



1. Introduction
Sediment stabilization by microbial binding has a key role in understanding the fossil record and the evolutionary palaeoecology of early metazoan life, particularly in the Ediacaran marine ecosystems and during the Proterozoic–Phanerozoic transition, i.e. times with widespread matgrounds (Seilacher, Reference Seilacher1999; Bottjer et al. Reference Bottjer, Hagadorn and Dornbos2000; Schieber et al. Reference Schieber, Bose, Eriksson, Sarkar, Schieber, Bose, Eriksson, Banerjee, Sarkar, Altermann and Catuneanu2007; Mata & Bottjer, Reference Mata and Bottjer2009). During the formation of microbially induced sedimentary structures (MISS; Noffke et al. Reference Noffke, Gerdes, Klenke and Krumbein1996, Reference Noffke, Gerdes, Klenke and Krumbein2001), changes in environmental parameters are key in controlling the compatibility potential of MISS-producing microorganisms (Bayet-Goll & Daraei, Reference Bayet-Goll and Daraei2020; Duperron & Scasso, Reference Duperron and Scasso2020). In addition to restricting environmental parameters, however, the effects of organism–matground interactions (Buatois & Mángano, Reference Buatois and Mángano2011a; Buatois et al. Reference Buatois, Netto, Mángano and Carmona2013) on the size, patterns and morphological diversity of MISS in various depositional environments should also be considered in the study of benthic microbial communities in deep time (Bayet-Goll et al. Reference Bayet-Goll, Daraei, Geyer, Bahrami and Bagheri2021 a, b). The early Cambrian diversification of marine benthos, known as the ‘Cambrian agronomic revolution’, enhanced infaunalization and promoted vertically oriented bioturbating organisms in shallow-marine ecosystems (Seilacher & Pflüger, Reference Seilacher, Pflüger, Krumbein, Peterson and Stal1994; Bottjer et al. Reference Bottjer, Hagadorn and Dornbos2000; Buatois & Mángano, Reference Buatois and Mángano2011 a, b), leading to the decrease of microbial matgrounds and their restriction to environments inhospitable for shallower-water bioturbators (e.g. deep-marine and/or transitional marine environments; Bayet-Goll et al. Reference Bayet-Goll, Daraei, Geyer, Bahrami and Bagheri2021 a). This restriction was further promoted during the Ordovician radiation (Sepkoski, Reference Sepkoski, Cooper, Droser and Finney1995; Droser & Finnegan, Reference Droser and Finnegan2003), known as the Great Ordovician Biodiversification Event (GOBE) (Servais & Harper, Reference Servais and Harper2018), when a considerable diversity increase occurred in deposit-, detritus- and suspension-feeders and grazers in shallow-water benthic communities (Servais et al. Reference Servais, Lehnert, Li, Mullins, Munnecke, Noutzel and Vecoli2008, Reference Servais, Owen, Harper, Kroger and Munnecke2010). For assessment of the influence of evolutionary radiations on matground facies, mapping the palaeoenvironmental distribution of trace fossils and MISS is essential, together with documenting the organism–matground interactions at the beginning of the Ordovician, when a wide variety of environmental conditions existed. It is likely that the factors controlling the spatiotemporal distribution of MISS around GOBE can be adapted to younger deposits as well when evolutionary radiations also impacted microbial mats distribution (e.g. Mata & Bottjer, Reference Mata and Bottjer2012).
The Shirgesht Formation, a Tremadocian to Floian shallow-marine succession in the Kalmard Block in Central Iran, is composed of offshore-to-foreshore shallowing-upward cycles (Figs 1 and 2) and contains diverse MISS and invertebrate trace fossils. High ichnodiversity across different sub-environments, from foreshore to lower-offshore / shelf settings, along an ancient wave-dominated depositional profile, makes the Shirgesht Formation ideal for studying the nature of bioturbation during the GOBE.Moreover, the distribution of the Shirgesht trace fossils and MISS can assist with understanding the effects of evolutionary radiations, particularly that of the Ordovician radiation, on the abundance and distribution of ‘physical-metazoan’ mixground and ‘physical-microbial’ matground ecosystems (Hagadorn & Bottjer Reference Hagadorn and Bottjer1999). An in-depth assessment of the trace fossils and MISS in the Shirgesht Formation may also clarify the link between early metazoans and palaeoenvironments as well as the shallow-marine ecological dynamics in the Early Ordovician. In a recent study, Bayet-Goll et al. (Reference Bayet-Goll, Buatois, Mángano and Daraei2021 b) showed the combined effect of environmental and evolutionary constraints on the migration of matground-dominated ecosystems in shallow- to deep-marine (turbidite fan) and estuarine settings of the Lower to Upper Ordovician Lashkerak and Ghelli formations (Alborz Mountains, northern Iran). That study discussed how matground-dominated ecosystems were distributed in submarine fans and related turbidite systems as well as estuarine environments, while the present study focuses on distribution of the same ecosystems along a wave-dominated depositional profile. Moreover, this study also addresses the environmental and stratigraphic distribution of shallow-marine ichnofabrics and MISS in the shoreface–offshore complex and barrier-island environments of the Ordovician Shirgesht Formation.

Figure 1. (a) Geological map of Iran with Central-East Iranian Microplate (CEIM) marked by a yellow box (modified from Aghanabati, Reference Aghanabati2004). (b) Stratigraphy of the lower Palaeozoic formations in the Alborz Basin and the Central-East Iran microplate.

Figure 2. (a) Geological map of the northern CEIM, with Tabas and Kalmard blocks marked in pink and the lower Palaeozoic marked in green; the yellow box indicates the study area. (b) Simplified map of the Kalmard region; the red stars are the locations of the study areas. (c, d) Panoramic views of the Kuh-e-Asheghan section, featuring the Precambrian Kalmard, Ordovician Shirgesht and Lower Carboniferous Gechal formations. (e) Panoramic views of the Kuh-e-Asheghan section, featuring the Kalmard and Shirgesht (members 1 to 3), Devonian Rahdar and Gechal formations.
2. Geological setting
After the Katangan orogeny in the late Precambrian, the cratonic area of the Iranian Plate and the Arabian Plate became a relatively stable continental platform that was dominated by epicontinental shelf deposition (Husseini, Reference Husseini1989; Geyer et al. Reference Geyer, Bayet-Goll, Wilmsen, Mahboubi and Moussavi-Harami2014). The Persian Cambro-Ordovician in the various basins of Iran attests to a passive margin tectonic setting (Geyer et al. Reference Geyer, Bayet-Goll, Wilmsen, Mahboubi and Moussavi-Harami2014), which apparently continued largely uninterrupted until the late Palaeozoic, with some orogenic uplifts across the Silurian–Devonian boundary (Berberian & King, Reference Berberian and King1981). The Cambro-Ordovician of northern, central and eastern Iran forms a more comprehensive succession than neighbouring Middle Eastern regions (Stöcklin et al. Reference Stöcklin, Ruttner and Nabavi1964), and thus is critical for the geological history of the region (Fig. 1a–b; Bayet-Goll et al. Reference Bayet-Goll, Geyer, Wilmsen, Mahboubi and Moussavi-Harami2014, Reference Bayet-Goll, Geyer and Daraei2018; Geyer et al. Reference Geyer, Bayet-Goll, Wilmsen, Mahboubi and Moussavi-Harami2014). The Ordovician rocks of Iran in most places consist of green shale, siltstone, and sandstone.
The Cambrian through Devonian strata represent Gondwana successions in the Kalmard Block of Central Iran (Ruttner et al. Reference Ruttner, Nabavi, Hajian, Bozorgnia, Eftekharnezhad, Emami, Flügel, Flügel, Haghipour, Iwao, Kahler, Ruttner-Kolisko, Sartener, Stepanov, Valeh, Walliser and Winsnes1968; Geyer et al. Reference Geyer, Bayet-Goll, Wilmsen, Mahboubi and Moussavi-Harami2014; Fig. 1c). A series of studies has been undertaken on the Shirgesht Formation, providing useful biostratigraphic information (Ruttner et al. Reference Ruttner, Nabavi, Hajian, Bozorgnia, Eftekharnezhad, Emami, Flügel, Flügel, Haghipour, Iwao, Kahler, Ruttner-Kolisko, Sartener, Stepanov, Valeh, Walliser and Winsnes1968; Hamedi et al. Reference Hamedi, Wright, Aldridge, Boucot, Bruton, Chaterton, Jones, Nicoll, Rickards and Ross1997; Aghanabati, Reference Aghanabati2004; Ghaderi et al. Reference Ghaderi, Aghanabati, Hamdi and Saeedi2009); based on the early referred papers, the age of the Shirgesht Formation has been set as the Early to Middle Ordovician (Fig. 2c–g).
3. Study area and methods
In this study, the Shirgesht Formation was examined in three sections (SHI-1 to SHI-3) (Fig. 2a–b), where stratigraphic columns were constructed based on sedimentological and ichnological data (Fig. 3).Textural and compositional properties were analysed in thin-section studies of 250 rock samples, using a polarizing (petrographic) microscope. This ichnological study is rooted in the concept of ichnofabric, which is defined as ‘all features of the texture and internal structure of the rock that result from bioturbation’ (Taylor & Goldring, Reference Taylor and Goldring1993). To this end, ichnofabric characterization was performed only in the field through investigating the most important ichnofabric criteria, including composition and distribution of trace fossils, intensity of bioturbation, abundance of individual ichnogenera and estimation of ichnodiversity, and the cross-cutting relationships between trace fossils, along with non-ichnological features such as primary sedimentary structures. Nature and cross-cutting relationships of trace fossils and primary sedimentary data were used to define the tiering pattern in different ichnofabrics. Regarding facies heterogeneity and stacking patterns of the facies, the distribution of MISS along a wave-dominated depositional profile was also documented.
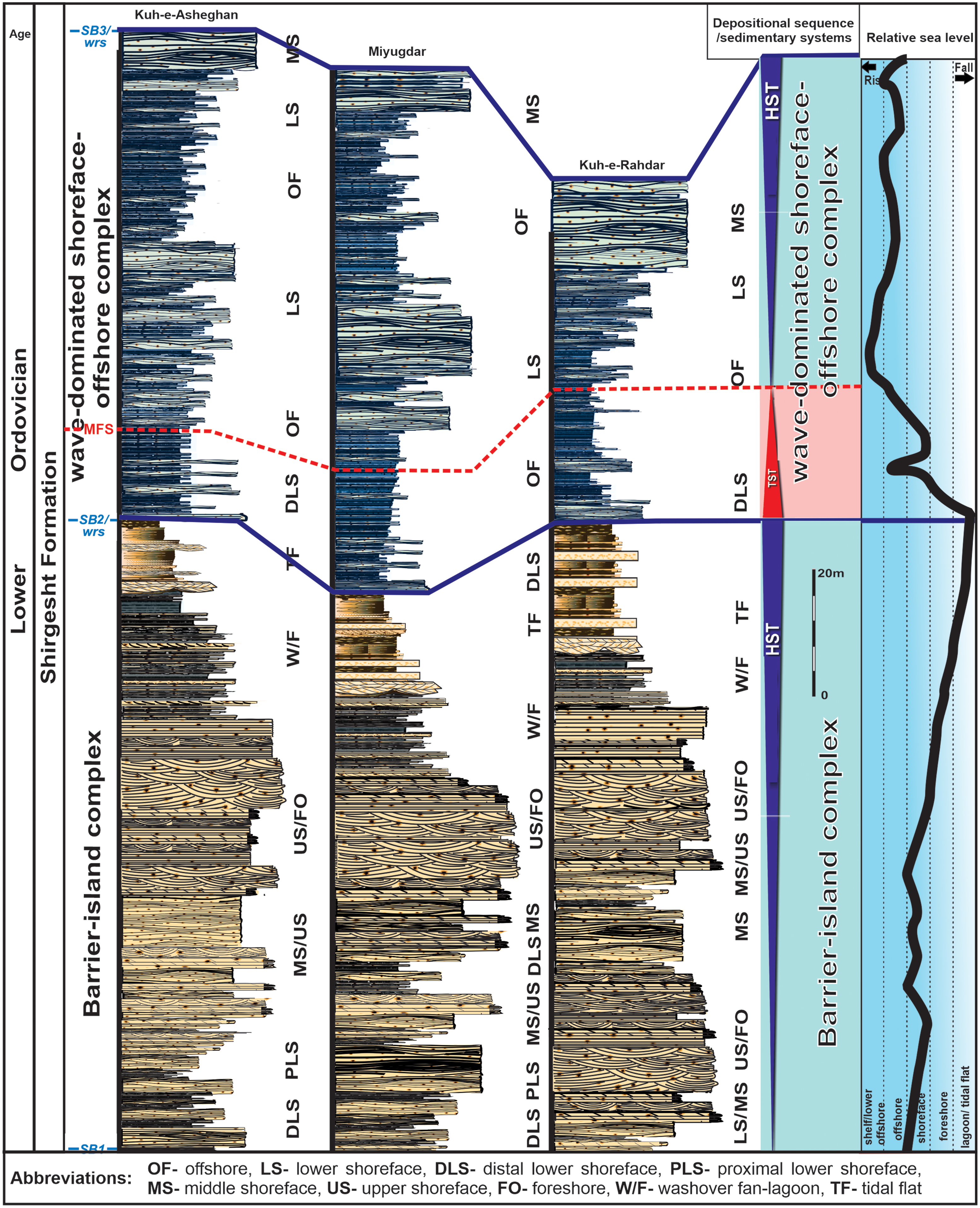
Figure 3. Composite log of the Shirgesht Formation in Central Iran, Kalmard Basin, with sedimentological and sequence stratigraphic characteristics.
4. Facies and depositional systems
A comprehensive litho-ichnofacies analysis of the Shirgesht Formation in the Kalmard Block of Central Iran was been recently carried out by Bayet-Goll et al. (Reference Bayet-Goll, Myrow, Aceñolaza, Moussavi-Harami and Mahboubi2016) and Bayet-Goll et al. (Reference Bayet-Goll, Uchman, Daraei and Neto de Carvalho2021 c). These studies show that the Shirgesht Formation formed a mixed carbonate–siliciclastic depositional system that consisted of a carbonate ramp (member 3), barrier-island (member 1) and shoreface–offshore (member 2) complexes (Fig. 4a–c). Stratigraphically, member 1 records a transition from offshore-transition to middle-shoreface, upper-shoreface, foreshore, tidal-inlet, interfingered washover-fan and lagoon, and intertidal-flat to supratidal settings, with a prograding architecture represented by a shallowing-upward facies pattern (Fig. 4b, e–g). After the onset of a global larger-scale lowering of sea level at the Cambrian–Ordovician boundary (Haq & Schutter, Reference Haq and Schutter2008) and during the Tremadocian, the wave-dominated shoreface (member 2) gave way to a barrier-island complex (member 1). High influx of siliciclastics associated with a slow and small-scale sea-level rise led to a gradual decrease in accommodation space and the development of a progradational facies architecture. Although most studies indicate that barrier-island complexes form during transgression (Cattaneo & Steel, Reference Cattaneo and Steel2003), the lack of evidence for backstepping of the facies belts and upward-decreasing thickness of sandstone layers (lagoon – tidal-flat facies), together with a progradational stacking pattern, collectively suggests the barrier-island complex of the Shirgesht Formation formed during a slow sea-level rise that was outpaced by the high rate of sedimentation (i.e. normal regression). In contrast, member 2 represents a retrogradational succession developed when the rapid eustatic sea-level rise during the Floian (Haq & Schutter, Reference Haq and Schutter2008) outpaced the local sediment supply. Member 2 shows retrogradational facies architecture from offshore-shelf and offshore-transition to distal lower-shoreface facies that in turn grades upwards into a progradational package that is characterized by a facies transition from offshore-transition – lower shoreface and proximal lower to middle shoreface facies (Fig. 4a, c). The thick progradational package formed due to the normal regression during sea-level highstand.

Figure 4. Field aspects of third-order depositional sequences of the Shirgesht Formation, Kalmard Basin. (a–c) Panoramic view of the barrier-island complex (member 1) and wave-dominated shoreface–offshore complex (Sequence 2). (d, e) Panoramic view of the Lower Member of the Shirgesht Formation representing the distal lower shoreface to upper shoreface/foreshore deposits. (f) Large-scale trough-cross stratified upper shoreface facies with tabular cross-stratification toward the top of the layer. (g) Interlaminated sandy coarse dolograinstone, silty dolomudstone and fine-grained sandstone, and dark mudstone deposited in intertidal to supratidal environments. Examples (a), (c) and (i) are from Kuh-e-Asheghan section, and examples (b), (f) and (g) are from Kuh-e-Rahdar section, whereas examples (h), (e) and (d) are from Miyugdar section.
The offshore facies is generally composed of variegated thinly interlaminated mudstone, shale and siltstone, sporadically interbedded with fine-grained sandstone. Lower parts of this facies are composed of mudstone-dominated heterolithic units and show an upward increase in the sandstone-dominated heterolithic units, which is interpreted as a progression from low-energy shelf / lower-offshore deposits below the storm wave base (SWB) to wave-agitated upper offshore deposits above the SWB. The sandstone beds are characterized by sharp, scoured bases, small-scale hummocky cross-stratification (HCS), parallel lamination, and/or symmetrical ripple marks at the top (Fig. 5a), which are hallmark features of event beds or tempestites (Myrow & Southard, Reference Myrow and Southard1996). The sandstones also tend to become thicker and more closely spaced up-section. Strongly fluctuating hydrodynamic conditions in the offshore setting were derived by the intermittent alternation of high-energy periodic storms and low-energy waning storm-energy conditions in a wave-dominated regime (Fig. 5b).

Figure 5. Field aspects of the Shirgesht Formation, Kalmard Block. (a (Heterolithic association of sharp-based sandstones/siltstones and shales in shelf / lower-offshore facies. (b (Heterolithic bedding in fine-grained sandstone, shale and muddy siltstone, with hummocky cross-stratification ripples, wave ripple cross-lamination and horizontal lamination, from upper offshore. (c (Offshore-transition – lower-shoreface facies with the upward decrease in bioturbation structures, diminishing and less abundant Trichophycus (CT ichnofabric), accompanied by the upward increase in abundance of event beds or tempestites. (d) The sandstone beds in lower and middle shoreface facies with HCS, horizontal lamination, and CT ichnofabric, and the upward increase in the thicknesses of sandstone layers. (e (A progradational package from offshore-transition – lower-shoreface and middle- to upper-shoreface facies. (f (Thick HCS bed in middle shoreface facies. (g (A shallowing-upward package from lower and middle shoreface grading upward to large-scale trough-cross stratified upper shoreface facies with tabular cross-stratification towards the top of the layer. (h (Tidal flat facies show a shallowing-upward trend beginning with lower-intertidal facies passing upward into upper-intertidal and supratidal facies. (i (Flaser-, wavy-, lenticular-bedded and thinly laminated heterolithic and rhythmical deposits in tidal flat facies. Examples (a), (c) and (i) are from Kuh-e-Asheghan section, and examples (b), (f) and (g) are from Kuh-e-Rahdar section, whereas examples (h), (e) and (d) are from Miyugdar section.
The offshore-transition to lower-shoreface facies consists of HCS in fine-grained sandstones that are sharp-based and interbedded with heavily bioturbated intervals mudstones (shale, siltstone, sandy siltstone) that decrease in thickness and abundance up-section (Fig. 5c–d). Deposition occurred below the fair-weather wave base (FWWB) and was interrupted by the deposition of thin, sharp-based sand beds with HCS and wave-ripple lamination, reflecting combined and oscillatory flows during storms (i.e. event beds or tempestites). The middle shoreface facies in this member is composed of thickly amalgamated sandstones with tabular to low-angle planar cross-stratification and HCS. Overall, the sedimentary structures show an upward decrease in oscillation ripple laminae and muddy intervals and a simultaneous increase in the thickness and abundance of planar lamination and HCS (Fig. 5e). Thick, amalgamated beds with HCS are interpreted as proximal storm deposits that formed in combined oscillatory and unidirectional currents (Fig. 5f). The scarcity of muddy interbeds between storm sandstone beds is consistent with deposition above the FWWB and continuous water agitation by waves. The upper shoreface/foreshore facies is composed of thick- to very thick-bedded, tabular and trough cross-bedded and sub-parallel-laminated sandstone beds that are capped by symmetrical to nearly symmetrical ripple marks (Fig. 5g). Amalgamation of sandstone beds and associated physical sedimentary structures suggests deposition under high-energy conditions as well as progressive sorting and sediment reworking by waves and currents in the upper shoreface/foreshore environment between the mean low-tide level and the FWWB. Large, high-angle cross-strata resulted from the migration of dunes under wave-generated onshore-directed sediment transport. The parallel lamination reflects high-energy swash and backswash transport, typical along the beach. The presence of oscillation ripple lamination and burrows at the top of these beds points to occasional brief drops in depositional energy levels (i.e. short periods of low wave energy).
The lagoonal facies is composed of interbedded fine-grained, well-sorted, medium-bedded sandstone, as well as dark mudstone, siltstone, and shale. The sandstone beds occur as both isolated beds and thick units of sandstone layers separated by very thin shale beds. Beds of this facies have basal erosional contacts and contain chert-pebble-bearing, massive to horizontal-laminated and low-angle cross-bedded sandstone beds that are locally covered by symmetrical ripple marks. Overall, the sedimentary structures suggest deposition in a wave-influenced lagoonal setting that was periodically influenced by storm-generated currents. Flooding events and periodic reworking by storms led to the transport of fine-grained sediments into a protected environment (e.g. lagoon) behind back-barrier islands to form washover fans. The texturally and structurally graded sandstone layers suggest rapid deposition in single events. The interbedding of mud- and sandstones also suggests deposition from suspension and gentle traction currents.
The intertidal-flat to supratidal facies shows a shallowing-upward trend that normally begins with massive dolomite and interlaminated sandy, coarsely crystalline dolograinstone, silty dolomudstone and laminated sandstone layers with planar to tabular cross-beds and flaser, lenticular and wavy to planar bedding, passing upward into dolomudstone (Fig. 5h–i). The latter are associated with exposure features, such as desiccation polygons or fenestral dolomite. Physical sedimentary structures include massive bedding, current-ripple cross-stratification and symmetrical ripple marks. Flaser- and wavy-bedded and thinly laminated heterolithic deposits indicate deposition on tidal flats. The mudstone-rich intervals, together with fenestral detrital dolomudstone, reflect deposition on a low-energy supratidal or mixed, upper-intertidal flat. The sandy coarse-crystalline dolograinstone beds and current-ripple-laminated sandstone layers with evidence of strong tidal currents suggest deposition on a higher-energy sandy, lower-intertidal flat.
5. Ichnofabrics
Ten ichnofabrics are recognized in both the barrier-island and shoreface–offshore complexes of the Shirgesht Formation. Some of the following ichnofabrics occur in various stratigraphic positions throughout the succession, whereas others only occur locally (e.g. Sk, CoSk, Th, CT ichnofabrics).
5.a. Skolithos ichnofabric (Sk)
The Sk ichnofabric occurs in clean sandy substrates of the upper shoreface/foreshore with trough and planar cross-stratified, fine- to medium-grained sandstones (Fig. 6a–b) and thick intervals of flaser-bedded dolomitic sandstone or planar to tabular cross-bedded beds in the intertidal flat facies (Fig. 6c–d). This ichnofabric shows highly variable bioturbation intensities (BI), ranging from BI 0 to 3. The Sk ichnofabric is characterized by its high abundance of primary sedimentary structures and a low-diversity ichnofauna with simple tiering pattern and colonization by single tracemakers. This ichnofabric is dominated by mid- to deep-tier, vertical dwelling burrows of suspension-feeders (Skolithos linearis) that are 5–10 mm wide and 20–120 mm long. A subordinate suite of middle to shallow tiers consisting of Diplocraterion, Arenicolites, Rosselia and Palaeophycus is also present. Evidence of intensive bioturbation (BI = 5–6), essentially resulting in a piperock ichnofabric, also occurs locally in both the upper shoreface and intertidal-flat facies.

Figure 6. Characteristics of the Sk and Pl ichnofabrics. (a, b) Piperock ichnofabric of vertical dwelling traces, upper shoreface facies. (c) CoSk ichnofabric associated with Sk ichnofabric in thick intervals of wavy to flaser-bedded dolomitic sandstone beds of intertidal flat facies, Conichnus (Co). (d) Sk ichnofabric in planar cross-bedded sandstone beds of the intertidal flat facies. (e) Planolites (arrows) and Bergaueria (Be) in lagoon facies with Pl ichnofabric. (f) Palaeophycus in tidal flat facies with Pl ichnofabric. (g) Synaeresis cracks in lagoon facies with Pl ichnofabric. Examples (a), (d) and (e) are from Kuh-e-Asheghan section, whereas examples (b), (c), (f) and (g) are from Kuh-e-Rahdar section.
5.b. Planolites ichnofabric (Pl)
The Pl ichnofabric occurs in silty and muddy substrates of the lagoon facies and wavy to lenticular-bedded, and thinly laminated heterolithic intervals of the tidal-flat facies. This ichnofabric is commonly associated with the crowded Rosselia (CR) and Sk ichnofabrics (see below and above, respectively). Sporadically distributed, highly burrowed, centimetre- to decimetre-thick units with CR and Sk occur throughout the mostly un-burrowed background of this ichnofabric. The Pl ichnofabric is characterized by single colonization, simple tiering pattern with low ichnodiversity, low bioturbation intensity (BI = 0–2), and dominated by simple feeding strategies of trophic generalists consistent with facies-crossing forms (MacEachern et al. Reference MacEachern, Bann, Bhattacharya and Howell2005). Its main constituent is represented by a few simple and unspecialized, shallow-tier, worm-like mobile deposit-feeding traces (Planolites and Palaeophycus) (Fig. 6e–f). A subordinate suite includes Diplichnites, Cruziana, Bergaueria and Helminthopsis. Cross-cutting relationships are not detected. Synaeresis cracks are preserved locally (Fig. 6g). MISS with high diversity and occurrence are widely present in host facies of this ichnofabric (see below).
5.c. Rosselia–Skolithos ichnofabric (RoSk)
The RoSk ichnofabric occurs in sandy substrates of the lower and middle shoreface facies, and shows shallow to deep tiers, reflecting multiple colonization events. The main constituent of this ichnofabric is middle- to lower-tier, funnel-shaped burrows of surface detritus-feeders (Rosselia and R. socialis) and deep-tier, suspension-feeding infaunal traces (Skolithos, Arenicolites) (Fig. 7a–b). A subordinate suite in Ro/Sk includes Bergaueria, Palaeophycus, Diplocraterion, Conichnus, Teichichnus, fugichnia and Monomorphichnus (Fig. 7c–d). The Rosselia-bearing bed-sets in the lower shoreface facies (BI = 1–3) consist of well-sorted, fine- to medium-grained, erosively based, HCS to massive sandstones intercalated with mudstone–siltstone beds. In longitudinal sections, Rosselia is characterized by straight to slightly curved, concentrically (sand- or mud-filled) lined cylinders that are 10–30 mm wide and 30–70 mm long. The upper bedding surfaces of sandstones show circular to elliptical endichnial burrows with a sand-filled tube, engulfed by concentric muddy laminae.

Figure 7. Characteristics of the RoSk and CRI ichnofabrics. (a, b) Funnel-shaped burrows of surface detritus-feeders (Rosselia; Ro) and suspension-feeding infaunal traces (Skolithos; Sk) and fugichnia (Fu), middle shoreface facies. (c) Diplocraterion, Palaeophycus (white arrows) and Skolithos (yellow arrows) in middle to upper shoreface facies. (d) Diplocraterion in middle shoreface facies. (e) Sandstone beds dominated by vertical dwelling burrows of Rosselia (CR ichnofabric), washover fan sandstone beds. (f) Top view of CR ichnofabric at the upper bedding surface showing a high concentration of circular, endichnial burrows of Rosselia. Examples (a), (b) and (c) are from Kuh-e-Asheghan section, and examples (d) and (f) are from Kuh-e-Rahdar section, whereas example (e) is from Miyugdar section.
5.d. Crowded Rosselia ichnofabric (CR)
The CR ichnofabric (sensu Nara, Reference Nara2002; Netto et al. Reference Netto, Tognoli, Assine and Nara2014) occurs in the washover fan sandstone beds within the fine-grained lagoonal deposits. CR is characterized by single colonization, simple tiering pattern with low ichnodiversity, high bioturbation intensity (BI = 4–5) and dense populations of funnel-shaped burrows of the surface detritus (Fig. 7e–f). A subordinate suite including sparse, deep to shallow tiers of Skolithos, Arenicolites, Palaeophycus and Bergaueria also occurs. In longitudinal sections, Rosselia is composed of vertical to sub-vertical, funnel-shaped burrows (maximum length 80 mm) with a sand-filled tube encircled by concentric muddy laminae. Compared with the RoSk ichnofabric, this washover fan facies with thick-bedded sandstone layers (in a position close to the barrier) is characterized by a considerable increase in the abundance of Rosselia to more than 30 burrows per m2, defining the CR ichnofabric (Nara, Reference Nara2002). However, in a position further from the barrier and close to the tidal flat, CR ichnofabric is replaced by microbial matgrounds (see below). Cross-cutting relationships or tiering levels are not observed in CR ichnofabric.
5.e. Conichnus–Skolithos ichnofabric (CoSk)
The CoSk ichnofabric is associated with the same facies as the Sk ichnofabric, but commonly occurs in planar-laminated to small cross-bedded, fine- to medium-grained dolomitic sandstones of the upper shoreface to foreshore facies and thick intervals of wavy to flaser-bedded dolomitic sandstone beds of the intertidal-flat facies. This ichnofabric contains shallow to deep tiers, reflecting multiple, successive-tier colonization events, and is characterized by low ichnodiversity, high bioturbation intensity (BI = 3–5) and dense populations of deep-tier, suspension-feeding infaunal traces (Fig. 8a–d). Intensive bioturbation (BI = 5–6), forming a piperock ichnofabric of vertical dwelling burrows, also occurs locally in the CoSk ichnofabric (Fig. 8c–d), which consists of deep-tier, conical to subconical vertical burrows with nested internal laminae (Conichnus); vertical or steeply inclined, cylindrical to sub-cylindrical burrows (Skolithos); and escape structures (fugichnia). The subordinate suite includes sparse, mid- to shallow-tier, Palaeophycus, Bergaueria and Rosselia. The depth of infaunal tiering or penetration depth of Conichnus (40–100 mm long) in CoSk is greater than that of Skolithos (20–70 mm long).

Figure 8. Characteristics of the CoSk and Th ichnofabrics. (a, b) Funnel-shaped burrows of Conichnus with nested internal laminae in wavy to flaser-bedded dolomitic sandstone beds of intertidal flat facies. (c) CoSk ichnofabric associated with Sk ichnofabric in wavy to flaser-bedded dolomitic sandstone beds of intertidal flat facies. (d) A high concentration of conical burrows of Conichnus, intertidal flat facies. (e, f) Branched networks of large Thalassinoides; branches are Y-shaped, commonly enlarged at the bifurcation point, middle to lower shoreface facies. (g) Bedding-plane view of fine-grained sandstone showing well-developed Thalassinoides burrows, in offshore-transition facies. (h) Firmground Glossifungites ichnofacies, the lower surfaces of beds characterized by pervasive branching burrow networks of decapod crustacean burrows, attributed to the substrate-controlled Glossifungites ichnofacies, in the upper offshore facies. (i) Thalassinoides burrows in offshore-transition facies with vertical shafts, sharp boundaries and filled with coarser sediments. Examples (a), (e), (f) and (g) are from Kuh-e-Asheghan section, and examples (b), (c), (d) and (i) are from Kuh-e-Rahdar section, whereas example (h) is from Miyugdar section.
5.f. Thalassinoides ichnofabric (Th)
The Th ichnofabric occurs at the base of storm-generated sandstone layers in the lower- and middle-shoreface, offshore-transition and upper-offshore facies. Commonly associated with the CrSkRo and Ro/Sk, the Th ichnofabric displays multiple, successive-tier colonization events (BI = 3–4) and is characterized by a moderate ichnodiversity, a large size of the burrows and a moderate bioturbation index (BI = 2–4). The main constituents of this ichnofabric are shallow- to mid-tier, endichnial or hypichnial, full-relief, smooth-walled burrow systems of decapod crustaceans (Thalassinoides) and shallow-tier, passive carnivore traces (Palaeophycus; Fig. 8e–h). The subordinate components are Planolites, Cruziana, Monomorphichnus, Diplichnites, Helminthopsis, Arenicolites and Skolithos. Thalassinoides is composed of three-dimensional burrow systems with predominantly horizontal galleries and vertical shafts. Horizons of branched networks of Thalassinoides in the lower to middle shoreface facies with unlined burrows and distinct burrow boundaries are 20–50 cm thick, and branch length ranges from 50 to 100 cm and diameter from 5 to 10 cm (Fig. 8g–h). The burrow fills may be similar to or different from the ambient rock in their petrological composition. Occasionally, the fill of Thalassinoides in the offshore-transition facies is dominated by coarse sand due to the passive infilling of the open burrow system (Fig. 8i). In these facies, branching burrow networks of decapod crustaceans are characterized by unlined, sharply bound shafts.
5.g. Cruziana–Skolithos–Rosselia ichnofabric (CrSkRo)
The host rocks of CrSkRo ichnofabric are mainly mud-dominated beds of the upper offshore, offshore-transition to distal lower shoreface facies that alternate with storm-generated sandstone beds (event beds or tempestites). Toward the shallower shoreface areas, with an increasing frequency of tempestites (lower and middle shoreface), CrSkRo is replaced by RoSk and Th ichnofabrics. Bioturbation is characterized by highly variable intensities, varying from BI 2 (event beds) to BI 6 (finer-grained interbeds). CrSkRo is characterized by multiple colonization events and complex tiering patterns by shallow- to deep-tier trace fossils. The suite in CrSkRo ichnofabric is dominated by a mixture of varied behavioural strategies. The mud-dominated beds contain locomotion-foraging trails (Cruziana furcifera, C. goldfussi, C. rugosa, C. yini), resting (Rusophycus, Bergaueria, Lockeia), walking (Diplichnites), sideways-grazing (Dimorphichnus) and swimming-grazing (Monomorphichnus), grazing (Helminthopsis), deposit-feeding (Planolites, Arthrophycus) and dwelling/deposit-feeding to passive carnivore traces (Trichophycus, Thalassinoides) (Fig. 9a–g). The storm-generated sandstone beds are dominated by suspension-feeding (Diplocraterion, Arenicolites, Skolithos) (Fig. 9a, f–h), surface detritus-feeding (Rosselia), passive carnivore (Palaeophycus) and escape traces. On a proximal-to-distal transect, with an increase in the number of storm sandstone beds showing large-scale HCS and internal erosion surfaces, the abundance of traces related to r-selected opportunistic species increases in these unstable substrates. Moreover, with a decrease of the mud-dominated beds, a considerable decrease in the abundance of arthropod burrows and trackways is observed.

Figure 9. Characteristics of the CrSkRo and CT ichnofabrics. (a, b) Cruziana furcifera, Rusophycus (Ru) and Bergaueria (Be), with small vertical burrows (Skolithos; Sk) and Arenicolites (Ar), offshore-transition facies. (c) Dimorphichnus, parallel series of sigmoidal or comma-shaped impressions arranged obliquely to the direction of movement, offshore-transition facies. (d) Lower bedding plane of a sandstone with a dense population of C. goldfussia, offshore-transition facies. (e) Arthophycus brongniartii with angular segments along the burrow, upper offshore facies. (f, g) Cross-cutting relationship of Trichophycus venosus with the background suite: Trichophycus (Tr) cross-cuts Cruziana isp. (Cr) and both of them cross-cut by deep-tier vertical suspension-feeding burrows, Arenicolites (Ar), Diplocraterion (Di) and Skolithos (Sk), offshore-transition to upper offshore facies. (h) Hypichnial view of Diplocraterion, offshore-transition facies. (i–j). Crowded Trichophycus (CT) ichnofabric with well-developed Trichophycus venosus in mudstone beds attached to the overlying sandstone tempestite, upper offshore facies. Examples (a), (b), (c), (g) and (j) are from Kuh-e-Asheghan section, whereas examples (d), (e), (f), (h), (i) and (j) are from Kuh-e-Rahdar section.
5.h. Crowded Trichophycus ichnofabric (CT)
The crowded Trichophycus (CT) ichnofabric (Bayet-Goll et al. Reference Bayet-Goll, Uchman, Daraei and Neto de Carvalho2021 c) occurs in the mudstone waning-storm beds in the lower-shoreface, offshore-transition and upper-offshore facies, and is commonly associated with the CrSkRo, CrHe and RoSk ichnofabrics. This ichnofabric is characterized by multiple, successive-tier colonization events with a high ichnodiversity and bioturbation intensity (BI = 4–5), and dense populations of large, mid-tier, straight to slightly curved burrows of Trichophycus venosus (Figs 5c–d, 9i–j). This trace shows variations in preservation from isolated forms in mudstone-dominated heterolithic facies of the lower offshore (endichnia) to unilobed casts in rhythmically interstratified bioturbated muddy beds of the upper offshore as well as HCS, hypichnic, sharp-based sandstones. The most common preservation form is horizontal to oblique, shallow, flattened burrows that mostly contain a retrusive spreite as gutter-shaped laminae (teichichnoid internal structure). Ranging between 10 and 30 mm, the diameter in individual burrows is usually constant, but the length is variable from 50 to 200 mm. The maximum penetration depth (without decompaction) in the isolated forms exceeds 10 cm below the colonization surface. Toward the shallower shoreface areas, with an increase in bed thickness and tempestite frequency, the length, diameter and density of burrows along with the penetration depth tend to decrease. In these areas, the muddy intervals with CT ichnofabrics largely occur within the upper part of the storm beds. Subordinate components are Cruziana, Rusophycus, Planolites, Monomorphichnus, Diplichnites, Bergaueria, Palaeophycus, Arenicolites and Skolithos (Fig. 9f–h). In many cases, T. venosus, as an elite burrow (sensu Bromley, Reference Bromley1996), cross-cuts arthropod burrows and trackways, leading to the obliteration of some shallow-tier structures. Toward the shallower areas, an overall decrease in the density of Trichophycus results in a better preservation of the shallow-tier background suites.
5.i. Cruziana–Helminthopsis ichnofabric (CrHe)
The CrHe ichnofabric is common in the rhythmically interstratified bioturbated muddy beds and thinly bedded, sharp-based sandstones of the upper offshore successions associated with the CT ichnofabric. Typically located between the CrSkRo and ChPl ichnofabrics, this ichnofabric shows shallow to mid-tiers, reflecting multiple colonization events represented by a high degree of bioturbation (BI = 3–5, averaging 4), a high ichnodiversity and high density of locomotion-foraging, walking, sideways-grazing and swimming-grazing, resting and deposit-feeding traces, and sparse feeding and grazing traces. The main constituent of CrHe ichnofabric in mudstone-dominated heterolithic beds is shallow- to middle-tier arthropod burrows and trackways (Cruziana furcifera, C. goldfussi, C. rugosa, Trichophycus venosus, Rusophycus, Monomorphichnus, Diplichnites, Dimorphichnus, Didymaulichnus) (Fig. 10a–f). Shallow-tier, epichnial or endichnial, shallow, endobenthic burrows of deposit-feeders (Planolites, Arthrophycus) are also dominant in this ichnofabric. The subordinate suite shows sparse, shallow- to deep-tier Bergaueria, Palaeophycus, Diplocraterion, Skolithos, Chondrites, Thalassinoides, Olivellites? and Helminthopsis (Fig. 10f–g). In this ichnofabric, microbial mats as restricted patches in the vicinity of bioturbated mixground are sporadically present on the upper bedding surfaces of fine-grained sandstone beds (see below).

Figure 10. Characteristics of the CrHe and ChPl ichnofabrics. (a) Cruziana goldfussi in upper offshore facies. (b) C. furcifera in upper offshore facies. (c) Diplichnites gouldi preserved as a concave epirelief in upper offshore facies. (d) Rusophycus in upper offshore facies. (e) Palaeophycus (Pl), Olivellites? (Ps) and Arthophycus (Ar) in upper offshore facies. (f) Lower bedding plane of a sandstone with a dense population of Rusophycus, upper offshore facies. (g) Hypichnial view of Diplocraterion, upper offshore facies. (h) Gordia (arrows) in the lower-offshore facies (ChPl ichnofabric). (i) Chondrites in lower-offshore facies, Miyugdar section. (j) Helminthopsis in lower-offshore facies. (k) Planolites montanus in the lower-offshore facies. Examples (b), (c), (d), (e), (h) and (k) are from Kuh-e-Asheghan section, whereas examples (a), (g), (i), and (j) are from Kuh-e-Rahdar section.
5.j. Chondrites–Planolites ichnofabric (ChPl)
The ChPl ichnofabric is found in heterolithic mudstone-dominated units of the lower-offshore facies below the SWB, and is represented by a nil to sparse bioturbation (BI = 0–2) in low-diversity ichnoassemblages. The tiering structure shows a single colonization and simple tiering pattern characterized by the occurrence of small, deep-tier endichnial deposit-feeder burrows (Chondrites and Zoophycos) and shallow-tier, vermiform deposit-feeder burrows (Planolites, P. montanus) (Fig. 10h–k). Shallow-tier, endichnial or hypichnial, surface and near-surface grazing trails (Helminthopsis, Gordia) are also dominant in this ichnofabric (Fig. 10h, j). The subordinate suite shows sparse, shallow-tier Bergaueria and Cruziana. In this ichnofabric, patches of microbial mats are common on the upper bedding surfaces of fine-grained sandstone beds (see below).
6. Distribution of microbially induced sedimentary structures (MISS)
As mentioned before, the progradationally stacked barrier–lagoonal facies of member 1 (lower Shirgesht Formation) grades upward to the transgressive, siliciclastic, open-shelf deposits of member 2. This transition represents the establishment of a shallow-marine, low-gradient, storm-dominated shelf. MISS occur commonly and in high diversity in this barrier-island complex, mainly in the tidal flat and back-barrier (lagoon and washover fan) facies. In the shoreface–offshore complex, MISS are dominant in the lower-offshore to shelf facies and rare in the lower parts of the upper offshore. Here, they occur as restricted patches in the vicinity of bioturbated mixgrounds. However, in both environments, the facies of the foreshore, tidal inlet, lower to upper shoreface, and offshore transition to upper offshore lack MISS. In the tidal flat and back-barrier facies of the barrier-island complex, features related to mat desiccation and shrinkage, and early microbial colonization such as shrinkage cracks, petee ridges, gas domes, radial gas- or fluid-escape structures (pseudofossil ‘Astropolithon’), Manchuriophycus-like shrinkage cracks or trough-confined cracks, palimpsest or mat-protected, mat-smoothed or multidirectional ripple marks, wrinkle structures and erosional pockets are observed. These MISS in the tidal-flat and back-barrier facies are commonly associated with the Sk, Pl, CR and CoSk ichnofabrics (but not on the same bedding surfaces). In other words, these facies are generally dominated by Sk, CR and CoSk ichnofabrics, sporadically interbedded with sandstone beds that contain microbial matgrounds.
Sand cracks and petee ridges, millimetric to centimetric in width, are developed preferentially on top of mud-free sandstone beds and are often developed in troughs or on crests of ripple marks (Fig. 11a–g). Shapes of these structures in plan view are variable including curved, rectangular, irregular polygonal, circular to semicircular, and sinusoidal to fusiforms. In the tidal-flat facies, there is an association between grazing traces and arthropod trackways (Planolites, Cruziana, Diplichnites), and sand cracks and petee ridges (Fig. 11c–d). The cracks are filled with structureless quartz sand, identical to the host rock. Petee ridges share the same geometrical variations of the sand cracks, but commonly occur more widely in the host facies (Fig. 11c–d, g–h). In different geometries of these structures, the width and height of the ridges are different, which is the result of variations in mat thickness (Bose & Chafetz Reference Bose and Chafetz2009). Sand cracks reflect the episodic alternation between mat growth and dehydration or desiccation of microbial mats (e.g. Gerdes et al. Reference Gerdes, Klenke and Noffke2000; Bose & Chafetz, Reference Bose and Chafetz2009). Moreover, the formation of petee ridges has been attributed to ‘buckling, doming and rupturing processes of microbial mats’ (e.g. Gerdes et al. Reference Gerdes, Klenke and Noffke2000; Schieber et al. Reference Schieber, Bose, Eriksson, Sarkar, Schieber, Bose, Eriksson, Banerjee, Sarkar, Altermann and Catuneanu2007). Gas domes (or dome-like elevations) and fluid-escape structures (‘Astropolithon’) are characterized by epichnial mounds with conical dome-like elevations on the upper surfaces of sandstone beds, bearing small, central depressions (Fig. 11h–k). The diameter of the domes is from 7 to 15 mm. Gas domes form preferentially due to gas accumulation in the sediment and beneath the coherent microbial mats (Noffke et al. Reference Noffke, Gerdes, Klenke and Krumbein2001). Manchuriophycus-like shrinkage cracks (3–10 mm wide) are characterized by sinuous, circular and triradiate cracks on upper bedding surfaces of sandstones in troughs of symmetrical to nearly symmetrical ripple marks (Fig. 12a–b). In some cases, these sinuous cracks are also developed on ripple crests atop rippled sandstones, roughly following ripple marks with straight or slightly sinuous crests (Fig. 12c–d). These structures are formed due to dehydration and shrinkage processes, and the close affinity of these structures with wave-ripple marks suggests a subaqueous origin (Schieber et al. Reference Schieber, Bose, Eriksson, Sarkar, Schieber, Bose, Eriksson, Banerjee, Sarkar, Altermann and Catuneanu2007; Noffke, Reference Noffke2010). On many bedding surfaces in the tidal-flat and back-barrier facies, ripple marks with different orientations are present in discrete patches (Fig. 12e–g). A periodic alternation between microbial growth, biostabilization of sediments and physical reworking by currents in the host sediment is a prerequisite for the formation of these structures (Pflüger, Reference Pflüger1999; Bottjer & Hagadorn, Reference Bottjer, Hagadorn, Schieber, Bose, Eriksson, Banerjee, Altermann and Catuneanu2007; Noffke, Reference Noffke2010). In some cases, all or parts of the ripple marks are covered by coherent microbial mats (Fig. 12g–h). Rarely, the coverage of ripple marks by microbial mats is so high that it masks the original ripple morphology (Fig. 12i–j).

Figure 11. Distribution of MISS along a wave-dominated depositional profile of the Shirgesht Formation. (a) Spindle-shaped sand cracks and triple junctions of cracks. (b) Reticulate, polygonal pattern of petee ridges. (c, d) Cross-cutting relationships between Cruziana-type scratches and reticulate pattern of petee ridges. (e) Sub-circular and circular microbial shrinkage cracks. (f) Curved, polygonal sand cracks developed in ripple-mark troughs and crests. (g). Palimpsest ripple marks on a bedding surface of sandstone with reticulate pattern of petee ridges. (h) Reticulate, polygonal pattern of petee ridges with probable gas dome (arrow). (i) Abundant small, probable gas domes. (j, k) Radial gas- or fluid-escape structures (= pseudofossil Astropolithon). Examples (c), (e), (h), (i), (j) and (k) are from Kuh-e-Asheghan section, and examples (a), (d) and (f) are from Kuh-e-Rahdar section, whereas examples (b) and (g) are from Miyugdar section.

Figure 12. Distribution of MISS along a wave-dominated depositional profile of the Shirgesht Formation. (a, b) Sinuous cracks confined to troughs of wave ripples (‘Manchuriophycus’) on the upper bedding plane of a sandstone bed. (c, d) Sinuous cracks that roughly follow the slightly sinuous- to straight-crested ripple marks. (e, f). Ripple marks with different orientations in discrete patches. (g) Multidirectional ripple marks associated with surface detritus-feeders burrows of Rosselia (Ro). (h) Large mat-smoothed ripple marks on bedding plane. (i, j). Ripple marks with dense microbial-mat covers masking the original shape of the ripple marks. Examples (a), (c), (d), (f) and (i) are from Kuh-e-Asheghan section, and examples (b), (e), (g) and (j) are from Kuh-e-Rahdar section, whereas example (h) is from Miyugdar section.
In the shoreface–offshore complex, wrinkle structures and circular to elliptical erosion marks (erosional remnants of mats) are seen in the lower-offshore to shelf facies and rarely in the upper offshore facies associated with the ChPl and CrHe ichnofabrics. The wrinkle structures range in preservation from (i) parallel, contorted and irregular forms to (ii) honeycomb-shaped networks with linear to winding ridges and (iii) round or flat-topped crests and pits separated by interconnected oblique, shallow, flat areas (Fig. 13a–d). The dimension of the networks varies between 4 mm, where finely wrinkled, to 12 mm, where coarsely wrinkled. These features may co-occur with shallow-tier, epichnial and endichnial trace fossils. In this regard, in these facies, there is an association between wrinkle structures and grazing traces and arthropod trackways (Planolites, Helminthopsis, Diplichnites, Rusophycus) (Fig. 13e–g). Toward the shallower areas of the upper-intertidal flat of the barrier-island complex, wrinkle structures and erosional remnants of mats co-occur with sand cracks (Fig. 13h). In most cases, these structures are associated with circular to elliptical erosional remnants. Various mechanisms have been suggested for the development and maintenance of these structures, including: (1) ductile deformation processes during the burial of microbial mats (Noffke et al. Reference Noffke, Gerdes and Klenke2003; Bottjer & Hagadorn, Reference Bottjer, Hagadorn, Schieber, Bose, Eriksson, Banerjee, Altermann and Catuneanu2007), (2) gas build-up or trapping of the gas accumulated underneath the mat (Porada & Bouougri, Reference Porada and Bouougri2007) and (3) the movement of microbial aggregates along a microbial-bound substrate by loose sand and waves (currents) with small-bed orbital amplitudes (Mariotti et al. Reference Mariotti, Pruss, Perron and Bosak2014).

Figure 13. Distribution of wrinkle structures and circular to elliptical erosional remnants in lower-offshore and tidal-flat – lagoon facies. (a) Wrinkle structures with short, flat-topped, winding crests, surrounding erosional remnants. (b) Honeycomb-shaped networks with linear to winding ridges. (c) Scattered patches of wrinkle structures (honeycomb-shaped networks) with circular to elliptical erosional remnants in lower offshore. (d) Wrinkle structures with parallel to irregular and linear ridges; flat-topped crests; lower offshore. (e) Cross-cutting relationship between patchy microbial mats (wrinkle structures) and Rusophycus (arrow). (f, g). Cross-cutting relationship between scattered patches of wrinkle structures and Planolites (Pl). (h) Scattered patches of wrinkle structures surrounding erosional remnants in intertidal flat facies co-occur with sand cracks. Examples (a), (c) and (f) are from Kuh-e-Asheghan section, and examples (b), (d), (g) and (h) are from Kuh-e-Rahdar section, whereas example (e) is from Miyugdar section.
Microscopic analysis of both facies reveals an alternation between sandy quartz layers and dark, wavy–crinkly laminae-bearing filamentous microstructures. Moreover, laterally continuous, wavy–crinkly dark laminae with a thickness of less than 0.5 mm are also present and in most cases alternate with light-grey, silty to very fine-grained sandy interlayers (Fig. 14a–d). These mat laminae contain a clayey matrix in which fine-grained quartz and muscovite grains are abundant but isolated (i.e. matrix-supported fabric; Fig. 14e–f). Oriented grains show strongly bimodal trends that are parallel to the bedding surface. The presence of floating detritus within the wavy, dark laminae indicates grain-trapping and binding of siliciclastic sediment by a benthic microbial mat (Gerdes et al. Reference Gerdes, Klenke and Noffke2000; Banerjee & Jeevankumar, Reference Banerjee and Jeevankumar2005). In addition, a preferred concentration of organic filaments and their diagenetic products (iron oxides and pyrite) along the wavy–crinkly dark laminae is also evident in thin-sections (Fig. 14g–h). Precipitation of diagenetic products was likely enhanced by the anaerobic decay of the buried mat material (cf. Schieber et al. Reference Schieber, Bose, Eriksson, Sarkar, Schieber, Bose, Eriksson, Banerjee, Sarkar, Altermann and Catuneanu2007).

Figure 14. Photomicrographs of microbial mats. (a–c) Alternation of sandy quartz and dark, wavy laminae; thin-section perpendicular to the bedding plane showing fine-grained sandstone and wavy crinkled laminae of microbial mats; ppl. (d, e) Alternation between sandy quartz layer and dark, wavy laminae-bearing filamentous microstructures; mat laminae characterized by abundant clay matrix, and floating grains of fine, isolated quartz (d) and mica (e). (f) Floating isolated quartz grains within the mat layers oriented parallel to the bedding plane. (g, h) Alternation of quartz-sands layer and dark, wavy laminae-bearing filamentous microstructures and its diagenetic products, pyrite (white arrows) and iron oxides (yellow arrow). Examples (a), (f) and (g) are from Kuh-e-Asheghan section, and examples (b), (d) and (h) are from Kuh-e-Rahdar section, whereas examples (c) and (e) are from Miyugdar section.
7. Discussion
7.a. Environmental and biogenic constraints on MISS distribution
The distribution of MISS in the sedimentary record is related to environmental factors (e.g. wave and current energy, substrate consistency, availability of nutrients, temperature, oxygenation, salinity and sedimentation rate) rather than the presence of a given sedimentary or ichnofacies (Schieber et al. Reference Schieber, Bose, Eriksson, Sarkar, Schieber, Bose, Eriksson, Banerjee, Sarkar, Altermann and Catuneanu2007; Bayet-Goll & Daraei Reference Bayet-Goll and Daraei2020). Moreover, Bayet-Goll & Daraei (Reference Bayet-Goll and Daraei2020) showed that environmental parameters can lead to changes in the type of mat-building benthic cyanobacteria and their role in depositional dynamics of transitional/shallow-marine depositional systems. Studies considering appropriate environments for the development of MISS (e.g. Noffke et al. Reference Noffke, Gerdes, Klenke and Krumbein2001, Reference Noffke, Gerdes and Klenke2003; Mata & Bottjer, Reference Mata and Bottjer2009, Reference Mata and Bottjer2012; Noffke, Reference Noffke2010; Mariotti et al. Reference Mariotti, Pruss, Perron and Bosak2014; Sarkar et al. Reference Sarkar, Banerjee, Samanta, Chakraborty, Mukhopadhyay and Singh2014; Davies et al. Reference Davies, Liu, Gibling and Miller2016; Bayet-Goll et al. Reference Bayet-Goll, Daraei, Geyer, Bahrami and Bagheri2021 a, b) show that environments ideal for extensive microbial mat formation are those where sedimentation rates are low and hydrodynamic energy is low to medium. However, within any environment with these overall characteristics, there are considerable variations with respect to hydrodynamic energy, and thus alternating domination of physical and biogenic processes is expected. For example, in tide- or wave-influenced environments, conditions ideal for MISS development are ample between the FWWB and SWB, because here the norm is variable wave energy coupled with episodic clastic sedimentation patterns (Eriksson et al. Reference Eriksson, Rautenbach, Wright, Bumby, Catuneanu, Mostert and van der Neut2009; Mata & Bottjer, Reference Mata and Bottjer2009, Reference Mata and Bottjer2012). Upon compiling the environmental distribution of MISS from 100 different Phanerozoic and 15 recent examples, Davies et al. (Reference Davies, Liu, Gibling and Miller2016, their table 1) demonstrated that most fossilized microbial mats were documented from storm- and tide-influenced environments, with only a few cases from deep-marine and fluvial settings. Compared to the shoreface–offshore complex, substrate stabilization by microbial activity in the barrier-island complex is more developed. The difference in the distributional patterns of MISS in the Shirgesht Formation is mainly related to differences in the biological parameters, such as increase in infaunal communities, and the environmental parameters, as well as the palaeo-ecological conditions through which they were formed (Figs 15 and 16).

Figure 15. Palaeoenvironmental model of the Shirgesht Formation, showing onshore–offshore ichnofabrics (modified from Bayet-Goll et al. Reference Bayet-Goll, Uchman, Daraei and Neto de Carvalho2021 c) and along-strike distribution of mixground and matground ecosystems.

Figure 16. Sedimentological model of the Shirgesht Formation, showing onshore–offshore gradients and the spatial distribution of facies in the barrier-island (a) and shoreface–offshore (b) complexes. The tidal flat, back-barrier (lagoon/washover fan) and lower-offshore setting were the optimum sub-environments for the development of MISS. FWWB = fair-weather wave base; SWB = storm wave base.
The disruption of mat fabrics with the tremendous increase in vertical partitioning of the infaunal and epifaunal communities in shallow-marine habitats, following the early Palaeozoic evolutionary radiations (e.g. Hagadorn & Bottjer, Reference Hagadorn and Bottjer1999; Buatois & Mangano, 2011a, b), led to the restriction of Shirgesht matgrounds to bioturbator-free environments such as the lower-offshore to shelf or tidal-flat and back-barrier environments, i.e. to environments inhospitable for bioturbators (see below). Although decline in bioturbation (resulting in depauperate ichnofaunal suites) likely increases microbial growth, fluctuating physico-chemical conditions are also critical for thriving microbial communities. The broad range of ichnofabrics indicates variable ecological conditions that also originated from fluctuating environmental conditions in these habitats (Fig. 15). It is noteworthy that both bioturbators and mat-forming microbes in these habitats are sensitive to physico-chemical changes, but the tracemakers have a greater ability to adapt by modifying their behaviour than the more conservative mat-forming microbes. In other words, mat-building microorganisms are limited to specific habitats and have less ability to adapt to changing environments.
7.a.1. Tidal-flat and back-barrier settings
In inhospitable environments such as tidal-flat and back-barrier facies of the barrier-island complex, the restriction of sediment mixing and bioirrigation by infaunal communities to shallow-tier, worm-like mobile deposit-feeding traces (Pl ichnofabric) led to a decrease in or even the removal of limitations on microbial growth. In these areas, the existence of environmental fluctuation in the form of physico-chemical stress on burrowing fauna (e.g. changes in salinity, nutrients, oxygen, clastic influx) led to a decline in tracemaker activity (i.e. rate of bioturbation). The largely un-burrowed nature of host facies with only Pl ichnofabric, the sporadic distribution of bioturbation structures, and an overall low ichnodiversity with a high abundance of simple and unspecialized, shallow-tier structures collectively suggest the inhabitation of tracemakers in a highly stressed depositional environment, possibly caused primarily by frequent salinity changes. In general, fluctuations in salinity, nutrients and oxygen evidently led to the decline in bioturbation and the development of depauperate ichnofaunal suites (Pl ichnofabric) in the lagoon (Bayet-Goll et al. Reference Bayet-Goll, Myrow, Aceñolaza, Moussavi-Harami and Mahboubi2016). This condition resulted in an ideal substrate, particularly at the interface of sandy and muddy layers, for microbial communities to blossom (Fig. 16). A few studies have reported the occurrence of matground facies in modern and ancient siliciclastic lagoonal settings (Horodyski et al. Reference Horodyski, Bloeser and Vonder Haar1977; Noffke, Reference Noffke2000; Mata & Bottjer, Reference Mata and Bottjer2012; Cuadrado et al. Reference Cuadrado, Perillo and Vitale2014; Bayet-Goll et al. Reference Bayet-Goll, Uchman, Daraei and Neto de Carvalho2021 b). According to studies by Noffke (Reference Noffke2000) and Mata & Bottjer (Reference Mata and Bottjer2012), compared to muddy lagoons, those lagoonal settings that are periodically associated with tractive sand deposition (i.e. high clastic influx) have had a higher potential for the development of translucent substrates with mat growth. In the Shirgesht lagoonal settings, suspended mud was the dominant sediment size. However, storm activity resulted in some sand being infiltrated into the environment, providing appropriate translucent quartz sand substrates for MISS to develop. According to Bayet-Goll et al. (Reference Bayet-Goll, Buatois, Mángano and Daraei2021 b), in such environments, during storms, appropriate substrates for mat growth are deposited (e.g. lagoonal washover sands; Noffke, Reference Noffke2000), whereas during low-energy conditions these substrates are covered by muddy deposits preventing MISS erosion. Variable hydrodynamical energy levels in the Shirgesht lagoons led to a discontinuous deposition owing to the alternation of physical and biogenic processes. These conditions caused mat-building benthic cyanobacteria to flourish. In other words, in the interfingering washover fan and lagoonal deposits, the effects of energy-level fluctuation recorded at several scales of rhythmic interlamination created fine sediments (silt and mud) that acted as a protective layer (blanket) on the substrate.
However, variations in sediment supply in the lagoon, possibly due to ephemeral changes in the tidal inlet position/dimension or removal of the barrier-island by storms, could have temporarily provided optimum conditions for the activity of biota such as resilient surface detritus-feeding polychaetes (CR ichnofabric). Increase in water turbidity because of sudden and higher-energy sediment influx, typical of washover fan deposits, could have assisted the development of CR ichnofabric. At the peak of wave activity and barrier destruction, the high rate of washover sand accumulation and the creation of unstable depositional substrates in the washover fan prohibited mat-building microorganisms and most tracemakers (e.g. filter- and suspension-feeding benthos) from colonizing the substrate, except for the Rosselia maker, which appears to have endured highly energetic conditions and sedimentation rates (Bayet-Goll et al. Reference Bayet-Goll, Knaust, Daraei, Bahrami and Bagheri2021 d). The presence of a highly stressed depositional substrate in washover sands forced the tracemaker of Rosselia to adjust its burrows to the conditions, resulting in the development of stacked segments. These adjustments prevented the organism from being buried by washover sands and maintained its connection with the sediment–water interface. Short-term colonization and stressed environments in host facies resulted in the development of CR ichnofabric (Nara, Reference Nara2002; Netto et al. Reference Netto, Tognoli, Assine and Nara2014; Bayet-Goll et al. Reference Bayet-Goll, Knaust, Daraei, Bahrami and Bagheri2021 d). High sedimentation rate and high turbidity levels in the water column near the sea-floor prevented mat-building microorganisms from affecting the substrate and enabling MISS to develop.
Toward the tidal flat, environmental factors (energy level, substrate moisture, texture, heterogeneity and its regular subaerial exposure) as well as the impact from tracemakers were the most important influences on the distribution of MISS (e.g. Noffke et al. Reference Noffke, Gerdes and Klenke2003; Tang et al. Reference Tang, Shi, Jiang, Wang, Noffke and Chafetz2012; Bayet-Goll & Daraei, Reference Bayet-Goll and Daraei2020; Bayet-Goll et al. Reference Bayet-Goll, Buatois, Mángano and Daraei2022). The presence of MISS such as shrinkage cracks and petee ridges in the upper tidal flat (mudflats and upper-intertidal flat) is linked to multiple generations of microbial mats and is attributed to cycles of mat-growth–exposure–cracking in the peritidal environment (Noffke et al. Reference Noffke, Beukes, Bower, Hazen and Swift2008). In other words, increase in the width and abundance of polygonal and reticulate shrinkage cracks and cracks formed along ripple-mark crests in the heterolithic intervals of the mudflats and upper-intertidal flat is attributed to lengthy subaerial exposure of the substrates and substantial shrinkage. However, in the supratidal facies, a significant decrease in the thickness and abundance of MISS is mainly attributed to successive reduction in moisture level and translucent substrates, and concomitant increase in mat decomposition (Gerdes et al. Reference Gerdes, Klenke and Noffke2000; Bayet-Goll & Daraei, Reference Bayet-Goll and Daraei2020). Observations on both the modern and ancient microbial mats in tidal flats suggest that the maximum activity of microbial communities and abundance of MISS are detected in mixed sand–mudflats (Gerdes et al. Reference Gerdes, Klenke and Noffke2000; Tang et al. Reference Tang, Shi, Jiang, Wang, Noffke and Chafetz2012; Cuadrado et al. Reference Cuadrado, Perillo and Vitale2014; Davies et al. Reference Davies, Liu, Gibling and Miller2016; Lakhdar et al. Reference Lakhdar, Soussi and Talbi2020). Essentially, one of the key factors triggering the growth of MISS-forming microbial mats in tidal rhythmites of the intertidal zone and the onset of biostabilization is the presence of non-continuous, episodic deposition of sand- and silt-sized clastic sediments (Mata & Bottjer, Reference Mata and Bottjer2009; Eriksson et al. Reference Eriksson, Sarkar, Banerjee, Porada, Catuneanu, Samanta, Seckbach and Oren2010; Bayet-Goll & Daraei, Reference Bayet-Goll and Daraei2020). In the upper-intertidal flat with tidal rhythmites, as a result of alternation of bedload transport and suspension fallout during slack-water periods (Dalrymple et al. Reference Dalrymple, Baker, Harris and Hughes2003), various structures related to MISS could form. In tidal rhythmites, trace fossils with simple feeding strategies of trophic generalists (Pl ichnofabric) are seen either associated with MISS or in sedimentary units hosting these beds. The association of Pl ichnofabric with MISS in the heterolithic intervals of these facies shows that the activity of microorganisms producing MISS was not completely restricted. In the lower-intertidal flat, a decrease in mud content, together with an increase in sedimentation rate and currents, resulted in the development of muddy siltstone–sandstone heterolithics with mostly flaser and subordinate wavy bedding. Considering the composition and distribution of trace fossils (including Sk and CoSk ichnofabrics) and the presence of bioturbated units, the lower-intertidal flat settings might have corresponded to high-stress environments that presumably led the tracemakers to adopt opportunistic strategies. As a result, the Sk and CoSk ichnofabrics can be attributed to opportunistic suspension-feeders in unstable and physically controlled substrates. Generally, in the lower-intertidal flat settings, the clastic influx was too high to allow mat-building microorganisms and most tracemakers to keep up with the accumulation rate and to rework previously deposited sediment. Occurrence of equilibrium structures (upward-shifting), such as Conichnus and escape traces in the CoSk ichnofabric, reflects tracemakers keeping up with accumulation rate by periodic upward shifting in an aggrading substrate (e.g. Savrda, Reference Savrda2002; Bayet-Goll et al. Reference Bayet-Goll, Uchman, Daraei and Neto de Carvalho2021 b).
7.a.2. Shoreface environments
In the foreshore environment, owing to the presence of strong currents and well-sorted, clean, quartzose sand with parallel lamination, opportunistic colonization of the substrate by the tracemakers of the Sk ichnofabric (and locally CoSk) occurred. These conditions also characterized the upper shoreface and tidal inlet settings where evidence for suspension-feeding and high-energy processes (e.g. trough and planar cross-bedding, parallel lamination, basal scouring) is common. In these facies zones, high sedimentation rates and erosion due to steady migration of bedforms, as well as sheet-flow conditions due to wave swash and backwash, were key in producing clean sandy substrates, precluding MISS development and promoting the formation of vertical domichnia. The high abundance of deep-tier, suspension-feeding traces with permanent connection to the sea floor and deep burrows of the Sk ichnofabric are indicative of inhabitation of tracemakers (lophophorate phoronids polychaetes; Pemberton & Frey, Reference Pemberton and Frey1984) in environments dominated by traction currents, frequent wave reworking and an intermittently high current velocity. In the upper part of the succession of the shoreface–offshore complex, the occurrence of multiple palaeocurrents resulting from the interaction of longshore currents and storm currents points to a mobile substrate that was inappropriate for benthic microbial communities (Fig. 16). The great length observed in the suspension-feeding traces suggests the presence of long, vermiform tracemakers that adapted to such environmental conditions, mainly by pulling themselves back into their burrows during turbulent events. The mixing of the water column by waves and currents, suspending food particles, and the general scarcity of endobenthic nutrition within clean sandy substrates of the Sk ichnofabric would have furnished an ideal environment for long vermiform tracemakers, which lived in vertical dwelling burrows (e.g. Droser, Reference Droser1991; Desjardins et al. Reference Desjardins, Mángano, Buatois and Pratt2010; Knaust et al. Reference Knaust, Roger and Thomas Curran2018). The establishment of dense, suspension-feeding infaunal communities characteristic of piperock occurs mainly in advection-dominated benthic substrates (Knaust et al. Reference Knaust, Roger and Thomas Curran2018). Moreover, the persistence of slow, continuous deposition conditions is also an important factor for piperock accumulation (McIlroy & Garton, Reference McIlroy and Garton2010). Such systems are prevalent in the upper shoreface and flaser-bedded sandstones of the intertidal flat (see above), which are composed of coarse-grained sediment with more efficient hydrological connections between the bottom and interstitial water (Knaust et al. Reference Knaust, Roger and Thomas Curran2018). However, the high rate of sedimentation, erosion, and upper flow regime conditions during wave swash and backwash in the foreshore likely explain the lack of dense piperock ichnofabrics in the Shiegesht Formation.
In the lower to middle shoreface, where shoaling and breaking waves and longshore currents generated a high-energy setting, thick aggradational beds with sharp erosional bases and HCS were produced (Fig. 16). In these conditions, again, parameters such as higher depositional energies, accumulation rates and the occurrence of erosion events and unstable mobile substrate, as reflected by a greater degree of bed amalgamation and a decrease in the time between successive storm events, prevented MISS formation and preservation. Under these circumstances, high-energy, storm-dominated settings with low endobenthic nutrient levels in the sandy substrates could have provided optimum conditions for the activity of detritus-feeding polychaetes and suspension-feeding infaunal tracemakers of the RoSk ichnofabric. The unstable conditions in these substrates forced the tracemakers to adjust their burrows, which resulted in the development of stacked segments in Rosselia. According to Bayet-Goll et al. (Reference Bayet-Goll, Knaust, Daraei, Bahrami and Bagheri2021 d), in the lower to middle shoreface areas, characterized by thick, amalgamated storm beds because of highly energetic conditions and high sedimentation rate as well as short-lived fair-weather conditions between storm events, only a very small amount of mud was deposited. In such situations, infaunal suspension-feeding animals, capable of readjusting their dwellings after sedimentation–erosion events, were able to survive. The tracemaker of Rosselia in the lower–middle shoreface was capable of maintaining a permanent connection to the sediment–water interface through upward migration, causing the development of vertically stacked forms to avoid burial.
Moreover, the storm-generated shoreface sandstone beds were unsuitable for the activity of shallow-tier deposit-feeders due to general paucity of endobenthic food and severe storm events. As a result, these substrates could have provided optimum conditions for the activity of r-selected organisms such as decapod crustaceans that generated traces in the Th ichnofabric. In this ichnofabric, the high abundance of burrow systems of dwelling and feeding ethology (Thalassinoides), together with evidence of the increasing degree of bioturbation, burrow size, penetration depth and complexity, is collectively indicative of inhabitation of tracemakers in highly energetic environments with high sedimentation rates and well-oxygenated water due to persistent wave agitation. Unlined burrows and sharp burrow boundaries point to unstable and physically controlled substrates. Unlike inhabiting organisms of softground communities, branching burrow networks of Thalassinoides were adapted to cope with deep erosional events or rapid sediment supply (Pemberton et al. Reference Pemberton, Zhou and MacEachern2001; Bayet-Goll et al. Reference Bayet-Goll, Myrow, Aceñolaza, Moussavi-Harami and Mahboubi2016).
7.a.3. Offshore environments
In the offshore transition to the upper offshore belt, a high degree of bioturbation and a higher abundance of fine-grained sediments than in the shoreface facies indicate the position of these facies below the FWWB, and also point to a long-lasting deposition in a low-energy setting. Sandstones with HCS are interpreted as products of occasional storms disturbing the substrate. As a result, muddy substrates rich in nutrients were developed between the FWWB and SWB, providing ideal conditions to enhance tracemaker activity and diversity. Typically, in this setting, homogeneous distribution of food, normal salinity and oxygenated water due to persistent wave agitation led to the development and preservation of trace fossils related to CrSkRo, Th, CT and CrHe ichnofabrics (Figs 15 and 16).
In the distal lower-shoreface / offshore-transition zone, based on the overall sedimentological characteristics and highly variable ichnological signatures, CrSkRo ichnofabric represents an environment with highly fluctuating hydrodynamic conditions. These fluctuations were driven by the intermittent alternation of high-energy storm events and low-energy, post-storm conditions, which mostly influenced the ichnological characteristics of the CrSkRo ichnofabric. The vertical partitioning of the habitat in this setting was caused by fluctuations in the physical character of the substrate. Such fluctuations cause the recurrence of ‘resident fair-weather communities’ (K-selected species) of the softground substrates with opportunistic colonizing strategies in unstable and shifting substrates (e.g. Pemberton et al. Reference Pemberton, Zhou and MacEachern2001; Sharafi et al., Reference Sharafi, Rodríguez-Tovar, Janočko, Bayet-Goll, Mohammadi and Khanehbad2022). K-selected equilibrium species show specialized feeding behaviour or long-term feeding activity via effective use of resources within the substrate (Bayet-Goll et al. Reference Bayet-Goll, Myrow, Aceñolaza, Moussavi-Harami and Mahboubi2016). Bioturbators with locomotion-foraging trails and deposit-feeding and grazing behaviours depend on the muddy, organic-rich substrates, and are attributed to resident fair-weather communities, which show equilibrium strategies in habitats with low physical stresses (Buatois & Mángano, Reference Buatois and Mángano2011 a). In contrast, in sandstone-dominated storm deposits with HCS and internal erosion surfaces, the emplacement and preservation of initial softground suites are reduced dramatically. The existence of deep-tier, suspension-feeding traces in event beds reflects the episodic generation of a community of opportunistic organisms persisting in an unstable and physically controlled environment (e.g. Pemberton et al. Reference Pemberton, Zhou and MacEachern2001; MacEachern & Bann, Reference MacEachern, Bann, Hampson, Steel, Burges and Dalrymple2008). Numerous studies have mentioned wrinkle structures at the top of tempestites elsewhere, typically in exceptional intervals in the Earth’s history, e.g. post-extinction intervals or the uniqueness of anachronistic Precambrian–Cambrian facies (e.g. Pruss et al. Reference Pruss, Fraiser and Bottjer2004; Twitchett, Reference Twitchett2006; Mata & Bottjer, Reference Mata and Bottjer2009). Conversely, in the offshore-transition facies of the Lashkerak Formation, the high diversity of trace fossils and high bioturbation in the mud-dominated beds with cohesive soft substrates, together with the existence of episodic generation of communities of opportunistic organisms in the event beds (e.g. Diplocraterion, Arenicolites, Rosselia, Skolithos, Thalassinoides, Monocraterion), prevented wrinkle structures being produced and preserved.
Another ichnofabric that caused the disruption of mat-fabrics in the offshore areas could be due to the presence of the Th ichnofabric. However, compared to shallower areas, a lower sedimentation rate and erosion of the sea-floor permitted multiple, successive-tier colonization of locomotion-foraging, deposit-feeding and grazing behaviours (e.g. Cruziana, Planolites, Helminthopsis) and opportunistic, suspension-feeding structures. In the offshore areas, below the FWWB, burrow networks of Thalassinoides (with evidence of unlined, uncompacted/sharp boundaries and passively filled burrows) are characteristic of the Thalassinoides-dominated Glossifungites surfaces (MacEachern and Pemberton Reference MacEachern and Pemberton1992; Bayet-Goll et al. Reference Bayet-Goll, Myrow, Aceñolaza, Moussavi-Harami and Mahboubi2016). In these areas, multiple overprinted softground communities and Thalassinoides networks of firmground communities represent a phase change from softground (early endobenthic deposit-feeders) to firmground substrates during depositional hiatuses after transgressive or regressive erosional events (e.g. Ghibaudo et al. Reference Ghibaudo, Grandesso, Massari and Uchman1996; Rodrìguez-Tovar et al. Reference Rodríguez-Tovar, Valera and López2007; Uchman et al. Reference Uchman, Pervesler, Hohenegger and Dominici2011).
In the upper offshore, in the vicinity of the SWB, more stable conditions with a low sedimentation rate as well as the prevalence of aerobic conditions are expected (Bayet-Goll et al. Reference Bayet-Goll, Myrow, Aceñolaza, Moussavi-Harami and Mahboubi2016). Because, in the vicinity of the SWB, episodic storm events are less frequent than in the lower-shoreface / offshore-transition zone, post-storm stable conditions are longer and promote not only the formation of muddy, organic-rich units but also the development of CT and CrHe ichnofabrics. High intensity and uniformity of burrowing in these ichnofabrics in post-storm intervals led to a rapid increase in the abundance, diversity and higher proportion of locomotion, grazing, deposit-feeding and surface detritus-feeding traces. Such infaunal and epifaunal reworking near the SWB did not allow the substrate stabilization by microbial activity. In the host deposits of CT ichnofabric, the close association of Trichophycus with muddy, organic-rich substrates points to the activity of tracemakers in stable habitats and muddy substrates above the SWB with a high nutrient content, low-flow regimes and well-oxygenated conditions, and hence, to a wider colonization window. According to Bayet-Goll et al. (Reference Bayet-Goll, Knaust, Daraei, Bahrami and Bagheri2021 d), Trichophycus resulting from resident fair-weather tracemakers is a common trace fossil in the archetypical Cruziana ichnofacies in the Shirgesht Formation, and reflects the activity of populations demonstrating a K-selected or climax strategy. Toward the shallower areas, near the FWWB (lower-shoreface / offshore-transition zone), because of short fair-weather conditions between storm events, the mud layers are thin and limited in lateral extent. Moreover, nutrient content is insufficient for supporting the producer of Trichophycus. This caused a decline in the length and diameter of burrows, as well as the density and the number of successively stacked causative burrows. Continuation of these conditions into shallower water caused CT to be replaced by RoSk and Th ichnofabrics.
The high abundance of resident fair-weather communities in CrHe ichnofabric is suggestive of tracemakers in softground substrates with homogeneous distribution of food, low sedimentation rate of coarse sandy material and presence of oxygen near the sediment–water interface, and hence of a wide colonization window. High ichnodiversity and uniform bioturbation is indicative of slow deposition and a continuous input of mud with nutrients under quiet, fully marine conditions that gave shallow-tier tracemakers enough time to keep up with the sedimentation rate (MacEachern et al. Reference MacEachern, Pemberton, Gingras, Bann and Miller2007; Buatois & Mángano, Reference Buatois and Mángano2011 a). Furthermore, vertical partitioning of the habitat was generated by fluctuations in hydrodynamic energy and nutrient level, oxygen content and sedimentation rate. The establishment of stable conditions near the SWB provided the right conditions for a suite of initial tier tracemakers (locomotion-foraging trails and early endobenthic deposit-feeding and grazing burrows), which reworked the sediment during early colonization of the substrate as shallow-burrowers. Subsequently, episodic biological activity, due to episodic increase in sedimentation rate and hydraulic energy during storms, permitted multiple, successive-tier colonization of the substrate by opportunistic, suspension-feeders (Skolithos) and deep-tier deposit-feeders (Chondrites).
In the lower offshore, below the SWB, the intensity of bioturbation caused MISS-generating microorganisms to shift toward the less bioturbated lower-offshore / shelf setting (Fig. 16). The development and preservation of wrinkle structures in the lower-offshore / shelf setting (dominated by the ChPl ichnofabric) is due to the low ichnodiversity, scattered burrowing and the absence of large burrows, together with complex trace systems. The strong impact of harsh environmental stress in these areas (e.g. shallow-water anoxia and euxinia; Pruss et al. Reference Pruss, Fraiser and Bottjer2004; Grice et al. Reference Grice, Cao, Love, Bottcher, Twitchett, Grosjean, Summons, Turgeon, Dunning and Jin2005) is important for the formation of microbial matgrounds. The cohesive substrate, well below the SWB, resulted in a decrease in the diversity, size and bioturbation degree so that bioturbation was restricted mainly to sediment surface and deep-tier deposit-feeding. In the basinward direction, the replacement of CrHe and CT ichnofabrics by deep-tier deposit-feeding and shallow, foraging burrows, along with a decrease in the size of burrows and ichnodiversity, suggests low oxygen content and periods of very rapid mud accumulation in the lower-offshore setting. The occurrence of thick, muddy units, the lack of wave-generated sedimentary structures, and the mostly non-burrowed nature of the host sediment of ChPl ichnofabric all indicate a highly stressed depositional environment, in which only tracemakers adapted to low oxygen content could have survived. However, the presence of shallow-tier, locomotion-foraging trails and deposit-feeding behaviours may be related to episodic storm events causing fluctuations in nutrient and oxygenation levels.
7.b. Linking MISS distribution to marine benthos evolution
The Cambrian–Ordovician evolutionary radiations, particularly involving the increase of suspension-feeding metazoans as well as progressive ecospace utilization (Mángano & Buatois, Reference Mángano and Buatois2016, Reference Mángano and Buatois2017; Buatois et al. Reference Buatois, Mángano, Minter, Zhou, Wisshak, Wilson and Olea2020), led to substrates changes in shallow-marine environments from ‘Ediacaran-style soft substrates’ into ‘Phanerozoic-style soft substrates’, probably with the development of mixed-layer substrates (soupy substrates homogenized by bioturbation; see Ekdale et al. Reference Ekdale, Muller and Novak1984). There was a shift of microbial matground-dominated ecosystems to more stressful settings after the Cambrian–Ordovician evolutionary radiations (Hagadorn & Bottjer, Reference Hagadorn and Bottjer1999; Mángano & Droser Reference Mángano, Droser, Webby, Droser, Paris and Percival2004; Mata & Bottjer Reference Mata and Bottjer2009; Buatois & Mángano, Reference Buatois and Mángano2011 a, b; Duperron & Scasso, Reference Duperron and Scasso2020). The Ordovician devastation of matgrounds and the development of mixed-layer substrates resulted from the increase in bioturbation intensity, ichnodiversity and burrow size. In both barrier-island and shoreface–offshore complexes of the Shirgesht Formation, the restriction of matgrounds to inhospitable environments such as lower-offshore / shelf or tidal-flat and back-barrier environments, is consistent with the lack of intense bioturbation (Pl and ChPl ichnofabrics) in these environments. The reason for this behaviour might have been related to ecological tolerance of producers of MISS to different environmental factors, and also the effect of increasing bioturbation intensity, ichnodiversity and deepening tiering structure of ichnofaunas derived from evolutionary changes in benthic faunas. Although the effects of environmental parameters on ecological tolerance of mat-building microorganisms and tracemakers were discussed earlier, changes in benthic fauna after the evolutionary radiations, as well as organism–matground interactions, are other important factors that should be considered.
During the GOBE, a substantial increase took place in the abundance and diversity of deposit-, detritus-, suspension-feeders and grazers in shallow-marine habitats, as well as suspension-feeders and predators in deeper marine communities (Servais et al. Reference Servais, Lehnert, Li, Mullins, Munnecke, Noutzel and Vecoli2008, Reference Servais, Owen, Harper, Kroger and Munnecke2010). This event, which is considered an important milestone in the evolutionary history of life, resulted in a vast change from the trilobite-dominated Cambrian Evolutionary Fauna to the post-Cambrian Evolutionary Fauna with a higher diversity and abundance (Finnegan & Droser, Reference Finnegan and Droser2008; Mángano et al. Reference Mángano, Buatois, Waisfeld, Muñoz, Vaccari and Astini2021). In shallow-marine Early Ordovician siliciclastic deposits, trilobites and trilobitomorphs were among the most common tracemakers (Seilacher, Reference Seilacher1999; Mángano & Droser, Reference Mángano, Droser, Webby, Droser, Paris and Percival2004; Mángano et al. Reference Mángano, Buatois, Wilson and Droser2016). An increase in the abundance and diversity of branched, burrow mazes and boxworks with sophisticated feeding strategies occurred during the Middle to Late Ordovician, but arthropods became less important as tracemakers (Mángano & Droser, Reference Mángano, Droser, Webby, Droser, Paris and Percival2004; Mángano et al. Reference Mángano, Buatois, Wilson and Droser2016). In the mixground ecosystems of the Shirgesht Formation with the Sk, CR, RoSk, CoSk, Th, CrSkRo, CT and CrHe ichnofabrics, vagile bilaterian metazoans, arthropod flattened trails, deep bioturbators (suspension-feeding metazoans, deposit- and detritus-feeders), and predators made up a large population of tracemakers, and therefore a widespread increase in ecospace utilization and tiering complexity took place in the substrates of the shallow-marine ecosystems. Consequently, because of the development of permanent and temporary dwelling burrows or infaunalization with vertical partitioning of the infaunal and epifaunal communities (i.e. tiering structures), and their greater role in sediment mixing and bioirrigation, appropriate conditions for the growth and development of matgrounds were not provided. As a result of the Ordovician radiation, progressive ecospace utilization by suspension-, deposit- and detritus-feeding producers of burrows such as Diplocraterion, Arenicolites, Skolithos, Rosselia, Teichichnus, Conichnus, Thalassinoides, Trichophycus and Arthrophycus (CR, RoSk, CoSk, Th, CrSkRo and CT ichnofabrics) can be detected in open-marine substrates. According to Mángano & Buatois (Reference Mángano and Buatois2016), after these evolutionary breakthroughs, mixing of the sediment occurred through increase in the exploitation of infaunal ecospace, especially in offshore areas.
A considerable enhancement in infaunalization is evidenced by the general occurrence of suspension-feeding infaunal communities (Sk ichnofabric). During the early Palaeozoic, in the sedimentary substrates of offshore to lower-shoreface settings, vagile bilaterian metazoans and arthropods were the major tracemakers of the resident fair-weather community. However, sandstones in the nearshore settings, particularly upper to middle shoreface/foreshore deposits, were influenced by dwelling, escape or equilibrium structures of suspension-feeding tracemakers (Bromley, Reference Bromley1996; Droser et al. Reference Droser, Hughes and Jell1994, Reference Droser, Jensen, Gehling and McIlroy2004; Mángano & Buatois, Reference Mángano and Buatois2017). The presence of Skolithos piperock as one of the most important ichnofabrics (in cases with a bioturbation depth of more than 1 m; e.g. Davies et al. Reference Davies, Herringshaw and Raine2009) is one of the main indicators of an increase in the exploitation of infaunal ecospace in nearshore environments. This increase occurred after the ‘Cambrian agronomic revolution’ (Seilacher & Pflüger, Reference Seilacher, Pflüger, Krumbein, Peterson and Stal1994) with the replacement of matgrounds by mixgrounds as a result of the increased depth/complexity of infaunal burrowing by metazoans (Seilacher, Reference Seilacher1999; Mángano & Buatois, Reference Mángano and Buatois2016). During Cambrian Stage 2, suspension-feeding infaunal communities developed as ecological drivers, causing the increase in diversity and frequency of deposit-feeding structures after Cambrian Stage 3 (Mángano & Buatois, Reference Mángano and Buatois2016).
The occurrence of equilibrium-adjustment behaviours of infaunal terebellid populations (synchronous upward movement by Rosselia producers) related to the CR and RoSk, ichnofabrics has also been considered responsible for the increase in progressive ecospace utilization (Desjardins et al. Reference Desjardins, Mángano, Buatois and Pratt2010; Bayet-Goll et al. Reference Bayet-Goll, Knaust, Daraei, Bahrami and Bagheri2021 d) that took place in the shallow-marine ecosystems of this formation. The importance of feeding structures of detritus-feeders (CR and RoSk ichnofabrics) and deposit-feeders (CT ichnofabric) is that such structures demonstrate complex morphological/behavioural patterns of trace fossils with systematic burrowing and sophisticated feeding behaviours, exploiting the infaunal ecospace. Despite its Early Cambrian appearance, Trichophycus remained uncommon until the Early Ordovician, and when compared to other burrowers, Trichophycus tracemakers may have only been local contributors to the GOBE infaunalization (Bayet-Goll et al. Reference Bayet-Goll, Uchman, Daraei and Neto de Carvalho2021 c). However, due to the Ordovician radiation, the producer of Trichophycus exerted an important role in the Shirgesht marine environment by increasing the complexity of tiering and the depth of bioturbation. In fact, a notable increase in exploitation of infaunal ecospace in the offshore settings is caused by the increase in the degree of ichnocomplexity and size of the burrow systems in CT and Th ichnofabrics, e.g. three-dimensional burrow systems represented by Trichophycus and branched networks of Thalassinoides (Bayet-Goll et al. Reference Bayet-Goll, Uchman, Daraei and Neto de Carvalho2021 c). Moreover, the strong impact of the Ordovician radiation on tracemakers of the resident fair-weather community (e.g. arthropods, vagile bilaterian metazoans) in shallow-marine habitats led to an increase in biological sediment mixing and irrigation. In the CrSkRo and CrHe ichnofabrics, trilobites and trilobitomorphs are considered dominant in a wide range of ethological categories (crawling, resting, walking, sideways grazing and swimming grazing). It is likely that mixing and reworking of sediment and thus disruption of matgrounds in the offshore settings was done mainly by trilobites and trilobitomorphs. In addition, a noticeable increase in the depth of burrow penetration and change from shallow-tier to mid-tier trace fossils attributable to trilobites have been documented after the evolutionary radiations in the Cambrian–Ordovician (Mángano & Buatois, Reference Mángano and Buatois2016, Reference Mángano and Buatois2017; Santos et al. Reference Santos, Mángano, Soares, Nogueira, Bandeira and Rudnitzki2017; Bayet-Goll et al. Reference Bayet-Goll, Daraei, Geyer, Bahrami and Bagheri2021 a). The dominance of the mid-tier trilobite trace fossils (i.e. Cruziana rugosa group: C. rugosa, C. furcifera, C. goldfussi) in CrSkRo and CrHe ichnofabrics could be considered as the evidence for the Ordovician infaunalization (cf. Santos et al. Reference Santos, Mángano, Soares, Nogueira, Bandeira and Rudnitzki2017). It should be mentioned that the development and preservation of wrinkle structures and circular to elliptical erosional remnants (between FWWB and SWB) occurred frequently in lower Cambrian facies, unlike in the Ordovician offshore-transition / upper-offshore facies that lacks MISS (e.g. Bailey et al. Reference Bailey, Corsetti, Bottjer and Marenco2006; Mata & Bottjer, Reference Mata and Bottjer2009, Reference Mata and Bottjer2012). Generally, because of the absence of middle-tier arthropod burrows and dwelling structures of suspension-feeders (e.g. Psilonichnus, Diplocraterion, Arenicolites, Skolithos) or deposit-feeders (e.g. Thalassinoides, Spongeliomorpha Rhizocorallium) before the ‘Cambrian agronomic revolution’ (Seilacher & Pflüger, Reference Seilacher, Pflüger, Krumbein, Peterson and Stal1994; Bottjer et al. Reference Bottjer, Hagadorn and Dornbos2000; Buatois et al. Reference Buatois, Netto, Mángano and Carmona2013), the offshore-transition / upper-offshore zone could have been associated with wrinkle structures (e.g. Bayet-Goll et al. Reference Bayet-Goll, Daraei, Geyer, Bahrami and Bagheri2021 a).
The Ordovician evolution of shallow-marine benthos likely had a profound impact on the intensity of bioturbation, and consequently MISS distribution, in the Shirgesht Formation as well. Consequently, these conditions in the shallow-marine part of the basin caused mat-building microorganisms to migrate into environments that experienced delayed colonization by burrowing organisms (Bayet-Goll et al. Reference Bayet-Goll, Daraei, Geyer, Bahrami and Bagheri2021 a, b), such as the lower-offshore / shelf or tidal-flat and back-barrier settings. Extensive matgrounds have recently been documented in deep-sea and hypersaline settings of the northern Gondwana platform, for example in the Ordovician of the Alborz Mountains of northern Iran (Lashkerak and Ghelli formations; Bayet-Goll et al. Reference Bayet-Goll, Buatois, Mángano and Daraei2021 b). Here, the early Palaeozoic evolutionary radiations have been shown to shift microbial matground-dominated ecosystems to deep-marine (Ghelli Formation; middle–upper Ordovician) and wave-dominated estuarine settings (Lashkerak Formation; lower to upper Ordovician). In progradationally stacked, deep-marine lobe complexes, patchily distributed wrinkle structures occur in the lobe-axis / off-axis and lobe-fringe / distal-fringe settings, associated with graphoglyptids and grazing traces. Moreover, generation of microbial mats in the wave-dominated shoreface–offshore complex was inhibited due to the establishment of abundant and diverse infaunas capable of sediment reworking. Overall, soft-bodied and skeletal organisms were forced to migrate into relatively inhospitable environments not only because of the increase in competition for ecospace utilization (e.g. resource acquisition) but also a more complex tiering linked to the establishment of the mixed-layer substrates in shallow-marine ecosystems, which resulted in the restriction of microbial communities to deep-sea and hypersaline settings. In these exceedingly inhospitable deeper-water environments, the reduced effects of sediment mixing and bioirrigation would have led to a decrease in or removal of restrictions on microbial growth (e.g. fewer grazing metazoans; Bayet-Goll et al. Reference Bayet-Goll, Buatois, Mángano and Daraei2021 b). Similarly, in the lower-offshore / shelf or tidal-flat and back-barrier settings of the Shirgesht Formation with evidence of microbial matground ecosystems, tracemakers with simple feeding strategies (e.g. trophic generalists; Pl ichnofabric) and deep-tier deposit-feeding or shallow, foraging burrows (ChPl ichnofabric) had negligible effect on sediment mixing and bioirrigation. Therefore, in these settings the restrictions for microbial growth were lifted.
8. Conclusions
This study investigates the shift of microbial matground-dominated ecosystems to environments inhospitable for bioturbators as a result of increasingly restricted environmental factors as well as advanced infaunalization after evolutionary radiations of the early Palaeozoic benthos (i.e. ‘Cambrian agronomic revolution’ and the GOBE). Based on sedimentological and ichnological evidence, barrier-island and wave-dominated shoreface–offshore complexes are identified in the shallow-marine depositional system of the lower–middle Ordovician Shirgesht Formation (Central Iran). These two complexes contain ichnological assemblages as well as ichnofabric and MISS (mat fabrics) distributions, which are shown to be linked to environmental parameters that operated at the scale of sub-environments. In both complexes, the restriction of matgrounds to inhospitable environments such as lower-offshore / shelf or tidal-flat and back-barrier settings is consistent with the lack of intense bioturbation (Pl and ChPl ichnofabrics) in these settings. Moreover, the facies of the foreshore, tidal-inlet, lower- to upper-shoreface, and offshore-transition / upper-offshore zones (as mixground ecosystems) lack MISS. It is shown that in the mixground ecosystems with Sk, CR, RoSk, CoSk, Th, CrSkRo, CT and the CrHe ichnofabrics, a widespread increase in ecospace utilization and vertical partitioning of the infaunal and epifaunal communities led to inadequate conditions for the growth and development of matgrounds. Moreover, the absence of MISS in the shallow-marine ecosystems is linked to the development of mixed-layer substrates and intensely bioturbated mixground substrates. This increase in bioturbation in the shallower waters is why microorganisms producing MISS shifted to and flourished in lower-offshore / shelf or tidal-flat and back-barrier settings in the early Palaeozoic. In the Shirgesht mixground ecosystems too, a considerable increase in vagile bilaterian metazoans, arthropods, deep bioturbators (suspension-feeding metazoans, deposit- and detritus-feeders) and predators is considered to have been the main reason for the lack of development and/or preservation of benthic microbial communities. In summary, because of the early Palaeozoic evolutionary radiations, the shallow-marine Shirgesht substrates underwent intense burrowing and reworking and this forced soft-bodied organisms to migrate into less hospitable environments, such as the lower-offshore / shelf settings with dysoxic to anoxic conditions or the tidal-flat / back-barrier settings with fluctuating salinities.
Acknowledgements
Financial support and field expenses were provided by the Department of Earth Sciences, Institute for Advanced Studies in Basic Sciences (IASBS), Research Grants 100/151/13302. I would like to thank reviewers Andrew Rindsberg (Alabama, USA), Alfred Uchman (Kraków, Poland) and the executive editor Emese M. Bordy (Cape Town, South Africa) for their helpful comments and constructive criticism that greatly improved this paper. Mehdi Daraei (Zanjan, IASBS) is gratefully acknowledged for improving the English of the manuscript.
















