


1. Introduction
The family Anguinae has three living genera: Pseudopus (Eurasia), Ophisaurus (Southeast Asia, North Africa and North America) and Anguis (Europe, Southwest Asia) (see reviews in Augé, Reference Augé2005; Gvoždík et al. Reference Gvoždík, Jandzík, Lymberakis, Jablonski and Moravec2010; Klembara, Hain & Dobiašová, Reference Klembara, Hain and Dobiašová2014). Ophisaurus fossils come from Eurasia North Africa and North America, while Anguis and Pseudopus are currently reported from Europe (Fejérváry-Lángh, Reference Fejérváry-Lángh1923; Holman, Reference Holman1970; Augé, Reference Augé2005; Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010; Klembara, Reference Klembara2012, Reference Klembara2015; Blain et al. Reference Blain, Agustí, López-Garcia, Haddoumi, Aouraghe, Hammouti, Pérez-Gonzáles, Chacón and Sala2013).
Cenozoic anguine fossils are mostly disarticulated with the exception of Ophisauriscus quadrupes skeletons and osteoderms from the Middle Eocene of Geiseltal (Kuhn, Reference Kuhn1940; Meszoely & Haubold, Reference Meszoely and Haubold1975; Klembara, Reference Klembara1981) and from Messel (Sullivan, Keller & Habersetzer, Reference Sullivan, Keller and Habersetzer1999), both in Germany. Later revisions, however, have questioned their classification as Anguinae (Sullivan, Keller & Habersetzer, Reference Sullivan, Keller and Habersetzer1999; Conrad, Reference Conrad2008; Conrad et al. Reference Conrad, Ast, Montanari and Norell2011). A further exception is Pseudopus laurillardi from the Early Miocene of Germany which has an articulated skull and well-preserved portions of the vertebral column and integument (Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010). This is the reference species for abundant disarticulated skeletal elements attributed to Pseudopus discovered at other Early and Middle Miocene localities in Central and Western Europe. Also recorded are: (1) a partly preserved skull of Ophisaurus acuminatus from the German lower Late Miocene (Jörg, Reference Jörg1965); (2) a specimen with a partly articulated skull from the Miocene of Catalonia (Ne Iberian Peninsula) briefly described by Bolet et al. (Reference Bolet, Delfino, Fortuny, Almécija and Alba2013); and (3) Ophisaurus sp. described from the Middle Miocene sediments of Slovakia by Klembara (Reference Klembara1986 a) which has a short vertebral column section and osteoderms. Recently, the cranial and postcranial remains of Ophisaurus spinari have been described from the late Middle Miocene of Austria (Böhme & Vasilyan, Reference Böhme and Vasilyan2014). Although not articulated, the parietal, several other cranial elements, vertebrae and osteoderms were attributed to this species by the authors (however, see below).
All other Cenozoic anguine species of Europe (see below), North Africa (Blain et al. Reference Blain, Agustí, López-Garcia, Haddoumi, Aouraghe, Hammouti, Pérez-Gonzáles, Chacón and Sala2013) and North America (Holman, Reference Holman1970; Estes, Reference Estes and Wellenhofer1983) were described on the basis of the disarticulated cranial elements. This is especially relevant for Ophisaurus species described mainly from parietals; and to a lesser extent to Pseudopus and Anguis descriptions. Recently, an incomplete parietal from the Late Eocene of Spain was attributed to Anguinae indet (Bolet & Evans, Reference Bolet and Evans2013). A further Paleogene find of parietal attributed to Ophisaurus comes from the Late Eocene of the Hampshire Basin of southern England. However, because this parietal exhibits no specific features it was described as Ophisaurus sp. (Klembara & Green, Reference Klembara and Green2010). From the same locality, the parietal of Headonhillia parva was also described (Klembara & Green, Reference Klembara and Green2010). An Oligocene species of the genus Ophisaurus is represented by O. roqueprunensis from the Early to Late Oligocene of France and Belgium (Augé, Reference Augé1992, Reference Augé2005; Augé & Smith, Reference Augé and Smith2009), later attributed to the genus Dopasia (Augé, Reference Augé2005) and reallocated to Ophisaurus by Čerňanský, Klembara & Müller, (Reference Čerňanský, Klembara and Müller2016). Within the new material of this species described from Belgium, a partially preserved parietal is present (Augé & Smith, Reference Augé and Smith2009). Four species of the genus Ophisaurus were described from the Early Miocene of the Czech Republic and Germany: O. fejfari, O. spinari and O. robustus from the locality Dolnice (MN 4) (Klembara, Reference Klembara1979, Reference Klembara1981) and O. holeci from the locality Merkur (MN 3) (Klembara, Reference Klembara2015). In O. holeci it was possible to associate the frontals with the parietals. Importantly, an O. spinari parietal was described from Germany (Čerňanský, Rage & Klembara, Reference Čerňanský, Rage and Klembara2015). Unfortunately, the parietal of O. acuminatus (Jörg, Reference Jörg1965) is not preserved. One partial parietal was described from the late Middle Miocene (MN 7–8) sediments of the Pannonian Basin in Hungary and determined as Ophisaurus cf. spinari (Hír et al. Reference Hír, Kókay, Venczel, Gál and Kessler2001). Later, more abundant material of anguines from the same region was described (Venczel & Hír, Reference Venczel and Hír2013). In this disarticulated material, several other parietals were described and also attributed to Ophisaurus cf. spinari (Venczel & Hír, Reference Venczel and Hír2013). Ophisaurus canadensis from the Late Miocene of Canada is based on one vertebra (Holman, Reference Holman1970).
Another partially preserved parietal was described from the Late Pliocene of Italy as Dopasia sp. (Delfino, Bailon & Pitruzzella, Reference Delfino, Bailon and Pitruzzella2011). Augé (Reference Augé2005) started to use the generic name Dopasia, originally used by Gray (Reference Gray1853) for the extant anguine D. gracilis from Southeast Asia, instead of Ophisaurus. Augé (Reference Augé2005) applied the name Dopasia for all extinct and extant species previously described as Ophisaurus from the Cenozoic of Eurasia and North Africa. However, the latest phylogenetic analysis of extant Anguinae showed that it would be more prudent to use the name Ophisaurus for all species living in Southeast Asia, North America and North Africa (Klembara, Hain & Dobiašová, Reference Klembara, Hain and Dobiašová2014).
Three Dopasia (=Ophisaurus) species, D. roqueprunensis, D. coderetensis and D. frayssensis from the Oligocene of France, were described on the basis of dentaries (Augé, Reference Augé2005). There is also one parietal attributed to D. coderetensis (Augé, Reference Augé2005); unfortunately its preservation is very poor and does not allow any detailed comparisons. The species D. coderetensis and D. frayssensis have been assigned to the genus Ophisauromimus by Čerňanský, Klembara & Müller (Reference Čerňanský, Klembara and Müller2016).
Crusafont Pairó & Vallalta (Reference Crusafont Pairó and Villalta1952) mentioned the record of the Ophisaurus skull from the Miocene locality Vallès-Penedès Basin in Catalonia; a revision of this find is needed. Bailon & Augé (Reference Bailon and Augé2012) described a new anguine genus and species, Ragesaurus medasensis from the Lower Pleistocene of Spain. This new taxon is based on one right dentary.
The recognition of two species of Pseudopus, P. ahnikoviensis (Klembara, Reference Klembara2012) and P. rugosus (Klembara, Reference Klembara2015; however, see below), from the Early Miocene locality Merkur (the Czech Republic) helped to re-evaluate Early Miocene anguines from Slovakia and the Czech Republic previously attributed to Ophisaurus on the basis of parietals (Klembara, Reference Klembara1986 a and see below).
The aims of this paper are: (1) to re-describe four previously reported Miocene species of Ophisaurus named on the basis of parietals from the Czech Republic and Germany on the basis of new material; (2) to describe new Ophisaurus, Anguis and Pseudopus specimens from the Early and Middle Miocene Czech and German localities through better understanding of their intraspecific and interspecific variability; and (3) to perform systematic revision of all Ophisaurus, Anguis and Pseudopus species previously described from the Cenozoic in Europe, North Africa and North America. Consequently, our aim is to present precise understanding of Ophisaurus, Anguis and Pseudopus diversity in the Cenozoic of Eurasia, North Africa and North America and to confirm their palaeobiogeography.
2. Materials and methods
The material described here is disarticulated. Several parietals were coated with ammonium chloride before taking photographs.
For comparisons, the skeletons of the following Anguinae species were used: Anguis fragilis (DE 14–21, 24, 25, 45–48); Ophisaurus ventralis (DE 34, 35, 38; AMNH 73057; UF 52539; CM 1411), O. attenuatus (DE 32, 33, 43, 44), O. compressus (DE 50), O. mimicus (DE 49), O. koellikeri (DE 30, 41), O. harti (DE 36, 37, 56, 57, 86; AMNH 34956), O. gracilis (DE 42), Pseudopus apodus (DE 1, 3–13, 22, 23, 29, 52–54, 58, 59; BSPG 1982 X 2383).
The terminology of the anatomical structures is from Fejérváry-Lángh (Reference Fejérváry-Lángh1923), Oelrich (Reference Oelrich1956), Klembara (Reference Klembara1979, Reference Klembara1981), Conrad (Reference Conrad2004), Evans (Reference Evans, Gans, Gaunt and Adler2008) and Klembara, Böhme & Rummel (Reference Klembara, Böhme and Rummel2010).
Institutional abbreviations. AMNH – American Museum of Natural History, New York, USA; BMNH, NHM – Natural History Museum, London, England; BSPG – Bayerische Staatssammlung für Paläontologie, Munich, Germany; CM – Carnegie Museum of Natural History, Pittsburgh, PA, USA; DE – Department of Ecology, Faculty of Natural Sciences, Comenius University in Bratislava, Slovakia; DP FNSP – Department of Palaeontology, Faculty of Natural Sciences, Charles University, Prague, Czech Republic; GM – Geiseltal Museum, Halle, Germany; IRsNB – Institut royal des sciences naturelles de Belgique, Bruxelles; MNHN – Muséum National d'Histoire Naturelle, Paris, France; NMA – Naturmuseum Augsburg, Germany; NMP Pb and Pv – National Museum, Prague, Czech Republic; ROM – Royal Ontario Museum, Toronto, Canada; SMNK (LNK) – Staatliches Museum für Naturkunde Karlsruhe (Landessamlungen für Naturkunde Karlsruhe), Germany; SNM PP – Slovak National Museum, Bratislava, Slovakia; UCBL – Université Claude Bernard, Lyon, France; UF – University of Florida, Gainesville, USA; UM MED – Université de Montpellier, France.
3. Systematic palaeontology
In this section, new specimens of Ophisaurus, Anguis and Pseudopus are described and the revision of all fossil species of these genera is given (see also Table 1).
Table 1. List of fossil Cenozoic species of Anguinae, their geological age and type localities. The species are listed in the same sequence as described in the text.

Order SQUAMATA Oppel, Reference Oppel1811
Infraorder ANGUIMORPHA Fürbringer, Reference Fürbringer1900
Family ANGUIDAE Gray, Reference Gray1825
Subfamily ANGUINAE Gray, Reference Gray1825
Genus Ophisaurus Daudin, Reference Daudin1803
Ophisaurus fejfari Klembara, Reference Klembara1979
Figures 1, 2a, b
1979 Ophisaurus fejfari Klembara, p. 164, pl. 1, fig. 1, pl. 2, fig. 1.
1981 Ophisaurus fejfari Klembara, p. 125.
1981 Ophisaurus spinari Klembara, p. 128, specimen DP FNSP 1023.
Holotype. Parietal DP FNSP 1013 (Klembara, Reference Klembara1979, pl. 1, fig. 1, pl. 2, fig. 1); Fig. 1a, b. Dolnice (near the city of Cheb, west Bohemia, Czech Republic), Early Miocene (Ottnangian), middle Orleanian, zone MN 4 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993).

Figure 1. Photographs of parietals of Ophisaurus fejfari in dorsal (a, c) and ventral (b, d) views. (a, b) DP FNSP 1013, holotype; (c, d) DP FNSP 1501. Numbers 1–3 designate distinguished features of O. fejfari discussed in the text.
Diagnosis. The Ophisaurus fejfari parietal is distinguished from that of other extinct Ophisaurus species described on the basis of parietals and all extant species of the genus in the following features and their combinations: (1) a distinct divergence of bases of supratemporal processes (the longitudinal line of the basal section of the supratemporal ridge of the supratemporal process continues to the frontal tab anteriorly); (2) the arcuate edge is wedge- or arch-shaped and its lateral section runs posteriorly close to the medial margin of the supratemporal process base; and (3) the parietal cranial crest is almost straight, except for its anteriormost section.
Referred specimens, localities and horizon. Parietals DP FNSP 1501, DP FNSP 1023, Dolnice (near the city of Cheb), Early Miocene (Ottnangian), middle Orleanian, zone MN 4 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993). Parietal BSPG 1997 XVI 466 (Fig. 2a, b), Griesbeckerzell 1a (Bavaria, south Germany), zone MN 5/6 (according to Böhme, Reference Böhme2002; Abdul-Aziz et al. Reference Abdul-Aziz, Böhme, Rocholl, Zwing, Prieto, Wijbrans, Heissig and Bachtadse2008).

Figure 2. (a, b) BSPG 1997XVI 466, photographs of parietals of Ophisaurus fejfari in dorsal (a) and ventral (b) views. (c, d) UF 57121, photographs of parietal of Ophisaurus attenuatus in dorsal (c) and ventral (d) views.
Description. The lateral margins of the parietal table as well as its ornamented surface converge posteromedially and the parietal is constricted at the level of the transition of the parietal table and supratemporal processes (Figs 1, 2a, b). The ornamented surface is longer than it is wide. The ornamentation consists of densely arranged distinct ridges, grooves and pits occupying most of the ornamented surface. Short ridges and grooves are present only at the periphery of the ornamented surface. The anterior end of the interparietal sulcus lies in the anterolateral corner of the ornamented surface. The ornamented surface consists of unpaired occipital and interparietal and paired lateral shields. The occipital shield has elliptical to quadrangular shape (Figs 1a, c, 2a). The pineal foramen lies immediately posterior to the mid-length of the ornamented surface. Measured in the median plane, the smooth area is slightly anteroposteriorly longer than the length of the occipital shield. The anterolateral process is well developed. The arcuate edge is wedge-shaped and runs posteriorly close to the medial margin of the supratemporal process base. The preserved basal portions of the supratemporal processes are massive and indicate a broad divergence of the supratemporal processes.
The parietal cranial crest forms a distinct straight wall, only its anterior section is low and turned anteromedially (Figs 1b, 2b). The frontal tab is moderately developed. The posterior tip at the junction of the posteriormost sections of the parietal cranial crest and juxtafoveal crest merges with the medial margin of the supratemporal process via a short, high and sharp postfoveal crest. The preserved anterior section of the ventrolateral ridge of the supratemporal process is massive and coincides with the lateral margin of the supratemporal process. The anterior end of the ventrolateral ridge lies at the level slightly anterior to the posteromedian margin of the floor of the parietal fossa. In the holotype parietal, there is a small, oval pit positioned posteriorly to the pineal foramen. The posterior margin of the floor of the parietal fossa is wedge-shaped. The position of the posteromedian margin of the parietal table varies slightly relative to the length of the juxtafoveal crest (cf. Fig. 1b, d).
The holotype (Fig. 1a, b) and DP FNSP 1501 (Fig. 1c, d) are of similar size. The parietal BSPG 1997 XVI 466 (Fig. 2a, b) is much larger than the other three parietals. The parietal BSPG 1997 XVI 466 differs from the holotype and DP FNSP 1501 in the following features: (1) large anterolateral processes of parietal table; (2) ornamentation of the anterolateral corner of the ornamented shield extends to the dorsal surface of the anterolateral process of the parietal table; and (3) quadrangular shape of occipital ornamented shield showing a slight variability in size and shape of the occipital shield. Probably the first two differences are connected with the larger size of the BSPG 1997 XVI 466 parietal. Unless better material is at our disposal, the parietal BSPG 1997 XVI is therefore tentatively assigned to Ophisaurus fejfari.
Comparisons. The parietal of Ophisaurus fajfari is characterized by these three features (the following numbers correspond to 1–3 in Fig. 1):
-
(1) A distinct divergence of bases of supratemporal processes (the longitudinal line of the basal section of the supratemporal ridge of the supratemporal process continues to the frontal tab anteriorly). In all other fossil species of Ophisaurus, the longitudinal line of the basal section of the supratemporal ridge of the supratemporal process continues to or lies close to the median point of the anterior margin of the parietal (cf. Figs 1b and 3b, 5d, 8b).
-
(2) The arcuate edge is wedge- or arch-shaped and its lateral section runs posteriorly close to the medial margin of the supratemporal process base.
-
(3) The parietal cranial crest is almost straight, except for its anteriormost section.

Figure 3. (a–d) Photographs of parietals of Ophisaurus spinari in dorsal (a, c) and ventral (b, d) views. (a, b) DP FNSP 2015, holotype; (c, d) DP FNSP 1025. (e, f) DE 35, photographs of parietals of Ophisaurus ventralis in dorsal (e) and ventral (f) views. Numbers 1–3 designate distinguished features of O. spinari discussed in the text.
The general shape of the parietal described as Dopasia coderetensis (Augé, Reference Augé2005, fig. 165a, b) from the Late Oligocene of France is somewhat similar to that of Ophisaurus fejfari. However, the parietal from France is poorly preserved and does not allow detailed comparisons.
Among the extant species of Ophisaurus, the O. fejfari parietal is most similar to that of O. attenuatus (Fig. 2c, d). The parietal of O. attenuatus shares three features with that of O. fejfari: (1) the lateral margins of the parietal table converge posteriorly and a distinct constriction is present at the transition of the parietal table and bases of supratemporal processes; (2) there is a distinct divergence of the bases of supratemporal processes, the same as is in O. fejfari (cf. Figs 1b, 2d); and (3) besides its anteriormost section, the parietal cranial crest is almost straight. In contrast, the parietal of O. attenuatus lacks the postfoveal crest and the anterior end of the interparietal sulcus lies medially to the anterolateral corner of the ornamented surface.
Ophisaurus spinari Klembara, Reference Klembara1979
Figures 3a–d, 4
1979 Ophisaurus spinari Klembara, p. 164, pl. 1, fig. 2, pl. 2, fig. 2.
1981 Ophisaurus spinari Klembara, p. 128, specimen DP FNSP 1029.
2011 Dopasia sp. Delfino, Bailon and Pitruzzella, p. 368, fig. 5C, D.
2015 Ophisaurus spinari Čerňanský, Rage & Klembara, p. 107, fig. 8A, B.
Reference Klembara2015 Ophisaurus spinari Klembara, p. 178, fig. 3A, B.
Holotype. Parietal DP FNSP 2015 (Klembara, Reference Klembara1979, pl. 1, fig. 2, pl. 2, fig. 2); Fig. 3a, b. Dolnice (near the city of Cheb), Early Miocene (Ottnangian), middle Orleanian, zone MN 4 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993).
Diagnosis. Ophisaurus spinari parietal differs from all extinct species of Ophisaurus described on the basis of parietals and extant species of Ophisaurus in the following features and combination of features: (1) the ornamented surface is quadrangular, longer than it is wide and its width is approximately the same along its entire anteroposterior length; (2) the occipital ornamented shield is triangular with an almost straight posterior margin; and (3) the posterior section of the parietal cranial crest has a concave course.
Referred specimens, locality and horizon. Parietals DP FNSP 1014, DP FNSP 1019, DP FNSP 1021, 1022, DP FNSP 1025 (Klembara, Reference Klembara1981, fig. 1; pl. 2, fig. 1), DP FNSP 1028, Dolnice (near the city of Cheb), Early Miocene (Ottnangian), middle Orleanian, zone MN 4 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993). Parietals NMA-2015-32/2196, Petersbuch 39 III, zone MN 5 (Bavaria) (Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010) (Fig. 4a, b); NMA-2015-36/2196, Petersbuch 62, zone MN 3 (Bavaria) (Abdul-Aziz et al. Reference Abdul-Aziz, Böhme, Rocholl, Zwing, Prieto, Wijbrans, Heissig and Bachtadse2008) (Fig. 4b, c).

Figure 4. Photographs of parietals of Ophisaurus spinari in dorsal (a, c) and ventral (b, d) views. (a, b) NMA-2015-32/2196; (c, d) NMA-2015-36/2196.
Description. The parietal table is covered by a quandrangular ornamented surface (Figs 3a, c, 4a, c). This ornamentation is densely arranged. The elongated grooves and ridges are present only at the periphery of the ornamented surface. The anterior end of the interparietal sulcus lies in the anterolateral corner of the ornamented surface. The occipital shield is triangular. Its posterior margin is almost straight. The pineal foramen lies immediately posterior to the mid-length of the ornamented surface. The length of the smooth area measured in the median plane approximates that of the occipital ornamented shield. The arcuate edge is arch-shaped and the anterolateral process is moderately developed.
The parietal cranial crest is deep and distinctly developed in its entire course (Figs 3b, d, 4b, d). Its anterior and middle sections have a convex course; only its posterior section is typically more or less distinctly concave. Laterally, the parietal cranial crest reaches the lateral margin of the parietal table (measured at the level of the pineal foramen). The frontal tab is distinct. The anterior end of the ventrolateral ridge of the supratemporal process lies anterior to the posteromedian margin of the floor of the parietal fossa. The ventrolateral ridge coincides with the lateral margin of the supratemporal process. The supratemporal articulation extends anteriorly to the level of the posteromedian margin of the parietal table (Fig. 4b). The paroccipital process of the opisthotic articulation has an elongated shape (Fig. 4b).
Comparisons. Three features characterize the parietal of Ophisaurus spinari (the following numbers correspond to 1–3 in Fig. 3):
-
(1) The ornamented surface of the parietal is quadrangular, longer than it is wide and its width is approximately the same along its entire anteroposterior length. A similar shape of the ornamented surface is present in extant Ophisaurus ventralis (Fig. 3e). However, in O. ventralis the ornamentation is indistinctly developed. The ornamented surface of the adult O. ventralis parietal consists of very indistinct and almost flat tubercles; between them, many small pits are present (e.g. CM 1411, DE 35). Longer ridges and grooves at the periphery of the ornamented shield are also very low and indistinct.
-
(2) The occipital ornamented shield is triangular with an almost straight posterior margin. Ophisaurus fejfari (Fig. 1c) and O. holeci (Figs 7a, c, 8a, c further below and Klembara, Reference Klembara2015) have a roughly similar shape of the occipital ornamented shield. However, in contrast to O. fejfari, the parietal of O. spinari (i) lacks a distinct constriction between the bases of the supratemporal processes and the parietal table; (ii) lacks distinct divergence of the supratemporal processes bases (as in O. fejfari, in which the longitudinal line of the basal section of the supratemporal ridge of the supratemporal process is continued with the frontal tab anteriorly, Fig. 1b); in all other fossil species of Ophisaurus, the longitudinal line of the basal section of the supratemporal ridge is continuous with or lies close to the median point of the anterior margin of the parietal, e.g. Fig. 3b); and (iii) its cranial crest is not straight in its middle and posterior sections. In contrast to O. holeci, in the parietal of O. spinari (i) the anterior end of the interparietal sulcus lies in the anterolateral corner of the ornamented surface; (ii) the ventrolateral ridge of the supratemporal process joins the parietal cranial crest anterior to the posteromedian margin of the floor of the parietal fossa; and (iii) the posterior section of the parietal cranial crest has a concave course.
-
(3) The posterior section of the parietal cranial crest has a concave course. This is in contrast to the straight course of this section of the parietal cranial crest in other species of Ophisaurus.
Delfino, Bailon & Pitruzzella (Reference Delfino, Bailon and Pitruzzella2011, fig. 5C, D) described a right portion of the parietal from the Late Pliocene of Italy as Dopasia sp. The parietal cranial crest of this parietal is well preserved and its posterior section is clearly concave. This feature is present only in Ophisaurus spinari (Figs 3b, d, 4b, d). The posterolateral portion of the ornamented surface of the parietal from Italy is rectangular, indicating that the posterior width of the ornamented surface is about the same as that of the anterior portion of the ornamented surface. This is also typical for O. spinari. Thus, the parietal from Italy described as Dopasia sp. (Delfino, Bailon & Pitruzzella, Reference Delfino, Bailon and Pitruzzella2011) is assigned to O. spinari here and it represents the youngest record of this species.

Figure 5. Photographs of parietals of Ophisaurus robustus in dorsal (a, c) and ventral (b, d) views. (a, b) DP FNSP 1018, holotype; (c, d) DP FNSP 1020. Numbers 1–4 designate distinguished features of O. robustus discussed in the text.
Special attention is required for the parietal attributed recently to Ophisaurus spinari from the late Middle Miocene of Austria (Böhme & Vasilyan, Reference Böhme and Vasilyan2014). The parietal from Austria exhibits features which are not preserved in similarly sized parietals of O. spinari (Figs 3a–d, 4; Klembara, Reference Klembara1979, 1981, Reference Klembara2015; Čerňanský, Rage & Klembara, Reference Čerňanský, Rage and Klembara2015). These features are:
-
(1) The ornamental surface of parietal gradually widens anteriorly. In O. spinari, the surface is rectangular and never exhibits distinct gradual widening of its anterior portion.
-
(2) The parietal table is distinctly anteriorly widened bilaterally. This widening is absent in O. spinari and all other Ophisaurus, Anguis and Pseudopus species.
-
(3) The anteroposterior length of the smooth area measured in the medial plane is less than the anteroposterior length of the occipital ornamented shield. This condition is typical for Anguis (Fig. 9c further below; see also Klembara, Reference Klembara1979, Reference Klembara2015).
-
(4) The parietal cranial crests diverge distinctly anteriorly and the anteriormost section of this crest runs laterally. In Ophisaurus spinari, the anteriormost section of this crest turns distinctly medially. Such course of the anterior section of the parietal crest as present in Austrian parietal is never observed in Ophisaurus, Anguis and Pseudopus.
-
(5) The presence of a muscular surface (facies muscularis). Although the lateral margins of the parietal plate of the Austrian parietal are slightly damaged, the presence of a narrow muscular surface is clearly distinguishable. The muscular surface is a typical feature of Pseudopus (Klembara, Reference Klembara1979, 1981, Reference Klembara2012, Reference Klembara2015; Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010).
-
(6) The supratemporal process has a smooth ventrolateral surface continuous with the muscular surface of the parietal table, and the ventrolateral ridge of the supratemporal process runs medial to the lateral margin of the supratemporal process. This ventrolateral surface/muscular surface continuity is typical for Pseudopus (Klembara, Reference Klembara1979, Reference Klembara1981, Reference Klembara2012, Reference Klembara2015; Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010).
The above analysis confirms that the parietal from the Austrian locality Gratkorn (Böhme & Vasilyan, Reference Böhme and Vasilyan2014) does not represent the Ophisaurus spinari species. The parietal from Austria exhibits a mixture of features of Ophisaurus, Anguis and Pseudopus, and two characters (2 and 4 above) are not exhibited by the parietals of any known anguine (Ophisaurus, Anguis and Pseudopus). A complete revision of the parietal from the late Middle Miocene of Austria (Böhme & Vasilyan, Reference Böhme and Vasilyan2014) is required.
Of all Recent Ophisaurus species, the parietal of O. spinari is most similar to that of O. ventralis (Fig. 3e, f). The parietal of O. ventralis shares two similar characters with that of O. spinari: (1) the ornamented surface is quadrangular, longer than it is wide and is of approximately equal width along its length; and (2) the occipital ornamented shield is triangular, with an almost straight posterior margin.
Ophisaurus robustus (Klembara, Reference Klembara1979)
Figures 5, 6
1979 Anguis robustus Klembara, p. 165, pl. 3, fig. 1, pl. 4, fig. 1.
1981 Ophisaurus spinari Klembara, p. 128, specimen DP FNSP 1020.
1981 Ophisaurus (?) robustus (cf. Anguis) Klembara, p. 130.
2013 Ophisaurus cf. spinari Venczel & Hír, p. 140, text-fig. 100d.
Holotype. Parietal DP FNSP 1018 (Klembara, Reference Klembara1979, pl. 3, fig. 1, pl. 4, fig. 1); Fig. 5a, b. Dolnice (near the city of Cheb), Early Miocene (Ottnangian), middle Orleanian, zone MN 4 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993).
Diagnosis. The parietal of Ophisaurus robustus differs from all extinct Ophisaurus species described on the basis of parietals and extant species of Ophisaurus in the following features and their combinations: (1) the ornamented shield is equal in length and width or wider than it is long; (2) the lateral margins of the lateral ornamented shields are distinctly rounded; (3) a small, elliptical occipital ornamented shield is present; and (4) the supratemporal articulation reaches the level or slightly anterior to the level of the posteromedian margin of the parietal table.
Referred specimens, locality and horizon. Parietals DP FNSP 1020, Dolnice (near the city of Cheb), Early Miocene (Ottnangian), middle Orleanian, zone MN 4 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993). Parietal NMA-2015-37/2196, Petersbuch 62, zone MN 3 (Bavaria) (Abdul-Aziz et al. Reference Abdul-Aziz, Böhme, Rocholl, Zwing, Prieto, Wijbrans, Heissig and Bachtadse2008) (Fig. 6).

Figure 6. Photographs of parietal NMA-2015-37/2196 of Ophisaurus robustus in dorsal (a) and ventral (b) views.
Description. The lateral margins of the ornamented surface of the parietal converge posteromedially and are distinctly rounded (Figs 5a, c, 6a). A more or less wide uncovered surface is present between the lateral margins of the ornamented surface and the parietal table. The ornamented surface is equal in length and width or wider than it is long. The anterior end of the interparietal sulcus lies in the anterolateral corner of the ornamented surface. The occipital shield is oval with the long axis running transversely. The ornamentation consists of pits, short grooves and faint ridges radiating from the centre of the ornamented surface. The position of the pineal foramen varies; it lies slightly anterior or posterior to the mid-length of the ornamented surface. The anterolateral process is moderately developed. The arcuate edge is arch-like.
The parietal cranial crest is well developed in its entire arch-like course. The postfoveal crest is absent. The ventrolateral ridge of the supratemporal process coincides with the lateral margin of the supratemporal process. The anterior end of the ventrolateral ridge of the supratemporal process joins the parietal cranial crest anterior to the level of the posteromedian margin of the parietal fossa floor. A small pit is present posterior to the pineal foramen. The frontal tab is moderately developed.
Comparisons. The Ophisaurus robustus parietal was originally described as Anguis robustus because of similarity of the parietals in both taxa, although that of O. robustus is more robustly constructed (Klembara, Reference Klembara1979). In addition, in Anguis the anteroposterior extent of the parietal occipital ornamented shield is greater than the anteroposterior length of the smooth area, when measured in the median plane (Fig. 9c and see below). Although there is a slight individual variability of this feature in A. fragilis, the smooth area is very short in most specimens of this species. This condition, typical for adult Anguis, is present in the juveniles of Ophisaurus (e.g. O. attenuatus, Klembara & Green, Reference Klembara and Green2010, fig. 3A, B).
The following features distinguish the Ophisaurus robustus parietal from all extinct Ophisaurus species described on the basis of parietals and Ophisaurus extant species (the following numbers correspond to 1–4 in Fig. 5):
-
(1) The ornamented surface of parietal is equal in length and width or wider than it is long. This is in contrast to the always more or less longer than wide ornamented surface in adult specimens of other Ophisaurus species.
-
(2) The lateral margins of the lateral ornamented shield are distinctly rounded.
-
(3) A small, elliptical occipital ornamented shield. A shield of similar morphology is present in Anguis fragilis (e.g. Klembara, Reference Klembara1979, pl. 3, fig. 2). However, the lateral margin of the lateral ornamented shield is straight in A. fragilis (Fig. 9c further below).
-
(4) The supratemporal articulation lies at or slightly anterior to the level of posteromedian margin of the parietal table.
The parietal NMA-2015-37/2196 (Fig. 6) represents the first record of Ophisaurus robustus in Germany.
The morphology of the ornamented surface of Ophisaurus robustus parietal is similar to that of small specimens of extant Ophisaurus; in them, the ornamented surface is anteriorly and posteriorly rounded (e.g. O. attenuatus, Klembara & Green, Reference Klembara and Green2010, fig. 3A). However, in small O. attenuatus the parietal is less robust, the smooth area is much shorter relative to the length of the occipital ornamented shield and the parietal cranial crest and juxtafoveal crest are very weakly developed relative to the adult specimen (Klembara & Green, Reference Klembara and Green2010, fig. 3). In contrast, known O. robustus parietals are well developed with an equally long anteroposterior smooth area and occipital ornamented shield measured in the median plane. All these features confirm that the O. robustus parietals are adult specimens.
Hír et al. (Reference Hír, Kókay, Venczel, Gál and Kessler2001) and Venczel & Hír (Reference Venczel and Hír2013, text-fig. 100d) described a well-preserved middle portion of the parietal from the late Middle Miocene of Hungary as Ophisaurus cf. spinari. This parietal differs from that of O. spinari (Fig. 3a, c) by posteromedially converging and posterolaterally rounded lateral margins of the ornamented shield. These conditions are typical for the Ophisaurus robustus parietal (Klembara, Reference Klembara1979) and Figures 5a, c, 6a, the ornamented surface of which has equal width and length or is slightly wider than it is long. Unfortunately, the anteriormost portion of the parietal from Hungary is missing, so it is impossible to measure its length. Besides this, the posterior section of the parietal cranial crest of the parietal of O. spinari is typically concave. In contrast, it is straight in the figured parietal from Hungary (Venczel & Hír, Reference Venczel and Hír2013, text-fig. 100d). Thus, this parietal from Hungary is assigned to O. robustus here and it represents the youngest record of this species.
Ophisaurus holeci Klembara, Reference Klembara2015
Figures 7, 8
1981 Ophisaurus spinari Klembara, p. 128, specimens DP FNSP 1026, DP FNSP 1027.
2015 Ophisaurus holeci Klembara, p. 172, figs 1A–C, 2.
Holotype. Parietal NMP Pb 02027 (Klembara, Reference Klembara2015, fig. 2A, B). Opencast mine Merkur-North (near the city of Chomutov, north Bohemia, Czech Republic), North Bohemian Basin, earlier part of the Early Miocene (Eggenburgian), lower Orleanian, zone MN 3 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993).
Diagnosis. The parietal of Ophisaurus holeci differs from all extinct Ophisaurus species described on the basis of parietals and extant Ophisaurus species in the following features and their combinations (for details, see Klembara, Reference Klembara2015): (1) the parietal and frontal ornamentation has short, pronounced anastomosing ridges and grooves and distinct pits; (2) the anterior end of the interparietal sulcus lies medial to the anterolateral corner of the ornamented surface; (3) the anterior end of the ventrolateral ridge of the supratemporal process joins the parietal cranial crest at or slightly posterior to the posteromedian margin of the parietal fossa floor; (4) the posterior section of parietal cranial crest is very low, especially at its junction with the ventrolateral ridge of the supratemporal process; (5) the supratemporal process is straight; (6) the base of the supratemporal process is mediolaterally narrow; (7) a short postfoveal crest is present; and (8) a long anterior section of parietal cranial crest is more or less distinctly concave or straight.
Referred specimens, locality and horizon. Parietals DP FNSP 1026 (Fig. 7a, b), ?DP FNSP 1027, ?DP FNSP 1029, Dolnice (near the city of Cheb), Early Miocene (Ottnangian), middle Orleanian, zone MN 4 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993). Parietal NMA-2007-69/2035, Petersbuch 39-III (Bavaria), zone MN 5 (Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010) (Fig. 7c, d). Parietals NMA-2015-33/2196 (Fig. 8a, b) and NMA-2015-35/2196 (Fig. 8c, d), Petersbuch 62, zone MN 3 (Bavaria) (Abdul-Aziz et al. Reference Abdul-Aziz, Böhme, Rocholl, Zwing, Prieto, Wijbrans, Heissig and Bachtadse2008).
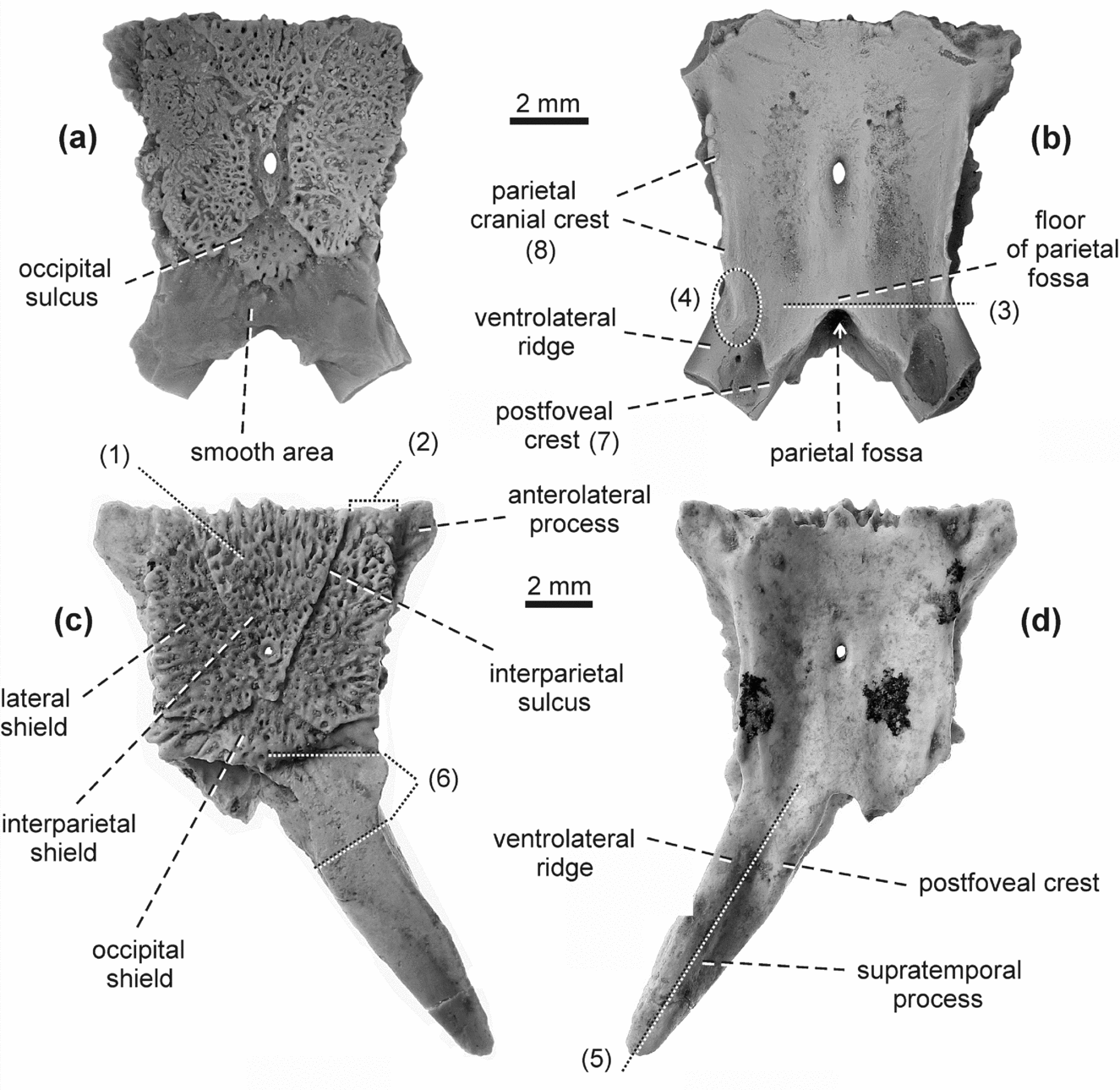
Figure 7. Photographs of parietals of Ophisaurus holeci in dorsal (a, c) and ventral (b, d) views. (a, b) DP FNSP 1026; (c, d) NMA-2007-69/2035. Numbers 1–8 designate distinguished features of O. holeci discussed in the text.

Figure 8. Photographs of parietals of Ophisaurus holeci in dorsal (a, c) and ventral (b, d) views. (a, b) NMA-2015-33/2196; (c, d) NMA-2015-35/2196.
Description. The parietal DP FNSP 1026 (Fig. 7a, b) and those from Germany (Fig. 8) are similar in size and morphology to those described recently (Klembara, Reference Klembara2015). Specimen NMA-2007-69/2035 (Fig. 7c, d) from Germany represents the largest parietal of Ophisaurus holeci. The ornamented surface is slightly longer than it is wide and its lateral margins converge slightly posteromedially. The anterior end of the interparietal sulcus lies medial to the anterolateral corner of the ornamented surface. The occipital ornamented shield is a more or less wide triangle with a slightly convex posterior margin. The anterolateral process is distinctly developed. A small pineal foramen lies immediately posterior to the mid-length of the ornamented shield. The right supratemporal process is completely preserved in NMA-2007-69/2035 (Fig. 7c, d) and is typically straight. The bases of the supratemporal processes are typically mediolaterally narrow relative to those of other species of Ophisaurus.
The posterior section of the parietal cranial crest is very indistinct, especially at the junction with the anterior end of the ventrolateral ridge of the supratemporal process. This junction lies at the level of or slightly posterior to the posteromedian margin of the parietal fossa floor. The ventrolateral ridge of the supratemporal process coincides with the lateral margin of the supratemporal process. A short postfoveal crest is present.
Comparisons. The parietal of Ophisaurus holeci is distinguished by the following features and their combinations (the following numbers correspond to 1–8 in Fig. 7):
-
(1) The parietal and frontal ornamentation has short, pronounced anastomosing ridges and grooves and distinct pits.
-
(2) The anterior end of the interparietal sulcus lies medial to the anterolateral corner of the ornamented surface. In O. fejfari, O. spinari and O. robustus, the anterior end of the interparietal sulcus lies in the anterolateral corner of the parietal ornamented shield. In the Recent O. attenuatus, the condition is almost the same as in O. holeci; however, O. holeci differs from O. attenuates in features 3–7 listed below.
-
(3) The anterior end of the ventrolateral ridge of the supratemporal process joins the parietal cranial crest at or slightly posterior to the posteromedian margin of the parietal fossa floor. In contrast, in all other species of Ophisaurus the anterior end of the ventrolateral ridge of the supratemporal process joins the parietal cranial crest anteriorly to the level of the posteromedian margin of the parietal fossa floor.
-
(4) The posterior section of the parietal cranial crest is very low, especially at its junction with the ventrolateral ridge of the supratemporal process. In all other species of Ophisaurus, the parietal cranial crest is a distinct ridge at its junction with the ventrolateral ridge of the supratemporal process.
-
(5) The supratemporal process is straight. In other species of Ophisaurus there is always an angle between the base and the posterior section of the supratemporal process.
-
(6) The base of the supratemporal process is mediolaterally narrow (the width of the base is less than half the width of the parietal table measured at the level of maximum constriction of the parietal table – at the level of the posterior portion of the occipital ornamented shield).
-
(7) A short postfoveal crest is present (a similar crest is present also in O. fejfari; however, the parietal of O. fejfari lacks characteristics 1–6 above).
-
(8) A long anterior section of the parietal cranial crest is more or less distinctly concave (see also Klembara, Reference Klembara2015, fig. 2B, D). In Ophisaurus fejfari, this section of the crest is straight and in O. spinari and O. robustus is convex.
The parietal NMA-2007-69/2035 (Fig. 7c, d) from Germany differs from smaller specimens by its slightly larger anterolateral process and wider occipital ornamented shield. This parietal represents the largest specimen of O. holeci, and together with two other parietals from Germany (Fig. 8) represents the first record of this species outside the Czech Republic.
The parietal of Ophisaurus holeci is similar to those of Ophisaurus roqueprunensis from the Early to Late Oligocene of France and Belgium (Augé & Smith, Reference Augé and Smith2009) and Ophisaurus sp. from the Late Eocene of the Hampshire Basin described by Klembara & Green (Reference Klembara and Green2010) in two features: (1) the anterior end of the ventrolateral ridge of the supratemporal process joins the posterior section of the parietal cranial crest slightly posteriorly to the level of the posteromedian margin of the parietal fossa floor; and (2) the posterior section of the parietal cranial crest is very low, especially at its junction with the posterolateral ridge of the supratemporal process. These two features are recognized only in these three taxa; the conditions in Ophisauriscus quadrupes from the Middle Eocene of Germany (Sullivan, Keller & Habersetzer, Reference Sullivan, Keller and Habersetzer1999) require revision in this respect. On the other hand, the parietals of Ophisaurus roqueprunensis and Ophisaurus sp. differ from the parietal of O. holeci by the two following characters: (1) the smooth area of the parietal of O. roqueprunensis and Ophisaurus sp. is slightly longer than the length of the occipital ornamented shield, when measured in the median plane; and (2) the postfoveal crest is absent in O. roqueprunensis and Ophisaurus sp. (Augé & Smith, Reference Augé and Smith2009; Klembara & Green, Reference Klembara and Green2010). These two latter features need to be revised also in Ophisauriscus quadrupes.
Ophisaurus acuminatus Jörg, Reference Jörg1965
Holotype. Partial skull SMNK-PAL.8561; Heweneg/Hegau (near city Öhningen), Germany.
Diagnosis. After Jörg (Reference Jörg1965).
Locality and horizon. Heweneg/Hegau (near city Öhningen), Germany; lower Late Miocene, zone MN 9 – MN 10.
Remarks. The general morphology of preserved cranial elements of Ophisaurus acuminatus corresponds to that in fossil or extant species of Ophisaurus. The maxillary and dentary teeth of O. acuminatus are most similar to those in Anguine morphotype 1 from the Early Miocene locality Merkur (MN 3; the Czech Republic) (Klembara, Reference Klembara2015). The teeth are recurved posteriorly and their apices bear anterior and posterior cutting edges. Although the parietal bone is missing in this species, the ornamented surfaces of frontal and nasal bones are preserved. Comparison of equal-sized bones of O. acuminatus with those of all other species of Ophisaurus described here shows that the ornamentation of the preserved skull roof bones in O. acuminatus is much more robustly developed.
Ophisaurus canadensis Holman, Reference Holman1970
Holotype. Dorsal vertebra ROM 7705, Upper Miocene, Kleinfelder Farm locality of the Wood Mountain Formation near Rockglen, Saskatchewan, Canada.
Diagnosis. After Holman (Reference Holman1970).
Locality and horizon. Kleinfelder Farm locality of the Wood Mountain Formation near Rockglen, Saskatchewan, Canada. Upper Miocene.
Remarks. According to Holman (Reference Holman1970), the dorsal vertebra is most similar to those of Recent North American Ophisaurus ventralis and O. attenuatus. However, it differs in having a rounded anterior portion of neural arch which extends almost as far as the anterior edges of prezygapophyses; prezygapophyses are not so dorsally extended (see Estes, Reference Estes and Wellenhofer1983). While additional material is required to confirm feature validity and justify the species existence, this dorsal vertebra documents Ophisaurus presence in the Late Miocene of North America and its distinction as the oldest currently recognized Ophisaurus record in North America.
Ophisaurus roqueprunensis Augé, Reference Augé1992
1992 Ophisaurus roqueprunensis Augé, p. 160, fig. 1A, B.
2005 Dopasia roqueprunensis Augé, p. 234.
2009 Dopasia roqueprunensis Augé & Smith, p. 152.
2016 Ophisaurus roqueprunensis Čerňanský, Klembara & Müller, p. 23.
Holotype. Left dentary IRScNB No. R120, Middle Oligocene (zone MP 23), Roqueprune, Phosphorites du Quercy, France.
Diagnosis. After Augé (Reference Augé1992); emended diagnosis (Augé & Smith, Reference Augé and Smith2009).
Localities and horizons. Early Oligocene, Boutersem, Hoogbutsel (MP 21), Belgium; Early Oligocene – Late Oligocene, Phosphorites du Quercy (MP 23 – MP 28), Roqueprune (MP 23), Belgarric (MP 25), Rigal-Jouet (MP 25), Pech du Fraysse (MP 28), France.
Remarks. This species is present from the Early to Late Oligocene of France and Belgium and is based on dentaries, maxillae, one incomplete parietal, dorsal and caudal vertebrae and numerous osteoderms (Augé & Smith Reference Augé and Smith2009).
Genus Anguis Linnaeus, 1758
Anguis rarus sp. nov.
Figure 9a, b
Holotype. Parietal NMA-2015-34/2196, Petersbuch 62 (Bavaria, south Germany), zone MN 3 (Fig. 9a, b).

Figure 9. (a, b) NMA 2015-34/2196, photographs of parietal of Anguis rarus sp. nov. in dorsal (a) and ventral (b) views; numbers 1–3 designate distinguished features of A. rarus discussed in the text. (c, d) DE 19, photographs of parietal of Anguis fragilis in dorsal (c) and ventral (d) views.
Diagnosis. The parietal of Anguis rarus sp. nov. differs from the parietal of A. fragilis by the following combination of features: (1) more diverging bases of supratemporal processes; (2) the anterior end of the interparietal sulcus lies in the anterolateral corner of the parietal ornamented shield; and (3) the anterior end of the supratemporal articulation reaches anterior to the level of the posteromedian margin of the parietal table.
Derivation of name. From the Latin rarus – rare, indicating very rare record of Anguis in the Miocene.
Locality and horizon. Petersbuch 62 (Bavaria), zone MN 3 (Abdul-Aziz et al. Reference Abdul-Aziz, Böhme, Rocholl, Zwing, Prieto, Wijbrans, Heissig and Bachtadse2008) (Fig. 9a, b).
Description. The parietal is almost completely preserved; only the posterior tip of the right supratemporal process is broken (Fig. 9a). The parietal table is anteroposteriorly elongated and its lateral margins converge slightly posteromedially. The anterolateral processes are moderately developed. The ornamented surface is longer than it is wide and covers most of the surface of the parietal table. Ornamentation consists of low tubercles and short ridges, and deep pits with longer ridges and grooves present only on the anterolateral margin of the ornamented surface. The anterior end of the interparietal sulcus lies in the anterolateral corner of the ornamented surface. The lateral margin of the parietal ornamented shield reaches the lateral margin of the parietal table. The occipital ornamented shield is triangular, but its posterior margin is slightly rounded. The smooth area of the parietal table immediately posterior to the ornamented surface is anteroposteriorly shorter than the anteroposterior length of the occipital shield (measured in the median plane). The supratemporal processes extend posterolaterally. The arcuate crest forms a shallow arch and its posterior end gradually passes into the medial margin of the supratemporal process.
The parietal cranial crest forms a moderate convex arch and reaches the lateral margin of the parietal table (Fig. 9b). The anterior section of the ventrolateral ridge of the supratemporal process coincides with the lateral margin of the supratemporal process. The anterior end of the ventrolateral ridge joins the parietal cranial crest anterior to the level of the posteromedian margin of the parietal fossa floor. The anterior end of the supratemporal articulation reaches the level of the posteromedian margin of the parietal table.
Comparisons. Specimen NMA-2015-34/2196 is the only parietal of Anguis from the Miocene and together with the vertebrae (Klembara, Reference Klembara1981) unambiguously documents the presence of Anguis in this geological period.
As already indicated earlier (Klembara, Reference Klembara2015), the parietals of Anguis and Ophisaurus are very similar, but they differ in one feature: in Anguis, the anteroposterior length of the smooth area lying immediately posterior to the ornamented surface of the parietal table is distinctly shorter than the anteroposterior length of the occipital ornamented shield (when measured in the median plane). This is quite apparent in both the extant Anguis and the parietal described here (Fig. 9a, c). Although this feature is evident in distinguishing the parietals of Ophisaurus and Pseudopus on the one hand and Anguis on the other, there is a slight variability as for the anteroposterior length of the occipital ornamented shield and the length of the smooth area of the parietal table (measured in the median plane). In most specimens of A. fragilis, the occipital ornamented shield is two or sometimes three times longer than the smooth area (Fig. 9c). In all specimens of A. fragilis studied here, however, the occipital shield is always slightly longer than the smooth area. Occasionally, in specimens of some Ophisaurus species the conditions may be similar to those of Anguis. For example, in the parietal NMA-2015-33/2196 of O. holeci the occipital ornamented shield is longer than the smooth area. This condition is mostly due to a secondary fusion of a small more posteriorly lying osteoderm to the posterior margin of the occipital ornamented shield (Fig. 8a). However, taking into account other parietal characters for comparison it is evident that the parietal NMA-2015-33/2196 cannot represent the species of Anguis.
The comparisons below do not include Anguis polgardiensis from the Late Miocene of Hungary (Bolkay, Reference Bolkay1913). As analysed recently (Klembara, Reference Klembara2015), it is not possible to decide unambiguously whether the incomplete parietal of this species represents Anguis or Ophisaurus.
The parietal of Anguis rarus sp. nov. exhibits no autapomorphy. However, it differs from the parietal of A. fragilis by the following combination of features (the following numbers correspond to 1–3 in Fig. 9a, b):
-
(1) Strongly diverging bases of the supratemporal processes. This is the same as in Ophisaurus fejfari (cf. Figs 1b and 9b); however, contrary to A. rarus, O. fejfari exhibits the following characteristics: (i) the lateral margins of the parietal table converge posteriorly and a distinct constriction is present at the transition of the parietal table and the bases of the supratemporal processes; (ii) the parietal cranial crest is almost straight, except for its anteriormost section; and (iii) a short postfoveal crest is present.
-
(2) The anterior end of the interparietal sulcus lies in the anterolateral corner of the parietal ornamented shield. In Anguis fragilis, the anterior end of the interparietal sulcus lies slightly medial to the anterolateral corner of the parietal ornamented shield (Fig. 9c and Klembara, Reference Klembara1979, pl. 3, fig. 2). The same condition as that of A. rarus is present in several species of Ophisaurus (see above) and Pseudopus ahnikoviensis (Klembara, Reference Klembara2012); however, in A. rarus (and A. fragilis) the anteroposterior length of the occipital ornamented shield is about twice the anteroposterior length of the smooth area (measured in the median plane).
-
(3) The anterior end of the supratemporal articulation reaches anterior to the level of the posteromedian margin of the parietal table. A similar condition is present in Ophisaurus robustus; however, the ornamented shield of the parietal of O. robustus has distinctly rounded anterolateral and posterolateral corners. This feature is absent in Anguis rarus.
Młynarski (Reference Młynarski1962) reported the presence of jaw fragments, osteoderms and vertebrae of Anguis at several Pliocene and Pleistocene localities in Poland. He figured the left dentary from the Pliocene locality Węże I and designated it as Anguis cf. fragilis (Młynarski, Reference Młynarski1962, pl. 14, fig. 3). Unfortunately, he gives no detailed description, and the posterior portion of the dentary is not visible on the photograph of this bone. Thus, it is not visible whether the dentary has the surangular spine or not. This spine is present in Ophisaurus, but absent in Anguis (Klembara, Hain & Dobiašová, Reference Klembara, Hain and Dobiašová2014). However, the morphology of the robust teeth, the position of the splenial spine at the level of the third tooth from posterior and a much longer coronoid process than the surangular process of the dentary could indicate that this dentary may belong to Ophisaurus. A dentary of similar morphology, including teeth, was described by Venczel (Reference Venczel2006) from the Late Miocene (MN 13) of Hungary. Anyway, the revision of the dentary from Poland is needed.
Augé (Reference Augé2005) and Rage & Augé (Reference Rage and Augé2010) described one left dentary from the Middle Eocene (MP 14) of Lissieu (France) as Anguis sp. (figured in Augé, Reference Augé2005, fig. 161). From the point of the determination, this dentary exhibits the following important features: (1) sharp, pointed teeth strongly recurved distally and lingually; (2) absence of a surangular spine; and (3) splenial spine lying at the level of the second tooth from the posterior. These features strongly indicate that this dentary is from Anguis.
Augé (Reference Augé2005) also described a parietal from the Middle Eocene (MP 16) locality Le Bretou in France as Anguis sp. However, (1) the smooth area of this parietal is long anteroposteriorly and much longer than that of the occipital ornamented shield; and (2) the parietal does not have a distinct parietal notch. These characteristics reflect partial similarity of this Middle Eocene parietal to the Late Eocene Headonhillia parva (Klembara & Green, Reference Klembara and Green2010).
?Anguis polgardiensis Bolkay, Reference Bolkay1913
1983 Ophisaurus pannonicus Estes, p. 141, fig. 34A.
Holotype. Not designated. Polgárdi, Bányahegy, Late Miocene, Hungary.
Diagnosis. After Bolkay (Reference Bolkay1913).
Referred specimens, locality and horizon. All specimens (parietal, 2 maxillae, fragment of dentary) of this species were lost (personal information of Professor L. Kordos of the Hungarian Geological Museum in Budapest, Hungary). Polgárdi, Bányahegy, Late Miocene (MN 13), Hungary.
Remarks. This species is based on one incomplete parietal, two maxillae and a fragment of the dentary. The morphology of this species parietal was recently revised by Klembara (Reference Klembara2015) who concluded that it is impossible to decide now whether this parietal belongs to Anguis or Ophisaurus. The teeth of maxilla and dentary described by Bolkay (Reference Bolkay1913) are more robust than those of A. fragilis, thus indicating the Ophisaurus affinity (Venczel, Reference Venczel2006).
Anguis stammeri Brunner, Reference Brunner1954
Holotype. Right dentary (BSPG; number not assigned), Late Pleistocene, Fuchsloch, Siegmannsbrunn, Germany.
Diagnosis. After Brunner (Reference Brunner1954, 1957) and Estes (Reference Estes and Wellenhofer1983).
Locality and horizon. Upper Pleistocene, Fuchloch, Siegmannsbrunn, Germany.
Remarks. This species is based on the right dentary. According to Brunner (Reference Brunner1957), the robust teeth differ from those of the extant Anguis fragilis. According to Młynarski (Reference Młynarski1962), the dentary of A. stammeri belongs to Ophisaurus. However, the teeth of the specimen described as Anguis cf. fragilis by Młynarski (Reference Młynarski1962) are similar to those of A. stammeri. Revision of A. stammeri is therefore required.
Genus Pseudopus Pallas, 1775
Pseudopus sp.
Figure 10
1986a Ophisaurus spinari Klembara, p. 189, figs 1, 2, pl. 1.
Referred specimens, locality and horizon. Parietal NMA-2007-86/2035, Petersbuch 73 (Bavaria), zone MN 7/8 (Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010), Fig. 10a, b; parietal SNM PP 1393, Devínska Nová Ves (west Slovakia), zone MN 6 (Klembara, Reference Klembara1986 a), Fig. 10c, d.

Figure 10. Photographs (a, b) and drawings (c, d) of parietals of Pseudopus sp. in dorsal (a, c) and ventral (b, d) views. (a, b) NMA-2007-86/2035; (c, d) SNM PP 1393 (from Klembara, Reference Klembara1986 a, pl. 1).
Description. The parietal table and bases of the supratemporal processes are well preserved in both specimens (Fig. 10). The ornamented surface has equal length and width or is slightly longer than it is wide. Its lateral margins converge slightly posteromedially. The ornamented surface is eroded, but short ridges, grooves and pits are easily recognizable. Several short grooves are clear at the periphery of the ornamented surface. The anterior end of the interparietal sulcus lies slightly medial to the anterolateral corner of the ornamented surface. The ornamentation of the lateral shield slightly extends to the surface of the anterolateral process of the parietal table. In NMA-2007-86/2035, the pineal foramen lies slightly anterior to the mid-length of the ornamented surface (Fig. 10a), whereas in SNM PP 1393 the pineal foramen lies slightly posterior to the mid-length of the ornamented surface (Fig. 10c). The occipital shield is triangular with a slightly convex posterior margin. The arcuate edge forms a shallow arch. The anterolateral process is moderately developed.
The parietal cranial crest forms a high and sharp wall. The muscular surface is present. The ventrolateral ridge of the supratemporal process is well developed. It joins the parietal cranial crest anterior to the posteromedian margin of the parietal fossa floor. There is a small, shallow pit posterior to the pineal foramen. A well-developed postfoveal crest is present.
Comparisons. Both parietals are very similar to that of Pseudopus laurillardi, but they have less distinctly developed anterolateral processes of the parietal table than those in P. laurillardi (Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010) when compared with similar size specimens. Distinctly extending anterolateral processes of the parietal are also present in P. ahnikoviensis, but the parietal ornamentation is completely different in the latter species (Klembara, Reference Klembara2012). The poorly developed anterolateral processes of the parietal are present in the Early Miocene P. rugosus (Klembara, Reference Klembara2015; however, see below), but Pseudopus sp. differs from P. rugosus in two features: (1) the ornamentation of the parietal comprises short ridges and grooves that are not so densely arranged as those in the parietal of P. rugosus; and (2) in ventral view, the pineal foramen lies anterior to the mid-distance between the anterior margin of the parietal table and the posteromedian margin of the parietal fossa floor, when measured in the median plane.
The parietals of Pseudopus sp. have similar geological age and are contemporaneous with the latest documented records of P. laurillardi (Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010). The poorly developed anterolateral processes are the only feature distinguishing Pseudopus sp. and P. laurillardi, so we have not created a new specific name for this anguine pending availability of more complete material.
Pseudopus confertus sp. nov.
2015 Pseudopus rugosus Klembara, p. 179, fig. 4.
Holotype. Frontal NMP Pv 10028; Merkur-North opencast mine (near Chomutov city, north Bohemia, Czech Republic).
Diagnosis. After Klembara (Reference Klembara2015).
Localities and horizons. Merkur-North opencast mine (near Chomutov), the earliest part of the Early Miocene (Eggenburgian), lower Orleanian, zone MN 3 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993).
Derivation of name. From the Latin confertus – squeezed-in, indicating the densely arranged ridges and grooves on the ornamented surface of the frontal and parietal.
Remarks. Recently, Klembara (Reference Klembara2015) described a new species of Pseudopus, P. rugosus, from the Merkur locality. Although Troschel (Reference Troschel1860) previously applied this name to a snake-like reptile from the Late Oligocene locality of Rott bei Bonn (Germany), it was also similar to the one he described as Thoracophis rugosus from the same locality (see Rage & Augé, Reference Rage and Augé1993). Troschel (Reference Troschel1860) further noted that the new animal from Rott is more like the modern Pseudopus lizard than a snake. This was probably the reason to replace the generic name Thoracophis with Pseudopus. Fejérváry-Láng (1923) and Estes (Reference Estes and Wellenhofer1983) suggested that the name Pseudopus rugosus could be a senior synonym of Ophisaurus moguntinus (Boettger, Reference Boettger1875), that is, Pseudopus laurillardi (Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010). But the specimen is presumably lost and no description, depiction or photograph exists. There is therefore doubt about which animal Troschel named P. rugosus. Theoretically, the animal he referred to as Pseudopus rugosus may be rediscovered, proving his appellation valid, and therefore we have created the new name, Pseudopus confertus sp. nov., for the Merkur anguine (Klembara, Reference Klembara2015, p. 179, fig. 4).
Pseudopus ahnikoviensis Klembara, Reference Klembara2012
Holotype. Parietal NMP Pb 02026. Opencast mine Merkur-North (near Chomutov).
Diagnosis. Pseudopus ahnikoviensis is distinguished from all other species of Pseudopus by the following features and the combination of features: (1) the length of a sulcus between the frontal and frontoparietal ornamented shields equals the length of the sulcus between the frontal and interfrontal shields; (2) the ornamented surface of the parietal consisting of densely arranged very fine ridges, grooves, and pits; (3) the ventrolateral ridge of the supratemporal process of the parietal coincides with the lateral margin of the supratemporal process; (4) the presence of the surangular spine of dentary; (5) the apices of crowns of robust bulbous teeth smooth; and (6) the posterior margin of the anterior inferior alveolar foramen formed by a splenial. For the analysis of these characteristics, see Klembara (Reference Klembara2012).
Localities and horizons. Opencast mine Merkur-North (near Chomutov), North Bohemian Basin, earlier part of the Early Miocene (Eggenburgian), lower Orleanian, zone MN 3 (Fejfar & Kvaček, Reference Fejfar and Kvaček1993).
Pseudopus laurillardi (Lartet, Reference Lartet1851)
Neotype. Left dentary MNHN Sa 23604. Sansan, Gers, France.
Diagnosis. After Klembara, Böhme & Rummel (Reference Klembara, Böhme and Rummel2010).
Localities and horizons. Sansan, Gers, France. Astaracien, zone MN 6, Middle Miocene (Augé & Rage, Reference Augé and Rage2000, p. 277, fig. 9).
Pseudopus pannonicus (Kormos, Reference Kormos1911)
Neotype. Mandible (number not assigned) deposited in the Hungarian Geological Institute, Budapest, Hungary.
Diagnosis. After Kormos (Reference Kormos1911) and Fejérváry-Lángh (Reference Fejérváry-Lángh1923). As for lower jaw, see Klembara, Böhme & Rummel (Reference Klembara, Böhme and Rummel2010).
Localities and horizons. Polgárdi 2 (Hungary), Upper Miocene (Turolian), zone MN 13 (Kormos, Reference Kormos1911; Tempfer, Reference Tempfer2009).
Genus Ophisauromimus Čerňanský, Klembara & Müller, Reference Čerňanský, Klembara and Müller2016
Ophisauromimus coderetensis (Augé, Reference Augé2005)
1986 Ophisaurus B Augé, p. 127, fig. 52.
2005 Dopasia coderetensis Augé, p. 237, fig. 156.
2015 Dopasia s.l. sp. Rage & Augé, p. 33.
2016 Ophisauromimus coderetensis Čerňanský, Klembara & Müller, p. 23, fig. 14.
Holotype. Right dentary (UCBL No. 97684), Upper Oligocene (MP 30), Coderet, France.
Diagnosis. After Augé (Reference Augé2005).
Localities and horizons. Itardies, Lower Oligocene (MP 23) and Coderet, Upper Oligocene (MP 30), Phosphorites du Quercy, France.
Remarks. This species is based on dentaries, maxilla fragments, one parietal and a mandible posterior portion. The parietal attributed to this species is so poorly preserved that comparisons are impossible (Augé, Reference Augé2005, fig. 165a, b). Recently Rage & Augé (Reference Rage and Augé2015) have described an anguine as Dopasia s.l. sp. on the basis of one dentary and several trunk and caudal vertebrae from the Early Oligocene (MP 22) Valbro locality of France (Quercy).
Ophisauromimus frayssensis (Augé, Reference Augé2005)
2005 Dopasia frayssensis Augé, p. 235, fig. 154.
2016 Ophisauromimus frayssensis Čerňanský, Klembara & Müller, p. 22.
Holotype. Right dentary NMHN No. PFR11003. Pech-du Fraysse, Phosphorites du Quercy, Upper Oligocene (MP 28), France.
Diagnosis. After Augé (Reference Augé2005) and Čerňanský, Klembara & Müller (Reference Čerňanský, Klembara and Müller2016).
Localities and horizons. Pech-du Fraysse and Pech Desse, Upper Oligocene (MP 28), Phosphorites du Quercy, France.
Remarks. This species is based on dentaries and dorsal vertebrae (Augé Reference Augé2005).
Genus Ragesaurus Bailon & Augé, Reference Bailon and Augé2012
Ragesaurus medasensis Bailon & Augé, Reference Bailon and Augé2012
aff. Ophisaurus Bailon (unpub. Ph.D. thesis, Univ. Paris, 1991)
Holotype. Right dentary UM MED-121. Meda Grande, îles Medas, Catalonia, Spain.
Diagnosis. After Bailon & Augé (Reference Bailon and Augé2012).
Locality and horizon. Lower Pleistocene; Meda Grande, îles Medas, Catalonia, Spain.
Remarks. The dentary of this Lower Pleistocene species is like that of the Oligocene species of Ophisauromimus in similar morphology of the intramandibular septum forming a large, perpendicular septum extending posteriorly to a small process, and the ventrolateral margin of the septum is free (Čerňanský, Klembara & Müller, Reference Čerňanský, Klembara and Müller2016). These features are not present in Ophisaurus. Ragesaurus medasensis differs from all other anguines in which the marginal dentition is preserved in the following features (Bailon & Augé, Reference Bailon and Augé2012): (1) the anterior inferior alveolar foramen is medially bordered by a small perpendicular septum; (2) the Meckelian groove is very narrow in the mid-length of the dentary in ventral view; and (3) the teeth are conical and their bases are not mediodistally widened; their apices are unstriated and their mesial and distal cutting edges are absent.
Genus Headonhillia Klembara & Green, Reference Klembara and Green2010
Headonhillia parva Klembara & Green, Reference Klembara and Green2010
Holotype. Parietal BMNH R13522. Bembridge Limestone Formation, Headon Hill, Isle of Wight, UK.
Diagnosis. Headonhillia is distinguished from other anguines on the basis of the following combination of features: (1) small size (the smallest of all the anguines; it approaches the size of Anguis fragilis and juveniles of Ophisaurus); (2) large smooth area of parietal table (twice the length of the occipital shield, and anteroposteriorly the longest of any anguine); and (3) short postfoveal crest which is about a quarter of the length of the posterior section of the parietal cranial crest. For the analysis of these characters, see Klembara & Green (Reference Klembara and Green2010).
Locality and horizon. Late Eocene, Ludian (Priabonian); Bed 6, Bembridge Limestone Formation, Headon Hill, Isle of Wight, UK.
Genus Ophisauriscus Kuhn, Reference Kuhn1940
Ophisauriscus quadrupes Kuhn, Reference Kuhn1940
Holotype. Almost complete specimen GM Ce VII-804; upper middle coal MP 13, Geiseltal, Grube Cecilie site VII (Ce VII), Germany.
Diagnosis. After Kuhn (Reference Kuhn1940) and Sullivan, Keller & Habersetzer (Reference Sullivan, Keller and Habersetzer1999).
Locality and horizon. Grube Cecilie site VII (Ce VII), upper middle coal MP 13, Geiseltal, Germany.
Remarks. Meszoely & Haubold (Reference Meszoely and Haubold1975) were the first to note the affinity of Ophisauriscus quadrupes to Anguinae. Klembara (Reference Klembara1981) came to a similar conclusion. Sullivan, Keller & Habersetzer (Reference Sullivan, Keller and Habersetzer1999) revised all finds assigned to Anguidae by Kuhn (Reference Kuhn1940) and assigned O. quadrupes as belonging to ?‘Anguinae’, the taxon based on plesiomorphies and thus no support for its homology. Conrad (Reference Conrad2008) and Conrad et al. (Reference Conrad, Ast, Montanari and Norell2011) questioned the affinity of O. quadrupes to Anguinae. Because there is no convincing anatomical evidence that would clearly pose O. quadrupes outside Anguinae, here it is included tentatively in Anguinae. However, a review of O. quadrupes is required to resolve its relationships.
It is necessary to emphasize that there are two features of the parietal which are similar in the Oligocene species Ophisaurus roqueprunensis (France and Belgium) and the Eocene taxa Ophisaurus sp., Headonhillia parva (England) and Ophisauriscus quadrupes (Germany): (1) the smooth area is longer than that of the occipital shield, when measured in the median plane; and (2) the anterior end of the ventrolateral ridge of the supratemporal process joins the posterior section of the parietal cranial crest at or slightly posterior to the level of the posteromedian margin of the parietal fossa floor. To evaluate these similarities, the revision of the skeletal material of O. quadrupes is needed.
4. Discussion
To summarize, there are six species of Ophisaurus coming from the Miocene of Europe and Canada. Four species of Ophisaurus from the Early Miocene of the Czech Republic and Germany are described on the basis of parietals: O. fejfari, O. spinari, O. robustus (Klembara, Reference Klembara1979, Reference Klembara1981; Čerňanský, Rage & Klembara, Reference Čerňanský, Rage and Klembara2015) and O. holeci (Klembara, Reference Klembara2015). The fifth species of Ophisaurus, O. acuminatus from the lower Late Miocene of Germany (Jörg, Reference Jörg1965), is based on partially articulated cranial elements without a parietal. The same is true for the sixth species, O. canadensis, based on one vertebra from the Late Miocene of Canada (Holman, Reference Holman1970). Recently, a new find of Ophisaurus, described as Dopasia (=Ophisaurus) sp., has been recorded in the Late Miocene of North Africa (Blain et al. Reference Blain, Agustí, López-Garcia, Haddoumi, Aouraghe, Hammouti, Pérez-Gonzáles, Chacón and Sala2013) and represents the earliest remains of this genus in Africa.
From the Oligocene of France, one parietal is present for Ophisaurus roqueprunensis (Augé, Reference Augé1992, Reference Augé2005; Augé & Smith, Reference Augé and Smith2009).
From the Eocene, a parietal of Ophisaurus sp. is described from the Late Eocene of the Hampshire Basin in southern England (Klembara & Green, Reference Klembara and Green2010).
Further isolated skeletal elements (mainly comprising dentaries, vertebrae and osteoderms) have been described from various Cenozoic localities of Europe and attributed with some certainty to the genus Ophisaurus (e.g. Ophisaurus sp., Rage & Sen, Reference Rage and Sen1976; Roček, Reference Roček1984; Augé, Reference Augé1990; Böhme, Reference Böhme1999, 2008; Augé & Rage, Reference Augé and Rage2000; Murelaga et al. Reference Murelaga, Suberbiola, Broin, Rage, Duffaud, Astibia and Badiola2002; Rage & Bailon, Reference Rage and Bailon2005; Venczel, Reference Venczel2006, Reference Venczel2007; Dopasia sp., Delfino, Bailon & Pitruzzella, Reference Delfino, Bailon and Pitruzzella2011; Blain et al. Reference Blain, Agustí, López-Garcia, Haddoumi, Aouraghe, Hammouti, Pérez-Gonzáles, Chacón and Sala2013; Venczel & Hír, Reference Venczel and Hír2015; Čerňanský, Klembara & Müller, Reference Čerňanský, Klembara and Müller2016). Dopasia s.l. sp. from the Oligocene of France is described on the basis of one dentary and several trunk and caudal vertebrae (Rage & Augé, Reference Rage and Augé2015); however, Čerňanský, Klembara & Müller (Reference Čerňanský, Klembara and Müller2016) consider these skeletal elements as belonging to a new genus Ophisauromimus.
There are four fossil species of the genus Pseudopus, P. laurillardi and P. pannonicus (Early Miocene to Pleistocene of Europe and Central Asia; for review see Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010) and P. ahnikoviensis and P. confertus from the Early Miocene of Merkur (Klembara, Reference Klembara2012, Reference Klembara2015), and two parietals of Pseudopus sp. from Germany and Slovakia are described herein.
Previously, two Cenozoic species of the genus Anguis were described: A. stammeri from the Late Pleistocene of Germany and A. polgardiensis from the Late Miocene of Hungary. However, A. stammeri requires revision and in the case of A. polgardiensis it is not clear whether this species belongs to Anguis or Ophisaurus. The third species is Anguis rarus sp. nov. described here from the Early Miocene of Germany.
Together, six species of Ophisaurus, four species of Pseudopus and one species of Anguis are recognized in the Miocene of Europe and Canada.
There are many isolated skeletal elements (mostly dentaries and vertebrae) and osteoderms described as cf. Anguis or Anguis sp. from various Cenozoic localities of Europe. The earliest come from the Early Eocene of France and Belgium (Augé, Reference Augé1990, Reference Augé2005; Rage & Augé, Reference Rage and Augé1993; Rage, Reference Rage2006). Dentaries, vertebrae and osteoderms designated as Anguis sp. were also described from the Early and Middle Eocene of France (Augé et al. Reference Augé, Duffaud, Broin, Rage and Vasse1997; Rage & Augé, Reference Rage and Augé2010). The vertebrae unambiguously belonging to Anguis come from the Early Miocene (MN 4) of Dolnice in the Czech Republic (Klembara, Reference Klembara1981, fig. 3C). It must be emphasized, however, that as in the case of Ophisaurus (see below) it is no certainty that these skeletal elements or their fragments from the Eocene really represent the genus Anguis. Only better-preserved specimens can help to resolve this taxonomic problem.
As previously mentioned, a detailed comparative analysis of the studied skeletal elements is required to differentiate clearly Anguis and Ophisaurus elements. According to the genetic data, the branching event separating Anguis and Pseudopus apodus is c. 9 Ma old (Macey et al. Reference Macey, Shulte II, Larson, Tuniyev, Orlov and Papenfuss1999). Unfortunately, Macey et al. (Reference Macey, Shulte II, Larson, Tuniyev, Orlov and Papenfuss1999) completely ignored the fossil record. Anyway, the presence of Anguis in the Early Miocene, that is, from c. 20 Ma (Klembara, Reference Klembara1981 and this paper), is confirmed.
Now we can conclude that the earliest find of the parietal attributable to Ophisaurus is represented by Ophisaurus sp. from the Late Eocene of the Hampshire Basin (Klembara & Green, Reference Klembara and Green2010), whereas the earliest finds of vertebrae most likely belonging to Ophisaurus are from the Early Eocene of France (cf. Ophisaurus, Augé, Reference Augé1990). However, as indicated above, it is not clear whether the genus Ophisaurus was already present in the Eocene and Oligocene. Only a more complete fossil record may elucidate this taxonomic problem. The earliest finds of Pseudopus are from the Early Miocene of the Czech Republic (Klembara, Reference Klembara2012, Reference Klembara2015); however, it is highly probable that the vertebrae and osteoderms described from the Late Oligocene of Herrlingen (Germany) belong to Pseudopus (Čerňanský, Klembara & Müller, Reference Čerňanský, Klembara and Müller2016). The presence of at least five species of Ophisaurus, four species of Pseudopus and one species of Anguis in the Miocene of Europe documents that anguines were most likely more diversified in this geological period than in preceding Cenozoic periods. While anguines from the Oligocene are rather small and have low diversity (Augé, Reference Augé2005; Augé & Smith, Reference Augé and Smith2009; Čerňanský, Klembara & Müller, Reference Čerňanský, Klembara and Müller2016), abundant osteoderms and vertebrae from the pre-Grande Coupure, i.e. Eocene, sediments of the Hampshire Basin are large, at least as large as in adult specimens of extant Pseudopus apodus (Klembara & Green, Reference Klembara and Green2010). Therefore more complete anguine material from the Eocene is required to understand their size and diversity in this warm period of the Cenozoic.
4.a. Palaeobiogeography
Currently several Ophisaurus species live in North America and Southeast Asia, and one species lives in North Africa and several Anguis species have recently been identified by genetic data in Europe and Southwest Asia (Gvoždík et al. Reference Gvoždík, Jandzík, Lymberakis, Jablonski and Moravec2010). However, Pseudopus apodus is the only species of Pseudopus surviving today. With regard to the distribution of fossil species of Ophisaurus in the Cenozoic, the Eocene and Oligocene species are restricted to Western Europe (France, England, Belgium and Spain) and Central Germany while Miocene species range from the Early Miocene of Germany (Čerňanský, Rage & Klembara, Reference Čerňanský, Rage and Klembara2015) and Central Europe (the Czech Republic; Klembara, Reference Klembara2015) to the Late Miocene O. canadensis of Saskatchewan (Canada) (Holman, Reference Holman1970). The youngest fossil records of Ophisaurus are reported from various Pliocene and Pleistocene localities of Europe (M. Böhme & A. Ilg, unpub. data, 2008, www.wahre.staerke.com) and the United States (Estes, Reference Estes and Wellenhofer1983). It is considered that Ophisaurus reached the North American continent at least in the Late Miocene, while several species persisted in the Middle and Late Miocene in Europe (e.g. O. acuminatus, lower Late Miocene of Germany; Ophisaurus robustus from the late Middle Miocene (MN 7–8) sediments of Hungary (Hír et al. Reference Hír, Kókay, Venczel, Gál and Kessler2001; Venczel & Hír, Reference Venczel and Hír2013). The oldest finding of Ophisaurus in North Africa also comes from the Late Miocene (Blain et al. Reference Blain, Agustí, López-Garcia, Haddoumi, Aouraghe, Hammouti, Pérez-Gonzáles, Chacón and Sala2013). In the Pliocene, Ophisaurus is present in southern Italy (Delfino, Bailon & Pitruzzella, Reference Delfino, Bailon and Pitruzzella2011 and this paper) and the youngest finds of Ophisaurus in Europe come from the Early Pleistocene (south of the Iberian Peninsula: Barranco León 5 and Fuente Nueva 3; Bailon & Blain, Reference Bailon and Blain2007).
No anguine fossil specimens were recorded in the Palaeocene, and it is surmised that the Anguinae ancestor came to Europe from North America via a land bridge connecting them (via Greenland) between the Late Cretaceous and Early Eocene (Klembara, Reference Klembara1981). In addition to Ophisaurus and Anguis, the anguine Headonhilia parva was also described in the Eocene (Klembara & Green, Reference Klembara and Green2010) and the further potential anguine, Ophisauriscus quadrupes, was present in the Middle Eocene of Germany (Kuhn, Reference Kuhn1940; Sullivan, Keller & Habersetzer, Reference Sullivan, Keller and Habersetzer1999; however, see Conrad, Reference Conrad2008 and Conrad et al. Reference Conrad, Ast, Montanari and Norell2011).
Such palaeogeographic distribution of Ophisaurus indicates that Ophisaurus probably originated in Western Europe where it is represented by Ophisaurus sp. from the Late Eocene of southern England (Klembara & Green, Reference Klembara and Green2010), although several skeletal elements probably belonging to Ophisaurus come from the Early Eocene of France (cf. Ophisaurus; Augé, Reference Augé1990). Later it spread to central and eastern regions of Europe. Because the Turgai Strait became dry land during the Early Oligocene (Rögl, Reference Rögl1998; Hou et al. Reference Hou, Sket, Fišer and Li2011), Ophisaurus might have spread to the east. Recent unique finds of several skeletal elements from the Middle Miocene and Early Pleistocene deposits of northern Kazakhstan document the presence of fossil Ophisaurus in Asia (Ophisaurus sp.; Vasilyan, Böhme & Klembara, Reference Vasilyan, Böhme and Klembara2016). The extant species of Ophisaurus of Southeast Asia are considered descendants of the European Neogene species of Ophisaurus. From East Asia, Ophisaurus most likely emigrated via the Bering Strait to North America as documented by the dorsal vertebra in the Late Miocene of Canada (Holman, Reference Holman1970). From here, Ophisaurus spread to the middle and southern territories of North America (Estes, Reference Estes and Wellenhofer1983). This indicates that Ophisaurus originated in Europe and only later migrated, via Asia, to North America. During the Miocene, Ophisaurus probably migrated to North Africa as documented by several cranial and postcranial remains in the Late Miocene deposits of Morocco (Blain et al. Reference Blain, Agustí, López-Garcia, Haddoumi, Aouraghe, Hammouti, Pérez-Gonzáles, Chacón and Sala2013). As suggested by Blain et al. (Reference Blain, Agustí, López-Garcia, Haddoumi, Aouraghe, Hammouti, Pérez-Gonzáles, Chacón and Sala2013), Ophisaurus may have entered North Africa during or prior to the Middle Miocene via the land bridge between the Afro-Arabian Plate and Eurasia established from the Early Miocene onwards. Only the one species, Ophisaurus (=Hyalosaurus) koellikeri (Günther, Reference Günther1873), survives in North Africa today.
Such a scenario of the evolution and palaeobiogeography of Ophisaurus and other anguines, although on the basis of a much limited fossil record, was first published in 1981 (Klembara, Reference Klembara1981). Macey et al. (Reference Macey, Shulte II, Larson, Tuniyev, Orlov and Papenfuss1999) came to the same palaeobiogeographic scenario of Ophisaurus evolution based on the genetic data. However, they ignored the results published previously (Klembara, Reference Klembara1981).
As previously mentioned, the earliest skeletal elements considered to be Anguis come from the Early Eocene of France and Belgium: Anguis sp., together with the remains of Ophisaurus (Augé, Reference Augé1990, Reference Augé2005). This means that the earliest record of this taxon is from Western Europe, similar to that of Ophisaurus (Rage & Augé, Reference Rage and Augé1993). In the Early Miocene, Anguis is already present in Germany (this paper) and the Czech Republic (Klembara, Reference Klembara1981). The recent morphological analysis of the parietal of A. polgardiensis from the Late Miocene of Hungary (Bolkay, Reference Bolkay1913) found that it was impossible to positively distinguish it as Anguis or Ophisaurus (Klembara, Reference Klembara2015). Later, Anguis is recorded in the Pliocene localities in Poland (Młynarski, Reference Młynarski1962) and the Plio-Pleistocene deposits at various localities of Western Europe (Bailon & Blain, Reference Bailon and Blain2007). All these recordings document that Anguis originated in Western Europe but later immigrated to Eastern European areas. Today, the most easterly occurrence of Anguis is in Southwest Asia (Georgia, Russia, Iran) (Gvoždík et al. Reference Gvoždík, Jandzík, Lymberakis, Jablonski and Moravec2010). Anyway, the presence of fossils strongly indicates that Western and Central Europe are the places of origin of Anguis and Ophisaurus.
The oldest species of Pseudopus, P. ahnikoviensis, comes from the Early Miocene (MN 3) locality in the Czech Republic (Klembara, Reference Klembara2012). More recently, the fragments of dentary and osteoderms belonging to Pseudopus sp. were identified from the German locality Wiesbaden–Amöneburg (MN 2) (Čerňanský, Rage & Klembara, Reference Čerňanský, Rage and Klembara2015). Pseudopus laurillardi (Augé & Rage, Reference Augé and Rage2000; Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010) comes from France and Germany. The oldest record is from the Early Miocene deposits (zone MN 4) of Germany, while the youngest finds of this species are from the late Middle Miocene of Germany (Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010). Here, Pseudopus sp. is described from Slovakia (zone MN 6) and south Germany (zone MN 7/8). The largest species of Pseudopus is P. pannonicus first described from Hungary (Kormos, Reference Kormos1911). This species ranges from the Late Miocene to the Early Pleistocene of Europe (e.g. Fejérváry-Lángh, Reference Fejérváry-Lángh1923; Bachmayer & Mlynarski, Reference Bachmayer and Mlynarski1977; Klembara, Reference Klembara1986 b; Rauscher, Reference Rauscher1992; Delfino & Bailon, Reference Delfino and Bailon2000; Tempfer, Reference Tempfer2004; Klembara, Böhme & Rummel, Reference Klembara, Böhme and Rummel2010). The recent species, Pseudopus apodus, was found in the Early and Late Pleistocene (M. Böhme & A. Ilg, unpub. data, 2008, www.wahre.staerke.com) and was more likely present in the Middle Pliocene (Zanclean, MN 15) of Slovakia (Klembara, Reference Klembara1986 b). It lives in arid and semi-arid territories from the Balkan region of Europe to Kazakhstan. As in Ophisaurus and Anguis, the fossil record of Pseudopus highlights west–east spreading of this genus from the Early Miocene to the Holocene. Macey et al. (Reference Macey, Shulte II, Larson, Tuniyev, Orlov and Papenfuss1999) suggested that the Ophisaurus generic name for O. apodus should be replaced by the original Pseudopus, but this had already been accomplished by Klembara (Reference Klembara1979).
Acknowledgements
We thankProf. O. Fejfar and Prof. Z. Roček (Charles University in Prague) and Dr B. Ekrt (National Museum, Prague) for the opportunity to study the material described in this paper. We also thank Dr K. Smith (Senckenberg Research Institute, Frankfurt) for allowing us the use of his extant anguines collection and Dr A. Bolet (Institut Català de paleontologia, Barcelona, Spain) for his useful comments on the earliest version of this paper. The photographs (Figs 2, 3e, f, 4, 6, 7c, d, 8, 9a, b, 10a, b) and (Figs 1, 3a–d, 5, 7a, b, 9c, d) were taken by J. Kotus (Bratislava) and Dr K. Dobiašová (Faculty of Natural Sciences, Bratislava) respectively. We thank Dr Andrew R. Milner (Natural History Museum, London) for useful discussion regarding taxonomic determinations and Dr R. J. Marshall (Sydney, Australia) and B. Sládečková (Košice, Slovakia) for grammar and stylistic review of the text. Finally, we thank the reviewers, Drs S. E. Evans (University College London) and J.-C. Rage (Muséum National d'Histoire Naturelle, Paris) for thoughtful reviews and suggestions.











