sundance Formation, record the largest Jurassic transgression and a regression, both of which occurred during a eustatic rise in sea level (Brenner & Peterson, Reference Brenner, Peterson, Caputo, Peterson and Franczyk1994). The transgression was controlled by the global sea level rise, but the regression occurred because increased tectonic activity to the west caused a high sedimentation rate that exceeded the rise in sea level (Brenner & Peterson, Reference Brenner, Peterson, Caputo, Peterson and Franczyk1994). More recently, Kvale et al. (Reference Kvale, Johnson, Mickelson, Keller, Furer and Archer2001) recognized three smaller-scale shallowing episodes within the upper Sundance Formation strata in north-central Wyoming (Bighorn Basin). These are punctuated by coquinas, which we have recognised in our field area as well. They have been attributed to storm deposits (Brenner & Davis, Reference Brenner and Davies1974) but have more recently been interpreted as sand waves generated by storm and tidal currents (Brenner, Swift & Gaynor, Reference Brenner, Swift and Gaynor1985).
This report is based in large part on our work in the upper Sundance Formation of central Wyoming. Our study area encompasses northern Natrona County and adjacent areas, including Alcova Reservoir (southern Natrona County) and the Gas Hills (eastern Fremont County; Fig. 2). Central Wyoming was far from the shoreline, in a deeper area of the Sundance Sea, and accumulated predominantly muddy facies of the Redwater Shale Member. The purpose of this paper is to describe the vertebrate fauna of the Redwater Shale Member based on more than a decade of field work and collecting in central Wyoming, and to present our interpretation of the palaeoecology of this interval of the Sundance Sea. We also compare the Redwater Shale Member vertebrate fauna to the more diverse and better known fauna of the Peterborough Member of the Oxford Clay Formation of England.
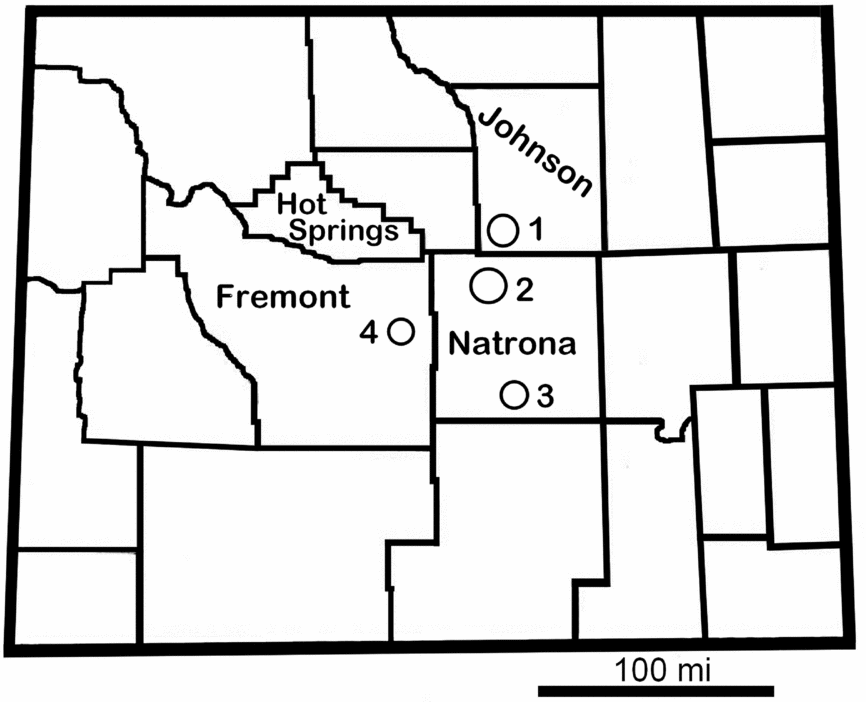
Figure 2. Outline map of Wyoming showing county boundaries. Our field areas included portions of Johnson, Fremont and Natrona counties. 1 – Wold Ranch; 2 – Thirty-three Mile Road area; 3 – Alcova Reservoir; 4 – Gas Hills. Scale bar represents 100 miles.
Institutional abbreviations. TATE – Tate Geological Museum, Casper College, Casper, WY; USNM – US National Museum, Smithsonian Institution, Washington, DC; UW – University of Wyoming, Laramie, WY; WDC – Wyoming Dinosaur Center, Thermopolis, WY.
2. Stratigraphy
The Sundance Formation ranges in age from middle Bathonian to Oxfordian (Kvale et al. Reference Kvale, Johnson, Mickelson, Keller, Furer and Archer2001). It lies unconformably on the Triassic Gypsum Springs Formation and is overlain by the Morrison Formation, river channel and floodplain deposits famous for a diverse dinosaur assemblage. The Windy Hill Sandstone (Pipiringos, Reference Pipiringos1968) occurs discontinuously between the terrestrial strata of the Morrison Formation and the marine Redwater Shale Member. It is a marine sandstone, oolitic in some locations, with cross-bedding, oscillation and current ripple marks, and pterosaur tracks (Johnson, Reference Johnson1992; Connely, Reference Connely2006). It indicates shallow marine deposition where the sedimentation rate could not keep up with the eustatic sea level rise (Brenner & Peterson, Reference Brenner, Peterson, Caputo, Peterson and Franczyk1994). Until recently, the Windy Hill sandstone was included in the upper Sundance Formation (Pipiringos, Reference Pipiringos1968; Johnson, Reference Johnson1992), but its occurrence above the J-5 unconformity and an interfingering relationship with the lower Morrison strata have led to its reassignment to the Morrison Formation (Brenner, Swift & Gaynor, Reference Brenner, Swift and Gaynor1985; Peterson, Reference Peterson, Caputo, Peterson and Franczyk1994). However, the lower Morrison strata are terrestrial, whereas Windy Hill strata are marine and thus at least environmentally more similar to the Sundance Formation.
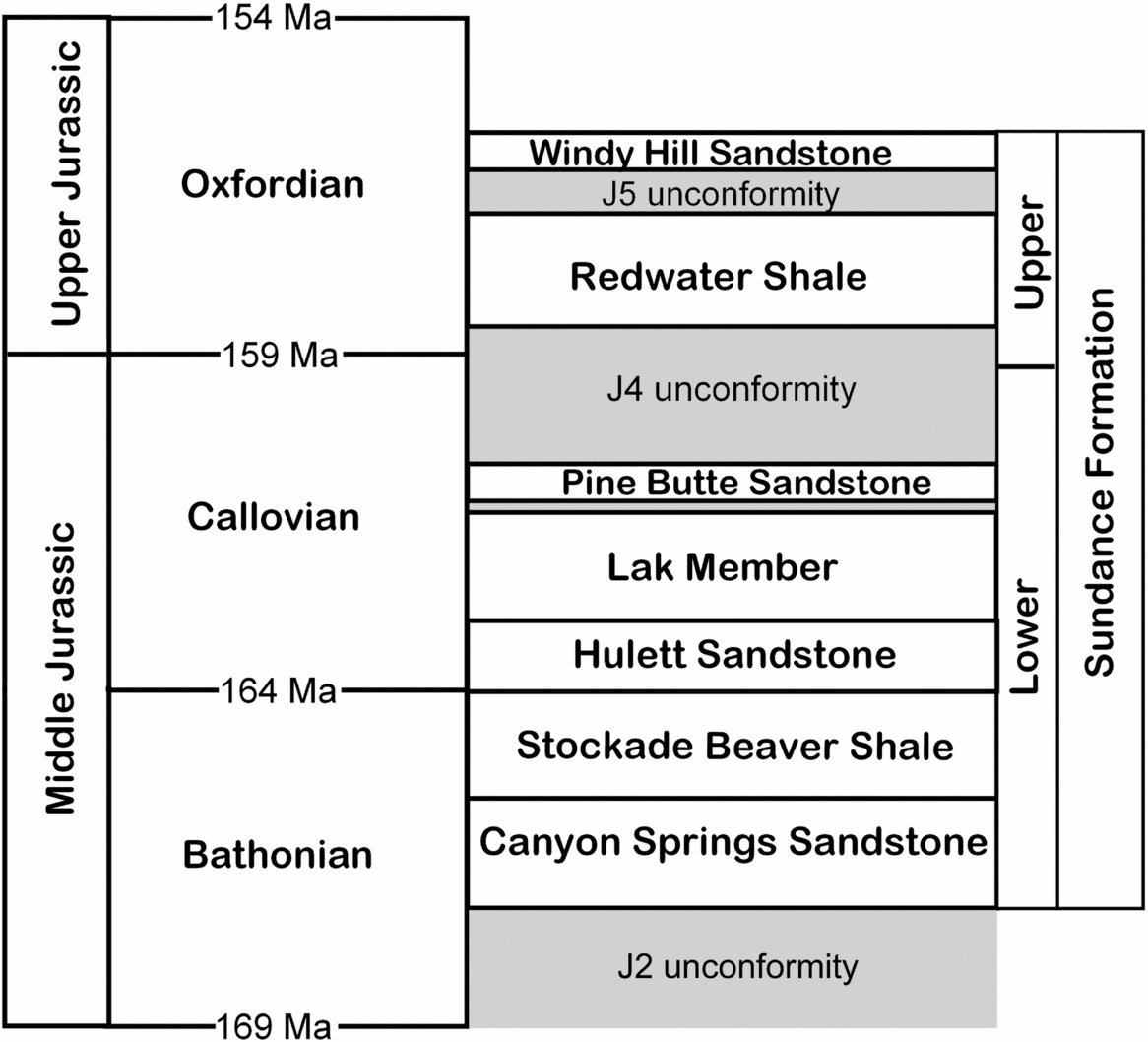
In south-central and northeastern Wyoming, the Sundance Formation has been subdivided into six members, from lower to upper: Canyon Springs Sandstone, Stockade Beaver Shale, Hulett Sandstone, Lak Member, Pine Butte Sandstone, and Redwater Shale (Fig. 3; Imlay, Reference Imlay1947; Robinson, Mapel & Bergendahl, Reference Robinson, Mapel and Bergendahl1964; Pipiringos, Reference Pipiringos1968; Johnson, Reference Johnson1992). The Redwater Shale Member has been recognized in central and north-central Wyoming by Imlay (Reference Imlay1980) and Johnson (Reference Johnson1992), and in Natrona County by Specht & Brenner (Reference Specht and Brenner1979). We have identified both the Pine Butte Sandstone and the Redwater Shale members in Natrona County, although the former varies greatly in thickness. Pipiringos (Reference Pipiringos1968) noted that belemnites are found only in the Redwater Shale Member, not in underlying strata of the Sundance Formation, and this is consistent with our identification of the belemnite-bearing strata as the Redwater Shale Member.
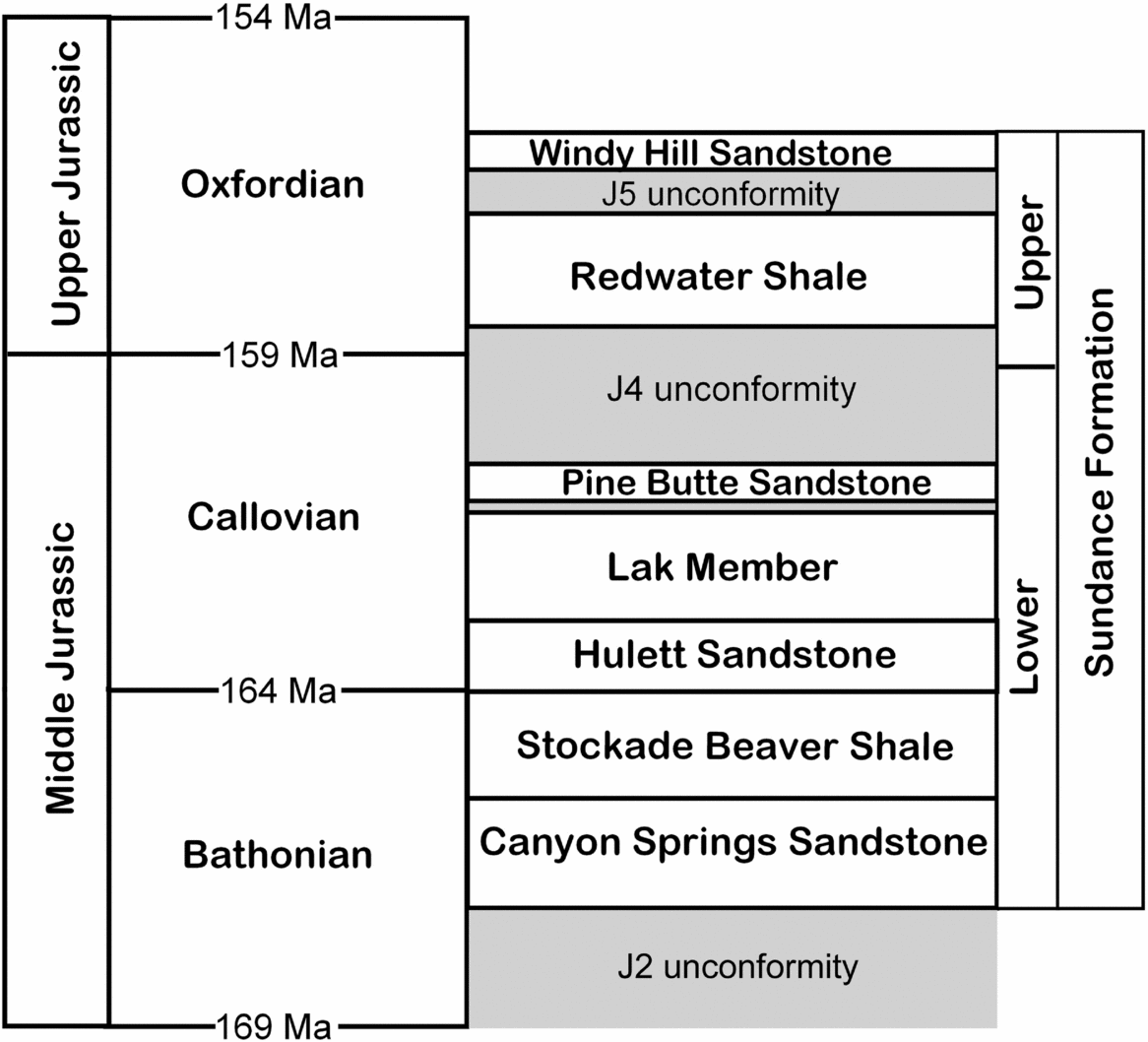
Figure 3. Members of the Sundance Formation showing the stratigraphical position of the Redwater Shale Member (Oxfordian), bounded by the J4 and J5 unconformities (modified from Johnson, Reference Johnson1992). The Windy Hill Sandstone is included as the upper member of the Sundance Formation, but others have included it in the overlying Morrison Formation (e.g. Peterson, Reference Peterson, Caputo, Peterson and Franczyk1994).
The Redwater Shale Member is a medium to dark grey, glauconitic shale with many thin beds of glauconitic sandstone. In northern Natrona County, three limestone coquinas occur, comprised of Camptonectes and/or Ostrea or Gryphaea shells, along with other bivalves, belemnites, and rare bone material. A thicker layer of green-grey cross-bedded, thin-bedded glauconitic sandstone occurs in the lower part of the Redwater Shale Member. Another distinctive tan to olive green fine sandstone with abundant trace fossils occurs higher in the section. Sandstones become more common towards the top of the Redwater Shale Member, reflecting the shallower water of the regressive phase. The soft shales of the Redwater Shale Member weather quickly to form vegetated slopes with outcrops of the resistant sandstones and limestone coquinas. Layers of iron-rich concretions also occur, and some of these concretions encase bone material, most often vertebrae and ribs.
The Redwater Shale Member is younger than the well known marine reptile fauna of the Peterborough Member of the Oxford Clay Formation (middle Callovian), England (Martill, Taylor & Duff, Reference Martill, Taylor and Duff1994; Martill & Hudson, Reference Martill and Hudson1991, text-fig. 1.2). Brenner & Peterson (Reference Brenner, Peterson, Caputo, Peterson and Franczyk1994) suggested an early to at least middle Oxfordian age for the Redwater Shale Member. Imlay (Reference Imlay1982) correlated the Redwater Shale Member in northeastern and southeastern Wyoming with the Cardioceras cordatum zone (upper lower Oxfordian). The Redwater Shale Member in the Bighorn Basin of north-central Wyoming, however, spans the Cardioceras cordatum and Perisphinctes plicatilis zones (upper lower Oxfordian to lower middle Oxfordian; Imlay, Reference Imlay1982). More recently, Kvale et al. (Reference Kvale, Johnson, Mickelson, Keller, Furer and Archer2001, figure 5) and McMullen (unpublished M.S. thesis, University of Georgia, 2012, figure 9) suggested a middle Callovian to middle Oxfordian age for the ‘upper’ Sundance Formation in north-central Wyoming (Bighorn Basin), but they included strata from the J3 to the J5 unconformities. The ammonites Cardioceras and Quenstedtoceras occur in the Redwater Shale Member at Alcova Reservoir in southwestern Natrona County (MR, pers. obs.), more in keeping with the Oxfordian age of Imlay (Reference Imlay1982).
3. Taphonomy of the marine reptiles
Reptilian remains are not confined to specific horizons. We have found specimens throughout the Redwater Shale Member, to within a couple of metres of the Windy Hill Sandstone (UW 34786; Massare et al. Reference Massare, Buchholtz, Kenney and Chomat2006). Thus the fauna represents an accumulation of skeletons over a fairly long interval of time. Most in situ material that we have found is ichthyosaurian. No complete skeletons have been found, although we have excavated a well-preserved skull and articulated anterior skeleton of an ichthyosaur (UW 24216). The skull was at nearly right angles to the bedding plane on which the articulated vertebral column rested, suggesting that the carcass landed on the sea floor in a ‘nose dive’, with the skull penetrating the soft substrate (Wahl, Reference Wahl2007b ). Articulated partial skeletons of both ichthyosaurs and plesiosaurs have been found (e.g. ichthyosaur UW 34653; plesiosaurs UW 15943, WDC SS01), although they are not common. Strings of articulated vertebrae and ribs, sometimes a substantial portion of the vertebral column, are more common (e.g. ichthyosaurs UW 15936, UW 24235, UW 19686, UW 34786). Disarticulated, associated bones have been collected in situ (e.g. ichthyosaurs UW 15936, TATE v-2038, UW 24802) and as float (e.g. plesiosaurs UW 15943, UW 24217; and ichthyosaurs UW 24234, UW 2). Isolated limb bones, pectoral elements, centra and ribs have also been collected as float.
The Redwater Shale Member does not typically form good outcrops, except for the limestone coquinas and thicker sandstone layers. Most areas in which it occurs are vegetated, and bones are often found as weathered float. Many of the more complete specimens are in irregularly shaped carbonate concretions within dark to medium grey shale layers or in sandstone lenses (F. R. O'Keefe, pers. comm. 2013). Some of the concretions include coarse shell hash (e.g. encasing UW 15936 and UW 34653), but a few are micritic (e.g. encasing UW 24216 and WDC SS-019). These concretions are distinct from the iron-rich concretions previously mentioned, which tend to be more ellipsoidal in shape and occur in distinct horizons. Both kinds of concretions can encase bone.
Burrows, probably formed by crustaceans, are associated with some specimens. UW 24216 had burrows running through the concretion along the ribs, and lobster carapace fragments (Eryma?) were found within the burrows by WRW as the specimen was prepared. One ichthyosaur (TATE v-2038) had sand-filled burrows in shale adjacent to the bones, as well as serpulid worm tubes in fine sediment immediately below the bones.
4. Palaeoenvironment
Based on the bivalve fauna, Wright (Reference Wright1974) considered the Redwater Shale Member to be a shallow, open shelf environment. The presence of belemnoids and crinoids would suggest normal marine salinity. Tang & Bottjer (Reference Tang and Bottjer1996) gave a depth estimate of less than 100 m. We suggest, however, that the depth was much less, in the order of 40–50 m, comparable to the 30–50 m depth that has been suggested for the Peterborough Member of the Oxford Clay Formation (Hudson & Martill, Reference Hudson, Martill, Tyson and Pearson1991).
S. K. McMullen (Unpublished M.S. thesis, University of Georgia, 2012) recognized two facies of the Redwater Shale Member in north-central Wyoming (Bighorn Basin), an offshore mudstone facies and a transitional facies of interbedded sandstone and mudstone. The former was interpreted as a low energy offshore environment that was below storm wave base. The latter was interpreted as a subtidal environment closer to the shoreline and frequently disturbed by waves (S. K. McMullen, unpublished M.S. thesis, University of Georgia, 2012). In northern Natrona County, layers of thin-bedded, cross-bedded sandstone occur in at least two horizons, and thin glauconitic sandstones occur throughout in the shale. Concretions around some ichthyosaur skeletons (UW 34653, UW 15963) contain coarse shell hash. So although shale is the dominant deposit, suggesting a quiet water environment below wave-base, frequent layers of fine sandstones and shell hash around skeletons indicate that the depths were near wave base. S. K. McMullen (Unpublished M.S. thesis, University of Georgia, 2012) indicated that the offshore facies had a higher abundance and diversity of vertebrates, but we have found vertebrates throughout the section in Natrona County, often immediately above (e.g. UW 15936, UW 19686) or below (e.g. UW 34786) thin beds of sandstone.
The Redwater Shale Member interval was a fairly rough water environment. Water-worn belemnites are fairly common, and water-worn bones or bone fragments occur as well. Andersson (Reference Andersson1979) described cobble layers of reworked concretions some of which were encrusted or bored by bivalves. He suggested that other cobble layers might be lag deposits from an interval of erosion. We have found cobbles that are encrusted by bivalves on the top and bottom, indicating currents or waves that were strong enough to turn them over. The limestone coquinas may also indicate rough water, or at least higher energy conditions. The coquinas have been interpreted as episodic storm deposits (Brenner & Davis, Reference Brenner and Davies1974), as storm-winnowed deposits on the higher areas of an irregular sea floor (Specht & Brenner, Reference Specht and Brenner1979), and as sand waves moved by tidal and storm currents (Brenner, Swift & Gaynor, Reference Brenner, Swift and Gaynor1985). All of those interpretations imply episodes of wave and current agitation reaching the seafloor.
Sedimentation rates were variable, but generally low as indicated by abundant glauconite throughout the Redwater Shale Member. Large carcasses on the sea floor may have served as baffles, accumulating coarser sediments within the shale. On one side of the limbs of the pliosaur Megalneusaurus rex, a thick sandstone lens occurs in the shale. It is possible that the carcass was exposed on the sea floor for some time before burial, blocking water flow so that coarser sediment was deposited on the leeward side. Concretions with shell hash suggest that this could have occurred with smaller carcasses as well. We have not, however, seen encrustations on bones of articulated vertebrate specimens, although encrusted belemnites occur. Another possible explanation is that the carcasses interfered with the movement of sand waves along the sea floor.
5. Fauna of the Redwater Shale Member
5.a. Macroinvertebrates
The invertebrate fauna of the Redwater Shale Member is dominated by molluscs. By far the most common invertebrate is the belemnoid Pachyteuthis densus. It occurs throughout the Redwater Shale Member, and has been found in the same horizons as vertebrate remains. A second, large belemnoid with a very deep phragmacone has been found (WDC JSI-22), and may represent a second, rare species. Doyle (Reference Doyle1987) suggested that Cylindroteuthis is also present. Belemnoids of the Redwater Shale Member have not been adequately studied, and so they may be more diverse than is currently thought. Belemnite guards with diameters of 1–2 mm, found in harvester ant mounds, suggest that the belemnoids could have been reproducing in the Sundance Sea. The lower Redwater Shale Member, in particular, has high concentrations of Pachyteuthis, and it is in these horizons that the anthills are covered with tiny guards. Belemnoids were a major food source for the reptiles because an ichthyosaur (UW 34653), both plesiosaur species (UW15943, UW 24215, and WDC SS-01), and the pliosaur (UW 4602) have been found with coleoid hooklets as gut contents (Massare & Young, Reference Massare and Young2005; Wahl, Ross & Massare, Reference Wahl, Ross and Massare2007; Wahl, Reference Wahl2012).
The ammonites Cardioceras and Quenstedtoceras occur in the Redwater Shale Member of central Wyoming. A cardiocerid ammonite jaw has been reported in the gut contents of a cryptocliedoid plesiosaur (WDC SS-001) from Hot Springs County (Wahl, Reference Wahl2012). Ammonite protoconchs can occasionally be found in harvester ant mounds in the lower Redwater Shale Member in northern Natrona County.
Wright (Reference Wright1974) identified seven bivalve species from the Redwater Shale Member in Wyoming and South Dakota, listed here in decreasing abundance: Camptonectes bellistriatus, Ostrea strigilecula, Gryphaea nebrascensis, Pleuromya newtoni, Tancredia sp., Meleagrinella curta, and Tancredia transversa. Camptonectes, Ostrea, Gryphaea, and Meleagrinella were epifaunal suspension feeders. Pleuromya and the two Tancredia species were infaunal, siphonate suspension feeders (Wright, Reference Wright1974). Kvale et al. (Reference Kvale, Johnson, Mickelson, Keller, Furer and Archer2001) reported Ostrea strigilecula, Vaugonia sturgisensis, Modiolis sp., Idonarca sp. as well as Meleagrinella curta and the two Tancredia species from the Redwater Shale Member equivalent in north-central Wyoming (Bighorn Basin). In both assemblages, the bivalve fauna are all suspension feeders, and a large proportion of them (Ostrea, Gryphaea, Meleagrinella, Camptonectes, Modiolis) are epifaunal (Wright, Reference Wright1974).
Pentacrinoid crinoids are the most common echinoderm in the Redwater Shale Member. We have found star-shaped crinoid columnals and stem pieces on harvester ant mounds in the lower Redwater Shale Member as well as elsewhere in the member. Crinoid fragments also make up the sediment of starved ripples and an encrinite in the lower Redwater Shale Member near Alkalai Creek in northern Natrona County (Wahl, Reference Wahl2007a ). Connely & Talbot (Reference Connely, Talbot, Woods and Lawlor2008) reported well preserved Isocrinus specimens in the Redwater Shale Member in southern Johnson County. Chariocrinus and Isocrinus have been identified from the Stockade Beaver Shale in the lower Sundance Formation (Hunter & Zonneveld, 1982; Koch, Reference Koch1972). Other echinoderms are rare. The starfish Eokainaster pewei has been reported from what might be the lower Redwater Shale Member of the Owl Creek Mountains (Miller & Unklesbay, Reference Miller and Unklesbay1943; Blake, Reference Blake1971) and from the Redwater Shale Member in the southern Bighorn Mountains (Connely & Talbot, Reference Connely, Talbot, Woods and Lawlor2008). Sea urchin spines can also be found in harvester ant mounds in the lower Redwater Shale Member. Sea urchin plates were found in the crinoid hash in the lower Redwater Shale Member and/or Pine Butte Member from Johnson County (Connely & Talbot, Reference Connely, Talbot, Woods and Lawlor2008) and at Como Bluff in southeastern Wyoming (MVC, pers. obs.). In addition, brittle stars are also known from the Stockade Beaver Shale (lower Sundance Formation) from these localities (Pipiringos, Reference Pipiringos1957; MVC, pers. obs).
Crustaceans were also a component of the benthic fauna of the Redwater Shale Member. WRW has found small pieces of crustacean carapaces (WDC JSI-10), which may be either Mecochirus or Eryma, in burrows associated with an ichthyosaur skeleton, UW 24216 (Wahl, Reference Wahl2008). A large piece of an Eryma carapace was found in the lower Redwater Shale Member (Wahl, Reference Wahl2004), and it has been reported from the equivalent Redwater Member of the Stump Formation in Utah (Feldman & Titus, Reference Feldmann and Titus2006).
5.b. Vertebrates
At least two kinds of chondrichthyan were present in the Sundance Sea. Wahl (Reference Wahl2005, Reference Wahl2007a ) described teeth, dermal denticles and dorsal fin spine denticles of a hybodont shark (UW 24218), possibly Hybodus or Asteracanthus, associated with the skeleton of a cryptocleidoid plesiosaur (UW 24215). Schaeffer & Patterson (Reference Schaeffer and Patterson1985) reported Hybodus from the Stockade Beaver Shale of the lower Sundance Formation. A hybodont shark tooth, as well as a long, thin tooth of a neoselachian shark, have been found in the coarse sands of starved ripples in the lower Redwater Shale Member (Wahl, Reference Wahl2007a ). Another single shark tooth was found in an encrinite in southern Johnson County (Connely & Talbot, Reference Connely, Talbot, Woods and Lawlor2008). Shark teeth are extremely rare, but it appears that at least two kinds of condrichthyan were present.
The osteichthyans Hulettia americana, Caturus dartoni, Lepidotes sp. and Occithrissops willsoni occur in the Stockade Beaver Shale (lower Sundance Formation; Fig. 3), Hulettia being the most common (Schaeffer & Patterson, Reference Schaeffer and Patterson1985). Dense, dark coloured bone and teeth, which can be attributed to some kind of osteichthyan fish, have been found in harvester ant mounds in the lower Redwater Shale Member (WRW, pers. obs). Similar dense bone fragments occur in the gut contents of Megalneusaurus (UW 4602) and Pantosaurus (WDC SS-01), and are associated with a partially articulated fragmentary skeleton of an ichthyosaur (TATE v-2038). Such fragments are neither common nor taxonomically identifiable, but they are clearly not reptilian bone fragments. The rough water environment and slow sedimentation rate may have inhibited preservation of small fish skeletons.
The most common vertebrate in the Redwater Shale Member is the ichthyosaur Ophthalmosaurus natans (Marsh, Reference Marsh1879). The ichthyosaur was first described as Sauranodon natans and later changed to Baptanodon natans when the genus name was found to be preoccupied (Marsh, Reference Marsh1880b ). Andrews (Reference Andrews1910) argued that Baptanodon was synonymous with Ophthalmosaurus, and most subsequent workers have agreed (but see Appleby, Reference Appleby1956). Recent phylogenetic analyses, however, have failed to recover a monophyletic Ophthalmosaurus (Druckenmiller & Maxwell, Reference Druckenmiller and Maxwell2010; Fischer et al. Reference Fischer, Maisch, Naish, Kosma, Liston, Joger, Krüger, Pardo Pérez, Tainsh and Appleby2012). Early workers recognized four additional species: B. discus (Marsh, Reference Marsh1880a ), B. marshi, Knight, Reference Knight1903; B. robustus, Gilmore, Reference Gilmore1906; and B. reedi, Gilmore, Reference Gilmore1907. Presently, only O. natans is considered valid (McGowan & Motani, Reference McGowan and Motani2003; Maisch & Matzke, Reference Maisch and Matzke2000), and we have not found any evidence to the contrary in the material that we have collected. Ophthalmosaurus natans can be distinguished from the closely related British species, O. icenicus, by the pattern of centrum shape change along the vertebral column. In particular, O. natans has longer centra relative to their height in the posterior dorsal and anterior caudal region than does O. icenicus (Massare et al. Reference Massare, Buchholtz, Kenney and Chomat2006). Appleby (Reference Appleby1956) also noted a difference in braincase features, especially the relative contribution of the supraoccipital and exoccipitals to the foramen magnum (but see discussion in McGowan & Motani, Reference McGowan and Motani2003, p. 111).
Specimens of Ophthalmosaurus fall into a range of size classes, including centra of a presumably voided embryo found as gut contents of a cryptocleidoid plesiosaur (O'Keefe et al. Reference O'Keefe, Street, Cavigelli, Socha and O'Keefe2009). The largest specimen we have collected has a skull length of 80 cm (Fig. 4), which would correspond to a total length, from the tip of the snout to the base of the tail fluke, of 3.2 m if we assume the same body proportions as O. icenicus (Andrews, Reference Andrews1910, text-figure 42). Massare et al. (Reference Massare, Buchholtz, Kenney and Chomat2006) estimated a preflexural length of the vertebral column of a large, composite specimen as 2.8 m. Again, assuming proportions similar to O. icenicus, this would suggest a length of about 3.5 m including the skull. Most specimens, however, are considerably smaller.

Figure 4. UW 24216 Ophthalmosaurus natans skull, approximately 80 cm long. The specimen settled to the sea floor apparently in a nose dive as the skull was at nearly a right angle to bedding and the post-cranial skeleton. Scale bar represents 10 cm.
Two long-necked cryptocleidoid plesiosaurs are known from the Redwater Shale Member, Pantosaurus striatus and Tatenectes laramiensis (O'Keefe & Wahl, Reference O'Keefe and Wahl2003a , Reference O'Keefe and Wahl b ). Both species are endemic to the Sundance Sea. Juvenile plesiosaur material has also been collected, although the material is insufficient for taxonomic identification (Wahl, Reference Wahl2006). It is likely, however, that at least one species of plesiosaur was breeding in the Sundance Sea.
The plesiosaur Tatenectes was originally described as ‘Cimoliosaurus’ laramiensis by Knight (Reference Knight1900), but the holotype is lost. It is the same taxon as ‘Tricleidus’ laramiensis (Mehl, Reference Mehl1912). O'Keefe & Wahl (Reference O'Keefe and Wahl2003b ) created a new genus for the species, erected a neotype based on the original description, and noted its close relationship to Kimmerosaurus. The relationship was confirmed by O'Keefe & Street (Reference O'Keefe and Street2009), who placed Tatenectes within the ‘Aristonectidae’, with Kimmerosaurus as a sister taxon. O'Keefe et al. (Reference O'Keefe, Street, Wilhelm, Richards and Zhu2011) estimated the adult body length as just under 3 m, although the length of the neck is uncertain. They noted that Tatenectes has a strongly dorsoventrally compressed body and the pelvic girdle has shifted anteriorly, reducing the distance between the limbs. The extreme compression and anteroposterior shortening gave Tatenectes an unusual body shape for a plesiosaur (O'Keefe et al. Reference O'Keefe, Street, Wilhelm, Richards and Zhu2011).
Pantosaurus striatus (Marsh, Reference Marsh1891), the second cryptocleidoid plesiosaur from the Redwater Shale Member, was originally described as Parasaurus striatus, but the generic name was changed because it was preoccupied (O'Keefe & Wahl, Reference O'Keefe and Wahl2003a ). The holotype (YPM 543) originally included teeth and cranial material, but that portion of the specimen has been lost (O'Keefe & Wahl, Reference O'Keefe and Wahl2003a ). The taxonomic affinities are made more difficult to ascertain because the holotype is a juvenile (O'Keefe & Wahl, Reference O'Keefe and Wahl2003a ). Overall, the taxon is less well known than Tatenectes. The cervical centra are ‘waisted’, where the middle of the centrum has a smaller diameter than the articular faces, and the centra are almost as long as they are wide, two features that are among those diagnostic of the species (Wilhelm & O'Keefe, Reference Wilhelm and O'Keefe2010). O'Keefe & Wahl (Reference O'Keefe and Wahl2003a ) noted that the centrum morphology was similar to that of Muraenosaurus, but specimens are too incomplete for a phylogenetic analysis. Muraenosaurus reedi Mehl, Reference Mehl1912, described from the Sundance Formation, is considered to be a junior synonym of P. striatus (O'Keefe & Wahl, Reference O'Keefe and Wahl2003a ), suggesting that earlier workers also noted a similarity to Muraenosaurus. Pantosaurus has more robust limb bones than Tatenectes (O'Keefe & Wahl, Reference O'Keefe and Wahl2003a ) and so may have been a somewhat larger species.
Megalneusaurus rex is the only pliosauromorph in the Redwater Shale Member, although the rhomaleosaurid Borealonectes is known from the Callovian of northern Canada (Sato & Wu, Reference Sato and Wu2008). The type specimen (UW 4602) was collected in 1895 and provisionally assigned to Cimoliosaurus (Knight, Reference Knight1895), but subsequently assigned to a new genus, Megalneusaurus (Knight, Reference Knight1898). Two hind limbs, one articulated and one disarticulated, are all that remain of the original holotype. A poorly preserved but articulated forelimb of the same individual was collected in 2008 from the original site (Wahl, Massare & Ross, Reference Wahl, Massare and Ross2010). The limbs do not provide useful taxonomic information, but the shape of the limbs suggest that Megalneusaurus may be a pliosaurid. It is the rarest of the marine reptiles, with only three other specimens known from the Redwater Shale Member, all of which are isolated, weathered bones collected as float (UW 24238-neural arch; WDC SS-019-propodial fragment; USNM 536978). The only in situ specimen of Megalneusaurus (UW 4602, the type specimen) occurs in the upper Redwater Shale Member, within 10 m of the top of the unit (Wahl, Ross & Massare, Reference Wahl, Ross and Massare2007), and so the pliosaur seems to have appeared during the regressive phase of the Redwater Shale Member. If we assume the same body proportions as for Liopleurodon (Robinson, Reference Robinson1976, figure 10A), the 106 cm partial forelimb that we collected (Wahl, Massare & Ross, Reference Wahl, Massare and Ross2010) would suggest a total length of 9.2 m for the holotype individual. Megalneusaurus appears to have been more wide ranging than the other plesiosaurs, being known from the Alaskan peninsula (Weems & Blodgett, Reference Weems and Blodgett1994), as well as from Wyoming.
6. Palaeoecology of the vertebrates
Ophthalmosaurus natans was a pursuit predator, but had a more undulatory swimming style than O. icenicus, based on the shape of the centra (Massare et al. Reference Massare, Buchholtz, Kenney and Chomat2006). It may not have been as fast and efficient a swimmer as O. icenicus of comparable size, but it was probably more maneuverable (Massare, Reference Massare, Maddock, Bone and Rayner1994; Massare et al. Reference Massare, Buchholtz, Kenney and Chomat2006). O. natans has slender, fairly pointed, but small teeth (Fig. 5), characteristic of the Pierce I guild of Massare (Reference Massare1987). For UW 24216, with a skull length of 80 cm, tooth crown heights range from 0.5 mm to 2 cm, with the largest preserved tooth being 3.5 cm high, including the root. The largest teeth appear to be anterior to the maxilla, and are smaller at the anterior end of the rostrum. Gut contents of UW 34653 confirm that the species consumed coleoid cephalopods, fairly soft prey as would be predicted from the tooth form (Massare, Reference Massare1987; Massare &Young, Reference Massare and Young2005). UW 24216 preserves teeth throughout the entire rostrum, although most are dislodged from their natural position. O. natans had a complete dentition as an adult, as asserted by Gilmore (Reference Gilmore1905, p. 99). Teeth can be seen along the entire length of the jaw in UW 24216. The specimen preserved teeth because the skeleton landed head first on the sea floor and the rostrum was driven into the sediment (Wahl, Reference Wahl2007b ). The encasing sediment held the teeth in place, even at the anterior end of the rostrum that was split open (Fig. 4). The reported lack of teeth or only anterior teeth in Ophthalmosaurus natans specimens (e.g. McGowan & Motani, Reference McGowan and Motani2003, p. 110) is probably an artefact of preservation.

Figure 5. UW 24216 Ophthalmosaurus natans teeth. (a) Teeth from the posterior portion of the rostrum (but anterior to the nares). Scale bar represents 3 cm. (b) Tooth from the anterior portion of the rostrum. Scale bar represents 2 cm.
Animals living in shallow water, such as the relatively shallow Sundance Sea, are continuously subjected to waves and current, which provide the stress to maintain and increase bone density (Sheldon, Reference Sheldon, Callaway and Nicholls1997). Rib cross-sections of UW 24216 show a thick layer of cortical bone and thick trabeculae (Fig. 6), somewhat similar to what has been reported for another ophthalmosaurid, Mollesaurus (Talevi & Fernández, Reference Talevi and Fernández2012). Bone microstructure thus suggests that O. natans was inhabiting the Sundance Sea, not migrating in for breeding or feeding. It also implies that it was not a deep diving species.

Figure 6. UW 24216 Ophthalmosaurus natans cross-section of ribs showing thick trebeculae. Rib cross-section changes shape from the head to the distal end. (a) is from a more proximal location on a rib than (b). Scale bars represent 1.5 cm.
However, the exceptionally large eye of Ophthalmosaurus natans has been interpreted as an adaptation for low light levels encountered in deep diving, perhaps as deep as 600 m or more (Motani, Rothschild & Wahl, Reference Motani, Rothschild and Wahl1999). Although O. icenicus may have been a deep diver, bone microstructure and the environment of the Sundance Sea suggest that it was not the case for O. natans. Humphries & Ruxton (Reference Humphries and Ruxton2002) argued that visual acuity, the ability to resolve fine details, is as important as sensitivity to low light levels. They suggested that ichthyosaurs may have been selective hunters of small prey at moderate depths, needing large eyes to distinguish among prey species. So possibly the large eye of O. natans allowed it to hunt in low light conditions at dawn or dusk, perhaps coinciding with a time of day when belemnoids were active. Nilsson et al. (Reference Nilsson, Warrant, Johnsen, Hanlon and Shashar2012), however, suggested that an eye size of about 9.0 cm diameter (3.0 cm pupil diameter) was the upper limit for optimal sensitivity and acuity. Larger eyes did not substantially improve either and so selection was probably for another aspect of vision. In the largest Ophthalmosaurus that we have found (UW 24216; Fig. 4), the outer diameter of the sclerotic ring is about 20 cm and the inner diameter (diameter of the opening) is 8 cm. Nilsson et al. (Reference Nilsson, Warrant, Johnsen, Hanlon and Shashar2012) hypothesized that selection for such very large eyes was to enhance long-range recognition of large objects, such as predators or potential mates. Or large eyes may have evolved to use visual signals to coordinate group hunting or for recognition of other ichthyosaurs (Humphries & Ruxton, Reference Humphries and Ruxton2002). In any event, an exceptionally large eye, as seen on O. natans, need not imply deep diving.
The cryptocleidoid plesiosaurs were about the same size as the ichthyosaurs. With its highly dorso-ventrally flattened body (O'Keefe et al. Reference O'Keefe, Street, Wilhelm, Richards and Zhu2011), Tatenectes was not a fast swimmer, and because of its long neck may have favoured an ambush strategy (Massare, Reference Massare1988, Reference Massare, Maddock, Bone and Rayner1994). Tatenectes had large gastralia that are pachyostotic along their midline, adaptations for stability in shallow water (Wahl, Reference Wahl and Hunter1999; Street & O'Keefe, Reference Street and O'Keefe2010). Thus like O. natans, Tatenectes was a shallow water species, and was probably living in the Sundance Sea rather than migrating in from the open ocean to the north. Its teeth were small, slender and pointed (O'Keefe & Street, Reference O'Keefe, Street, Cavigelli, Socha and O'Keefe2009), and it occupied the Pierce I guild (Massare, Reference Massare1987). Coleoid hooklets were found associated with a partial Tatenectes skeleton (UW 15943), so belemnoids probably made up a major portion of its diet.
The body shape of Pantosaurus is not known, but if it had the typical cryptocleidoid body shape (small head and fairly long neck), Pantosaurus would have also tended towards an ambush style of attack, or at least slower swimming speeds (Massare, Reference Massare1988, Reference Massare, Maddock, Bone and Rayner1994). Pantosaurus may have been a more opportunistic predator than Tatenectes, as specimens have been found with gut contents that include coleoid hooklets (UW24215 and WDC SS-01), an ammonite jaw (WDC SS-01), small fragments of fish bone (WDC SS-01), and a scavenged ichthyosaur embryo (O'Keefe et al. Reference O'Keefe, Street, Cavigelli, Socha and O'Keefe2009). However, its tooth form is unknown.
The pliosaur Megalneusaurus was much larger than the other reptiles. It was the fastest predator in the sea, if for no other reason than its large size. It could have easily consumed juveniles and subadults of the other reptiles. Bite marks are known on a small plesiosaur propodial (UW 24219, Fig. 7; Wahl, Reference Wahl2006), although neither the species of the victim nor the predator can be determined. Stomach contents have been recovered for the type specimen (UW4602) of Megalneusaurus, and these consist of coleoid hooklets, as occurs with the other reptiles, along with a few rare fish bone fragments (Wahl, Ross & Massare, Reference Wahl2007). Megalneusaurus appeared towards the end of the Redwater Shale Member deposition, when the Sundance Sea was experiencing the last regressive phase. Because it is so rare, it may be that the pliosaur was a visitor rather than an inhabitant of the area, but this is impossible to determine with any certainty. However, Megalneusaurus is the only marine reptile in the Redwater Shale Member fauna to occur outside of Wyoming (Weems & Blodgett, Reference Weems and Blodgett1994).

Figure 7. UW 24219, a portion of a small plesiosaurian propodial showing bite marks, four circular depressions, at the distal end. Specimen on display at the Tate Geological Museum, Casper College, Casper, WY, USA. Scale bar represents 5 cm.
7. Comparison with the Peterborough Member, Oxford Clay Formation
One of the best known and diverse marine vertebrate faunas of the Jurassic is that of the Peterborough Member of the Oxford Clay Formation. It is mainly Middle Callovian in age (upper Calloviense Zone through the Athleta Zone; Martill & Hudson, Reference Hudson, Martill, Tyson and Pearson1991, text-fig.1.2), older than the Redwater Shale Member by a few million years. The deposition of the Peterborough Member was estimated to have been a period of just two million years, based on the average duration of ammonite zones (Martill, Taylor & Duff, Reference Martill, Taylor and Duff1994). By comparison, the Redwater Shale Member spans one ammonite zone (Imlay, Reference Imlay1982) but the duration has been estimated as at least four million years (Kvale et al. Reference Kvale, Johnson, Mickelson, Keller, Furer and Archer2001), and perhaps as much as eight million years (Brenner & Peterson, Reference Brenner, Peterson, Caputo, Peterson and Franczyk1994). In spite of the shorter time frame, the Peterborough Member has a much higher diversity of vertebrates and invertebrates, although some of the same genera are present in both faunas. This discussion will focus on the vertebrates.
A major difference between the Redwater Shale Member and the Peterborough Member is that the latter has a much more diverse fish fauna, with 27 genera, including both chondrichthyans and osteichthyans (Martill, Taylor & Duff, Reference Martill, Taylor and Duff1994). In fact, fish are the most abundant vertebrates in the Peterborough Member (Martill & Hudson, Reference Hudson, Martill, Tyson and Pearson1991). Even if we assume that all of the fish species in the Stockade Beaver Shale survived to Redwater Shale Member time, only five fish genera are known (Schaeffer & Patterson, Reference Schaeffer and Patterson1985). Four of the lower Sundance Formation genera (Caturus, Lepidotes, Hybodus, and Ischyodus) are also found in the Peterborough Member, and the first two genera are among the most common fish fossils (Martill, Taylor & Duff, Reference Martill, Taylor and Duff1994). The most common fish in the lower Sundance Formation is Hulettia, which appears to be endemic to the Sundance Sea (Schaeffer & Patterson, Reference Schaeffer and Patterson1985). Notably, remains of the giant suspension feeding fish, Leedsichthys, which are frequent in the Peterborough Member (Martill, Taylor & Duff, Reference Martill, Taylor and Duff1994), are absent in the Redwater Shale Member. Thus fish made up a much more significant component of the food web in the Peterborough Member, and probably were prey for many marine reptiles. Additionally, the relationships among the fish species themselves were quite complex (Martill, Taylor & Duff, Reference Martill, Taylor and Duff1994, figure 7). In contrast, it appears that belemnoids were the major food source for the reptiles in the Redwater Shale Member, as indicated by preserved gut contents and their abundance compared with that of other possible prey species.
An ophthalmosaurid ichthyosaur is a major component of both faunas. Based on the collection of specimens from the Thirty-three Mile Road area in northern Natrona County (Fig. 2), ichthyosaurs make up at least 58% the Redwater Shale Member fauna (Table S1, online Supplementary Material available at http://journals.cambridge.org/geo), whereas they are only about a quarter (23%) of the Peterborough Member fauna (Martill & Hudson, Reference Hudson, Martill, Tyson and Pearson1991, text-figure 8.1). Only one species, Ophthalmosaurus icenicus, is recognized from the Peterborough Member (A. M. Kirton, unpublished Ph.D. thesis, Newcastle upon Tyne, 1983; but see Appleby, Reference Appleby1956). With its deep caudal fin and streamlined body, O. icenicus was an efficient pursuit predator and could probably sustain higher speeds than O. natans of comparable size (Massare, Reference Massare1988; Massare et. al. Reference Massare, Buchholtz, Kenney and Chomat2006). Ophthalmosaurus natans had a more flexible vertebral column that suggests greater maneuverability but slower sustained swimming speed (Massare et al. Reference Massare, Buchholtz, Kenney and Chomat2006). The difference in swimming styles suggests a difference in how prey was located and captured. Although no gut contents are preserved, Martill, Taylor & Duff (Reference Martill, Taylor and Duff1994) argued that the diet of O. icenicus was likely soft cephalopods, in part because of the very slender, elongate rostrum. Preserved gut contents indicate that O. natans also fed on cephalopods, specifically belemnoids, although O. icenicus had many more species to select from. Martill, Taylor & Duff (Reference Martill, Taylor and Duff1994) considered O. icenicus to be a deep diving form and thought that it may have been a migrant, ranging well beyond the Oxford Clay basin. This is in contrast to our interpretation of O. natans as predominantly shallow water species that was endemic to the shallow Sundance Sea.
A major difference in diversity occurs in the Plesiosauria, with more than three times as many species in the Peterborough Member than in the Redwater Shale Member (Martill & Hudson, Reference Martill and Hudson1991). The Peterborough Member preserves six species (assigned to four genera) of long-necked plesiosaurs, compared to two species in the Redwater Shale Member. The long-necked plesiosaurs are 38% of the Peterborough fauna (Martill & Hudson, Reference Martill and Hudson1991, text-figure 8.1) about the same proportion as in the Redwater Shale Member (39%; Table S1, online Supplementary Material available at http://journals.cambridge.org/geo). Martill, Taylor & Duff (1994; table 1) reconstructed the Peterborough plesiosaurs as surface dwelling invertebrate feeders (Cryptoclidus, Tricelidus) and more generalist surface feeders (Muraenosaurus). In addition, the very rare plesiosaur Pachycostasaurus has been interpreted as a slow swimmer that fed on nektobenthic prey such as arthropods, cephalopods, or ganoid fish (Cruickshank, Martill & Noé, Reference Cruickshank, Martill and Noè1996). The Redwater Shale Member is similar to the Peterborough Member in the two common feeding types of long-necked plesiosaurs, although it had far fewer species.
In addition, the Peterborough Member includes six species of pliosauromorphs, representing five genera and comprising 12% of the fauna (Martill & Hudson, Reference Martill and Hudson1991, text-figure 8.1; Ketchum & Benson, Reference Ketchum and Benson2011). These include large opportunistic carnivores (Liopleurodon, Simolestes) as well as generalists, fish and cephalopod feeders (Peloneustes, ‘Pliosaurus’; Martill, Taylor & Duff, Reference Martill, Taylor and Duff1994), and a rare, gracile longirostrine form (Marmornectes; Ketchum & Benson, Reference Ketchum and Benson2011) that may have been a more specialized feeder. The Redwater Shale Member preserves a single rare species of pliosaur, comprising less than 3% of the fauna (Table S1, online Supplementary Material available at http://journals.cambridge.org/geo). By virtue of its size, Megalneusaurus was probably an opportunistic predator that was capable of preying upon the other marine reptiles, although the only preserved gut contents indicate a diet of mainly cephalopods (Wahl, Ross & Massare, Reference Wahl2007). Overall, the Plesiosauria, especially the pliosauromorphs, were much more ecologically and taxonomically diverse in the Peterborough Member than in the Redwater Shale Member. Moreover, the plesiosaurs comprise 50% of the reptilian fauna of the Peterborough Member, twice as abundant as the ichthyosaurs. In the Redwater Shale Member, the ichthyosaurs are much more abundant than the plesiosaurs (Table S1, online Supplementary Material available at http://journals.cambridge.org/geo).
The most noticeable difference between the reptilian faunas, however, is the lack of marine crocodiles in the Redwater Shale Member. Two families of marine crocodiles, metriorhynchids and teleosauids, make up 22% of the Peterborough Member specimens (Martill & Hudson, Reference Martill and Hudson1991 text-figure 8.1). Martill & Hudson (Reference Martill and Hudson1991) listed two species of the teleosaurid genus Steneosaurus, but a recent phylogenetic analysis by Young et al. (Reference Young, Andrade, Desojo, Beatty, Steel, Fernández, Sakamoto, Ruiz-Omeñaca and Schoch2012) suggested that there are at least three species. The metriorhynchids include four species, representing the genera Metriorhynchus, Gracilineustes, Suchodus, and the rare hypercarnivore Tyrannoneustes (Young et al. Reference Young, Andrade, Brusatte, Sakamoto and Liston2013). The marine crocodiles occupied the Crunch, Pierce II, and Cut guilds of Massare (Reference Massare1987; Young et al. Reference Young, Brusatte, Ruta and Andrade2010), and thus were quite ecologically diverse. The complete absence of these ambush predators in the Redwater Shale Member fauna is puzzling, especially as they also occur in the Middle Jurassic of South America (e.g. Gasparini, Cichowolski & Lazo, Reference Gasparini, Cichowolski and Lazo2005; Fernández & Herrera, Reference Fernández and Herrera2009). A metriorhynchid crocodile has been reported from the Middle Jurassic (Aalenian–Bajocian) Snowshoe Formation of eastern Oregon, however, the area is part of the Izee terrane, which was accreted in the Upper Jurassic (E. W. Wilberg, unpublished Ph.D. thesis, University of Iowa 2012). Thus a marine crocodile inhabited the eastern Pacific in the Middle Jurassic, but it did not necessarily inhabit the North American coast.
It is possible that the marine crocodiles were excluded because of cool temperatures associated with the flow of water from the northern boreal ocean. M. Ploynoi (Unpublished M.S. thesis, Wichita State College, Kansas, 2007) estimated an average water temperature of 15–17°C for the Sundance Sea from oxygen isotope measurements on belemnites. Hudson & Martill (Reference Hudson, Martill, Tyson and Pearson1991) cited temperatures of 17–20°C for the Peterborough Member, similarly based on pelagic species. More recently, Anderson et al. (Reference Anderson, Popp, Williams, Ho and Hudson1994) calculated a mean temperature of 15°C from belemnites, 17°C from the bivalve Gryphaea, and 21°C from the ammonite Kosmoceras, although the latter may be 3–4°C too high because of possible fractionation. Thus oxygen isotope analyses do not indicate a substantially different water temperature for the Redwater Shale Member compared to the Peterborough Member. Although the Redwater Shale Member might have been a few degrees cooler, it is unlikely to have affected the distribution of crocodiles. Crocodiles can tolerate those temperatures, although a mean annual temperature of 25–35°C is the preferred range (Markwick, Reference Markwick1998). Young et al. (Reference Young, Brusatte, Ruta and Andrade2010) suggested that the Oxfordian expansion of the metriorhynchids failed to reach the Sundance Sea before the final regression occurred. Or perhaps the Redwater Shale Member was too far from the shoreline to be a suitable habitat for marine crocodiles (Hudson & Martill, Reference Hudson, Martill, Tyson and Pearson1991, fig. 3).
Five percent of the fauna of the Peterborough Member is represented by terrestrial species, dinosaurs that washed in, and an additional 1% are pterosaurs (Martill & Hudson, Reference Martill and Hudson1991, text-figure 8.1). No bones of terrestrial animals have been found in the Redwater Shale Member, although pterosaur tracks occur in the overlying Windy Hill Sandstone (Connely, Reference Connely2006). Wood fragments occur in Natrona County, but they are extremely rare. They have also been reported from western nearshore strata (Brenner & Davies, Reference Brenner and Davies1974). This probably indicates that the Redwater Shale Member environment, at least in central Wyoming, was an open but shallow ocean, much farther from the shoreline than the Peterborough Member.
Figures 8 and 9 depict the trophic structure of the Redwater Shale Member and the Peterborough Member, respectively. Belemnoids form the basis of the food web for marine reptiles in the Redwater Shale Member (Fig. 8), whereas fish are a major component, if not the most important food source for the reptiles, in the Peterborough Member (Fig. 9). This difference in the trophic structure probably contributed to the difference in diversity of the marine reptiles. The marine reptiles are much more diverse in the Peterborough Member because of the more diverse and abundant assemblage of potential prey. Not only were there more fish, but there were more belemnoid and ammonoid species as well. Of course, this does not address the question of why so many more prey species were available in the Peterborough Member (see Conclusion below).
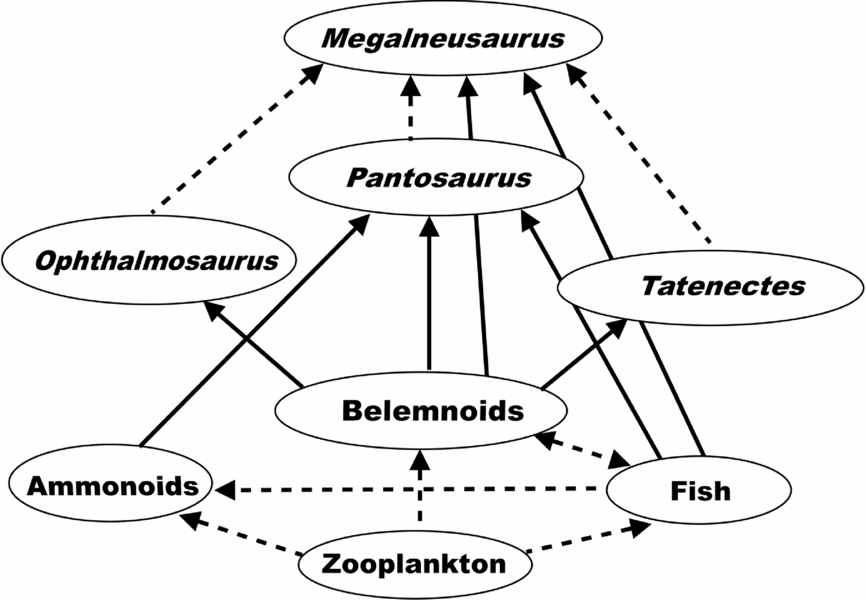
Figure 8. Food web for the Redwater Shale Member of the Sundance Formation. Dashed lines indicate inferred relationships. Solid lines indicate relationships supported by preserved stomach contents.

Figure 9. A simplified food web for the Peterborough Member of the Oxford Clay Formation, modified from Martill, Taylor & Duff (Reference Martill, Taylor and Duff1994). Unlike Figure 8, solid lines do not indicate relationships supported by preserved stomach contents. All relationships are inferred. Star indicates a rare taxon. Very rare taxa (e.g. Simolestes, Pachycostasaurus, Marmornectes) are omitted.
8. Conclusion
A number of non-biological factors may contribute to the lower diversity of vertebrates in the Redwater Shale Member as compared to the Peterborough Member. The Redwater Shale Member lacks any quarry or shoreline exposures, but instead forms vegetated slopes with few outcrops. Moreover, the Redwater Shale Member has not been collected for as long a time by as many workers as the Peterborough Member. Our research, and that of O'Keefe and his students, represents the first focus on the marine reptiles of the Sundance Formation since pioneering work in the late nineteenth and early twentieth centuries by O. C. Marsh, W. C. Knight, W. Reed, and C. W. Gilmore. Martill & Hudson (Reference Martill and Hudson1991, text-figure 8.1) estimated the relative proportions of marine reptiles of the Peterborough Member from over 1000 museum specimens. By comparison, the number of reptilian specimens from the Redwater Shale Member is probably less than 20% of that number. Thus the limited exposures and limited collecting may contribute to the lower diversity of the Redwater Shale Member. Preservation bias may come into play as well, especially in terms of the fish preservation. The Sundance Sea was probably close to wave base depth for much of its duration, and rough water, frequent storm deposits, and erosion may have reduced the likelihood of preservation of articulated skeletons of small vertebrates such as fish.
These factors, however, are inadequate to explain the difference in diversity. The Redwater Shale Member has a depauperate fauna even taking the aforementioned biases into account. Fewer prey species limited the diversity of the large reptilian predators. It is not uncommon for physically stressful environments to have lower diversity, although low habitat heterogeneity could also reduce diversity (Therriault & Kolasa, Reference Therriault and Kolasa2000). The rough-water environment of the Sundance Sea may have exceeded the tolerances of potential prey species. More likely, however, the outlet to the north limited the number of species that could migrate into the Sundance Sea. Martill, Taylor & Duff (Reference Martill, Taylor and Duff1994) thought that at least some of the Peterborough fauna were migrants from deeper water (see also Hudson & Martill, Reference Hudson, Martill, Tyson and Pearson1991, fig. 3). A more extensive connection to the open ocean may have allowed a greater variety of species to frequent the Oxford Sea. By comparison, the Redwater Shale Member fauna includes mainly shallow water species, and the relatively narrow outlet to the north may have been a bottleneck for open ocean species trying to enter the Sundance Sea.
The low diversity of pelagic invertebrates, fish, and benthic invertebrates in the Redwater Shale Member is probably indicative of low primary productivity in the pocket sea, perhaps due to poor circulation, distance from the shoreline, or low sedimentation rates. By comparison, the Peterborough Member is dominated by bituminous shales, and the high carbon content, up to 10% in some strata, indicates very high primary productivity (Duff, Reference Duff1975; Hudson & Martill, Reference Hudson, Martill, Tyson and Pearson1991). Notably, vertebrate remains occur in shales with the highest carbon content, suggesting an association with times of highest primary productivity (Hudson & Martill, Reference Hudson, Martill, Tyson and Pearson1991). It is probably this higher primary productivity that allowed the Peterborough Member to attain a very high diversity of both invertebrates and vertebrates. The low diversity in the Redwater Shale Member probably reflects a much lower primary productivity, as well as restricted migration from the open ocean to the north.
Acknowledgements
Field work has been aided by an army of Tate Museum volunteers, several undergraduate field assistants from SUNY Brockport and Wellesley College, and more recently, volunteers from the Wyoming Dinosaur Center. Colleagues have joined us in the field over the years, and we greatly appreciated their company, assistance, and discussions. We thank the Bureau of Land Management for supporting our research through permits for surface collection and excavations, including PA02-WY-075, PA02-WY-076, PA05-WY-130, PA05-WY-131, and PA08-WY-165 issued to JAM. We are grateful to the University of Wyoming and the Tate Geological Museum for serving as repositories for the specimens collected on BLM administered land. We also thank the Wyoming Dinosaur Center for the use of their fossil preparation lab. Finally we thank Erin Maxwell, Mark Young and Robin O'Keefe for their thoughtful reviews and suggestions. Some field work by JAM was partially funded by Scholarly Incentive grants from SUNY Brockport.









{kind=link}