


1. Introduction
The Jurassic redbeds in the Sichuan Basin have yielded an important turtle fauna. Their earliest discovery is dated back to the 1950s. In 1951, a series of turtle shells were discovered during the construction of the railway between the cities of Chengdu and Chongqing (Chengyu railway) and the clearing away of building foundations in Datianwan, in the suburb of Chongqing. Young and Chow studied the material and created three monotypic genera (Chengyuchelys, Tienfuchelys and Sinaspideretes) and three new species of Plesiochelys (Young & Chow, Reference Young and Chow1953). The taxonomy of these turtles in Young & Chow's work (Reference Young and Chow1953) is summarized as follows:
Amphichelyidia
Baenidae?
Chengyuchelys
C. baenoides
Plesiochelyidae
Plesiochelys
P. latimarginalis
P. radiplicatus
P. chungkingensis
Tienfuchelys
T. tzuyangensis
Pleurodira
Trionychidae
Sinaspideretes
S. wimani
All these specimens are from the redbeds of the Xiashaximiao or Shangshaximiao formations, dated as Middle Jurassic and Late Jurassic, respectively. But for most of them, the exact locality and horizon are unknown; they may come from either the Middle Jurassic Xiashaximiao Formation or the Late Jurassic Shangshaximiao Formation. Of these taxa, we are particularly interested in Chengyuchelys baenoides in the present paper. Other turtles will be reviewed in separate papers.
All subsequent discoveries of Middle Jurassic turtles in the Sichuan Basin are from the Dashanpu locality, Zigong, Sichuan Province (Fig. 1). Since the initial discovery in 1972, to date Dashanpu has yielded about 20 turtle shells. The first study on Dashanpu turtles by Ye in 1982 includes three shells (CMNH C.V. 00678, CMNH C.V. 00679 and IVPP V6507). Ye created Chengyuchelys zigongensis on the basis of CMNH C.V.00678, referred IVPP V6507 to Chengyuchelys baenoides and CMNH C.V. 00679 to Chengyuchelys sp. (Ye, Reference Ye1982). In 1987, Fang studied four shells from Dashanpu, erected Chengyuchelys dashanpuensis on the basis of one of them (field number I-T19), referred I-Z1 to Chengyuchelys zigongensis and two others (I-L1 and I-L2) to Chengyuchelys sp. (Fang, Reference Fang1987). Later in his second paper on Dashanpu turtles, Ye reviewed all specimens from Dashanpu available to him at that time, including about 20 shells housed in the Zigong Dinosaur Museum (Ye, Reference Ye1990b). The family of Chengyuchelyidae was created on that occasion (in Chinese) and also in a separate paper in English published in the same year (Ye, Reference Ye1990a). The systematics of the Chengyuchelyidae and specimens from Dashanpu are repeated in Ye's monograph on Chinese fossil and recent turtles (Ye, Reference Ye1994). Some years later, Ye and Pi reported three shells from Dashanpu (one of them was mentioned in Ye, Reference Ye1990a) and created Sichuanchelys, a second genus of Chengyuchelyidae (Ye & Pi, Reference Ye and Pi1997).

Figure 1. Map showing the location of Dashanpu in the Zigong region and the Chengyu Railway.
In recent years, turtle specimens from the Middle Jurassic Dashanpu locality have been inventoried (Peng et al. Reference Peng, Ye, Gao, Shu and Jiang2005) and two specimens housed in the Institute of Vertebrate Palaeontology and Palaeoanthropology, Beijing (IVPP) collection have been re-described (Danilov & Parham, Reference Danilov and Parham2008). Danilov & Parham (Reference Danilov and Parham2008) noted that there are no shared derived characters uniting Chengyuchelys and Sichuanchelys in the Chengyuchelyidae and considered Chengyuchelys dashanpuensis as ‘Chengyuchelys’ dashanpuensis. Brinkman, Li & Ye (Reference Brinkman, Li, Ye, Li, Wu and Zhang2008) in a book on Chinese reptiles followed mostly previous work, but moved Chengyuchelys dashanpuensis to Sichuanchelys because of the wide vertebral scutes. However, the type series of Chengyuchelys baenoides has never been re-examined.
The history of studies on Dashanpu turtles and the type series of Chengyuchelys baenoides is summarized in Table 1. The turtle specimens described by Young and Chow in 1953, contrary to their usual citation as being in the IVPP collection (Young & Chow, Reference Young and Chow1953; Meylan & Gaffney, Reference Meylan and Gaffney1992; Ye, Reference Ye1994; Peng et al. Reference Peng, Ye, Gao, Shu and Jiang2005; Brinkman, Li & Ye, Reference Brinkman, Li, Ye, Li, Wu and Zhang2008; Danilov & Parham, Reference Danilov and Parham2008), are all housed in the Chongqing Museum of Natural History. Here we present a review of the Middle Jurassic turtle fauna from the Sichuan Basin. The type series of Chengyuchelys baenoides is re-examined. The aim of the present work is to make a detailed description of the specimens in order to review the validity of the taxa and their phylogenetic relationships in a contemporary systematic framework.
Table 1. History of studies on the Middle Jurassic turtle specimens from the Sichuan Basin

In this paper we use traditional nomenclature (ICZN) for turtle taxa (Gaffney & Meylan, Reference Gaffney, Meylan and Benton1988; Gaffney, Reference Gaffney1996) instead of rank-free nomenclature (Joyce, Parham & Gauthier, Reference Joyce, Parham and Gauthier2004), followed in the paper of Danilov & Parham (Reference Danilov and Parham2008). Anatomical terms of the shell follow Zangerl (Reference Zangerl, Gans, Bellairs and Parsons1969).
2. Geological background of Dashanpu
The Sichuan Basin, formerly the Sichuan Plateau on the east coast of Tethys, was formed during the Late Triassic Indosinian orogeny. It is situated in the eastern part of the former Sichuan Province, which has been divided into Sichuan Province and the Municipality of Chongqing since 1997. The Jurassic continental deposits are extensively developed in the basin, reaching 1000–2000 m in thickness. From the bottom to the top, the Jurassic sediments are divided into seven formations: the Lower Jurassic Zhenzhuchong and Ziliujing formations, the Middle Jurassic Xintiangou and Xiashaximiao formations, and the Upper Jurassic Shangshaximiao, Suining and Penglaizhen formations. Dashanpu, in the Zigong region, is situated in the southern part of the Sichuan Basin (Fig. 1). The Xiashaximiao Formation in the Zigong area is composed of 90–217 m of purplish red mudstones with 2–3 grey, yellowish grey fine- to medium-gained feldspathic quartzitic sandstone intercalations. The unit is characteristic of alternative fluvial and lacustrine facies deposited under a subtropical–tropical climate, showing relative warm and moist ecological conditions. It overlies unconformably the Middle Jurassic Xintiangou or Ziliujing Formation and underlies conformably the Upper Jurassic Shangshaximiao Formation. In Dashanpu, the sandstones in the lower part of the Xiashaximiao Formation have yielded abundant vertebrate remains belonging to the Shunosaurus fauna, including selachians, bony fishes, amphibians, turtles, plesiosaurs, crocodiles, pterosaurs, mammal-like reptiles and theropod, sauropod, ornithopod and stegosaur dinosaurs (Dong, Reference Dong1992; Peng et al. Reference Peng, Ye, Gao, Shu and Jiang2005; Wang et al. Reference Wang, Liang, Kan, Li, Zhu and Ji2008).
3. Material
The specimens examined in this work are listed in Table 1, which includes all available specimens from the Middle Jurassic of the Sichuan Basin housed in three important collections: Zigong Dinosaur Museum (ZDM), Chongqing Museum of Natural History (CMNH) and Institute of Vertebrate Palaeontology and Palaeoanthropology, Beijing (IVPP).
The first-hand comparisons are made with the Late Jurassic turtles from the Sichuan Basin, which are referred to ‘Plesiochelys’ or Xinjiangchelys: ‘Plesiochelys’ latimarginalis Young & Chow, Reference Young and Chow1953, ‘P’. radiplicatus Young & Chow, Reference Young and Chow1953, ‘P’. chungkingensis Young & Chow, Reference Young and Chow1953, ‘P’ jingyanensis Ye & Fang, Reference Ye and Fang1982, Tienfuchelys tzuyangensis Young & Chow, Reference Young and Chow1953 (Young & Chow, Reference Young and Chow1953; Ye, Reference Ye1963, Reference Ye1986b, Reference Ye1994; Ye & Fang, Reference Ye and Fang1982; Peng & Brinkman, Reference Peng and Brinkman1993; Matzke et al. Reference Matzke, Maisch, Ge and Pfretzschner2004, Reference Matzke, Maisch, Sun, Pfretzschner and Stöhr2005; Peng et al. Reference Peng, Ye, Gao, Shu and Jiang2005; Brinkman, Li & Ye, Reference Brinkman, Li, Ye, Li, Wu and Zhang2008); Xinjiangchelys junggarensis Ye, Reference Ye1986a and Xinjiangchelys? oshanensis (Ye, Reference Ye1973, Reference Ye1986a; Peng & Brinkman, Reference Peng and Brinkman1993; Brinkman, Li & Ye, Reference Brinkman, Li, Ye, Li, Wu and Zhang2008).
Other comparison materials are from the literature: Condorchelys antiqua Sterli, Reference Sterli2008 (Sterli, Reference Sterli2008; Sterli & de la Fuente, Reference Sterli and de la Fuente2010); Dinochelys whitei Gaffney, Reference Gaffney1979 (Gaffney, Reference Gaffney1979; Brinkman, Stadtman & Smith, Reference Brinkman, Stadtman and Smith2000); Eileanchelys waldmani Anquetin, Barrett, Jones, Moor-Fay & Evans, Reference Anquetin, Barrett, Jones, Moor-Fay and Evans2008 (Anquetin et al. Reference Anquetin, Barrett, Jones, Moor-Fay and Evans2008; Anquetin, Reference Anquetin2010); Glyptops plicatulus (Cope, Reference Cope1877) (Gaffney, Reference Gaffney1979); Heckerochelys romani Sukhanov, Reference Sukhanov2006 (Sukhanov, Reference Sukhanov2006); Hylaeochelys latiscutata (Owen) Lydekker, Reference Lydekker1889 (Milner, Reference Milner2004); Indochelys spatulata Datta, Ghosh & Das, Reference Datta, Manna, Ghosh and Das2000 (Datta et al. Reference Datta, Manna, Ghosh and Das2000); Kayentachelys aprix Gaffney, Hutchison, Jenkins & Meeker, Reference Gaffney, Hutchison, Jenkins and Mekker1987 (Gaffney et al. Reference Gaffney, Hutchison, Jenkins and Mekker1987); Kirgizemys hoburensis (Sukhanov & Narmandakh, Reference Sukhanov and Narmandakh1974) (Sukhanov, Reference Sukhanov, Benton, Shishkin, Unwin and Kurochkin2000; Danilov et al. Reference Danilov, Averianov, Skutchas and Rezvyi2006); Mongolochelys efremovi Khosatzky, Reference Khosatzky1997 (Khosatzky, Reference Khosatzky1997); Ordosemys liaoxiensis (Ji, Reference Ji, Ren, Lu, Guo and Ji1995) (Tong, Ji & Ji, Reference Tong, Ji and Ji2004); Palaeochersis talampayensis Rougier, de la Fuente & Arcucci, Reference Rougier, Fuente and Arcucci1995 (Rougier, de la Fuente & Arcucci, Reference Rougier, Fuente and Arcucci1995; Sterli, de la Fuente & Rougier, Reference Sterli, de la Fuente and Rougier2007); Plesiochelys solodurensis (Bräm, Reference Bräm1965); Pleurosternon bullockii (Owen) Lydekker & Boulenger, Reference Lydekker and Boulenger1887 (Milner, Reference Milner2004); Proganochelys quenstedti Baur, Reference Baur1887; Proterochersis robusta Fraas, Reference Fraas1913 (Gaffney, Reference Gaffney1990); Siamochelys peninsularis Tong, Buffetaut & Suteethorn, Reference Tong, Buffetaut and Suteethorn2002 (Tong, Buffetaut & Suteethorn, Reference Tong, Buffetaut and Suteethorn2002); Xinjiangchelys junggarensis, X. chowi, X. qiquensis, X . tianshanensis (Ye, Reference Ye1986a; Kaznyshkin, Nalbandyan & Nessov, Reference Kaznyshkin, Nalbandyan and Nessov1990; Peng & Brinkman, Reference Peng and Brinkman1993; Nessov, Reference Nessov1995; Matzke et al. Reference Matzke, Maisch, Ge and Pfretzschner2004, Reference Matzke, Maisch, Sun, Pfretzschner and Stöhr2005; Brinkman, Li & Ye, Reference Brinkman, Li, Ye, Li, Wu and Zhang2008).
4. The problem of Chengyuchelys baenoides Young & Chow, Reference Young and Chow1953
The type species of Chengyuchelys, C. baenoides, was based on two shells: CMNH C.996 (V708 in Young & Chow, Reference Young and Chow1953, holotype), an incomplete shell with articulated carapace and plastron; and CMNH C.997 (V710 in Young & Chow, Reference Young and Chow1953, paratype), a shell of a juvenile in which the carapace is mostly preserved as an internal mould, articulated with an incomplete plastron. Both specimens came from sites along the Chengyu Railway and based on the matrix surrounding the fossils, it is supposed that they are from the same stratigraphical layer and close localities, and considered Late Jurassic to Cretaceous in age (Young & Chow, Reference Young and Chow1953). Young & Chow (Reference Young and Chow1953) indicated the presence of a pair of mesoplastra that is narrowed in the middle and included in the abdominal scute. Based on the presence of the mesoplastron, the general morphology of the shell, as well as the structure of the bridge, which resemble Baena from North America, the taxon was tentatively placed in the family Baenidae (Young & Chow, Reference Young and Chow1953).
The re-examination of these two specimens by one of us (H. T.) reveals that both CMNH C.996 and CMNH C.997 lack the mesoplastron. CMNH C.996 has a well-preserved hyoplastron–hypoplastron suture that can be followed from the broken right margin to the complete left bridge. However, the supposed posterior suture of the mesoplastron (Young & Chow, Reference Young and Chow1953, plate 1, fig. 2) is absent (Fig. 2a–d). The case is similar in CMNH C.997: a clear suture between the hyoplastron and hypoplastron can be followed from the left to the right, but no other suture is visible between it and abdominofemoral sulcus (Fig. 2e–g), contrary to Young & Chow (Reference Young and Chow1953, fig. 1).

Figure 2. Type series of Chengyuchelys baenoides Young & Chow, Reference Young and Chow1953. (a–d) CMNH C.996 (holotype, V. 709 in Young & Chow, Reference Young and Chow1953), shell in dorsal (a, b) and ventral (c, d) views; (e–g) CMNH C.997 (paratype, V.710 in Young & Chow, Reference Young and Chow1953), shell in dorsal (e) and ventral (f, g) views. Scale bar = 50 mm.
Comparisons with other turtles from the Middle and Upper Jurassic beds of the Sichuan Basin provide additional information. CMNH C.996 is quite different from turtles from the Middle Jurassic of Dashanpu, but presents the following features shared with the Late Jurassic turtles from the Sichuan Basin previously referred to ‘Plesiochelys’ and Xinjiangchelys: third to fifth marginals extending onto the costals; fourth vertebral only slightly wider than the third vertebral; wide anterior and posterior lobe; entoplastron relatively broad with straight posterolateral margins; humeropectoral sulcus close to the base of the anterior lobe; pectoral slightly shorter than abdominal. CMNH C.997 differs from CMNH C.996 in the absence of the midline groove on the carapace, the shape of the plastral lobes, especially the posterior lobe which is long and narrow, and the more anteriorly placed humeropectoral sulcus. Thus, both CMNH C.996 and CMNH C.997 belong to the Xinjiangchelyidae. We refrain from more precise assignment of these specimens pending revision of the Xinjiangchelyidae. Based on their close resemblance to the Late Jurassic turtles of the Sichuan Basin, these specimens likely come from the Upper Jurassic Shangshaximiao Formation.
The close examination of the Middle Jurassic turtles from Dashanpu shows that most of them do have a pair of mesoplastra, which meet on the midline. A new genus Bashuchelys is erected below with ‘Chengyuchelys’ zigongensis as the type species. A new species Bashuchelys youngi is also erected for turtles from Dashanpu that have been previously placed in the species Chengyuchelys baenoides Young & Chow, Reference Young and Chow1953, which have a pair of mesoplastra that is narrowed at the midline and covered mostly by the abdominal scute.
5. Systematic palaeontology
Order TESTUDINES Linnaeus, Reference Linnaeus1758
Infraorder CRYPTODIRA Cope, Reference Cope1868
Family Bashuchelyidae n. fam.
Diagnosis. Primitive Cryptodira with the following combination of characters: one pair of mesoplastra present (1), which are meeting on the midline (2), with the hyoplastron–mesoplastron suture very close to the pectoroabdominal sulcus in the middle (3); regular and elongate neural series (4); wide fourth vertebral scute (5); sulcus between third and fourth vertebrals on sixth neural (6); plastron sutured to carapace (7); bridge fan-shaped with long axillary and inguinal buttresses (8); epiplastra midline contact present (9); humeropectoral sulcus lying far anterior to axillary notch (10); pectoral scute notably longer than abdominal (11); femoroanal sulcus omega-shaped, reaching or extending onto the hypoplastron (12); four inframarginals (13). Differs from Kayentachelyidae and Indochelyidae by characters 3, 4, 8, 9 and 12. Differs from Heckerochelyidae by characters 4, 6, 7, 11 and 12. Differs from Xinjiangchelyidae by characters 1–3.
Included genera. Bashuchelys n. gen., Chuannanchelys n. gen.
Genus Bashuchelys n. gen.
Type species. Bashuchelys zigongensis (Ye, Reference Ye1982).
Etymology. Bashu means Bashan (Ba Mountain) in the Chongqing area and Shushui (the river system on the plain of Sichuan); alternative Chinese name for Sichuan–Chongqing area; chelys: Greek for turtle.
Diagnosis. A genus of the Bashuchelyidae with shell length up to 250 mm; carapace elongate oval and low domed (1); 8 neurals (2); 2 suprapygals (3), the first larger and wider than the second (4); narrow first to third vertebrals (5); fourth vertebral scute twice as wide as it is long and notably wider than the third vertebral (6); anterior edge of the plastron with one anterior intergular projection and a pair of lateral gular projections (7); short epiplastra midline contact (8); entoplastron leaf-shaped with serrated posterolateral margins (9); one pair of intergulars extending deeply onto the entoplastron (10) (modified from Danilov & Parham, Reference Danilov and Parham2008). Differs from Chuannanchelys by characters 5–7, 9 and 10.
Distribution. Middle Jurassic of the Sichuan Basin, China.
Included species. B. youngi n. sp.; B. zigongensis (Ye, Reference Ye1982).
Bashuchelys youngi n. sp.Figure 3
1953 Chengyuchelys baenoides Young & Chow, p. 217, figs 1, 2; pl. 1, figs 1, 2.
1963 Chengyuchelys baenoides Young & Chow; Ye, p. 8.
1982 Chengyuchelys baenoides Young & Chow; Ye, p. 286, fig. 2.
1990a Chengyuchelys baenoides Young & Chow; Ye, p. 34, fig. 1.
1990b Chengyuchelys baenoides Young & Chow; Ye, p. 305.
1994 Chengyuchelys baenoides Young & Chow; Ye, p. 5, fig. 2.
2005 Chengyuchelys baenoides Young & Chow; Peng et al., p. 37, fig. 23.
2008 Chengyuchelys baenoides Young & Chow; Danilov & Parham, p. 307, 316, fig. 2.
2008 Chengyuchelys baenoides Young & Chow; Brinkman, Li & Ye, p. 35, fig. 35.

Figure 3. Bashuchelys youngi n. gen., n. sp. from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. ZDM 3007 (holotype), shell in dorsal (a, b) and ventral (c, d) views. Scale bar = 50 mm.
Etymology. In honour of Yang Zhong-Jian (C. C. Young) who first studied Jurassic turtles from the Sichuan Basin.
Holotype. ZDM 3007 (field number ZDM 1), a shell with articulated damaged carapace and almost complete plastron.
Referred material. IVPP V6507, anterior portion of a shell with articulated carapace and plastron, and fragments of left pectoral girdle and humerus; ZDM 3003, a shell with articulated carapace and plastron, carapace surface damaged, plastron lacking anterior and posterior ends; ZDM3006 (field number ZDM 7), a shell with articulated damaged carapace and plastron; ZDM 3008 (field number ZDM 3), posterior half of plastron.
Type locality and horizon. Dashanpu, Zigong, Sichuan Province, China; Xiashaximiao Formation, Middle Jurassic.
Diagnosis. A species of Bashuchelys, different from B. zigongensis in the mesoplastron being strongly narrowed in the middle and mostly included in the abdominal scute, except laterally.
Measurements. See Table 2.
Table 2. Measurements (in mm) of turtle specimens from the Middle Jurassic Xiashaximiao Formation, Sichuan Basin

Description and comparisons. The carapace is low and has a narrow oval outline. A light midline depression is visible in ZDM 3003 and ZDM 3006, but not in IVPP V6507 and ZDM 3007. As preserved in IVPP V6507, radiating ridges are present on the vertebral region. Other specimens have a worn or damaged shell surface; thus no obvious ornamentation is visible. A shallow cervical notch is preserved in IVPP V6507 (Danilov & Parham, Reference Danilov and Parham2008), which is smaller than in ‘Plesiochelys’ jingyanensis (Ye & Fang, Reference Ye and Fang1982).
The carapace is damaged in all specimens; the best preserved is IVPP V6507 and ZDM 3007. The nuchal is complete only in IVPP V6507. It is trapezoidal and as wide as the first vertebral scute. In IVPP V6507, only the first, second and anterior end of the third neurals are preserved, and the outline of the fourth through seventh in ZDM 3007 and the first to sixth neurals in ZDM 3003 can be traced out. The first neural, as preserved in IVPP V6507 and ZDM 3003, is hexagonal with short posterolateral sides. The second neural is rectangular in ZDM 3003 and contacts the second costal plate only, while the second neural in IVPP V6507 has five sides, with a short posterolateral side contacting the right third costal plate. The third to sixth neurals are all elongate and hexagonal with short anterolateral sides. This elongated neural pattern differs from the short and irregular neurals of Kayentachelys (Gaffney et al. Reference Gaffney, Hutchison, Jenkins and Mekker1987) and Indochelys (Datta et al. Reference Datta, Manna, Ghosh and Das2000). The suprapygal and pygal are not preserved in any specimen. The first costal plate, as preserved in IVPP V6507 and ZDM 3006, is short as in many primitive turtles. It is shorter than the second costal plate and contacts the first to third peripherals. The second costal has almost parallel anterior and posterior margins, while the third costal is longer laterally than medially in IVPP V6507, ZDM 3006 and ZDM 3007. The peripherals are better preserved in ZDM 3006 and IVPP V6507. There are 11 peripherals. The first peripheral is almost triangular with a short contact with the first costal. The second peripheral is roughly as long as wide. The third to seventh peripherals are longer than wide. The eighth and ninth peripherals are mesiolaterally expanded. The tenth and eleventh peripherals are clearly smaller than the ninth. This peripheral morphology is similar to that of Bashuchelys zigongensis. In comparison, in ‘P.’ jingyanensis the second to sixth peripherals are shorter and the seventh to eleventh peripherals are mesiolaterally expanded. There is a narrow lateral carapacial fontanelle between the peripherals and costals in IVPP V6507, presumably a juvenile character. This fontanelle is absent in all other specimens (ZDM 3003, ZDM 3006, ZDM 3007).
The cervical scute is preserved only in IVPP V6507; it is triple as reported by Danilov & Parham (Reference Danilov and Parham2008). The first vertebral is complete in IVPP V6507, and partly preserved in ZDM 3003, ZDM 3006 and ZDM 3007. It is wide and hexagonal, being as wide as the nuchal and contacting the second marginal laterally. The second vertebral is roughly as long as it is wide (IVPP V6507) or slightly longer than wide (ZDM 3003). The third vertebral is as wide as the second, as seen in ZDM 3003 and ZDM 3007. The fourth vertebral scute, although only partly preserved in ZDM 3006 and ZDM 3007, is much wider than the third vertebral scute, as in B. zigongensis (Ye, Reference Ye1982). The fifth vertebral is again narrow, as seen in ZDM 3006. The intervertebral sulci pass through the first neural (IVPP V6507, ZDM 3003), the third neural (ZDM 3003, ZDM 3007) and likely the sixth neural (ZDM 3007), as in B. zigongensis, Kayentachelys aprix and Eileanchelys waldmani, in contrast to Pleurosternon bullockii and Glyptops plicatulus. The first to third pleural scutes are almost as long as wide. The fourth pleural scute is very narrow as in B. zigongensis, a consequence of the enlarged fourth vertebral scute. The first to seventh marginals are preserved in IVPP V6507. Most marginals are preserved in ZDM 3006, and only a few of them are visible in ZDM 3003 and ZDM 3007. The second to fourth marginals are restricted in the peripherals. The fifth marginal extends onto the second costal and the seventh marginal extends onto the fourth costal plate in IVPP V6507 and ZDM 3003, but not in ZDM 3006. As preserved in IVPP V6507, the sixth marginal is clearly narrower than the fifth and the seventh, with its medial sulcus matching the lateral carapacial fontanelle border. In comparison, all lateral marginals are restricted in the peripherals in Kayentachelys aprix and Sichuanchelys chowi, and the fourth through seventh marginals extend onto the costals in ‘P.’ jingyanensis and other xinjiangchelyids.
The plastron is sutured to the carapace in ZDM 3007, ZDM 3006 and ZDM 3003, with the anterior margin reaching or slightly beyond the anterior rim of the carapace. IVPP V6507 has a loose plastron–carapace attachment (Danilov & Parham, Reference Danilov and Parham2008). The whole morphology of the plastron is similar to that of B. zigongensis. The plastron is elongated, with both anterior and posterior lobes, which are long and have a narrow end. The posterior lobe is longer than the anterior lobe and without an anal notch. The bridge is roughly as long as the posterior lobe, narrow and fan-shaped. As in B. zigongensis, the axillary buttress is long, reaching the second peripheral. The inguinal buttress touches the eighth peripheral. As preserved in IVPP V6507 and ZDM 3007, the plastron has an anteriorly directed intergular projection in the middle and a pair of the gular projections that are directed laterally. Such projections are absent in other Jurassic turtles (e.g. Kayentachelys aprix, Heckerochelys romani). Three musk ducts are visible on the right side in ZDM 3007, on or close to the plastron–carapace suture.
The epiplastron, completely preserved only on the right side of ZDM 3007 and left side of IVPP V6507, has a short midline suture to its mate, preventing the exposure of the entoplastron on the plastron front margin. The entoplastron is complete in IVPP V6507 and nearly so in ZDM 3007. It is leaf-shaped, with serrated posterolateral margins. The entoplastron separates the posterior two thirds of the epiplastron and does not reach the anterior border of the plastron, in contrast to Kayentachelys aprix and Indochelys spatulata. The mesoplastron is well preserved in ZDM 3007, ZDM 3003 and ZDM 3006, with clear sutures. It is incomplete in IVPP V6507, lacking the left part. ZDM 3008 has the mesoplastron mostly preserved as an imprint on the internal mould. The pair of mesoplastra meets on the midline but is strongly narrowed in the middle. Laterally its length in the bridge region is about three times its midline length or more. The hyoplastron–mesoplastron suture is clearly convex posteriorly. The mesoplastron–hypoplastron suture is straight in ZDM 3007 and convex anteriorly in ZDM 3003, ZDM 3006 and IVPP V6507. The hypoplastron–xiphiplastron suture is convex posteriorly.
The gular is short and wide in ZDM 3007 and roughly as long as it is wide in IVPP V6507. The pair of intergulars is clearly longer than the gulars and extends deeply onto the entoplastron in both IVPP V6507 and ZDM 3007. The humeropectoral sulcus is only slightly convex posteriorly, lying posterior to the entoplastron and well anterior to the axillary notch. The pectoroabdominal sulcus is convex posteriorly in the middle and very close to the hyoplastron–mesoplastron suture. Laterally, it turns sharply in the posterolateral direction, crossing the hyoplastron–mesoplastron suture. The midline length of the femoral scute is longer than that of the abdominal. The abdominofemoral sulcus is convex anteriorly. The anal scute has an omega-shaped anterior margin and is overlapping the hypoplastron in ZDM 3007, ZDM 3003 and ZDM 3008, while in ZDM 3006 the anal scute is restricted to the xiphiplastron, and its anterior sulcus does not reach the hypoplastron–xiphiplastron suture. As preserved in ZDM 3007, ZDM 3003 and ZDM 3006, four inframarginals are present, the fourth being the largest.
Bashuchelys zigongensis (Ye, Reference Ye1982) Figures 4a–d, 5
1982 Chengyuchelys zigongensis Ye, p. 283, fig. 1.
1987 Chengyuchelys zigongensis Ye; Fang, p. 67, pl. 6.
1990a Chengyuchelys zigongensis Ye; Ye, p. 34, fig. 2.
1990b Chengyuchelys zigongensis Ye; Ye, p. 305.
1994 Chengyuchelys zigongensis Ye; Ye, p. 8, fig. 3.
2005 Chengyuchelys zigongensis Ye; Peng et al., p. 38, fig. 24.
2008 Chengyuchelys zigongensis Ye; Brinkman, Li & Ye, p. 36, fig. 36.
2008 Chengyuchelys zigongensis Ye; Danilov & Parham, p. 316.
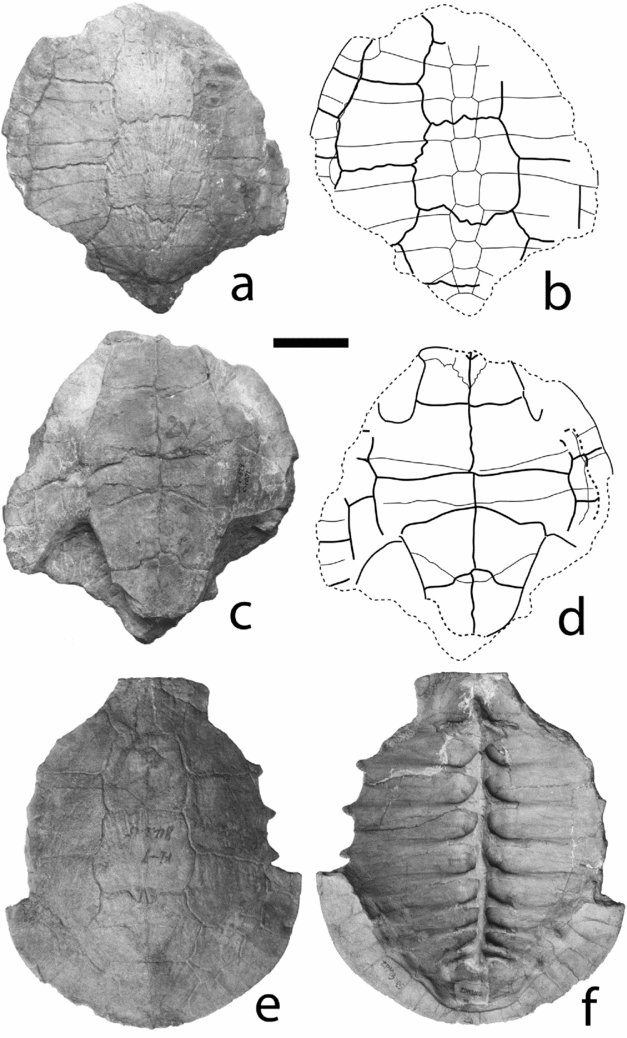
Figure 4. (a–d) Bashuchelys zigongensis (Ye, Reference Ye1982) from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. CMNH C.1001 (holotype), shell in dorsal (a, b) and ventral (c, d) views; (e, f) Bashuchelys sp. from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. ZDM 3005, carapace in dorsal (e) and ventral (f) views. Scale bar = 50 mm.

Figure 5. Bashuchelys zigongensis (Ye, Reference Ye1982) from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. ZDM 3012, shell in dorsal (a, b) and ventral (c, d) views. Scale bar = 50 mm.
Holotype. CMNH C.1001 (C. V. 00678 in Ye, Reference Ye1982), an incomplete shell with articulated carapace and plastron.
Referred material. ZDM 3012 (field number: I-Z1), an almost complete shell with articulated carapace and plastron, slightly deformed.
Type locality and horizon. Dashanpu, Zigong, Sichuan Province, China; Xiashaximiao Formation, Middle Jurassic.
Diagnosis. A species of Bashuchelys, different from B. youngi in the pair of mesoplastra that is not narrowed in the middle and covered by both pectoral and abdominal scutes.
Measurements. See Table 2.
Description and comparisons. CMNH C.1001 lacks the anterior and posterior end of the carapace. The carapace of ZDM 3012 is complete, and has an elongate oval outline. In comparison, ‘Plesiochelys’ latimarginalis and other xinjiangchelyids have a wider carapace (Young & Chow, Reference Young and Chow1953; Ye & Fang, Reference Ye and Fang1982; Ye, Reference Ye1986a; Peng & Brinkman, Reference Peng and Brinkman1993; Matzke et al. Reference Matzke, Maisch, Ge and Pfretzschner2004, Reference Matzke, Maisch, Sun, Pfretzschner and Stöhr2005). The carapace is low domed. The midline depression is absent in both CMNH C.1001 and ZDM 3012. The cervical notch is absent as preserved in ZDM 3012. The complete lateral margin of ZDM 3012 shows no gutter formed by the anterolateral peripherals, in contrast to Xinjiangchelys junggarensis (Peng & Brinkman, Reference Peng and Brinkman1993), while a light gutter is present on the bridge peripherals, as in IVPP V6507 (Danilov & Parham, Reference Danilov and Parham2008). The carapace surface is decorated by some radiating ridges in the vertebral region and in the anterolateral part, which are lighter than that of ‘Plesiochelys’ radiplicatus (Young & Chow, Reference Young and Chow1953). The surface of the plastron is rough.
The nuchal is preserved in ZDM 3012, which is trapezoidal with a long free margin. There are eight neurals in CMNH C.1001; the first is not complete. In ZDM 3012, the complete series of neurals is preserved, but the sutures of the third to fifth neurals are not all discernible. In CMNH C.1001, the first neural is likely rectangular; it does not contact the second costal plate. The second to eighth neurals are all hexagonal, with short anterolateral sides. They are all longer than wide except the seventh. In ZDM 3012, the first neural appears to be hexagonal, with short posterolateral sides to contact the second costal plates. The sixth and seventh neurals are longer than wide with short anterolateral sides. The eighth neural is nearly as long as wide. There are two suprapygals in ZDM 3012 and only part of the first suprapygal is preserved in CMNH C.1001. The first suprapygal is larger and wider than the second and contacts the eleventh peripheral laterally. The pygal is nearly complete in ZDM 3012; it is wider than long, with the anterior suture convex backward. The first costal plate as preserved in ZDM 3012 is short and narrower than the second costal, as in many primitive turtles. The second and third costals are slightly wider laterally than medially, as in IVPP V6507 (Danilov & Parham, Reference Danilov and Parham2008). Only the left second to sixth peripherals are preserved in CMNH C.1001. The complete series of 11 peripherals are preserved in ZDM 3012. The first peripheral is four-sided, with the length of its free margin twice that of its contact with the first costal plate. The second and third peripherals are as long as wide. The fourth to seventh peripherals are narrow. The eighth and ninth peripherals are mesiolaterally expanded. The tenth and eleventh peripherals are clearly smaller than the eighth and the ninth. In comparison, Xinjiangchelys junggarensis has the seventh through eleventh peripherals mesiolaterally expanded (Ye, Reference Ye1986a; Peng & Brinkman, Reference Peng and Brinkman1993).
As preserved in ZDM 3012, there is a single cervical scute which is wider than long, unlike the triple cervical in IVPP V6507 (Danilov & Parham, Reference Danilov and Parham2008). The cervical has posteriorly convergent lateral sulci; as a result its free margin is longer than its sulcus to the first vertebral scute. The first to third vertebrals are moderate in width, as in ‘P.’ latimarginalis (Young & Chow, Reference Young and Chow1953), and different from the very large vertebrals of Sichuanchelys chowi (Ye & Pi, Reference Ye and Pi1997) and Kayentachelys aprix (Gaffney et al. Reference Gaffney, Hutchison, Jenkins and Mekker1987). The first vertebral scute is complete in ZDM 3012. It is slightly wider than long and as wide as the second vertebral. The second and the third vertebrals are narrower anteriorly than posteriorly in ZDM 3012. In CMNH C.1001, the second vertebral is as wide anteriorly as posteriorly. The third vertebral is slightly wider than the second in both CMNH C.1001 and ZDM 3012. The fourth vertebral is the widest, which is much wider than the third one and nearly twice wider than it is long. This vertebral scute pattern is present in both CMNH C.1001 and ZDM 3012, and similar to Bashuchelys youngi. The fifth vertebral, as preserved in ZDM 3012, is almost as wide as the third vertebral and clearly wider than the first suprapygal. It covers the posterior part of the eighth neural and eighth costals, two suprapygals, the anterior part of the pygal and the eleventh peripheral plates. The intervertebral sulci pass through the first, the third, the sixth and the eighth neural plates, respectively. The sulcus passing the sixth neural is straight in ZDM 3012, but serrated and convex posteriorly in CMNH C.1001. The first pleural scute covers the first to fourth peripherals, but does not extend onto the nuchal, contrary to IVPP V6507 (Danilov & Parham, Reference Danilov and Parham2008). The second pleural scute is larger than the first and the third pleurals and notably wide, as indicated by Ye (Reference Ye1982). It is as long as wide and contacts the fourth to sixth peripherals. The third pleural scute is slightly longer than wide, and covers the seventh to ninth peripherals. The fourth pleural scute is very narrow; it covers the ninth and tenth peripherals. As preserved in ZDM 3012, there are 12 marginal scutes. The first to the third and the eighth to the twelfth marginals are much narrower than the corresponding peripherals, although the fourth to seventh marginals are nearly as wide as the peripherals. In comparison, the medial tips of the marginals extend onto costal two to five in Xinjiangchelys junggarensis (Peng & Brinkman, Reference Peng and Brinkman1993) and all marginals are narrower than peripherals in Kayentachelys aprix (Gaffney et al. Reference Gaffney, Hutchison, Jenkins and Mekker1987) and Sichuanchelys chowi.
The plastron is sutured to the carapace in both CMNH C.1001 and ZDM 3012, in contrast to IVPP V6507 (Danilov & Parham, Reference Danilov and Parham2008) and Xinjiangchelys junggarensis (Peng & Brinkman, Reference Peng and Brinkman1993). The plastron is elongate, with a long fan-shaped bridge and narrow axillary and inguinal notches. The plastron of Xinjiangchelys junggarensis is shorter, with shorter anterior and posterior lobes. The morphology of the anterior lobe is comparable to that of Bashuchelys youngi, although its anterior end is slightly damaged in both CMNH C.1001 and ZDM 3012. The gular projection is preserved on the left side of ZDM 3012 and the right side of CMNH C.1001. The intergular projection, as seen in Bashuchelys youngi, is not preserved in any specimen. The anterior margin of the plastron extends slightly beyond the anterior margin of the carapace, as in IVPP V6507. The anterior lobe is shorter than the posterior lobe. In ZDM 3012, the posterior lobe is long and narrow, with posteriorly convergent lateral margins. The posterior lobe in CMNH C.1001 is wider than that of ZDM 3012. The anal notch is absent. The axillary buttress is long, reaching the posterior part of the second peripheral. The inguinal buttress reaches the eighth peripheral.
The incomplete epiplastron is preserved on the right side of CMNH C.1001 and left side of ZDM 3012. It has a short contact with its mate. The entoplastron is complete in ZDM 3012 and lacks the anterior end in CMNH C.1001. It is leaf-shaped, with serrated sutures and a pointed posterior end. It does not reach the anterior margin of the plastron. The hyoplastron contacts the second through fifth peripherals. In CMNH C.1001, the mesoplastron is well preserved, with clear anterior and posterior sutures. In ZDM 3012, the sutures are more difficult to follow, especially the anterior one. In both specimens, the mesoplastron is large and not narrowed in the middle, covered by both pectoral and abdominal scutes. However, its anterior suture lies very close to the pectoroabdominal sulcus, as in Heckerochelys romani (Sukhanov, Reference Sukhanov2006). In Glyptops plicatulus, Dinochelys whitei, Kayentachelys aprix, Pleurosternon bullockii and Helochelydra anglica, the anterior border of the mesoplastron lies at about same distance to the pectoroabdominal sulcus as its posterior border (Gaffney, Reference Gaffney1979; Gaffney et al. Reference Gaffney, Hutchison, Jenkins and Mekker1987; Milner, Reference Milner2004). The mesoplastron contacts the fifth and sixth peripherals. The hypoplastron contacts the sixth through eighth peripherals. The hypoplastron–xiphiplastron suture is strongly convex posteriorly in both specimens.
The pair of intergulars extends deeply onto the entoplastron in both specimens as in Bashuchelys youngi. The humeral is long, with a straight humeropectoral sulcus, which lies far anterior to the axillary notch as in Bashuchelys youngi. The pectoral scute is clearly longer than the abdominal scute. In comparison, the humeropectoral sulcus lies close to the base of the anterior lobe and the pectoral and abdominal have a similar length in Xinjiangchelys junggarensis and ‘P.’ latimarginalis. The femoral is as long as the pectoral. The femoroanal sulcus is roughly omega-shaped, reaching the hypoplastron–xiphiplastron suture in ZDM 3012, and extending onto the hypoplastron in CMNH C.1001. As preserved in CMNHC.1001 and the left side of ZDM 3012, there are four inframarginals. The midline sulcus is straight in ZDM 3012 and slightly sinuous in CMNH C.1001.
Bashuchelys sp.
1990b Chengyuchelys sp. Ye, p. 306.
1994 Chengyuchelys sp. Ye, p. 12.
2005 Chengyuchelys sp. Peng et al., p. 42.
Referred material. ZDM 3004, incomplete carapace lacking peripherals; ZDM 3005 (Fig. 4e, f) and ZDM 3013, two incomplete carapaces lacking anterior and lateral peripherals; ZDM 3016, incomplete carapace with damaged surface, lacking some peripherals.
Locality and horizon. Dashanpu, Zigong, Sichuan Province, China; Xiashaximiao Formation, Middle Jurassic.
Description. The carapace is oval. The almost complete right margin in ZDM 3016 shows slightly upturned anterolateral margins with the carapace being wider posteriorly than anteriorly. ZDM 3005 and ZDM 3016 have a well-preserved anterior margin of the carapace, showing that the cervical notch is absent. The carapace surface has radiating ridges on the vertebral region in all four specimens. The free rib end is present on the lateral end of the costals in ZDM 3005. The sutures are not all discernible. The cervical is preserved in ZDM 3005; it is wider than long. The first to third vertebrals are moderate in width in ZDM 3005, ZDM 3013 and ZDM 3016, while the third vertebral in ZDM 3004 is wider. In all four specimens, the fourth vertebral scute is notably wider than other vertebrals. There is an additional small scute between the second and third vertebrals in ZDM 3004. As preserved in ZDM 3005 and ZDM 3016, the sulcus between the third and fourth vertebrals is W-shaped and lying on the sixth neural. The anterior and lateral marginals are longer than wide, and the posterior marginals are square. The sulci between the marginal and pleural scutes are visible on the second costal in ZDM 3005, and on the fourth costal in ZDM 3004 and ZDM 3005.
The inner surface of the carapace is exposed in ZDM 3005 (Fig. 4f). The first thoracic rib extends laterally to half the width of the first costal as in Xinjiangchelys junggarensis (Peng & Brinkman, Reference Peng and Brinkman1993; Brinkman, Li & Ye, Reference Brinkman, Li, Ye, Li, Wu and Zhang2008), in contrast to Ordosemys leios (Brinkman & Peng, Reference Brinkman and Peng1993). All rib heads are large and triangular. The axillary buttress reaches the lateral margin of the first costal, while the inguinal buttress inserts on the limit of the sixth costal and eighth peripheral.
Remarks. These specimens are referable to Bashuchelys in that the fourth vertebral scute is notably wider than the other vertebrals, closely resembling the two Bashuchelys species described above. As the two species of Bashuchelys are distinct only by the mesoplastron, these specimens are referred to Bashuchelys without specific assignment. ZDM 3004 and ZDM 3005 have been identified as Chengyuchelys sp. in Peng et al. Reference Peng, Ye, Gao, Shu and Jiang2005.
Genus Chuannanchelys n. gen.
Chuannanchelys dashanpuensis (Fang, Reference Fang1987) Figure 6
1987 Chengyuchelys dashanpuensis Fang, p.65, figs on p. 66, pl. VI, figs I-T19.
1990b Chengyuchelys dashanpuensis Fang; Ye, p. 306.
1994 Chengyuchelys dashanpuensis Fang; Ye, p.10, fig. 4.
2005 Chengyuchelys dashanpuensis Fang; Peng et al., p. 41, fig. 26.
2008 ‘Chengyuchelys’ dashanpuensis Fang; Danilov & Parham, p. 316.
2008 Sichuanchelys dashanpuensis (Fang); Brinkman, Li & Ye, p. 37, fig. 38.

Figure 6. Chuannanchelys dashanpuensis (Fang, Reference Fang1987) from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. ZDM 3011 (holotype), shell in dorsal (a, b) and ventral (c, d) views. Scale bar = 50 mm.
Etymology. Genus name from Chuannan (south Sichuan).
Holotype. ZDM 3011 (I-T19 in Fang, Reference Fang1987), an almost complete shell with articulated carapace and plastron, lacking part of peripherals.
Type locality and horizon. Dashanpu, Zigong, Sichuan Province, China; Xiashaximiao Formation, Middle Jurassic.
Diagnosis. A genus of the Bashuchelyidae, carapace length about 200 mm, with the following combination of characters: carapace elongated oval and low (1) with shallow cervical notch (2); all vertebrals broad, second to fourth vertebrals almost twice as wide as long (3); plastron with wide anterior lobe, and short and wide posterior lobe (4); one pair of reduced mesoplastra meeting on the midline and included entirely in the abdominal scute (5); epiplastron squarish (6) with long midline contact (7); entoplastron diamond-shaped and slightly longer than wide (8); intergulars excluded from entoplastron (9). Differs from Bashuchelys by characters 3–9.
Measurements. See Table 2.
Description and comparisons. Although the shell is well preserved, some sutures are not discernible. The carapace is slightly deformed. It is elongate oval and low. The cervical notch is wide and shallow, larger than in Bashuchelys spp., but clearly smaller than in Sichuanchelys chowi. There are some light wrinkles on the carapace surface, within the vertebral scutes.
The nuchal is trapezoid. There are eight neurals, with the neural formula of 6<4>6<6<6<6<6<6. They are all longer than wide, in contrast to mostly short and wide neurals in Kayentachelys aprix and Indochelys spatulata. The suprapygals and pygal are not discernible. There are eight costal plates. The first costal is shorter than the second. The third costal is wider laterally than medially as in Bashuchelys spp. The peripherals are not all discernible. The first peripheral is roughly rectangular with the second being as long as wide. Other peripheral sutures are not visible.
The cervical is short and wide, with its width two times its length, similar to Kayentachelys aprix. In contrast to Glyptops plicatulus, Bashuchelys spp. and xinjiangchelyids, all five vertebrals are wide as in Kayentachelys aprix, Indochelys spatulata, Heckerochelys romani, Dinochelys whitei and Eileanchelys waldmani. The first vertebral is wider than the nuchal and has a rather long contact with the second marginal. The second through fourth vertebrals are nearly twice as wide as long, different from Bashuchelys spp. in which only the fourth vertebral is wide. There are 12 marginals. The first marginal is longer than wide. The second marginal is as long as wide. The third through seventh marginals are elongate and the ninth to eleventh marginals are square.
In contrast to Heckerochelys romani, the plastron is sutured to the carapace as in Kayentachelys aprix and Eileanchelys waldmani. In comparison with Bashuchelys spp., the plastron of Chuannanchelys dashanpuensis is wider with wider anterior and posterior ends. The anterior lobe is longer than the posterior lobe but shorter than the bridge. The anterior lobe has a wide front end and smooth anterior border, without gular and intergular projections as seen in Bashuchelys youngi. The bridge is fan-shaped as in Bashuchelys spp. and xinjiangchelyids. The posterior lobe is wider and shorter than in Bashuchelys. The axillary and inguinal buttresses are long as in Bashuchelys. Three musk ducts are visible on the left side, on the plastron–carapace suture. A fourth one may be present (see Fig. 6d).
The epiplastron is large and squarish, with the transverse epiplastron–hyoplastron suture, unlike Sichuanchelys chowi. The entoplastron is ovoid, with a narrow posterior end. It is different from the leaf-shaped entoplastron of Bashuchelys spp. and the very small one of Sichuanchelys chowi. The hyoplastron is very long. A pair of mesoplastra is present, meeting on the midline. The mesoplastron is narrowed in the middle and entirely included in the abdominal scute, even laterally, in contrast to Bashuchelys youngi and Sichuanchelys chowi, in which the mesoplastron extends onto the pectoral laterally. The hypoplastron–xiphiplastron suture is convex posteriorly.
There are one pair of gulars and one pair of intergulars. In contrast to Bashuchelys spp. and Sichuanchelys chowi, the posterior border of the gulars and intergulars form a straight sulcus and are placed just before the entoplastron as in xinjiangchelyids. The humeropectoral sulcus is posterior to the entoplastron and lying far anterior to the axillary notch as in Bashuchelys spp. The pectoral scute is clearly longer than the abdominal. The femoroanal sulcus is omega-shaped and extends onto the hypoplastron as in Bashuchelys spp. and most xinjiangchelyids. Four inframarginals are preserved on the left side, the last one being the largest.
Family Xinjiangchelyidae Nessov in Kaznyshkin, Nalbandyan & Nessov, Reference Kaznyshkin, Nalbandyan and Nessov1990
Protoxinjiangchelys salis n. gen. et n. sp. Figure 7
1990b Chengyuchelys zigongensis Ye, p. 306.
1994 Chengyuchelys cf. zigongensis Ye, p. 9.
2005 Chengyuchelys zigongensis Ye; Peng et al., p. 40, fig. 25E–F.

Figure 7. Protoxinjiangchelys salis n. gen., n. sp. from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. ZDM 3009 (holotype), shell in dorsal (a, b) and ventral (c, d) views. Scale bar = 50 mm.
Etymology. Genus name: proto, first or primitive in Latin, in reference of its primitive status relative to Xinjiangchelys. Species name: salis, salt in Latin, in reference of the underground salt mine in the Zigong area.
Holotype. ZDM 3009, a complete shell with articulated carapace and plastron.
Type locality and horizon. Dashanpu, Zigong, Sichuan Province, China; Xiashaximiao Formation, Middle Jurassic.
Diagnosis. Primitive xinjiangchelyid with carapace length of about 200 mm; carapace oval and low (1), with shallow cervical notch (2); all vertebral scutes wide (3); plastron sutured to carapace (4), with narrow and long anterior and posterior lobes (5); epiplastron squarish with long midline contact (6); entoplastron elongate diamond-shaped (7); pair of intergulars excluded from entoplastron (8); humeropectoral sulcus lying far anterior to axillary notch (9); pectoral scute longer than abdominal (10); femoroanal sulcus reaching hypoplastron–xiphiplastron suture (11). Differs from bashuchelyids in the absence of the mesoplastron; differs from Xinjiangchelys by the characters 3, 4, 9 and 10; differs from Tienfuchelys by the characters 3, 7, 9 and 10; differs from Siamochelys by the characters 3, 4, 7, 9 and 10.
Measurements. See Table 2.
Description and comparisons. The carapace is slightly crushed, with the periphery regions damaged and restored. The plastron has the left bridge damaged.
The carapace is low and has an oval outline, which is wider than in Bashuchelys spp. The cervical notch is shallow and wide, comparable to that of Chuannanchelys dashanpuensis, but shallower than in ‘Plesiochelys’ jingyanensis. The carapace surface has no ornamentation. The nuchal is partly preserved, which appears to be similar to that of Chuannanchelys dashanpuensis. The first to fifth and incomplete sixth neurals are preserved, with the neural formula of 4>6>6>6>6>6. The first through fifth neurals are all elongate. The remaining neurals are not discernible, nor are the suprapygals and pygal. The costal plates are not all distinguishable. The first costal plate is short as in Bashuchelys spp. The peripheral plate sutures are not visible, except the medial suture of the seventh and eighth peripherals; they are slightly wider than the corresponding marginals.
The cervical is not preserved. The morphology of the vertebrals closely resembles that of Chuannanchelys dashanpuensis. The first and fifth vertebrals are narrower than the second vertebral. The second to fourth vertebrals are all clearly wider than long, being almost twice as wide as long. The intervertebral sulci pass through the first, the third and sixth neurals. The pleural scutes are longer than wide, except the second one which is square.
The morphology of the plastron resembles that of Bashuchelys spp. The plastron is sutured to the carapace. It is narrow and elongate as in Bashuchelys spp., but longer and narrower than in ‘P.’ jingyanensis. The anterior lobe is long and extends slightly beyond the anterior carapace margin as in Bashuchelys youngi. It is longer than the posterior lobe. The anterior margin of the plastron is slightly damaged, the gular and intergular projections as seen in Bashuchelys youngi seem to be absent; the margin is scallop-shaped as in ‘P.’ jingyanensis. The bridge is wide, fan-shaped and moderate in length. It is slightly longer than the anterior lobe. The posterior lobe is narrow, without an anal notch. The squarish epiplastron has a long midline suture to its mate. The entoplastron is diamond-shaped and longer than wide, with the anterolateral rims clearly shorter than the posterolateral ones; the posterolateral margin is straight on the right and only slightly undulated on the left, in contrast to Bashuchelys spp. The hyoplastron is longer than hypoplastron at the midline. The mesoplastron is absent.
The gular is smaller than the intergular. The posterior margin of gulars and intergulars is a straight sulcus situated at the anterior margin of the entoplastron as in ‘P.’ jingyanensis. In comparison, the intergulars in Bashuchelys spp. extend deeply onto the entoplastron. The humeropectoral sulcus is convex backwards, lying posterior to the entoplastron and far anterior to the axillary notch as in Bashuchelys spp. In ‘P.’ jingyanensis, this sulcus is close to the axillary notch. The pectoral is much longer than the abdominal at the midline, and similar in length to the femoral. The femoroanal sulcus is convex anteriorly and reaches the hypoplastron–xiphiplastron suture, but does not cut it. Four narrow inframarginal scutes are preserved on the right side, the last being the largest.
Protoxinjiangchelys sp.
1982 Chengyuchelys sp. Ye, p.286.
1990b Chengyuchelys sp. Ye, p.306.
1994 Chengyuchelys sp. Ye, p. 10.
2005 Chengyuchelys sp. Peng et al., p. 42.
Referred material. CMNH C.1002 (C.V. 00679 in Ye, Reference Ye1982), an incomplete shell with articulated carapace and plastron.
Remarks. Ye (Reference Ye1982) reported an incomplete shell with articulated carapace and plastron (C. V. 00679) from Dashanpu and referred it to Chengyuchelys sp. This specimen, now under the number of CMNH C.1002, is catalogued as being from the Late Jurassic of Jiulongpo, Chongqing. The specimen has broad vertebrals, although the third and fourth vertebrals are slightly wider than the second vertebral, different from Protoxinjiangchelys salis and Chuannanchelys dashanpuensis. The sulcus between the third and fourth vertebrals is on the fifth neural instead of on the sixth neural as in Bashuchelys spp., Protoxinjiangchelys salis and Chuannanchelys dashanpuensis. As in Bashuchelys spp. and Protoxinjiangchelys salis, the plastron has long anterior and posterior lobes; the humeropectoral sulcus lies far anterior to the axillary notch, the pectoral is longer than the abdominal, and the femoroanal sulcus is omega-shaped and extending onto the hypoplastron. The mesoplastron is absent as in Protoxinjiangchelys salis, in contrast to Bashuchelys spp. and Ch. dashanpuensis. Based on these characters, CMNH C.1002 is likely from the Middle Jurassic Dashanpu locality as indicated originally by Ye (Reference Ye1982). It is referred to Protoxinjiangchelys sp. because of the absence of the mesoplastron.
Family Sichuanchelyidae n. fam.
Diagnosis. Same as for Sichuanchelys.
Genus Sichuanchelys Ye & Pi, Reference Ye and Pi1997
Type species. S. chowi Ye & Pi, Reference Ye and Pi1997.
Emended diagnosis. Primitive turtle of small size, with a carapace length about 160 mm (1); carapace low, as long as wide and tapering posteriorly (2), with a large and deep cervical notch (3) and a shallow midline depression (4); shell surface without obvious ornamentation (5); most neurals elongate hexagonal with short anterolateral sides (6); peripherals very broad (7); vertebral scutes extremely broad, and second and third vertebrals with width being nearly three times length (8); pleural scutes narrow (9); all marginals narrow, lateral marginal scutes extremely narrow (10); plastron broad and sutured to carapace (11); anterior border of plastron slightly notched and made of a pair of weak intergular projections (12); one pair of mesoplastra present, which is narrowed in the midline and included mostly in the abdominal scute except laterally (13); epiplastron with very short midline contact (14) and long posterolateral process (15); entoplastron small and triangular and not reaching the anterior plastral margin (16); one pair of intergulars extending onto the entoplastron (17); anal scute included in the xiphiplastron (18); four inframarginals (19). Differs from Bashuchelys by the characters 1–3, 7–10, 12, 15, 16 and 18. Differs from Protoxinjiangchelys and Chuannanchelys by characters 2–4, 7, 10, 12, 14–18. Differs from Kayentachelys and Indochelys by characters 1–3, 6, 7, 10, 13–16.
Distribution. Middle Jurassic of Sichuan; ?Middle to Late Jurassic of Xinjiang.
Sichuanchelys chowi Ye & Pi, Reference Ye and Pi1997 Figures 8–10
Holotype. ZDM 3014, a complete shell with articulated carapace and plastron; part of carapace surface damaged.

Figure 8. Sichuanchelys chowi Ye & Pi, Reference Ye and Pi1997 from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. ZDM 3014 (holotype), shell in dorsal (a, b) and ventral (c, d) views. Scale bar = 50 mm.

Figure 9. Sichuanchelys chowi Ye & Pi, Reference Ye and Pi1997 from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. ZDM 3017, shell in dorsal (a, b) and ventral (c, d) views. Scale bar = 50 mm.
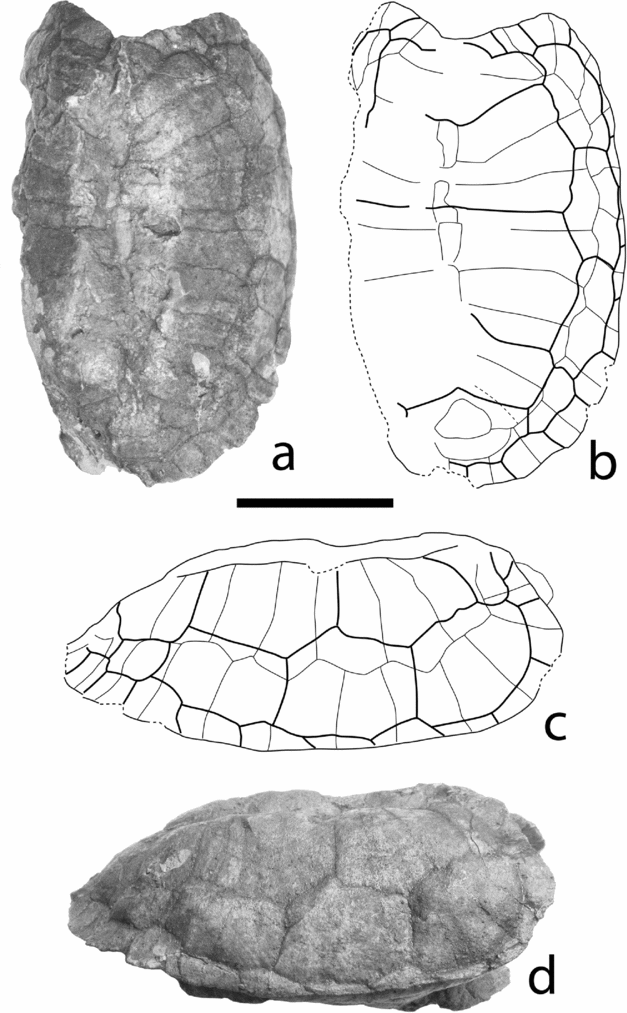
Figure 10. Sichuanchelys chowi Ye & Pi, Reference Ye and Pi1997 from the Middle Jurassic Xiashaximiao Formation, Dashanpu, Zigong. ZDM 3001, shell in dorsal (a, b) and right lateral (c, d) views. Scale bar = 50 mm.
Referred material. ZDM 3001, complete but deformed shell with articulated carapace and plastron; ZDM 3017, almost complete shell with articulated carapace and plastron, left part of carapace surface damaged; ZDM 3018, almost complete shell with articulated carapace and plastron, posterior peripheral region of carapace damaged.
Type locality and horizon. Dashanpu, Zigong, Sichuan Province; Xiashaximiao Formation, Middle Jurassic.
Diagnosis. Only species, same as for genus.
Measurements. See Table 2.
Description and comparisons. Carapace is low and broad, with a deep and large cervical notch and a slight midline depression. The carapace outline is similar to that of Kayentachelys aprix (Gaffney et al. Reference Gaffney, Hutchison, Jenkins and Mekker1987), but the posterior end of Sichuanchelys is narrower and its cervical notch is much more developed. The posterior edge of the carapace is slightly serrated, as preserved in ZDM 3001 and ZDM 3017. The shell surface is smooth without any ornamentation.
Although all specimens are almost complete shells, sutures are not always clearly preserved. The nuchal is preserved in all specimens. Although incomplete in most specimens (ZDM 3014, ZDM 3017, ZDM 3018), its shape can be easily reconstructed. The nuchal is trapezoidal, its width being twice its midline length, with the posterolateral margins strongly convergent backwards. The shape of the nuchal in ZDM 3001 is different; it is wider and shorter with an almost straight posterior margin. The second to fifth neurals are preserved in ZDM 3001, the second to sixth neural are preserved in ZDM 3014 and the first to fourth neurals are preserved in ZDM 3017. The first neural is rectangular and wide. The second through sixth neurals are elongated hexagonal with short sides in front. The remaining neurals are not preserved. The neural formula is thus 4<6>6>6>6>6. As preserved in ZDM 3001 and ZDM 3017, two suprapygals are present. The first suprapygal is roughly triangular and does not reach the peripheral, in contrast to Bashuchelys zigongensis. The second suprapygal is clearly wider than the first and contacts the eleventh peripheral and pygal. The pygal, as preserved in ZDM 3001 and ZDM 3017, is wider than long. In comparison, Kayentachelys aprix has broad neurals of irregular length and shape, and the first two suprapygals are smaller than the third suprapygal. The first costal plate is well preserved on the right side of ZDM 3001 and on the left side of ZDM 3017. It is large and clearly longer than the second costal, in contrast to Bashuchelys spp., Chuannanchelys dashanpuensis, Kayentachelys aprix and other primitive turtles in which the first costal is as long as the second costal. Eight costals are present in Sichuanchelys chowi in contrast to nine costals in Kayentachelys. Eleven peripherals are present, all very broad. The peripherals in Sichuanchelys chowi are broader than that of Kayentachelys aprix; the first to sixth peripherals extend medially to half the width of the pleural scutes; the seventh and eighth peripherals extend medially to two thirds of the pleural width. The ninth and tenth peripherals are almost as wide as the corresponding marginal plus the fourth pleural scute, with their medial suture matching or close to the vertebral–pleural sulcus (ZDM 3001) or extending onto the fourth vertebral scute (ZDM 3017). The tenth and eleventh peripherals, as well as the pygal, extend onto the fifth vertebral scute.
The cervical scute is complete in ZDM 3017. It is small and slightly wider than long. The vertebrals are all extremely wide, the second to fourth vertebrals being almost three times as wide as long. The first vertebral scute has a particular shape in that it is clearly longer in the midline than laterally, with its posterior border strongly convex backwards, different from the nearly straight posterior sulcus of the first vertebral in Kayentachelys aprix. All pleural scutes are narrow, especially the fourth; their width decreases from the front to the back. Twelve marginal scutes are present. They are all narrow, particularly the third through eighth, with their width being one third of the corresponding peripheral, and included in the peripherals except the first and the twelfth.
The plastron is sutured to the carapace, with the anterior margin extending to the anterior margin of the carapace. The plastron in ZDM 3014 and ZDM 3018 is broad, with short anterior and posterior lobes, while the plastron of ZDM 3001 and ZDM 3017 appears to be narrower. The anterior rim of the plastron is turned dorsally smoothly, with a pair of light swellings formed by the intergulars, as in Kayentachelys aprix. The anal notch is absent. The axillary and inguinal buttresses are long.
The epiplastron shape can be traced out in ZDM 3017 and ZDM 3014. It is an anteromedially-posterolaterally elongate element, with a very short midline suture to its mate before the entoplastron. Its posterior contact with the entoplastron and hyoplastron is a straight, long and oblique suture. This shape of the entoplastron is comparable to sinemydid/macrobaenid Ordosemys liaoxiensis and Kirgizemys hoburensis, but different from Kayentachelys aprix and most other turtles, which have an anteriolaterally directed epiplastron–hyoplastron suture. The entoplastron is a small triangular plate with a very narrow posterior end. The mesoplastron is well preserved in ZDM 3014, with clear sutures. The mesoplastra meet on the midline; it is short on the midline and becomes much longer laterally, with the lateral length being twice the midline length. It is mostly included in the abdominal scute except laterally on the bridge. The hyoplastron–mesoplastron suture is slightly convex posteriorly. The hypoplastron and xiphiplastron have a similar length. The hypoplastron–xiphiplastron suture is slightly convex posteriorly.
As preserved in ZDM 3014 and ZDM 3017, the intergular is paired, covering the anterior part of the entoplastron. The gular is wide and short, shorter than the intergular. The humeropectoral sulcus is placed far behind the entoplastron and clearly anterior to the axillary notch as in Bashuchelys spp. and Kayentachelys aprix, in contrast to Xinjiangchelys. The pectoroabdominal sulcus is very close to the hyoplastron–mesoplastron suture as in Bashuchelys spp. The femoroanal sulcus is convex anteriorly but not reaching the hypoplastron–xiphiplastron suture. The midline sulcus is straight. As preserved in ZDM 3014 and ZDM 3017, four inframarginal scutes are present.
Sichuanchelys sp.
1990b Chengyuchelys baenoides Young & Chow; Ye, p. 305.
1990b Chengyuchelyidae indet. Ye, p. 307, fig. 1.
1994 Chengyuchelys baenoides Young & Chow; Ye, p. 7.
1994 Chengyuchelyidae indet. Ye, p. 12.
2005 Chengyuchelyidae gen. et sp. indet. Peng et al., p. 45, fig. 29.
2008 Sichuanchelys sp. indet. Danilov & Parham, p. 312, fig. 3.
Referred material. IVPP V8805, a shell of a juvenile; ZDM 3002, posterior half of a carapace.
Locality and horizon. Dashanpu, Zigong, Sichuan Province, China; Xiashaximiao Formation, Middle Jurassic.
Remarks. IVPP V8805 has been referred to Chengyuchelys baenoides by Ye (Reference Ye1990b). Danilov & Parham (Reference Danilov and Parham2008) redescribed the specimen and referred it to Sichuanchelys sp. based on the extremely wide vertebral scutes and narrow mesoplastra.
ZDM 3002 was described as Chengyuchelyidae indet. by Ye (Reference Ye1990b) and Chengyuchelyidae gen et sp. indet. by Peng et al. (Reference Peng, Ye, Gao, Shu and Jiang2005). The second through fifth neurals are elongate and hexagonal, with short anterolateral sides. The second to fourth vertebrals are very broad, the third one being the broadest. The intervertebral sulcus between the third and fourth vertebrals lies on the fifth neural. The seventh and eighth marginals are elongate and narrower than the corresponding peripherals. This specimen is tentatively referred to Sichuanchelys based on the extremely broad vertebrals.
6. Phylogenetic relationships of Dashanpu turtles
The turtle assemblage of the Middle Jurassic Dashanpu locality contains five species belonging to four genera and three families. Three genera (Bashuchelys, Chuannanchelys and Protoxinjiangchelys) are closely related to one another, and share a combination of primitive and derived characters. Among them, the regular elongate neural series, fan-shaped bridge with long axillary and inguinal buttresses and omega-shaped femoroanal sulcus reaching or extending onto the hypoplastron are apomorphical relative to other primitive turtles. The sulcus between the third and the fourth vertebrals on the sixth neural is considered primitive since it is widely spread in primitive turtles such as Kayentachelys aprix, Indochelys spatulata, Eileanchelys waldmani and Condorchelys antiqua. The humeropectoral sulcus lying far anterior to the axillary notch and the pectoral scute notably longer than the abdominal are two characters that are related to one another. It is derived relative to Proganochelys quenstedti and Heckerochelys romani and appears in bashuchelyids and also in Kayentachelys aprix. Despite the narrow first to third vertebral scutes, Bashuchelys is more primitive relative to Chuannanchelys and Protoxinjiangchelys in the gular and intergular projections, leaf-shaped entoplastron, short epiplastra midline contact and pair of intergulars extending deeply onto the entoplastron. Although the mesoplastron in B. youngi and B. zigongensis is different from one another, it appears more reduced than in Kayentachelys aprix, Heckerochelys romani, Glyptops plicatulus and Pleurosternon bullockii (Table 3) in that the hyoplastron–mesoplastron suture is very close to the pectoroabdominal sulcus in the middle. The gular and intergular projections are primitive features that are present in the Late Triassic turtles Proganochelys quenstedti and Proterochersis robusta, which differ in that the intergular projections are paired or triple (Gaffney, Reference Gaffney1990). The leaf-shaped entoplastron in Bashuchelys spp. closely resembles that of Kayentachelys aprix and Indochelys spatulata. However, Bashuchelys is more advanced relative to Kayentachelys and Indochelys in the epiplastra meeting on the midline, preventing the entoplastron from being exposed on the plastral front margin; eight elongate and regular neurals; and narrow first to third vertebrals (Gaffney et al. Reference Gaffney, Hutchison, Jenkins and Mekker1987; Datta et al. Reference Datta, Manna, Ghosh and Das2000). As none of specimens has the dorsal surface of the plastron exposed; the epiplastron dorsal process remains unknown in Bashuchelys.
Table 3. Comparisons of turtles from the Middle Jurassic of the Sichuan Basin and other primitive turtles

Two Bashuchelys species are united by a combination of primitive and derived characters (see diagnosis of the genus). As many carapace features are unknown in B. youngi because of the poor preservation of that part, the two species of Bashuchelys are distinguished from one another mainly by the morphology of the mesoplastron. Better preserved material is needed to provide more distinctive characters. B. youngi n. sp. is, however, more derived than B. zigongensis (Ye, Reference Ye1982) in the more strongly reduced mesoplastron that narrows in the middle and is included mostly in the abdominal scute, and in the fifth marginal extending onto the second costal and the seventh marginal extending onto the fourth costal, although this character is present in two specimens among four (IVPP V6507 and ZDM 3003). Both species of Bashuchelys have the plastron sutured to the carapace. Danilov & Parham (Reference Danilov and Parham2008) reported a loose carapace–plastron attachment in IVPP V6507, a specimen referred to ‘Chengyuchelys baenoides’ (here referred to Bashuchelys youngi). Our observations reveal that all other Bashuchelys specimens examined have a sutured carapace–plastron attachment. Despite its large size, IVPP V6507 may be a juvenile, or neotenous because of the presence of the carapacial lateral fontanelles.
The trio of Bashuchelys, Chuannanchelys and Protoxinjiangchelys share several apomorphic characters with the Late Jurassic turtles from the Sichuan Basin previously referred to ‘Plesiochelys’ and Xinjiangchelys from Northern China and Central Asia: a fan-shaped bridge with long axillary and inguinal buttresses and an omega-shaped femoroanal sulcus reaching or extending onto the hypoplastron. In addition, the first thoracic rib in Bashuchelys extends to half the width of the first costal as in Xinjiangchelys. The close relationship between Bashuchelys (Chengyuchelys) and Xinjiangchelys has been noted by Danilov & Parham (Reference Danilov and Parham2008).
‘Chengyuchelys’ dashanpuensis is moved to a new genus Chuannanchelys. Danilov & Parham (Reference Danilov and Parham2008) considered the retention of this species in the genus Chengyuchelys as untenable. It was transferred to Sichuanchelys by Brinkman et al. (Reference Brinkman, Li, Ye, Li, Wu and Zhang2008) because of the reduced mesoplastron covered entirely by the abdominal scute and the wide vertebral scutes. We exclude this species from Sichuanchelys since it lacks the autapomorphic features of the latter such as the wide peripheral plates, all marginals narrow, particularly the lateral ones which are extremely narrow, and a small triangular elongate entoplastron and elongate epiplastron with extremely short midline contact. As mentioned above, Chuannanchelys is closely related to Protoxinjiangchelys and Bashuchelys. Despite the wider plastron and presence of a mesoplastron, Chuannanchelys is similar to Protoxinjiangchelys in the wide vertebral scutes, the diamond-shaped entoplastron with short anterolateral borders, long epiplastra midline contact, and gular and intergular posterior margins forming a straight sulcus, lying before the entoplastron. Except for the first character, all others are derived relative to Bashuchelys and present in ‘Plesiochelys’ from the Sichuan Basin and Xinjiangchelys.
Protoxinjiangchelys is included in the family Xinjiangchelyidae based on the absence of a mesoplastron and other plastral features. Peng & Brinkman (Reference Peng and Brinkman1993) described a partial carapace with almost complete plastron (IVPP V9537–10) from the Late Jurassic of Penfengshan in the Junggar Basin as Xinjiangchelys sp. This specimen is more primitive relative to other Xinjiangchelys spp. but resembles Protoxinjiangchelys in the narrow plastron and position of humeropectoral sulcus.
Sichuanchelys was originally included in the family Chengyuchelyidae because of the shell outline, the smooth surface, the neural shape, the presence of a pair of mesoplastra meeting on the midline, a wide bridge and the presence of 3–4 inframarginals (Ye & Pi, Reference Ye and Pi1997). This classification is followed by other authors (Peng et al. Reference Peng, Ye, Gao, Shu and Jiang2005; Brinkman, Li & Ye, Reference Brinkman, Li, Ye, Li, Wu and Zhang2008). Based on the shell outline and scalation pattern, Danilov & Parham (Reference Danilov and Parham2008) recognized the close similarity between Sichuanchelys and Kayentachelys. The following characters are shared by Sichuanchelys and Kayentachelys: (a) wide carapace with wide cervical notch; (b) very wide vertebral scutes; (c) narrow lateral marginals which are clearly narrower than the peripherals; (d) small intergular projections and (e) the sulcus between the third and fourth vertebrals on the sixth neural. With the exception of the first character, all others appear to be primitive. The wide vertebrals are present in most Triassic to Middle Jurassic turtles, including Proganochelys, Proterochersis, Hecherochelys, Indochelys, Eileanchelys and Chondorchelys. The narrow lateral marginals are like Proganochelys and Proterochersis. The intergular projections seem to be remnants from primitive turtles such as Proganochelys or Proterochersis, and are also present in Bashuchelys. As mentioned above, the sulcus between the third and fourth vertebral on the sixth neural is widespread in primitive turtles. Sichuanchelys is more advanced relative to Kayentachelys in the following aspects: (1) eight regular and elongate neurals; (2) entoplastron not exposed on the plastral front margin; (3) mesoplastron reduced; (4) huge cervical notch; (5) all marginals narrow; (6) all peripherals mesiolaterally expanded; (7) reduced entoplastron; (8) long first costal plate; and (9) long first vertebral scute with posteriorly convex posterior border. The last six characters are autapomorphic for Sichuanchelys and distinct from Kayentachelys and also Bashuchelys. However, Sichuanchelys retains a primitive epiplastron that is anteromedially-posterolaterally elongated, with an oblique and straight suture with the entoplastron and hyoplastron. This condition is reminiscent of Proganochelys and Proterochersis, and also similar to Heckerochelys, some sinemydids/macrobaenids, cheloniids and Platysternon. In Kayentachelys, Bashuchelys and most other turtles, the epiplastron is squarish, and the epiplastron–entoplastron and epiplastron–hypoplastron suture forms an angle.
We performed the phylogenetic analysis with the addition of the four taxa from the Middle Jurassic of the Sichuan Basin (Bashuchelys, Chuannanchelys dashanpuensis, Protoxinjiangchelys salis and Sichuanchelys chowi) in the data matrix of Joyce (Reference Joyce2007) according to parameters used therein (15 ordered characters and three excluded ‘rogue’ taxa), and modified by Danilov & Parham (Reference Danilov and Parham2006, Reference Danilov and Parham2008). The latter modifications include a DNA-based constraint tree for living taxa (Danilov & Parham, Reference Danilov and Parham2006: p. 577, fig. 4), the addition of two taxa, Yehguia tatsuensis (Ye, Reference Ye1963) and Chengyuchelys (based on observations of IVPP-V6507 and published data on ‘C.’ zigongensis), and one new character ‘Marginal B’. Here we remove Chengyuchelys from the analysis (see Section 4) and do not use a constraint tree as it has no influence on the position of the examined taxa. Based on our new observations, three additional characters have been added: ‘Anal B’, ‘Pectoral B’ and ‘Gular B’ (see Appendix 1). In addition, one more taxon (Xinjiangchelys tianshanensis) is included. This makes a total of 74 taxa and 140 characters. Forty-two characters in Bashuchelys (B. zigongensis + B. youngi), 40 characters in Chuannanchelys dashanpuensis, 41 characters in Protoxinjiangchelys salis and 40 characters in Sichuanchelys chowi have been coded (Appendix 2). The cladistic analysis using NONA ver. 2 and Winclada ver. 1.00.08 by Ratchet algorithm with 1000 iterations resulted in 36 trees of 478 steps (CI = 0.36, HI = 0.64, RI = 0.72, Fig. 11).
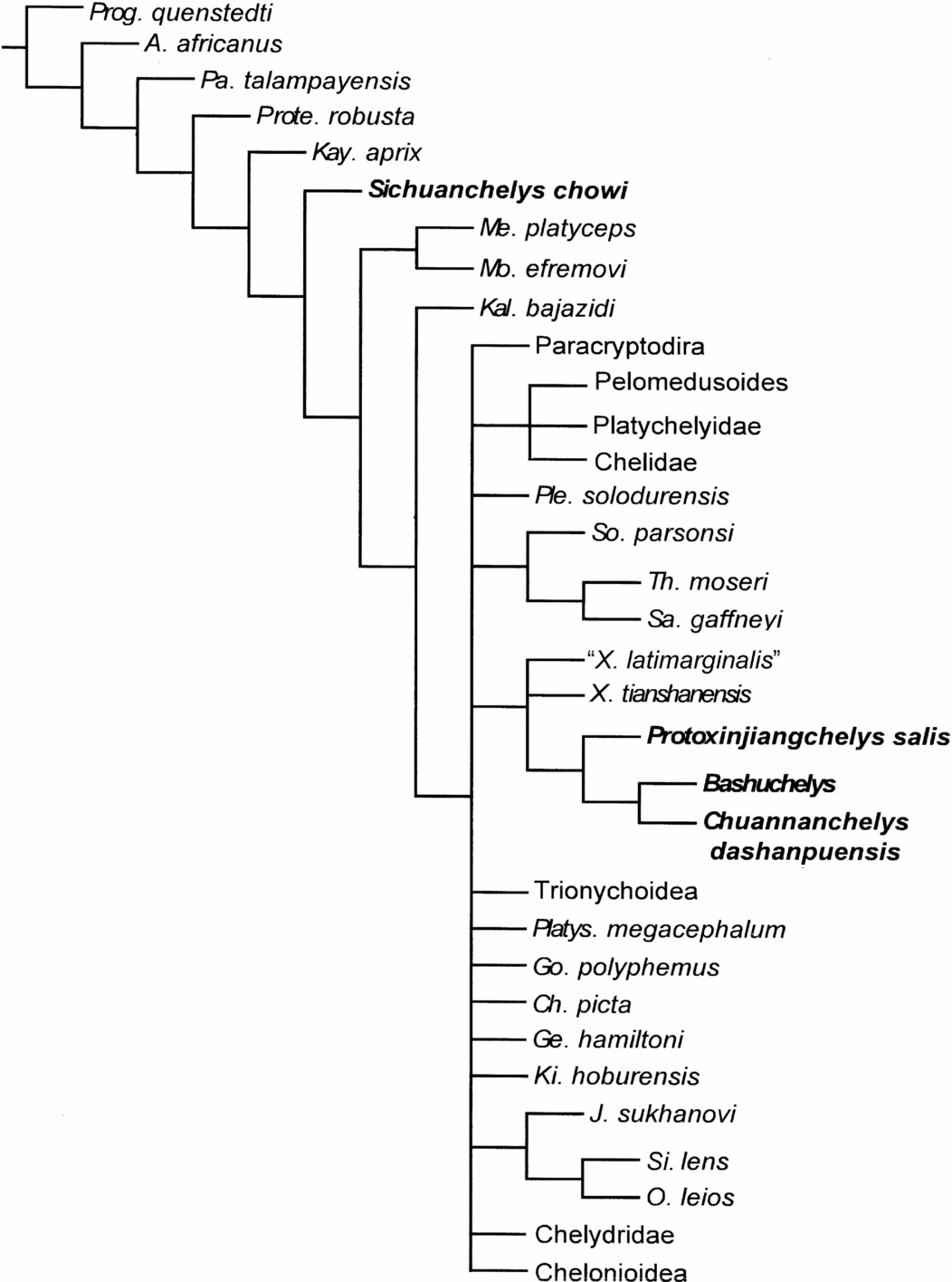
Figure 11. Strict consensus of 36 most parsimonious trees showing the phylogenetic position of Middle Jurassic turtles from Dashanpu recovered by our cladistic analysis. The following taxa have been condensed for space reasons and simplicity: Chelidae, Chelonioidea, Chelydridae, Paracryptodira, Pelomedusoides, Platychelyidae, Trionychoidea, Pleurodira. Abbreviations: A. – Australochelys; Ch. – Chrysemys, Ge. – Geoclemys, Go. – Gopherus, J. – Judithemys; Kal. – Kallokibotion; Kay. – Kayentachelys; Ki. – Kirgizemys; Me. – Meiolania; Mo. – Mongolochelys; O. – Ordosemys; Pa. – Palaeochersis; Platys. – Platysternon; Ple. – Plesiochelys; Prog. – Proganochelys; Prote. – Proterochersis; Sa. – Santanachelys; Si. – Sinemys; So. – Solnhofia; Th. – Thalassemys; X. – Xinjiangchelys.
Our cladogram shows two important points concerning the turtles studied herein. First, bashuchelyids are closely related to xinjiangchelyids, similar to the finding of Danilov & Parham (Reference Danilov and Parham2008), although relationships of bashuchelyids + xinjiangchelyids with most other turtles (Paracryptodira, Pleurodira and Eucryptodira) are unresolved, whereas in Danilov & Parham (Reference Danilov and Parham2008) Chengyuchelys + Xinjiangchelys latimarginalis were placed among stem cryptodires (correspond to Eucryptodira in this paper). Second, Sichuanchelys is placed in a more basal position, only one step more advanced than Kayentachelys. Sichuanchelys is united with turtles more advanced than Kayentachelys by a single unambiguous synapomorphy, Entoplastron A, 1 (anterior entoplastral process absent, medial contact of epiplastra present). The clade bashuchelyids + xinjiangchelyids is united with 11 unambiguous synapomorphies, all of which are considered homoplasic in our analysis: Basisphenoid B, 1 (paired pits on ventral surface of basisphenoid present); Extragular A, 0 (intergular present); Anal A, 0 (anals only cover parts of the xiphiplastra); Cervical Vertebra B, 0 (posterior cervicals without strongly developed ventral keels); Cervical Vertebra C, 0 (cervical centra VIII is not significantly shorter than VII); Cervical Articulation A, 0 (cervical central articulations not formed); Cleithrum A, 1 (cleithra (dorsal processes of epiplastra) present); Scapula C, 1 (glenoid neck on scapula present); Marginal B, 1 (marginals overlap onto costals); Anal B, 1 (omega-shaped femoroanal sulcus); and Gular B, 1 (weak or absent extension of intergulars onto the entoplastron). Among the above mentioned characters only shell ones are known in all members of the clade, whereas characters of the skull and non-shell postcrania are owing to the presence of X. tianshanensis in this clade. The clade (Protoxinjiangchelys (Bashuchelys + Chuannanchelys)) is united by two unambiguous synapomorphies, which are also considered homoplasic in our analysis: Vertebral C, 0 (position of sulcus between third and fourth vertebrals on sixth neural) and Pectoral B, 1 (pectoral longer than abdominal). Bashuchelys and Chuannanchelys are united by two characters: Neural A, 1 (neural formula 6>4<6<6<6<6) and Mesoplastron A, 0 (one pair of mesoplastra present that fully hinders any contact between the hyoplastra and hypoplastra). Thus, contrary to expected, the bashuchelyids are at a more advanced position relative to the xinjiangchelyids in the cladogram. This shortcoming regarding the bashuchelyids position is probably due to the high homoplasy in the dataset and the large amount of missing data in the specimens from the Sichuan Basin. The early age and the primitive characters such as the presence of a mesoplastron and sutured plastron–carapace attachment in bashuchelyids relative to Xinjiangchelys would support their more basal position as ((Bashuchelys + Chuannanchelys) (Protoxinjiangchelys (Xinjiangchelys + more advanced taxa))) or (Bashuchelys (Chuannanchelys (Protoxinjiangchelys (Xinjiangchelys + more advanced taxa)))). The further discovery of skull material will provide crucial information about the phylogenetic relationships of Middle Jurassic turtles from the Sichuan Basin.
7. Conclusion
By the absence of a mesoplastron and other shell characters, both the holotype and paratype of Chengyuchelys baenoides Young & Chow, Reference Young and Chow1953 belong to the family Xinjiangchelyidae and probably come from the Upper Jurassic Shangshaximiao Formation. The family Chengyuchelyidae is thus abandoned. The Middle Jurassic Dashanpu turtle assemblage represents the most diverse turtle fauna of that period and contains five species, belonging to four genera and three families. This assemblage is composed of two entities: Bashuchelys spp., Chuannanchelys and Protoxinjiangchelys on the one hand and Sichuanchelys on the other hand, with the former as the dominant group. Our study supports the close relationship between bashuchelyids and xinjiangchelyids suggested by Danilov & Parham (Reference Danilov and Parham2008). Protoxinjiangchelys, the most primitive representative of the Xinjiangchelyidae, represents the intermediate form between bashuchelyids and the more advanced xinjiangchelyids. Sichuanchelys has no shared apomorphic characters with bashuchelyids but is close to Kayentachelys. The whole assemblage appears to be endemic to the Sichuan Basin at genus level until the presence of Sichuanchelys in the Jurassic Shishugou Formation of the Junggar Basin, Xinjiang is confirmed (Brinkman & Matzke, Reference Brinkman, Matzke and Braman2009). Although closely related, this Middle Jurassic turtle fauna is distinct from the Late Jurassic turtle assemblage of the Sichuan Basin in its relict nature and absence of representatives of the Polycryptodira.
Together with the abundant Late Jurassic turtle remains, the Sichuan Basin has produced one of the most important Jurassic turtle assemblages in a key period of turtle evolution. The inland lake ecosystem of the Sichuan Basin under the subtropical–tropical climate during the Jurassic period probably favoured the development of the basal eucryptodiran turtles.
Acknowledgements
We thank Zheng Fang (IVPP, Beijing), Zhou Songlin and Tong Jiangbo (CMNH, Chongqing) and Shu Chunkang (ZDM, Zigong) for access to the specimens in their care, James Parham for help with phylogenetic analysis and discussion and two anonymous reviewers for their helpful comments.
Appendix 1. Details of new characters added to the matrix of Joyce (Reference Joyce2007)
Anal B. Character definition: femoroanal sulcus omega-shaped: 0 = absent; 1 = present. Codings: Hypothetical ancestor, 0; Proganochelys quenstedti, 0; Proterochersis robusta, 0; Palaeochersis talampayensis, 0; Australochelys africanus, ?; Kayentachelys aprix, 0; Sichuanchelys chowi, 0; Meiolania platyceps, ?; Mongolochelys efremovi, 0; Kallokibotion bajazidi, 0; Platychelys oberndorferi, 0; Caribemys oxfordiensis, ?; Notoemys laticentralis, 0; Elseya dentata, 0; Chelodina oblonga, 0; Phrynops geoffroanus, 0; Erymnochelys madagascariensis, 0; Pelomedusa subrufa, 0; Podocnemis expansa, 0; Dorsetochelys delairi, ?; Pleurosternon bullockii, 0; Glyptops plicatulus, 0; Dinochelys whitei, 0; Neurankylus eximius, 0; Trinitichelys hiatti, ?; Plesiobaena antiqua, 1; Boremys pulchra, 0; Baena arenosa, 1; Chisternon undatum, 1; Portlandemys mcdowelli, ?; Plesiochelys solodurensis, 0; Solnhofia parsonsi, 0; Thalassemys moseri, 0; Santanachelys gaffneyi, ?; Bashuchelys, 1; Chuannanchelys dashanpuensis, 1; Protoxinjiangchelys salis, 1; Xinjiangchelys latimarginalis, 1; ‘X.’ tianshanensis, 1; Kirgizemys hoburensis, 0; Judithemys sukhanovi, 0; Dracochelys bicuspis, ?; Sinemys lens, ?; Ordosemys leios, 0; Toxochelys latiremis, ?; Caretta caretta, 0; Chelonia mydas, 0; Mesodermochelys undulatus, –; Dermochelys coriacea, –; Protochelydra zangerli, 0; Macroclemys temminckii, 0; Chelydra serpentina, 0; Platysternon megacephalum, 0; Mongolemys elegans, 0; Gopherus polyphemus, 0; Chrysemys picta, 0; Geoclemys hamiltonii, 0; Emarginachelys cretacea, 0; Baptemys wyomingensis, 0; Dermatemys mawii, 0; Hoplochelys crassa, 0; Staurotypus triporcatus, 0; Sternotherus odoratus, 0; Kinosternon flavescens, 0; Zangerlia neimongolensis, 0; Basilemys variolosa, 0; Adocus beatus, 0; Yehguia tatsuensis, 0; Peltochelys duchastelli, 0; Sandownia harrisi, ?; Apalone spinifera, –; Lissemys punctata, –; Anosteira ornata, -; Carettochelys insculpta, –.
Pectoral B. Character definition: Pectoral significantly longer than abdominal: 0 = absent; 1 = present. Codings: Hypothetical ancestor, 0; Proganochelys quenstedti, 0; Proterochersis robusta, 1; Palaeochersis talampayensis, ?; Australochelys africanus, ?; Kayentachelys aprix, 0; Sichuanchelys chowi, 0; Meiolania platyceps, ?; Mongolochelys efremovi, 0; Kallokibotion bajazidi, 0; Platychelys oberndorferi, 1; Caribemys oxfordiensis, 1; Notoemys laticentralis, 0; Elseya dentata, 0; Chelodina oblonga, 0; Phrynops geoffroanus, 0; Erymnochelys madagascariensis, 0; Pelomedusa subrufa, 0; Podocnemis expansa, 0; Dorsetochelys delairi, ?; Pleurosternon bullockii, 0; Glyptops plicatulus, 0; Dinochelys whitei, 0; Neurankylus eximius, 0; Trinitichelys hiatti, 0; Plesiobaena antiqua, 0; Boremys pulchra, 1; Baena arenosa, 0; Chisternon undatum, 0; Portlandemys mcdowelli, ?; Plesiochelys solodurensis, 0; Solnhofia parsonsi, ?; Thalassemys moseri, 0; Santanachelys gaffneyi, ?; Bashuchelys, 1; Chuannanchelys dashanpuensis, 1; Protoxinjiangchelys salis, 1; Xinjiangchelys latimarginalis, 0; ‘X.’ tianshanensis, 0; Kirgizemys hoburensis, 0; Judithemys sukhanovi, 1; Dracochelys bicuspis, ?; Sinemys lens, ?; Ordosemys leios, ?; Toxochelys latiremis, ?; Caretta caretta, 0; Chelonia mydas, 0; Mesodermochelys undulatus, –; Dermochelys coriacea, –; Protochelydra zangerli, –; Macroclemys temminckii, –; Chelydra serpentina, –; Platysternon megacephalum, 0; Mongolemys elegans, 0; Gopherus polyphemus, 0; Chrysemys picta, 0; Geoclemys hamiltonii, 0; Emarginachelys cretacea, –; Baptemys wyomingensis, –; Dermatemys mawii, –; Hoplochelys crassa, –; Staurotypus triporcatus, –; Sternotherus odoratus, –; Kinosternon flavescens, –; Zangerlia neimongolensis, 0; Basilemys variolosa, 0; Adocus beatus, 0; Yehguia tatsuensis, 0; Peltochelys duchastelli, 0; Sandownia harrisi, ?; Apalone spinifera, –; Lissemys punctata, –; Anosteira ornata, –; Carettochelys insculpta, –.
Gular B. Character definition: Extension of gulars (intergulars in this paper) on the entoplastron: 0 = strong; 1 = weak or absent. Codings: Hypothetical ancestor, 0; Proganochelys quenstedti, 0; Proterochersis robusta, 0; Palaeochersis talampayensis, ?; Australochelys africanus, ?; Kayentachelys aprix, 0; Sichuanchelys chowi, 0; Meiolania platyceps, 0; Mongolochelys efremovi, 1; Kallokibotion bajazidi, 0; Platychelys oberndorferi, 0; Caribemys oxfordiensis, 0; Notoemys laticentralis, 0; Elseya dentata, 0; Chelodina oblonga, 0; Phrynops geoffroanus, 0; Erymnochelys madagascariensis, 1; Pelomedusa subrufa, 0; Podocnemis expansa, 0; Dorsetochelys delairi, ?; Pleurosternon bullockii, 0; Glyptops plicatulus, 1; Dinochelys whitei, 1; Neurankylus eximius, 0; Trinitichelys hiatti, 0; Plesiobaena antiqua, 1; Boremys pulchra, 1; Baena arenosa, 0; Chisternon undatum, 1; Portlandemys mcdowelli, ?; Plesiochelys solodurensis, 0; Solnhofia parsonsi, ?; Thalassemys moseri, ?; Santanachelys gaffneyi, ?; Bashuchelys, 0; Chuannanchelys dashanpuensis, 1; Protoxinjiangchelys salis, 1; Xinjiangchelys latimarginalis, 1; ‘X.’ tianshanensis, 1; Kirgizemys hoburensis, 1; Judithemys sukhanovi, ?; Dracochelys bicuspis, ?; Sinemys lens, ?; Ordosemys leios, ?; Toxochelys latiremis, ?; Caretta caretta, ?; Chelonia mydas, ?; Mesodermochelys undulatus, –; Dermochelys coriacea, –; Protochelydra zangerli, 1; Macroclemys temminckii, 1; Chelydra serpentina, 1; Platysternon megacephalum, 1; Mongolemys elegans, 1; Gopherus polyphemus, 0; Chrysemys picta, 0; Geoclemys hamiltonii, 0; Emarginachelys cretacea, 0; Baptemys wyomingensis, 0; Dermatemys mawii, 0; Hoplochelys crassa, 0; Staurotypus triporcatus, 0; Sternotherus odoratus, –; Kinosternon flavescens, –; Zangerlia neimongolensis, 1; Basilemys variolosa, 1; Adocus beatus, 1; Yehguia tatsuensis, 1; Peltochelys duchastelli, 0; Sandownia harrisi, ?; Apalone spinifera, –; Lissemys punctata, –; Anosteira ornata, –; Carettochelys insculpta, –.
Appendix 2. Characters coded for taxa added to the matrix of Joyce (Reference Joyce2007) with modifications of Danilov & Parham (Reference Danilov and Parham2008)
Sichuanchelys chowi: Carapace A, 0; Carapace B, 0; Neural A, 0; Peripheral A, 1; Peripheral B, 0; Costal A, 0; Costal B, 0; Costal C, 0; Cervical A, 0; Supramarginal A, 2; Marginal B, 0; Vertebral A, 1; Vertebral B, 0; Vertebral C, 0; Plastron A, 0; Plastron B, 0; Plastron C, 0; Entoplastron A, 1; Entoplastron C, 1; Entoplastron D, 0; Entoplastron E, 0; Epiplastron A, 0; Mesoplastron A, 0; Xiphiplastron A, 0; Xiphiplastron B, 0; Plastral Scutes A, 0; Plastral Scutes B, 0; Gular A, 0; Extragular A, 0; Extragular B, 0; Extragular C, 1; Intergular A, 0; Humeral A, 0; Pectoral A, 0; Abdominal A, 0; Anal A, 0; Inframarginal A, 0.
Bashuchelys: Carapace A, 0; Carapace B, 0; Nuchal B, 0; Neural A, 1; Peripheral A, 1; Peripheral B, 0; Costal A, 0; Costal C, 0; Cervical A, 2; Supramarginal A, 2; Marginal B, 1; Vertebral A, 1; Vertebral B, 1; Vertebral C, 0; Plastron A, 0; Plastron B, 0; Plastron C, 0; Entoplastron A, 1; Entoplastron C, 1; Entoplastron D, 0; Entoplastron E, 0; Epiplastron A, 0; Hyoplastron A, 0; Mesoplastron A, 0; Hypoplastron A, 0; Xiphiplastron A, 0; Xiphiplastron B, 0; Plastral Scutes A, 0; Plastral Scutes B, 0; Gular A, 0; Extragular A, 0; Extragular B, 0; Extragular C, 0; Intergular A, 0; Humeral A, 0; Pectoral A, 0; Abdominal A, 0; Anal A, 1; Inframarginal A, 0.
Chuannanchelys dashanpuensis: Carapace A, 0; Carapace B, 0; Neural A, 1; Peripheral B, 0; Costal A, 0; Costal B, 0; Costal C, 0; Cervical A, 0; Supramarginal A, 2; Vertebral A, 1; Vertebral B, 1; Vertebral C, 0; Plastron A, 0; Plastron B, 0; Plastron C, 0; Entoplastron A, 1; Entoplastron C, 1; Entoplastron D, 0; Entoplastron E, 0; Epiplastron A, 0; Hyoplastron A, 0; Mesoplastron A, 0; Hypoplastron A, 0; Xiphiplastron A, 0; Xiphiplastron B, 0; Plastral Scutes A, 0; Plastral Scutes B, 0; Gular A, 0; Extragular A, 0; Extragular B, 0; Extragular C, 1; Intergular A, 0; Humeral A, 0; Pectoral A, 0; Abdominal A, 0; Anal A, 1; Inframarginal A, 0.
Protoxinjiangchelys salis: Carapace A, 0; Carapace B, 0; Neural A, 0; Peripheral B, 0; Costal A, 0; Costal B, 0; Costal C, 0; Supramarginal A, 2; Marginal B, 1; Vertebral A, 1; Vertebral B, 1; Vertebral C, 0; Plastron A, 0; Plastron B, 0; Plastron C, 0; Entoplastron A, 1; Entoplastron C, 1; Entoplastron D, 0; Entoplastron E, 0; Epiplastron A, 0; Hyoplastron A, 0; Mesoplastron A, 2; Hypoplastron A, 0; Xiphiplastron A, 0; Xiphiplastron B, 0; Plastral Scutes A, 0; Plastral Scutes B, 0; Gular A, 0; Extragular A, 0; Extragular B, 0; Extragular C, 1; Intergular A, 0; Humeral A, 0; Pectoral A, 0; Abdominal A, 0; Anal A, 1; Inframarginal A, 0.
Xinjiangchelys tianshanensis: Quadrate A, 1; Quadrate B, 1; Quadrate C, 2; Quadrate F, 1; Pterygoid A, 1; Pterygoid B, 1; Pterygoid C, 1; Pterygoid D, 1; Pterygoid G, 0; Supraoccipital A, 1; Supraoccipital B, 0; Opisthotic A, 1; Basisphenoid A, 0; Basisphenoid B, 1; Stapedial Artery A, 1; Stapedial Artery B, 0; Jugular Foramina A, 1; Canalis Caroticum A, 3; Dentary A, 0; Carapace A, 0; Carapace B, 0; Nuchal A, 1; Nuchal B, 0; Neural A, 0; Peripheral A, 1; Peripheral B, 0; Costal A, 0; Costal B, 0; Costal C, 0; Cervical A, 0; Supramarginal A, 2; Marginal B, 1; Vertebral A, 1; Vertebral B, 1; Vertebral A, 1; Vertebral B, 1; Vertebral C, 1; Plastron A, 1; Plastron B, 0; Plastron C, 0; ; Entoplastron A, 1; Entoplastron B, 1; Entoplastron C, 1; Entoplastron D, 0; Entoplastron E, 0; Epiplastron A, 0; Hyoplastron A, 0; Mesoplastron A, 2; Hypoplastron A, 0; Xiphiplastron A, 0; Xiphiplastron B, 0; Plastral Scutes A, 0; Plastral Scutes B, 0; Gular A, 0; Extragular A, 0; Extragular B, 0; Extragular C, 1; Intergular A, 0; Humeral A, 0; Pectoral A, 0; Abdominal A, 0; Anal A, 1; Inframarginal A, 0; Cervical Rib A, 1; Cervical Vertebra A, 1; Cervical Vertebra B, 0; Cervical Vertebra C, 0; Cervical Articulation A, 0; Dorsal Rib A, 2; Dorsal Rib B, 0; Dorsal Rib C, 1; Caudal A, 1; Cleithrum A, 1; Scapula A, 1; Scapula B, 1; Scapula C, 1; Coracoid A, 1; Pelvis A, 0; Ilium A, 1; Ilium D, 0; Ischium A, 1; Hypoischium A, 1.














