


1. Introduction
One of the most important scientific challenges in the quest for better knowledge of the Cambrian explosion is the appearance of primitive arthropoda in the fossil record, the dominant phylum in the recent biosphere. In this regard, the records of primitive trilobites are of great importance; not only do they allow us to understand the emergence of arthropoda, but they also constitute a group of guide fossils that allows intercontinental correlation to be established.
Middle lower Cambrian successions (provisional Cambrian Series 2) containing a trilobite record are found at several localities in Spain. This has enabled a trilobite zonation to be established for the Ovetian Stage, a regional stage characterized by the presence of primitive trilobites as well as archaeocyathans. This stage is widely recognized in Spain, France, Portugal, Germany, Morocco and Italy (Sdzuy, Reference Sdzuy1971; Gonçalves, Reference Gonçalves1982; Liñán, Perejón & Sdzuy, Reference Liñán, Perejón and Sdzuy1993; Liñán et al. Reference Liñán, Gozalo, Palacios, Gámez Vintaned, Ugidos, Mayoral, Gibbons and Moreno2002, Reference Liñán, Perejón, Gozalo, Moreno-Eiris and Oliveira2004; Gozalo et al. Reference Gozalo, Liñán, Palacios, Gámez Vintaned and Mayoral2003; Pillola et al. Reference Pillola, Gámez-Vintaned, Dabard, Leone, Liñán, Chauvel, Matteucci, Carboni and Pignatti1994 ; Pillola, Leone & Loi, Reference Pillola, Leone and Loi1995; Elicki, Reference Elicki1997; Perejón & Moreno-Eiris, Reference Perejón and Moreno-Eiris2006). From a perspective of evolution and intercontinental correlation, the most important Ovetian trilobite successions in Spain are located in the provinces of Córdoba (Liñán et al. Reference Liñán, Dies, Gámez Vintaned, Gozalo, Mayoral and Muñiz2005) and Badajoz (Liñán et al. Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b ) in the south of the Iberian Peninsula; Toledo (Liñán & Gámez-Vintaned, Reference Liñán and Gámez-Vintaned1993; Jensen, Palacios & Martí Mus, Reference Jensen, Palacios and Mus2010) and Salamanca (Rodríguez et al. Reference Rodríguez Alonso, Alonso Gavilán, Liñán, Gámez-Vintaned, Pérez García, Rodríguez Alonso and Alonso Gavilán1995) in the central region; and León (Sdzuy, Reference Sdzuy1961) in the north.
The agraulid trilobites of the Ovetian Stage (Series 2) in Spain are revised in this work as a result of recently found material. They are restricted to siliciclastic facies (ranging from coarse sandstone and microconglomerate to siltstone), the reason for their scarce fossil record. They have been found in two of the Spanish successions mentioned above. One set of specimens came from the locality of Los Barrios de Luna in the Cantabrian Cordillera of the province of León, and an additional specimen came from La Rinconada in the province of Salamanca.
From a geological standpoint (Lotze, Reference Lotze1945; Fig. 1), Los Barrios de Luna is located in the Cantabrian Zone while La Rinconada lies in the Galician–Castilian Zone (included in some papers in the so-called Central–Iberian Zone, sensu Julivert et al. Reference Julivert, Fontboté, Ribeiro and Nabais Conde1972).
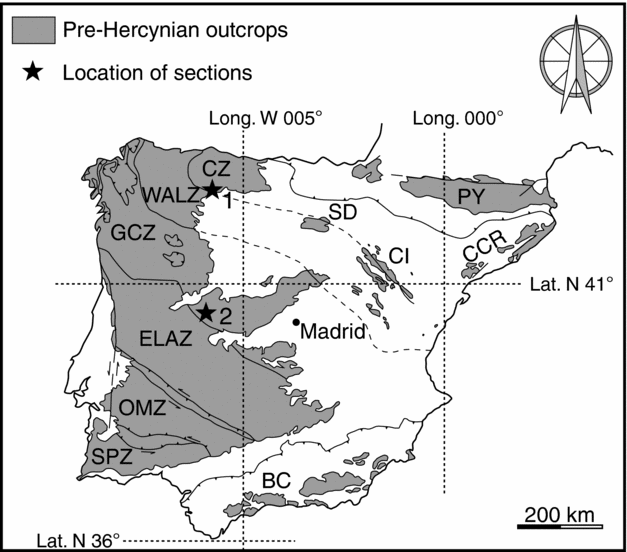
Figure 1. Tectonostratigraphic zones of the Iberian Massif (mostly after Lotze, Reference Lotze1945) and other pre-Hercynian outcrops of Iberia (SD, CI, PY, CCR and BC), with location of Los Barrios de Luna (star 1) and La Rinconada (star 2). CZ – Cantabrian Zone; WALZ – West Asturian – Leonese Zone; GCZ – Galician–Castilian Zone; ELAZ – East Lusitanian –Alcudian Zone; OMZ – Ossa–Morena Zone; SPZ – South Portuguese Zone; SD – Sierra de la Demanda; CI – Cadenas Ibéricas (Iberian Chains); PY – Pyrenees; CCR – Catalonian Coastal Ranges; BC – Betic Cordillera.
2. Stratigraphy
The Cambrian stratigraphy of Los Barrios de Luna was described by Comte (Reference Comte1937a , Reference Comte b , Reference Comte1938, Reference Comte1959) who established the following units from bottom to top: La Herrería, Láncara, Oville and Barrios formations. A synthesis is found in Aramburu et al. (Reference Aramburu, Bernárdez, Gozalo, Gutiérrez-Marco and Liñán2006).
We have made a detailed litho- and biostratigraphical study of the La Herrería Formation (Corduban–Ovetian in age) at Los Barrios de Luna through a natural section running south–north, located to the east of the village. It starts on a dirt road near the television repeater tower and continues along road CL-626, eventually reaching the Los Barrios de Luna reservoir. The section was named ‘Los Barrios de Luna 1’ (BL1) by Aramburu et al. (Reference Aramburu, Bernárdez, Gozalo, Gutiérrez-Marco and Liñán2006) and Gámez Vintaned et al. (Reference Gámez Vintaned, Liñán, Mayoral, Gozalo, Zamora and Fernández-Martínez2006b ). Other sections spanning the La Herrería Formation in the vicinity are those called BL2 and BL3, both located along road LE-3422 (running south–north on the western slope of the valley, from the junction with LE-34321 as far as the Los Barrios de Luna reservoir). The geographic and geological location is shown in Figure 2.

Figure 2. Geology of the areas (a) near Los Barrios de Luna (after Aramburu et al. Reference Aramburu, Arbizu, Gutiérrez Marco, Méndez-Bedia, Rábano and Truyols1996) and (b) La Rinconada (after Martín Herrero et al. Reference Martín Herrero, Ugidos Meana, Nozal Martín and Pardo Alonso1990), with location of the studied sections.
The La Herrería Formation forms a complete outcrop throughout section BL1, although it is faulted at both its base and top (Fig. 3). It is composed of alternating sandstone, siltstone, shale (grey and red) and microconglomerate. At the top of the section Lotze (Reference Lotze1961) and Sdzuy (Reference Sdzuy1961) described ‘fossil point 3’ (or ‘FP3’) containing a fossil assemblage comprising the trilobites Lunolenus lunae Sdzuy, Reference Sdzuy1961 and Metadoxides richterorum Sdzuy, Reference Sdzuy1961, as well as the trace fossil Astropolichnus hispanicus (Crimes et al. Reference Crimes, Legg, Marcos, Arboleya, Crimes and Harper1977; reported as Astropolithon Dawson by Seilacher in Lotze, Reference Lotze1961). These two authors also described ‘fossil point 2’ (or ‘FP2’) with Agraulos antiquus Sdzuy, Reference Sdzuy1961 and Lunolenus prior Sdzuy, Reference Sdzuy1961, located at levels 19m below FP3. In beds further below in the formation, they also described ‘fossil point 1’ (or ‘FP1’) containing Lunolenus? lotzei Sdzuy, Reference Sdzuy1961, but they did not provide an accurate stratigraphical position for this faunule. In this paper we accurately locate all three of Lotze and Sdzuy's points (fossils assemblages) and their detailed biostratigraphic positions (Fig. 3). Lotze (Reference Lotze1961) and Sdzuy (Reference Sdzuy1961) also cited Lunolenus lunae Sdzuy, Reference Sdzuy1961, Dolerolenus formosus Sdzuy, Reference Sdzuy1961 and Metadoxides armatus (Meneghini, Reference Meneghini1881) at the top of the La Herrería Formation in section BL2, corresponding to their ‘fossil point 4’ (or ‘FP4’). They considered the fossil point 4 trilobite assemblage as an equivalent of fossil point 3, although their fossil contents differ in some respects.

Figure 3. Stratigraphic column of the upper part of the La Herrería Formation, at a lower interval of the Los Barrios de Luna 1 (BL1) section (province of León, northern Spain).
The agraulids that we collected were derived from a new fossiliferous level in the La Herrería Formation, comprising microconglomeratic coarse-grained sandstone. It was located in section BL1, 20m below FP2, containing Agraulos antiquus Sdzuy, Reference Sdzuy1961 (Fig. 3). The new material is assigned to the same species but included in the new genus Lunagraulos.
The early Cambrian succession of the La Rinconada section is composed (in ascending stratigraphic order) of the Aldeatejada Formation, the Tamames Sandstone and the Tamames Limestone (Rölz, Reference Rölz1975; Díez Balda, Reference Díez Balda1986), cropping out at the northern flank of the Tamames Syncline. The trilobite material studied here was found in a level of siltstone located in the lower part of the Tamames Sandstone and is described here as the new species Lunagraulos tamamensis n. gen. n. sp. (Fig. 4). It was recorded immediately above the ichnotaxa Dactyloidites cabanasi (Meléndez in Cabanás, Reference Cabanás1966) and Astropolichnus hispanicus (Crimes et al. Reference Crimes, Legg, Marcos, Arboleya, Crimes and Harper1977).

Figure 4. Stratigraphy of the Cambrian near La Rinconada (province of Salamanca, central Spain). After Díez Balda (Reference Díez Balda1986).
A younger (middle Marianian), lower Cambrian unit, the Endrinal Shale, crops out to the east of La Rinconada at the core of the Endrinal Syncline (García de Figuerola & Martínez García, Reference García de Figuerola and Martínez García1972). No Cambrian rocks younger than the Endrinal Shale crop out in this area of the Galician–Castilian Zone.
3. Systematic palaeontology
The new material studied in this work is deposited in the Museo de Ciencias Naturales de la Universidad de Zaragoza (University of Zaragoza Museum of Natural Sciences), Spain – formerly known (until 2012) as the Museo Paleontológico de la Universidad de Zaragoza (University of Zaragoza Museum of Palaeontology) – in repository MPZ 2014/208 to MPZ 2014/213. Additionally, the holotype of Serrania verae Liñán Guijarro, Reference Liñán Guijarro1978 (type species of the genus Serrania Liñán Guijarro, Reference Liñán Guijarro1978) is currently deposited in the same museum as MPZ 2014/214.
Sdzuy's material is housed at the Geomuseum der Westfälische Wilhelms-Universität Münster (Geomuseum of the University of Muenster), Germany, formerly known (until 2007) as the Geologisch-Palaeontologisches Institut und Museum der Westfälische Wilhelms-Universität Münster, in repository L 3315, L 3317 and L 3318 (Sdzuy, Reference Sdzuy1961).
Family AGRAULIDAE Raymond, Reference Raymond1913
Genus Lunagraulos n. gen.
Type species: Agraulos antiquus Sdzuy, Reference Sdzuy1961
Derivatio nominis: Composite name that combines the River Luna, in whose valley it was first found, with the former genus determination.
Diagnosis: Redlichiina with a smooth cranidium with prominent glabella; eyes well marked only in holaspis specimens, which have arched and tenuous ocular ridges. Facial suture with the parameter S (Liñán Guijarro, Reference Liñán Guijarro1978) cutting the eye lobe (secant), convergent, parallel to the anterior and posterior suture branches and also cutting the posterior branch of the suture (Fig. 5). Preocular area convex, at a lower level than the glabella; anterior margin slightly arched to pointed. Thorax with at least ten segments, directed backwards. Micropygous. Librigena unknown.

Figure 5. (a) Reconstruction of the dorsal exoskeleton of Lunagraulos antiquus. (b) Cranidium of Lunagraulos antiquus (after Sdzuy, Reference Sdzuy1961) showing the characteristic S parameter (Liñán Guijarro, Reference Liñán Guijarro1978).
Comparison: The closest genus is Agraulos Hawle and Corda, Reference Hawle and Corda1847 from the traditional middle Cambrian rocks, showing a closely similar cranidium; the morphology of the thorax is, however, different (the pleural points of Agraulos are blunt and directed forwards); furthermore, the S parameter of the facial suture is tangent and therefore does not cut the eye lobe or the posterior branch of the facial suture; finally, the preocular area has a central enlargement which is absent from Lunagraulos. The genus Proampyx Frech, Reference Frech1897 is also similar; however, it differs in having well-marked eyes and a preocular furrow coinciding with a high relief of the cranidium. Another quite similar genus is Giordanella Bornemann, Reference Bornemann1891 from Sardinia, Italy (Pillola, Reference Pillola1991), but this genus has a rectangular glabella and the S parameter does not cut the palpebral lobe. The ptychoparioid trilobite assigned to Glabrella? Lermontova, Reference Lermontova, Vologdin, Lermontova, Yavorsky and Janischevsky1940, from lower Cambrian deposits of Antarctica (Palmer & Gatehouse, Reference Palmer and Gatehouse1972), shows some similarities with Lunagraulos but its preocular area is very short and it lacks a prominent and well-defined glabella. The Chinese genus Paragraulos Lu, Reference Lu1941, from upper lower Cambrian deposits, has a distinct anterior border and furrow, as well as very different eyes and facial suture. Plesiagraulos Chang, Reference Chang1963, from middle Cambrian deposits of China (Stage 5), differs in having convergent anterior branches of the facial suture, and also a straight and depressed anterior border. The two genera and their species were revised by Yuan & Li (Reference Yuan and Li1999).
Lunagraulos antiquus (Sdzuy, Reference Sdzuy1961)
Figures 5; 6a–k, r
v. 1961 Agraulos antiquus n. sp.; Sdzuy, p. 623–625, plate 22, figs 17–19; plate 23, figs 1–6; Abb. 35.

Figure 6. (a–k, r) Lunagraulos antiquus (Sdzuy, Reference Sdzuy1961). (a) Münster collection L 3318; (b–d) holotype, Münster collection L 3315; (e) Münster collection L 3317 (all specimens of the Münster collection are internal moulds); (g–h) MPZ 2014/211, latex cast (note the preservation in a microconglomerate); (f) MPZ 2014/210, latex cast; (i–k) MPZ 2014/209 (j, lateral view; k, frontal view), latex cast (note the microconglomerate grain size); (r) MPZ 2014/208, latex cast of external mould. (l–o) Lunagraulos tamamensis n. gen. n. sp., MPZ 2014/213 (l, retrodeformed), latex cast of external mould. (p) Serrania verae Liñán Guijarro, 1978, holotype, internal mould MPZ 2014/214. (q) Lemdadella linaresae Liñán Guijarro, 1978, internal mould, MPZ 2002/223. (s) Bigotina bivallata Cobbold, Reference Cobbold1935, internal mould MPZ 2002/80.
v. 2006 Agraulos antiquus; Aramburu et al., p. 36.
New material: Six cranidia, preserved in microconglomeratic coarse-grained sandstone. In addition, Sdzuy (Reference Sdzuy1961) studied and figured seven cranidia and one nearly complete specimen lacking the librigena. The material is not tectonically distorted.
Remarks: Sdzuy (Reference Sdzuy1961) provided a very detailed description of this species. The new cranidia now studied (from level BL1/22) are relatively large and are considered to represent adult specimens (Fig. 6f–k, r). Their characteristics are identical to the specimens described by Sdzuy (Fig. 6a–e) and sampled at a level located some 20m above in the same section (level BL1/23). The specimens in the two assemblages appear with either a pointed or rounded anterior margin, and they also show differences in the axial length of the preocular area in relation to the axial length of the glabella, an intraspecific variation already cited by Sdzuy (Reference Sdzuy1961).
Measurements: The holotype (Fig. 6b–d) measures 12mm along the axis. The largest specimen (Fig. 6i, k) measures 15mm along the axis. The axial length of the preocular area is between 1:4 and 1:2.5 of that of the glabella.
Biostratigraphy: Sdzuy (Reference Sdzuy1971) included L. antiquus in the Ovetian Stage due to its stratigraphic position in the middle part of a transgressive siliciclastic lithosome (the La Herrería Formation) which constitutes the base of the Cambrian succession in northern Spain, and also considering the similar locations of basal Cambrian units elsewhere in Europe. The La Herrería Formation is stratigraphically located right above the Precambrian–Cambrian boundary. The Ovetian age for Lunagraulos antiquus is also congruent with the associated ichnofossil record, as discussed in Section 4. Sdzuy considered the assemblage Lunolenus-Dolerolenus-Metadoxides as upper Ovetian in nature but, given that Lunagraulos antiquus is found below this assemblage (Fig. 5), we cannot definitively rule out an early Ovetian age for the species.
Lunagraulos tamamensis n. gen. n. sp.
(Fig. 6l–o)
v. 1995 Giordanella? sp. Rodríguez Alonso et al., p. 22.
Derivatio nominis: After the village of Tamames, province of Salamanca, central Spain.
Material: After extensive sampling, only one well-preserved adult cranidium was found in a greenish siltstone. The specimen is somewhat tectonically distorted.
Diagnosis: Lunagraulos species characterized by having the preocular and preglabellar areas located on a plane below that of the glabella, forming a gently sloping, flattened ramp that finishes in a curved anterior margin (Fig. 6m). Eye small, distinct and having an ocular ridge. The palpebral lobe and the anterior branch of the suture are similar in length.
Description: Cranidium with arched anterior margin. Preocular field located on a plane below that of the glabella, forming a gently sloping, flattened ramp, somewhat curved near the margin. Glabella prominent and trapezoidal in shape, with a poorly developed occipital furrow. Facial suture with the three branches of similar length. S parameter divergent, secant and subparallel to the anterior-posterior suture branches. Posterior furrow distinct and directed forwards; posterior margin arched.
Comparison: Preocular area forms a flattened ramp, curved anterior furrow and smaller palpebral area distinguish this species from L. antiquus, in which the anterior furrow of the cranidium is typically straight and well distinct. Moreover, the adult stage of L. tamamensis shows eyes with an ocular ridge and a palpebral lobe.
Measurements: The cranidium is 8mm in length along the axis; the ratio of the axial length of the preocular area to that of the glabella is 0.29.
Biostratigraphy: The Tamames Sandstone lies with an apparent conformity on top of the Aldeatejada Formation (upper part of the Schist–Greywacke Complex). According to Díez Balda & Fournier Viñas (Reference Díez Balda and Fournier Viñas1981), it contains the late Neoproterozoic – lower Cambrian acritarchs Synsphaeridium sp. and Micrhystridium dissimilare Volkova, Reference Volkova and Rozanov1969. Material attributed to the latter taxon was revised by Vidal et al. (Reference Vidal, Palacios, Gámez-Vintaned, Díez Balda and Grant1994, fig. 16e, g, h) who transferred it to Heliosphaeridium sp., interpreted by these authors as late Vendian (Ediacaran) in age. The Aldeatejada Formation also contains the early Cambrian trace fossil Treptichnus? ichnosp. near its upper part (Rodríguez Alonso et al. Reference Rodríguez Alonso, Alonso Gavilán, Liñán, Gámez-Vintaned, Pérez García, Rodríguez Alonso and Alonso Gavilán1995), which supports a late Vendian (Ediacaran) – early Corduban (Terreneuvian) age for this unit.
Lunagraulos tamamensis is found in the lower part of the Tamames Sandstone, in levels immediately above the ichnotaxa Dactyloidites cabanasi (Meléndez in Cabanás, Reference Cabanás1966) and Astropolichnus hispanicus (Crimes et al. Reference Crimes, Legg, Marcos, Arboleya, Crimes and Harper1977), reported as Haentzschelinia sp.? and Astropolithon hispanicus, respectively, by Díez Balda (Reference Díez Balda1986). The observed biostratigraphic range for D. cabanasi in Spain is Corduban – lower Ovetian (see following section). A. hispanicus is considered by Pillola et al. (Reference Pillola, Gámez-Vintaned, Dabard, Leone, Liñán, Chauvel, Matteucci, Carboni and Pignatti1994) as indicative of the Ovetian Stage. Both the Corduban and the Ovetian are regional Mediterranean stages (Liñán, Perejón & Sdzuy, Reference Liñán, Perejón and Sdzuy1993; Liñán et al. Reference Liñán, Gozalo, Palacios, Gámez Vintaned, Ugidos, Mayoral, Gibbons and Moreno2002). An early Ovetian age is therefore considered here for Lunagraulos tamamensis n. gen. n. sp.
4. Age and correlation
The biostratigraphic distribution of Lunagraulos antiquus (Sdzuy, Reference Sdzuy1961) in Los Barrios de Luna coincides with that of the north-western Gondwanan ichnospecies Astropolichnus hispanicus, which was considered a low lower Cambrian index fossil by Crimes et al. (Reference Crimes, Legg, Marcos, Arboleya, Crimes and Harper1977, p. 114). The biostratigraphic utility was later confirmed by Pillola et al. (Reference Pillola, Gámez-Vintaned, Dabard, Leone, Liñán, Chauvel, Matteucci, Carboni and Pignatti1994) who assigned an Ovetian age for this ichnospecies. The Ovetian is the second stage in the biochronological scale first proposed by Sdzuy (Reference Sdzuy1971) and Liñán, Perejón & Sdzuy (Reference Liñán, Perejón and Sdzuy1993) for the Cambrian successions of Spain, a scale which is widely employed in studies of the lower Cambrian Mediterranean region.
In the Cadenas Ibéricas (Iberian Chains) of north-eastern Spain, Astropolichnus hispanicus is recorded in the Embid Formation below the trilobites Thoralaspis sp. and Metadoxides? sp. of the basal Jalón Formation (Sdzuy, Reference Sdzuy1987). In the Toledo Mountains of central Spain, it is recorded below the trilobite species Granolenus midi Jago in Courtessole & Jago, Reference Courtessole and Jago1980 (Moreno, Vegas & Marcos, Reference Moreno, Vegas and Marcos1976; Liñán & Gámez-Vintaned, Reference Liñán and Gámez-Vintaned1993; Liñán et al. Reference Liñán, Perejón, Gozalo, Moreno-Eiris and Oliveira2004; Jensen, Palacios & Martí Mus, Reference Jensen, Palacios and Mus2010). The mentioned trilobite genera are considered to be late Ovetian in age (Liñán et al. Reference Liñán, Gozalo, Palacios, Gámez Vintaned, Ugidos, Mayoral, Gibbons and Moreno2002, Reference Liñán, Perejón, Gozalo, Moreno-Eiris and Oliveira2004).
In the Cantabrian Mountains of northern Spain, Lunagraulos antiquus (Sdzuy, Reference Sdzuy1961) is recorded in levels located stratigraphically below those containing species of Dolerolenus and Metadoxides. These two trilobite genera have also been recorded from the lower part of the lower Cambrian deposits of Sardinia, Italy, by Pillola (Reference Pillola1991). The species Metadoxides armatus (Meneghini, Reference Meneghini1881) is present in both countries. The Metadoxides–Dolerolenus assemblage has generally been assigned to the Mediterranean upper Ovetian Sub-stage (Sdzuy, Reference Sdzuy1971; Pillola et al. Reference Pillola, Gámez-Vintaned, Dabard, Leone, Liñán, Chauvel, Matteucci, Carboni and Pignatti1994; Pillola, Leone & Loi, Reference Pillola, Leone and Loi1995; Liñán et al. Reference Liñán, Gozalo, Palacios, Gámez Vintaned, Ugidos, Mayoral, Gibbons and Moreno2002, Reference Liñán, Perejón, Gozalo, Moreno-Eiris and Oliveira2004), as used for the north-western Gondwanan margin, characterizing the N2 and N3 zones (Dolerolenus courtessolei and Dolerolenus longioculata biozones, respectively) of Sardinia (Pillola, Reference Pillola1991; Pillola, Leone & Loi, Reference Pillola, Leone and Loi1995). Since Metadoxides armatus is characteristic of the Dolerolenus longioculata biozone (or N3 trilobite zone), we suggest that the lowermost stratigraphic record of L. antiquus is coeval with the D. courtessolei biozone (Pillola, Reference Pillola1991), an equivalent of the N2 trilobite zone of Sardinia (Pillola, Leone & Loi, Reference Pillola, Leone and Loi1995). Pillola (Reference Pillola1991) correlated the two Dolerolenus biozones of Sardinia with the former Yunnanaspis Zone of China (today known as Yiliangella–Yunnanaspis trilobite Zone), which is the base of the Tsanglangpuan Stage (Lin, Reference Lin, Zhou and Zhen2008). A tentative correlation of Lunagraulos antiquus with the mentioned Chinese zone is therefore suggested.
However, Perejón et al. (Reference Perejón, Fröhler, Bechstädt, Moreno-Eiris and Boni2000) studied the Sardinian archaeocyathans included in the N2 trilobite zone of Pillola, Leone & Loi (Reference Pillola, Leone and Loi1995) and came to the conclusion that the N2 Zone can be correlated with the lower part of the Botoman Stage of Siberia, which in turn was parallelized with Spanish archaeocyathan Zone VI of the upper Ovetian Substage by Perejón & Moreno-Eiris (Reference Perejón and Moreno-Eiris2006) (Fig. 7).

Figure 7. Biochronology of the species of Lunagraulos and lower Cambrian zonation in Spain based on trilobites, archaeocyathans and trace fossils.
In summary, a late Ovetian (early Botoman) age is suggested for Lunagraulos antiquus.
Lunagraulos tamamensis n. gen. n. sp. is recorded 500m below the Ovetian bigotinid trilobite Serrania sp. at La Rinconada, province of Salamanca, central Spain, and its occurrence coincides with the first record of Astropolichnus hispanicus in the region (Rodríguez Alonso et al. Reference Rodríguez Alonso, Alonso Gavilán, Liñán, Gámez-Vintaned, Pérez García, Rodríguez Alonso and Alonso Gavilán1995; Fig. 4).
Another ichnospecies associated with Lunagraulos tamamensis in the Salamanca area is Dactyloidites cabanasi (Meléndez in Cabanás, Reference Cabanás1966), an ichnotaxon typical of the lower Ovetian Lemdadella linaresae trilobite Zone in Sierra Morena, province of Córdoba, southern Spain (Gámez Vintaned et al. Reference Gámez Vintaned, Liñán, Mayoral, Dies, Gozalo and Muñiz2006a ). D. cabanasi is recorded in association with Psammichnites gigas (Torell, Reference Torell1868) in Corduban rocks ( = Terreneuvian; Landing et al. Reference Landing, Peng, Babcock, Geyer and Moczydlowska-Vidal2007) from the Toledo Mountains in central Spain (Seilacher & Gámez-Vintaned, Reference Seilacher, Gamez-Vintaned and Cherchi1995; Palacios et al. Reference Palacios, Gámez Vintaned, Fernández-Remolar and Liñán1999). Furthermore, the record of L. tamamensis well below the trilobite Serrania sp. in the same section of La Rinconada (Rodríguez Alonso et al. Reference Rodríguez Alonso, Alonso Gavilán, Liñán, Gámez-Vintaned, Pérez García, Rodríguez Alonso and Alonso Gavilán1995) clearly demonstrates an earliest Ovetian age for this species, and a probable equivalence with the interval of archaeocyathan Zone I–II described by Perejón (Reference Perejón1986, Reference Perejón1989, Reference Perejón1994) and Perejón and Moreno-Eiris (Reference Perejón and Moreno-Eiris2006). It is noteworthy that Liñán et al. (Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b ) identified the biochronological position of the early bigotinid trilobites Serrania palaciosi Liñán et al., Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b and Serrania? gordaensis Liñán et al., Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b at the same interval (Fig. 7). Consequently, Lunagraulos tamamensis may actually be one of the oldest trilobite species in Gondwana.
4.a. The record of the oldest trilobites
The intercontinental correlation of the oldest trilobites is a priority matter for the International Subcommission on Cambrian Stratigraphy (ISCS), in order to define the Stage 2 – Stage 3 boundary and the Series 1 – Series 2 boundary for the Cambrian System (Peng & Babcock, Reference Peng, Babcock, Peng, Babcock and Zhu2001).
In southern Europe, the trilobite Bigotina bivallata Cobbold, Reference Cobbold1935 was first described in Carteret, Normandy, northern France. This species, typical of the Ovetian Stage (Bigotina bivallata zone), was later found to be recorded below Lemdadella linaresae Liñán Guijarro, Reference Liñán Guijarro1978 in the Sierra de Córdoba mountain range of southern Spain, but still coexisted for a short time with the latter (Liñán et al. Reference Liñán, Dies, Gámez Vintaned, Gozalo, Mayoral and Muñiz2005) within archaeocyathan Zone III (which was tentatively correlated by Perejón & Moreno-Eiris, Reference Perejón and Moreno-Eiris2006 with the lower Atdabanian or A2 of Siberia). The trilobite cf. Abadiella bourgini Hupé, Reference Hupé1953 has been reported in the central part of Spain from the Pusa Formation of the Toledo Mountains (Jensen, Palacios & Martí Mus, Reference Jensen, Palacios and Mus2010). The age of these trilobites is a subject of controversy. Palacios et al. (Reference Palacios, Gámez Vintaned, Fernández-Remolar and Liñán1999) cited bigotinid trilobites in Robledo del Buey and supposed a latest Tommotian or Atdabanian age for it. These bigotinid trilobites were later assigned an early Corduban age (the Mediterranean parastratotype of the Terreneuvian Series, after Landing et al. Reference Landing, Peng, Babcock, Geyer and Moczydlowska-Vidal2007) by Liñán et al. (Reference Liñán, Gozalo, Palacios, Gámez Vintaned, Ugidos, Mayoral, Gibbons and Moreno2002) and Gozalo et al. (Reference Gozalo, Liñán, Palacios, Gámez Vintaned and Mayoral2003) due to its stratigraphical record under the ichnofossil Cruziana ichnosp. of the Azorejo Formation. Gubanov (Reference Gubanov2002) cited the small shelly fossils Aldanella plana and Oelandiella korobkovi (Nemakit–Daldynian to Tommotian) in the Pusa Formation but did not include the reference localities. Jensen, Palacios & Martí Mus (Reference Jensen, Palacios and Mus2010) later figured non-biohermal archaeocyathans and the small shelly fossils Pelagiella sp. and Cupitheca sp. together with the trilobite cf. Abadiella bourgini in the Robledo del Buey section, as well as Rusophycus cf. avalonensis in the Los Alares section, which they considered to be older than the Robledo del Buey assemblage in base of the lithostratigraphic correlation. These authors considered this assemblage of early Ovetian (archaeocyathan zones I–III) or late Ovetian (archaeocyathan zones IV–VII) age, but tentatively as late early Ovetian age. A. Yu. Zhuravlev (pers. comm., 2009) recognized the archaeocyathan Capsulocyatus sp. (at that moment dated as Zone I of Perejón, Reference Perejón1994 or Atdabanian A1) in the Robledo del Buey assemblage. In Sierra de Córdoba, Pelagiella sp. and the Tommotian–Botoman cf. Watsonella sp. (cf. Heraultia sp.; S. Bengtson, pers. comm. 1976) together with non-biohermal archaeocyathans have been reported 20m below the Archaeocyathan Zone I (the lower part of the Ovetian Stage) at the base of the Pedroche Formation from Las Ermitas section (Liñán Guijarro, Reference Liñán Guijarro1978: plate I, figs 1 and 2, respectively; Brasier, Reference Brasier, Cowie and Brasier1989). These data suggest an earliest Ovetian (Atdabanian A1) age for the Robledo del Buey assemblage or a latest Corduban (Tommotian) age, as was previously assigned by Palacios et al. (Reference Palacios, Gámez Vintaned, Fernández-Remolar and Liñán1999).
The trilobite species Lemdadella tioutensis Sdzuy in Liñán & Sdzuy, Reference Liñán and Sdzuy1978 from the High Atlas mountains in Morocco was included by these authors in the Fallotaspis tazemmourtensis Zone (or Zone 1); these levels were considered as an equivalent of the Lemdadella linaresae Zone (see also Liñán et al. Reference Liñán, Dies, Gámez Vintaned, Gozalo, Mayoral and Muñiz2005). Nevertheless, another tentative correlation between the Fallotaspis tazemmourtensis Zone of Morocco and the Bigotina bivallata Zone of Iberia has been made by Landing et al. (Reference Landing, Geyer, Brasier and Bowring2013). Sdzuy (Reference Sdzuy1981) figured a bigotinid from Amouslek in the Anti-Atlas mountains in Morocco and included it in his trilobite zone 0, to which he also assigned the species Hupetina antiqua Sdzuy, Reference Sdzuy1978, Eofallotaspis prima Sdzuy, Reference Sdzuy1978 and Eofallotaspis tioutensis Sdzuy, Reference Sdzuy1978 from the locality of Tiout in the High Atlas mountains. Pillola (Reference Pillola1993) revised Bigotina bivallata Cobbold, Reference Cobbold1935 from the northern French locality of Carteret in Normandy and elsewhere, and identified the bigotinid from zone 0 of Morocco (sensu Sdzuy, Reference Sdzuy1981) as Bigotina bivallata which he also placed in horizon T2 of the Tiout section (Pillola, Reference Pillola1993, p. 861), 10m above the record of Hupetina antiqua and below the first record of Eofallotaspis prima (horizon T4), Eofallotaspis tioutensis (horizon T10), Lemdadella tioutensis (horizon T16) and Fallotaspis tazemmourtensis (horizon T17) sensu Sdzuy (Reference Sdzuy1978, fig. 2b). Fallotaspis tazemmourtensis was later found in horizon T14 by Geyer & Landing (Reference Geyer and Landing2006, p. 58), supporting the correlation suggested by Liñán & Sdzuy (Reference Liñán and Sdzuy1978). The same opinion is shared by Liñán et al. (Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b ). The correlation of the trilobite Zone 0 (=Eofallotaspis zone) of Morocco with the Bigotina bivallata Zone of southern Europe (base of archaeocyathan Zone III) allows a good correlation between the Issendalenian (sensu Geyer, Reference Geyer1990) and Ovetian stages.
In Avalonia (western Newfoundland and southern Great Britain), the first trilobite records belong to the Callavia Zone which is correlated with the uppermost Atdabanian (Zhuravlev, Reference Zhuravlev1995).
The oldest trilobite from Antarctica is Lemdadella antarctica Palmer & Rowell, Reference Palmer and Rowell1995 originally included in the lower Ovetian Substage. Liñán et al. (Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b ) proposed a correlation of that level with the Lemdadella linaresae – Lemdadella perejoni zones of the lower Ovetian Substage in Spain.
In China, the oldest trilobites are represented by species of Abadiella which represent the beginning of the Nangaoan Stage (Peng & Babcock, Reference Peng, Babcock, Peng, Babcock and Zhu2001; Peng, Reference Peng2003). The Chinese Abadiella Zone has been correlated with the oldest trilobite level of Australia containing Abadiella (Bengtson et al. Reference Bengtson, Conway Morris, Cooper, Jell and Runnegar1990) and is considered equivalent in age to the upper Atdabanian Substage of Siberia (Zhuravlev, Reference Zhuravlev1995; Geyer & Shergold, Reference Geyer and Shergold2000).
According to Repina (Reference Repina and Taylor1981) and Pegel (Reference Pegel2000), the oldest trilobites in Siberia are represented by the genus Profallotaspis recorded at the base of the Atdabanian Stage (A1). Furthermore, Bigotinella and Bigotina species are found in the lower Atdabanian Substage (Suvorova, Reference Suvorova1960; Repina, Reference Repina1969; Egorova, Reference Egorova1983; Zhuravleva, Reference Zhuravleva, Rozanov and Sokolov1984). A correlation between the lower Atdabanian Substage and the lowermost part of the Ovetian Stage was therefore proposed by Liñán et al. (Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b ) due to the presence of trilobites. The same correlation is supported by archaeocyathans (Perejón & Moreno-Eiris, Reference Perejón and Moreno-Eiris2006).
In Baltica, the first trilobites are olenellids from the Schmidtiellus mickwitzi assemblage zone (Bergström, Reference Bergström and Taylor1981; Ahlberg, Reference Ahlberg1984; Ahlberg, Bergström & Johansson, Reference Ahlberg, Bergström and Johansson1986). Its correlation in Norway and Sweden is well established by acritarchs and trilobites (Vidal & Nystuen, Reference Vidal and Nystuen1990; Moczydlowska, Reference Moczydlowska1991; Ahlberg & Bergström, Reference Ahlberg and Bergström1993) and correlated with the lowermost part of the Atdabanian Stage (Moczydlowska, Reference Moczydlowska1991; Zhuravlev, Reference Zhuravlev1995).
In Laurentia, the oldest trilobites known are olenellids of the Fritzaspis Zone which belong to the uppermost part of the Begadean Stage, which is correlated with the lower Atdabanian Substage (A2; Hollingsworth, Reference Hollingsworth2005a , Reference Hollingsworth b , Reference Hollingsworth2007). A new correlation of the Fritzaspis Zone with the oldest trilobites of Spain, Siberia and Morocco was proposed by Hollingsworth (Reference Hollingsworth, Rábano, Gozalo and García Bellido2008).
Finally, the oldest trilobites of Argentina are from the Olenellus Zone of the Dyeran Stage from Laurentia (Bordonaro, Reference Bordonaro, Gutiérrez-Marco, Saavedra and Rábano1992, Reference Bordonaro2003) and are considered equivalent to the Botoman Stage.
A tentative intercontinental correlation of the first trilobite records is shown in Figure 8. It is broadly demonstrated and accepted that the oldest appearance of trilobite species (body fossils) were preceded by that of trilobite trace fossils in the fossil record, which typically appear as the following succession of ichnogenera: Monomorphichnus/Dimorphichnus/Diplichnites – Rusophycus – Cruziana. Regarding the Spanish Cambrian stratigraphic successions, Rusophycus ichnosp. precedes the oldest trilobites Serrania? gordaensis and Bigotina bivallata from SW and SE Sierra Morena (Liñán et al. Reference Liñán, Dies, Gámez Vintaned, Gozalo, Mayoral and Muñiz2005, Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b ); Cruziana ichnosp. precedes the appearance of Lunagraulos tamamensis in central Spain (this work); Rusophycus fasciculatus (Seilacher, Reference Seilacher, Crimes and Harper1970) (=R. avalonensis Crimes & Anderson, Reference Crimes and Anderson1985) and Cruziana cantabrica Seilacher, Reference Seilacher, Crimes and Harper1970 precede the oldest trilobite Thoralaspis n. sp. in the Cadenas Ibéricas (Iberian Chains) (Liñán et al. Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned, Mayoral, Chirivella Martorell, Esteve, Zamora, Zhuravlev and Andrés2008a ); Rusophycus cf. avalonensis has been reported to precede the trilobite cf. Abadiella bourgini (Jensen, Palacios & Martí Mus, Reference Jensen, Palacios and Mus2010); and Rusophycus ichnosp. precedes the oldest trilobite Lunolenus? lotzei in the Cantabrian Mountains (Gámez Vintaned et al. Reference Gámez Vintaned, Liñán, Mayoral, Gozalo, Zamora and Fernández-Martínez2006b ).

Figure 8. Intercontinental correlation of the oldest trilobite records (after Liñán et al. Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b ).
An explanation for the timing of the trilobitic trace and body fossils appearance described above is provided by the scenario in which flooding of extensive areas during the first stages of the worldwide Cambrian transgression initially led to conglomeratic and then to coarse-grained, sandy marine sedimentation. The preservation potential of trace fossils in basal Cambrian sandstones is high, while body fossil occurrences are very limited or exceptional as for Lunagraulos antiquus (Sdzuy, Reference Sdzuy1961) analysed here.
The lithological and taphonomic characteristics of lowermost Cambrian successions may therefore explain why the first trilobite trace fossil assemblages are abundant and older than the first trilobite body fossils, and also why the first appearances of the latter are diachronous and represented by different taxa in different palaeocontinents.
5. Conclusions
A study of trilobites from the lowermost Cambrian siliciclastic formations of northern and central Spain (the La Herrería Formation and the Tamames Sandstone, respectively), which are located below the lower Cambrian carbonate lithosome (Zamarreño, Reference Zamarreño1978), shows that the species of the family Agraulidae are also among the oldest trilobites in the world, and that Lunagraulos tamamensis may be equivalent in age to the oldest trilobite records of southern Spain, Siberia and Baltica.
The two species of Lunagraulos are found in microconglomeratic sandy and silty facies and represented by well-preserved smooth cranidia and even one complete specimen. They were therefore adapted to high- to medium-energy environments, where they probably developed opportunistic strategies living on coarse-grained substrates. Their presence in coarse siliciclastic facies may explain the scarcity of specimens in the geological record, and highlights the exceptionality of these findings.
As the Cambrian transgression progressed, biohermal palaeoenvironments were developed nearly worldwide and Gondwanan trilobite genera such as Bigotina, Serrania, Lemdadella, Eoredlichia, Eofallotaspis, Hupetina, Parabadiella and Fallotaspis appeared; they have short stratigraphical distributions, probably as a result of rapid evolution in the new habitats, characterized by carbonated and mixed siliciclastic-carbonated substrates. As a result it is possible to establish phylogenetic lineages starting from these first appearances, which are more precise than those made for the oldest trilobites (as for the Bigotina–Lemdadella–Eoredlichia lineage and the Serrania species lineage; Jell, Reference Jell2003; Liñán et al. Reference Liñán, Gozalo, Dies Álvarez, Gámez Vintaned and Zamora2008b ). These trilobite lineages are useful for erecting phylozonations for middle early Cambrian times (Liñán et al. Reference Liñán, Gámez Vintaned, Gozalo, Dies and Mayoral2006) and allow a better intercontinental correlation (Figs 7, 8).
The presence of the species Lunagraulos antiquus (Sdzuy, Reference Sdzuy1961) and Lunagraulos tamamensis n. gen. n. sp. in lower Cambrian successions contributes to increase the knowledge of the primitive trilobites, which are observed to be surprisingly well diversified in different provinces and facies since their first records.
Acknowledgements
This is a contribution to the CGL2011/24516 Project supported by the Spanish Ministry of Science and Innovation and to the E17 Research Group of the Autonomous Government of Aragón. Thanks are due to Ms Isabel Pérez Urresti (technician of the University of Zaragoza) who assisted us with the figures, and to Dr Markus Bertling (Geomuseum der Westfälische Wilhelms-Universität Münster) who provided replicae of the type material of Agraulos antiquus Sdzuy, Reference Sdzuy1961. Two anonymous referees are thanked for their constructive reviews.








