


1. Introduction
Graptolites are extinct colonial hemichordates, generally considered to be closely related to the present-day pterobranchs. They range in age from the middle of the Cambrian to the Carboniferous. The graptolites include the exclusively planktonic graptoloids, the largely benthic dendroids and also the benthic crustoids, tuboids, cameroids and dithecoids (Rickards & Durman, Reference Rickards, Durman, Bassett and Deisler2006). The graptoloids are the focus of this account. They provide the primary means of correlation of Ordovician and Silurian strata in the UK, and are fundamental to resolving the stratigraphical and structural architecture of these rocks (e.g. Zalasiewicz, Reference Zalasiewicz2001), which were laid down in sedimentary basins on the margins of the Palaeozoic Iapetus Ocean. Major outcrops are in the Southern Uplands of Scotland, the Lake District and the Howgill Fells of northern England, and Wales and the Welsh Borderland. Outside Great Britain, graptolites are important also in early Devonian successions.
The British graptolite biozonal and subzonal schemes are shown in Figures 1 and 2. In the early Ordovician, planktonic dendroids, and then the dichograptid graptoloids, were the most important groups. These were succeeded by the diplograptids, which were dominant throughout the rest of the Ordovician. Dicranograptids and nemagraptids were also important elements at various times in the mid- to late Ordovician. (Such terms are used here in a general sense; higher-level graptolite taxonomy is discussed in more detail in Mitchell, Reference Mitchell1987 and Mitchell et al. Reference Mitchell, Goldman, Klosterman, Maletz, Sheets and Melchin2007).
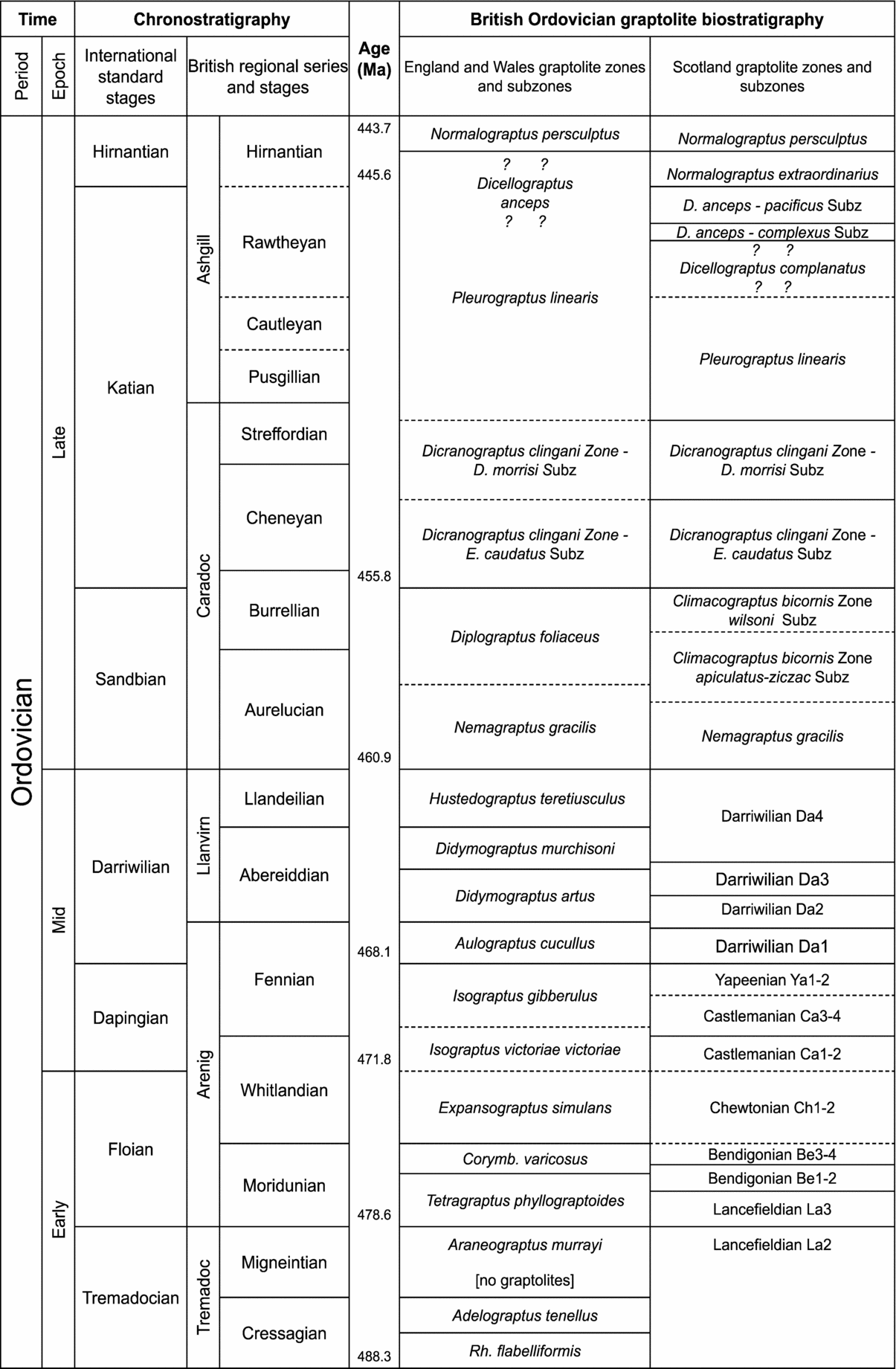
Figure 1. British Ordovician graptolite biozones and subzones. Zonal scheme based on that proposed by Fortey et al. (Reference Fortey, Harper, Ingham, Owen and Rushton1995), and adopted in the Ordovician Correlation Report of Fortey et al. (Reference Fortey, Harper, Ingham, Owen, Parkes, Rushton and Woodcock2000), with subsequent modification to the Arenig to early Llanvirn by Cooper et al. (Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004), refinement of the Caradoc by Bettley, Fortey & Siveter (Reference Bettley, Fortey and Siveter2001), while the Caradoc/Ashgill section of England and Wales shows the modified correlations suggested by Rickards (Reference Rickards2002). Chronostratigraphy and radiometric dates after Ogg, Ogg & Gradstein (Reference Ogg, Ogg and Gradstein2008).

Figure 2. British Silurian graptolite biozones and subzones. Zonal scheme follows Rickards (Reference Rickards1976, depicted) with modifications from Loydell (Reference Loydell1992–1993a), Zalasiewicz (Reference Zalasiewicz1994), Loydell & Cave (Reference Loydell and Cave1993, Reference Loydell and Cave1996), Zalasiewicz & Williams (Reference Zalasiewicz and Williams1999) and those proposed herein. Chronostratigraphy and radiometric dates after Ogg, Ogg & Gradstein (Reference Ogg, Ogg and Gradstein2008).
Following near-extinction during the latest Ordovician glaciation, a few species of diplograptids survived into the earliest Silurian. These gave rise to the monograptids, which diversified to dominate the Silurian and Devonian seas worldwide (although in Great Britain, graptolites disappeared from the shallowing marine basins late in the Ludlow). Normal diplograptids persisted for a short while into the Silurian, while retiolitid (‘meshwork’) graptolites were locally common and survived into the Ludlow. The morphologically diverse and rapidly evolving monograptids provide a fine resolution for the Silurian, with graptolite zones lasting, on average, well under a million years (Rickards, Reference Rickards1976, Reference Rickards, Holland and Bassett1989; Zalasiewicz, Reference Zalasiewicz1990; Hughes, Reference Hughes1995; Melchin, Cooper & Sadler, Reference Melchin, Cooper, Sadler, Gradstein, Ogg and Smith2004); by contrast, the duration of graptolite zones in the British Ordovician averages c. 2 Ma (Rushton, Reference Rushton1990, cf. Cooper & Sadler, Reference Cooper, Sadler, Gradstein, Ogg and Smith2004).
2. Palaeoecology, provincialism and distribution
Graptolites are commonly held to be ‘ideal’ zone fossils, because they were widely distributed in marine waters and so not bound by facies. The situation, though, is not as simple as this. The graptoloids were probably holoplanktonic, although there is still much debate about whether they floated more or less passively (e.g. Bulman, Reference Bulman1964; see also discussion in Rigby & Rickards, Reference Rigby and Rickards1990 and Palmer & Rickards, Reference Palmer and Rickards1991) or actively propelled themselves through the water (e.g. Kirk, Reference Kirk1978; Bates & Kirk, Reference Bates and Kirk1984, Reference Bates and Kirk1985; Rickards et al. Reference Rickards, Rigby, Rickards and Swales1998; Melchin & DeMont, Reference Melchin and DeMont1995).
Planktonic graptolites have long been interpreted as largely ‘open ocean’ dwellers, common in offshore pelagic and hemipelagic sequences (‘graptolite facies’), and rare or absent in shallow water deposits (‘shelly facies’). Subsequent elaborations of this general observation included suggestions that graptolites were subject to depth control, with near-surface and deep-living taxa (e.g. Berry & Boucot, Reference Berry and Boucot1972; Bates & Kirk, Reference Bates and Kirk1984; Erdtmann, Reference Erdtmann and Bassett1976) or that they were controlled by ‘water mass specificity’ with particular assemblages of taxa adapted to particular conditions of temperature and chemistry (Finney, Reference Finney1986), while competition from other (soft-bodied) macrozoo-plankton may also have restricted their occurrence (Zalasiewicz, Reference Zalasiewicz2001). Finney & Berry (Reference Finney and Berry1997) disputed the notion of graptolites as truly ocean-going, noting their absence from many deep-water, anoxic deposits, and suggested that they were largely confined to the region of the outer shelf and continental slope.
Provincialism in the graptolites was particularly marked in the Ordovician, with well-defined ‘Atlantic’ and ‘Pacific’ provinces between which it is often difficult to correlate. This may have been due in part to pronounced climatic gradients (Skevington, Reference Skevington1974), the Atlantic province representing temperate, and the Pacific representing equatorial, waters. More local provincialism is also common, however, with apparently coeval faunas of markedly different composition being reported from different regions of the USA interior (Finney, Reference Finney1986), and from the western and eastern parts of the Welsh Basin (e.g. compare the faunal successions in Hughes, Reference Hughes1989 and Zalasiewicz, Reference Zalasiewicz1992a). Provincialism is less pronounced in the Silurian (Melchin, Reference Melchin1989), and the British biozonal system can be applied for much of the Silurian throughout much of the world, with relatively minor modifications (see Koren’ et al. Reference Koren’, Lenz, Loydell, Melchin, Štorch and Teller1996 and Melchin, Cooper & Sadler, Reference Melchin, Cooper, Sadler, Gradstein, Ogg and Smith2004).
3. Taxonomy
One of the fundamental constraints on graptolite biostratigraphy is the ability to discriminate consistently between taxa. Many graptolites have a complex morphology, with many identifiable features that can be measured and tabulated. Nevertheless, the identification of graptolites is not by any means universally straightforward, partly because of the difficulty of assessing levels of intraspecific variation and partly because of preservational factors (see below).
Furthermore, in some early Ordovician dichograptids, while new thecae were being added to the growing tips of the colony, the early-formed thecae continued to grow; this gave rise to mature rhabdosomes that have a markedly different appearance from juvenile ones (Williams & Stevens, Reference Williams and Stevens1988, p. 49). Later Ordovician diplograptid taxa are often difficult to classify because the phylogenetically significant patterns of the very earliest growth stages (e.g. Mitchell, Reference Mitchell1987) can be recognized only in very well-preserved material.
The Silurian monograptids are arguably more tractable. The development of a single stipe led to a great diversity in rhabdosome shape, with different types of straight, curved or plane- to helically spiralled forms (Figs 11–19). This was accompanied by the evolution of a wide range of thecal types, many of which can be recognized even in indifferently preserved material. The Silurian monograptids were thus very ‘expressive’ morphologically. Evolution is easy to see in them, although it is uncertain whether it was actually more rapid than in the less easily interpreted Ordovician taxa.
Taxonomic uncertainties continue to be the most severe constraint upon the use of graptolites in biostratigraphy. Many species are poorly understood or inadequately described, and those, particularly in the older literature, can ‘mutate’ in subsequent descriptions to become, in the words of N. F. Hughes, imprecise ‘balloon’ taxa. There are many instances of the type material of a ‘classical’ species being too poorly preserved, or inadequately described, to serve as a reference specimen by modern standards. It has been common, also, for several distinct taxa to be described, at various times, as the same species. Conversely, Loydell (Reference Loydell, Hailwood and Kidd1993b, pp. 330–1) has shown how Stomatograptus longus Obut, 1949 has, over time, acquired five additional species names (junior synonyms). There remains much taxonomic ‘housekeeping’ to do, much of it straightforward but time-consuming. Better definition of taxa, for instance by the wholesale refiguring of type material (Zalasiewicz et al. Reference Zalasiewicz, Rushton, Hutt and Howe2000; Zalasiewicz & Rushton, Reference Zalasiewicz and Rushton2008) should lead to greater refinement in graptolite biostratigraphy.
4. Preservation
The widespread occurrence of graptolites in British early Palaeozoic successions is due to a phenomenon which is absent from recent oceans: that of prolonged, widespread periods of sea-floor anoxia (Page et al. Reference Page, Zalasiewicz, Williams, Popov, Williams, Haywood, Gregory and Schmidt2007 and references therein). Thus, graptolites are typically found in finely laminated hemipelagic deposits (‘graptolite shales’) laid down in anoxic conditions that excluded benthos. In Britain, graptolites are generally rare or absent in rocks that were laid down under oxygenated sea-floor conditions and that were colonized, and bioturbated, by a benthic fauna (e.g. Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
British Ordovician and Silurian deep-water sequences characteristically show an alternation of oxic facies (‘barren beds’) and anoxic facies (‘graptolite shales’) (Rickards, Reference Rickards1964; Cave, Reference Cave, Harris, Holland and Leake1979; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). These are most clearly seen in condensed, pelagic deposits such as the Moffat Shale Group of Scotland and the Skelgill Beds of the Lake District, where individual graptolite biozones tend to be only a few metres thick. In coarser clastic successions, such as the kilometres-thick Silurian turbidite deposits of the Welsh Basin, the ‘graptolite shales’ take the form of millimetre- to centimetre-thick units between individual turbidites.
The relative proportion of ‘graptolite shales’ and ‘barren beds’ exerts a strong control on the preservation of graptolites, and hence on the resolution of graptolite biostratigraphy. In the Caradoc of Scotland and Wales, for instance, the clingani Biozone is locally preserved within continuously anoxic facies, and this has enabled detailed range charts (e.g. Williams, Reference Williams1982a; Zalasiewicz, Rushton & Owen, Reference Zalasiewicz, Loydell and Štorch1995) to be constructed, allowing the incomings and extinctions of species to be established, thus offering the prospect of very detailed correlation. The complanatus Biozone of the lower Ashgill, by contrast, is preserved in the Southern Uplands of Scotland only as two 5 cm thick beds within a sequence of ‘barren beds’ several metres thick. In Wales, the complanatus Biozone has not been recognized, probably due to an absence of anoxic facies of that age.
In highly expanded successions, such as the Skiddaw Group of the Lake District, graptolites can be rare simply through dilution by clastic sediment. Most of the Skiddaw graptolites have been found not at exposure, but by searching through extensive screes (Cooper et al. Reference Cooper, Rushton, Molyneux, Moore and Webb1995, Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004). The Manx Slates of the Isle of Man are the equivalents of the Skiddaw Group but, as there are few screes on the Isle of Man, graptolites have only ever been found on two occasions, a century apart (Bolton, Reference Bolton1899; Rushton, Reference Rushton1993). Further, one ‘Manx Slate’ lithofacies was recently shown to contain late Wenlock graptolites (Howe, Reference Howe, Woodcock, Quirk, Fitches and Barnes1999), necessitating a radical revision of geological interpretations of that island.
Graptolites are preserved either diagenetically flattened or in partial to full relief. Flattened specimens, common in condensed black shale successions, may originally have been encased in gelatinous ‘marine snow’ or microbial mats on the sea floor (Jones, Zalasiewicz & Rickards, Reference Jones, Zalasiewicz and Rickards2002). The pyritized relief material which is common, for instance, in the Llandovery turbiditic sequences of Wales, generally shows more morphological information than can be gleaned from flattened material. However, relief material may occasionally be more difficult to identify than the ‘simple’ flattened silhouettes on which many specific descriptions were originally based; for example, the characteristic apertural spines of the zone fossil Stimulograptus sedgwickii are normally not visible on relief specimens, being either broken off or embedded in the rock matrix. Graptolites that can be dissolved from limestones or cherts to provide exquisitely preserved isolated specimens are rare in British sequences (a notable exception is the Balclatchie fauna from Scotland: Bulman, Reference Bulman1944–1947); however, little use has been made of the graptolite fragments that are encountered while preparing samples for conodonts or chitinozoans.
5. Graptolite biozones
The distinctiveness and usefulness of graptolite assemblages were recognized midway through the nineteenth century, notably by Joachim Barrande (Reference Barrande1850) in Bohemia, James Hall (Reference Hall1865) in Canada, Gustav Linnarsson (Reference Linnarsson1871) in Sweden, and Charles Lapworth (Reference Lapworth1878) in the Southern Uplands of Scotland. Barrande and Lapworth both worked in areas that are now realized to be characterized by many structural dislocations and repetitions. Barrande believed that in any area, these repeated assemblages were environmentally controlled, successively ‘colonizing’ it whenever conditions were right. Lapworth interpreted his assemblages to be temporally restricted and used them to demonstrate the multiple structural repetitions of the Southern Uplands, thus simultaneously solving a major controversy of British regional geology and providing one of the most precise correlative tools in geology.
Lapworth's graptolite biozones, as modified by Elles & Wood (Reference Elles and Wood1901–1918) are essentially those in use today, though further refined and subdivided (e.g. Rickards, Reference Rickards1976; Loydell, Reference Loydell1992–1993a; Loydell & Cave, Reference Loydell and Cave1996). They are broadly assemblage biozones, named after a species, ideally with a restricted vertical range and a wide horizontal distribution (see Rickards, Reference Rickards1995). A name-giving species may be restricted to its biozone, with its incoming being used to define the zonal base (e.g. Monograptus riccartonensis, ‘Monograptus’ crispus). It may, equally, range outside ‘its’ biozone, either above its upper boundary (e.g. Spirograptus turriculatus, Monograptus firmus) or exceptionally below it (e.g. Bohemograptus). What are important in the recognition of a biozone are the total assemblage and the incoming species. Strictly speaking, they are thus the Oppel biozones of the North American Stratigraphic Code, but in practice, assemblage biozones are essentially the same. Where the base of a biozone is defined by the incoming of more than one species, it is realized that these species may not appear exactly synchronously, but in practical biostratigraphical collecting they commonly seem to do so, particularly in condensed sequences.
Graptolite biozones, like biozones generally and unlike chronostratigraphical units, are not fixed within sections by ‘golden spikes’. They may, though, have type sections, where they were originally described, enabling clear, original definitions and forming the starting point for subsequent redefinition either there or elsewhere in the world.
An alternative to the use of assemblage biozones is the use of evolutionary lineages. These are not so widely applied in the case of graptolites, largely because the use of assemblage biozones is so successful. Their use is also constrained by the relatively small amount of detailed work that has been done on graptolite evolutionary lineages, particularly with regard to determining whether these lineages show gradualistic or punctuational change. In gradualistic lineages, arbitrary ‘snips’ of a continuum must be selected, while punctuational lineages are ‘naturally’ broken up into discrete taxon ranges. It might be said that the latter show evolution to be working in a ‘digital’ fashion compared to the ‘analogue’ mechanism of the former. Both punctuational and gradualistic modes of evolution may be inferred in the graptolite record. Punctuation may be invoked where distinctive species, such as Aulograptus cucullus, seem to ‘appear’ globally without any trace of a direct ancestor, though this observation must be viewed in the knowledge that the evolution of such species may have taken place ‘elsewhere’ (and perhaps slowly), that is, in some marine basin that has not yet been located. Gradualistic modes of evolution have also been recorded (e.g. Urbanek, Reference Urbanek1966; Sudbury, Reference Sudbury1958); it should be remembered, however, that even ‘established’ lineages are only hypotheses.
There have been several modifications of graptolite biozones. Numerical notation, giving biozones numbers rather than names, has been used by some authors. This practice is a corruption of Elles & Wood (Reference Elles and Wood1901–1918), who numbered their biozones as well as naming them. Elles and Wood, though, were simply providing a count. The numbering of biozones is not as well standardized as are names, and the use of different numerical schemes for different regions is a recipe for chaos. Numerical notation is best avoided.
Interregna are low-diversity levels that may have value in practical correlation, even defined on the occurrence of long-ranging species (and thus on the absence of other species). A familiar example is the Wenlock Gothograptus nassa/Pristiograptus dubius Interregnum established by Jaeger (Reference Jaeger1959). P. dubius is one of the longest-ranging graptolite species known (early Wenlock to late Ludlow), while G. nassa has been recorded from significantly below the interregnum up to the basal Ludlow (though outside Britain this species seems to be almost confined to its biozone: Porębska, Kozłowska-Dawidziuk & Masiak, Reference Porębska, Kozłowska-Dawidziuk and Masiak2004). Thus, certain identification of this interregnum hinges on the recognition of the underlying or overlying biozone. The nassa/dubius level has been subsequently treated and referred to as a biozone by most workers. Other ‘interregna’ have been recognized (e.g. a low diversity interval dominated by monograptids with hooked thecae in the upper turriculatus Biozone of central Wales: Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) and used informally. Some well-established biozones, also, effectively comprise low-diversity intervals separating more diverse and distinctive biozonal assemblages. For example, the use of the peltifer Biozone in Scotland was defined on the basis of few incoming taxa, but many species of the underlying gracilis Biozone are absent, while those defining the overlying wilsoni Biozone have not yet appeared. The difficulty of recognizing the peltifer Biozone led Williams et al. (Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004) to propose an alternative biozonal arrangement for the lower Caradoc interval, as discussed below.
The duration of graptolite biozones has been assessed in a number of ways. Average duration is estimated by dividing radiometric estimates for the duration of periods or epochs by the number of biozones (e.g. Hughes, Reference Hughes1995). Ordovician biozones are significantly longer, averaging some 2 Ma (Rushton, Reference Rushton1990), than Silurian biozones, which average < 1 Ma, and < 0.5 Ma if subzones and informal subdivisions are taken into account (Zalasiewicz, Reference Zalasiewicz1990). Within-period inequalities in length have been assessed by using radiometric dates that constrain epoch rather than period boundaries; in this way, Hughes (Reference Hughes1995) estimated average biozone durations, respectively, of 1.0, 0.44, 1.0 and 1.43 Ma for the Llandovery, Wenlock, Ludlow and Přídolí epochs. Finer-scale estimates have been made by using thicknesses of distal, deep-sea graptolitic mudrocks, assuming sedimentation rates to be roughly constant, and cross-checking by comparing sections in different parts of the world, and then applying radiometric age constraints. In this way, Churkin, Carter & Johnson (Reference Churkin, Carter and Johnson1977) estimated the persculptus Biozone as < 1 Ma in duration, while the gracilis Biozone apparently represents some 8 Ma.
Some of the most precise correlation achieved has been effected by bypassing the zonal concept altogether, and cross-correlating the first and last appearances of individual species in selected well-studied sections. Cooper & Lindholm (Reference Cooper and Lindholm1990) used such data to effect graphical correlation of early to mid-Ordovician sections; they showed that, globally, the appearances and disappearances of graptolites form a coherent pattern in time, enabling subdivision into about 50 time-slices within a 20 Ma interval. Further refinement of this approach has involved, for example, the ‘constrained optimization’ (CONOP) method described by Cooper & Sadler (Reference Cooper, Sadler, Gradstein, Ogg and Smith2004) that aimed at extracting the maximum information from individual species ranges, and that further refined estimates of the durations of individual biozones. Within depositional basins or graptolite provinces, such methodologies should see progessive advances in precision; between provinces, endemism and diachronous species ranges will limit progress.
6. British graptoloid biostratigraphy: a summary of previous work
Charles Lapworth (Reference Lapworth1878) first applied the concept of graptolite zones in Britain while working in the Southern Uplands of Scotland. Having examined a large number of sections, and following the Swedish work of Linnarsson (Hamilton, Reference Hamilton2001), he was able to subdivide the Hartfell and Birkhill Shale formations into eleven units based on distinct lithological differences, and found that these corresponded with differences in their accompanying graptolite assemblages which he called ‘zones’. He provided detailed lithological logs of the best sections, as well as a chart outlining the vertical ranges of all observed taxa from the sections studied. This was to be the model of procedure for all future biostratigraphical work involving graptolites. In the following year, Lapworth (Reference Lapworth1879–1880a) extended his study of graptolite zones to include all British and international material, identifying twenty zones for the Late Cambrian to Silurian interval. He provided range charts for the graptolite genera and species then known, and suggested that these could form a basis for detailed temporal subdivision of early Palaeozoic strata. His work was immediately put to the test by the Geological Survey of Scotland and was found to be of the greatest value (Rushton, Reference Rushton2001a).
These zones were recognizable in other parts of Britain, as later papers indicated, notably in the Stockdale Shales (Marr & Nicholson, Reference Marr and Nicholson1888) and the Skiddaw Slates (Marr, Reference Marr1894) of the Lake District. Key biostratigraphical work in Wales and the Welsh Borderland increased the number of known British graptolite species and biozones. Elles's (Reference Elles1900) study of the Wenlock Shales of the Welsh Borderland, Wood's (Reference Wood1900) on the Lower Ludlow Formation of the Welsh Borderland, and Herbert Lapworth's (Reference Lapworth1900) work on the Silurian sequence at Rhayader in mid-Wales, all provide detailed lithostratigraphies combined with species range charts; new biozones were introduced and existing ones altered, reflecting distinct local variations in the composition of the graptolite assemblages. Wood (Reference Wood1900) subdivided part of the Ludlow succession at Long Mountain into subzones. These workers also compared their range charts and faunal lists with information available from the rest of Britain and abroad, particularly Sweden.
‘A monograph of British graptolites’ (Elles & Wood, Reference Elles and Wood1901–1918) is a complete account of all the British planktonic graptolite species and subspecies known at that time (apart from Rhabdinopora and Corynoides), providing descriptions and figures of each taxon. The stratigraphical ranges of all the taxa were collated into a biostratigraphy comprising 36 graptolite biozones and subzones. This was the first time that one study on British graptolites had unified the taxonomy, biostratigraphy and chronostratigraphical importance of the group to such an extent. The monograph continues to provide the basis for British graptolite biostratigraphy, and its influence on global correlation remains profound, although many of the individual taxon descriptions and ranges have been revised in subsequent studies.
Graptolites were now established as a fundamental part of British Early Palaeozoic stratigraphical studies (e.g. Jones, Reference Jones1909, Reference Jones1947; Davies, Reference Davies1929; Jones & Pugh, Reference Jones and Pugh1935). Later, Sudbury (Reference Sudbury1958) beautifully demonstrated patterns of fine-scale evolution in the Llandovery triangulate monograptids of the Rheidol Gorge, Wales. Toghill (Reference Toghill1968a) updated the graptolite assemblages and biozones of the early Silurian Birkhill Shale Formation of Scotland. Studies on the Silurian strata of Northern England included accounts of the Wenlock and Ludlow (Rickards, Reference Rickards1967) and Llandovery (Rickards, Reference Rickards1970) graptolites of the Howgill Fells and of the Lake District (Hutt, Reference Hutt1974–1975). The results of these studies were collated into a major revision of British Silurian graptolite ranges and zonal descriptions (Rickards, Reference Rickards1976). No corresponding update of Elles & Wood's Ordovician range charts was produced prior to the one we provide in this report, although Jackson (Reference Jackson1962) provided a synthesis of the Arenig biostratigraphy of the Skiddaw Group of the Lake District.
Subsequent work includes Williams's descriptions of the classic Dob's Linn section (1982b, 1988), which became the Ordovician–Silurian boundary stratotype (Bassett, Reference Bassett1985), and in the Moffat and Girvan districts (Williams, Reference Williams1982a, Reference Williams1987, Reference Williams1994). Zalasiewicz (Reference Zalasiewicz1984, Reference Zalasiewicz1986) described early Ordovician graptolite material from North Wales, and Fortey & Owens (Reference Fortey and Owens1978, Reference Fortey and Owens1987) from South Wales.
Major British Geological Survey (BGS) mapping programmes in Wales, the Lake District and southern Scotland were all underpinned by graptolite biostratigraphy; large collections were made and described, and the biozonation itself was considerably refined. Rushton's (in Cooper et al. Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004) reorganization of Arenig graptolite biostratigraphy in the Skiddaw Group formed part of this work, as did the report of graptolites of Arenig age in the Ballantrae ophiolite complex in SW Scotland (Stone & Rushton, Reference Stone and Rushton1983). Studies of younger Ordovician strata included those of Zalasiewicz (Reference Zalasiewicz1992a), Zalasiewicz, Rushton & Owen (Reference Zalasiewicz, Rushton and Owen1995), Cave & Rushton (Reference Cave and Rushton1996) and Williams et al. (Reference Williams, Davies, Waters, Rushton and Wilby2003a, Reference Williams and Zalasiewicz2004), while the compilation of Scottish records by Rushton et al. (Reference Rushton, Stone and Hughes1996) demonstrated the continued importance of graptolite work in the stratigraphy and structure of the Southern Uplands of Scotland.
Remapping of central Wales by BGS prompted refinement of Llandovery and Wenlock graptolite biostratigraphy. There were studies of early to mid-Llandovery (Rhuddanian–Aeronian) sections and graptolite assemblages (Zalasiewicz, Reference Zalasiewicz1992b, Reference Zalasiewicz1996; Zalasiewicz & Tunnicliff, Reference Zalasiewicz and Tunnicliff1994). Spectacular progress was made in late Llandovery (Telychian) successions (Loydell, Reference Loydell1991a, Reference Loydell1992–1993a, Reference Loydell, Hailwood and Kidd1993b; Loydell & Cave, Reference Loydell and Cave1993, Reference Loydell and Cave1996; Zalasiewicz, Reference Zalasiewicz1994; Zalasiewicz, Loydell & Štorch, Reference Zalasiewicz, Loydell and Štorch1995; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). For example, the former Monograptus turriculatus Biozone was split into two separate biozones, each with several component subzones, and the former Monoclimacis crenulata Biozone into four separate biozones. Work on Elles's (1900) original Wenlock biozonal localities near Builth Wells (Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999; Williams & Zalasiewicz, Reference Williams and Zalasiewicz2004) resulted in a reorganization of graptolite biozones for that part of the stratigraphical column. Nevertheless, substantial sections of British early Palaeozoic biostratigraphy remain relatively neglected, for example, the Ludlow successions, and further refinement may be expected. The current biozonal schemes for the British Ordovician and Silurian are given in Figures 1 and 2.
7. Notes on the range charts
Two separate graptoloid range charts are given for the British Ordovician, one representing England and Wales (Figs 3–7) and the other Scotland (Figs 8–10), because significant regional differences in graptolite faunas exist. The separation of Scotland from England and Wales by the Iapetus Ocean resulted in faunal provincialism during the Ordovician, and for graptolites, this remained the case even upon the closure of Iapetus at the end of the period, probably as a result of environmental factors (Zalasiewicz, Rushton & Owen, Reference Zalasiewicz, Rushton and Owen1995). Fewer such problems exist in the Silurian, and a single sequence of graptolite biozones has been recognized throughout the UK (Figs 11–19). Sources for the ranges are provided (Tables 1–3) and, for ease of use, an index to taxa (Table 4).
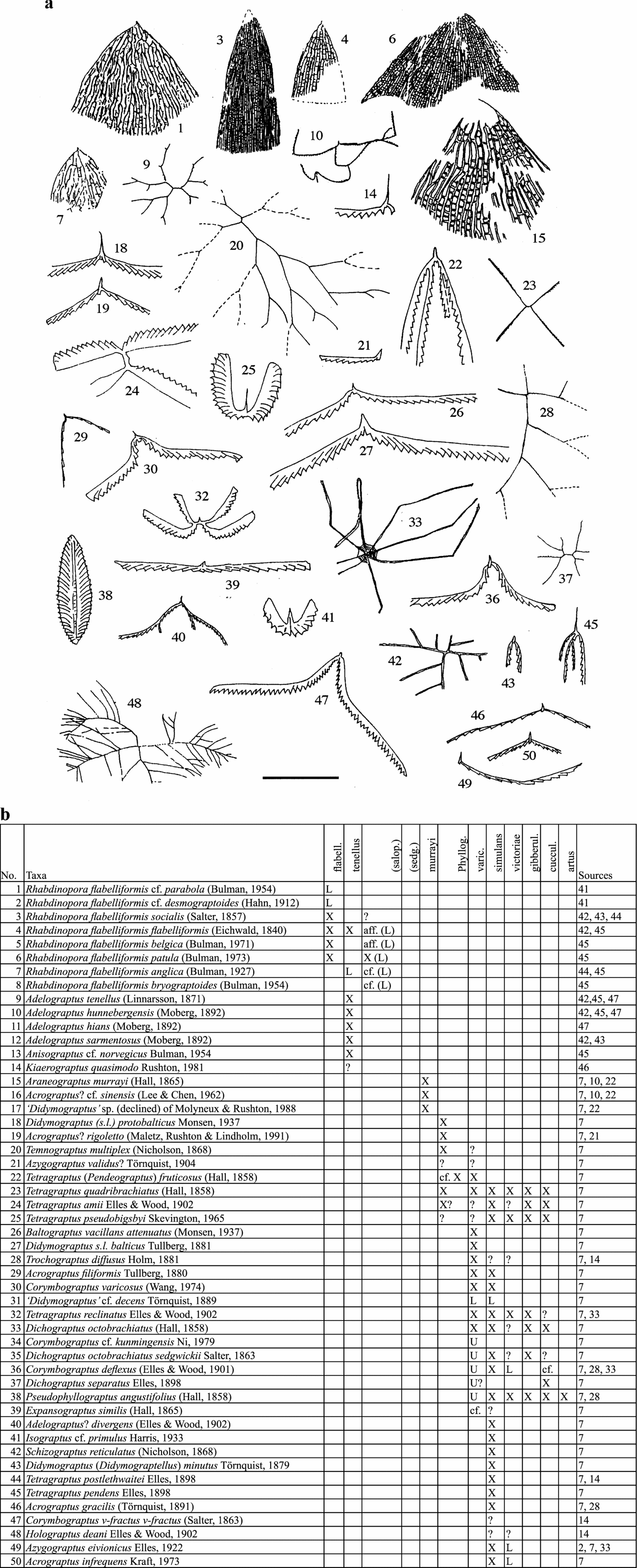
Figure 3. Ordovician graptolites (a) and their stratigraphical ranges (b) in England and Wales, flabelliformis to artus biozones. Scale bar represents 10 mm except for: 1, 23, 32, 40, 45, 47 (20 mm); 3, 4, 6, 7, 9, 10, 15, 23, 37, 42 (40 mm); 20, 28 (57 mm); 33 (80 mm); and 48 (320 mm). Key to symbols used in range charts: X – present; A – abundant; L – present in lower part; M – present in middle part; U – present in upper part; cf. – similar to but not certainly identified; aff. – related to but not identical; ? – doubtful; ?? – very doubtful; < – range extends lower; > – range extends higher.
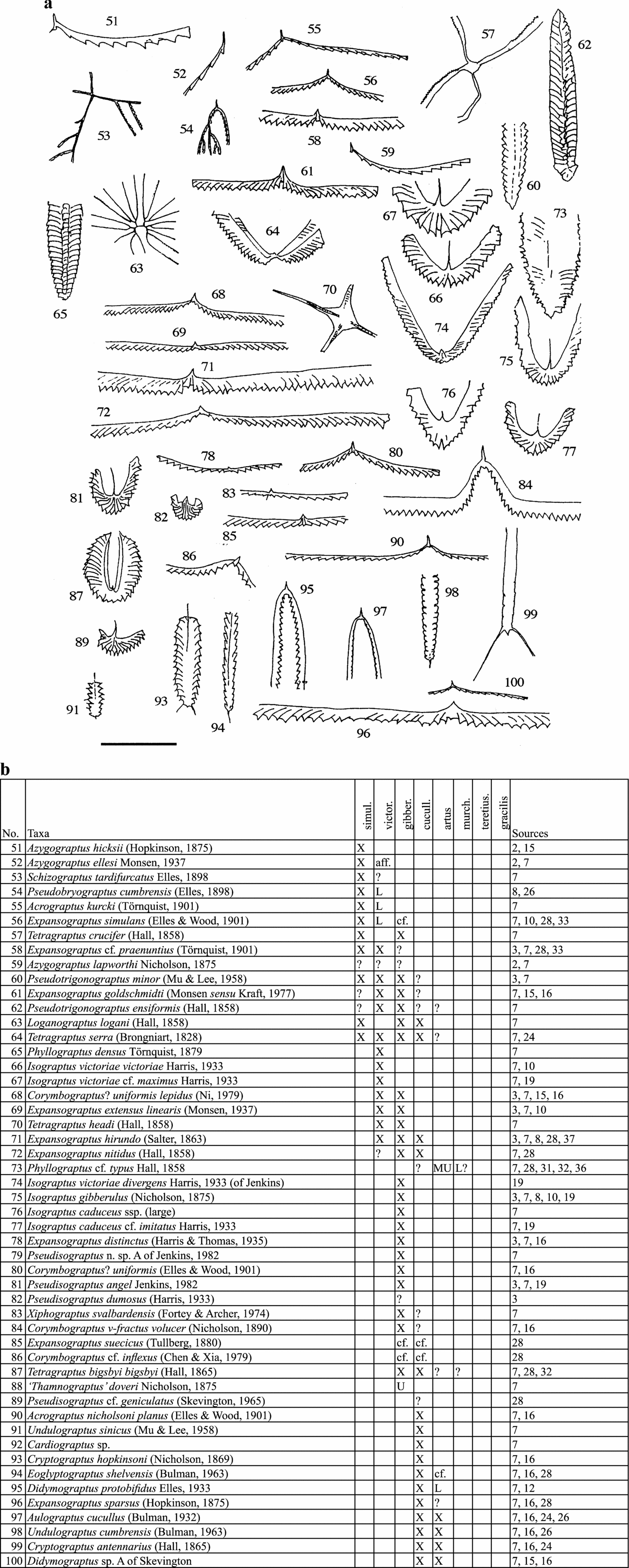
Figure 4. Ordovician graptolites (a) and their stratigraphical ranges (b) in England and Wales, simulans to gracilis biozones. Scale bar represents 10 mm except for: 54, 64 (20 mm); 53, 57, 63, 70 (40 mm). For key to symbols in range charts, see Figure 3.

Figure 5. Ordovician graptolites (a) and their stratigraphical ranges (b) in England and Wales, cucullus Biozone to caudatus Subzone. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 6. Ordovician graptolites (a) and their stratigraphical ranges (b) in England and Wales, murchisoni to persculptus biozones. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 7. Ordovician graptolites (a) and their stratigraphical ranges (b) in England and Wales, foliaceus to persculptus biozones. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 8. Ordovician graptolites (a) and their stratigraphical ranges (b) in Scotland, Lancefieldian 2 to wilsoni biozones. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 9. Ordovician graptolites (a) and their stratigraphical ranges (b) in Scotland, gracilis to linearis biozones. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 10. Ordovician graptolites (a) and their stratigraphical ranges (b) in Scotland, caudatus Subzone to persculptus Biozone. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.
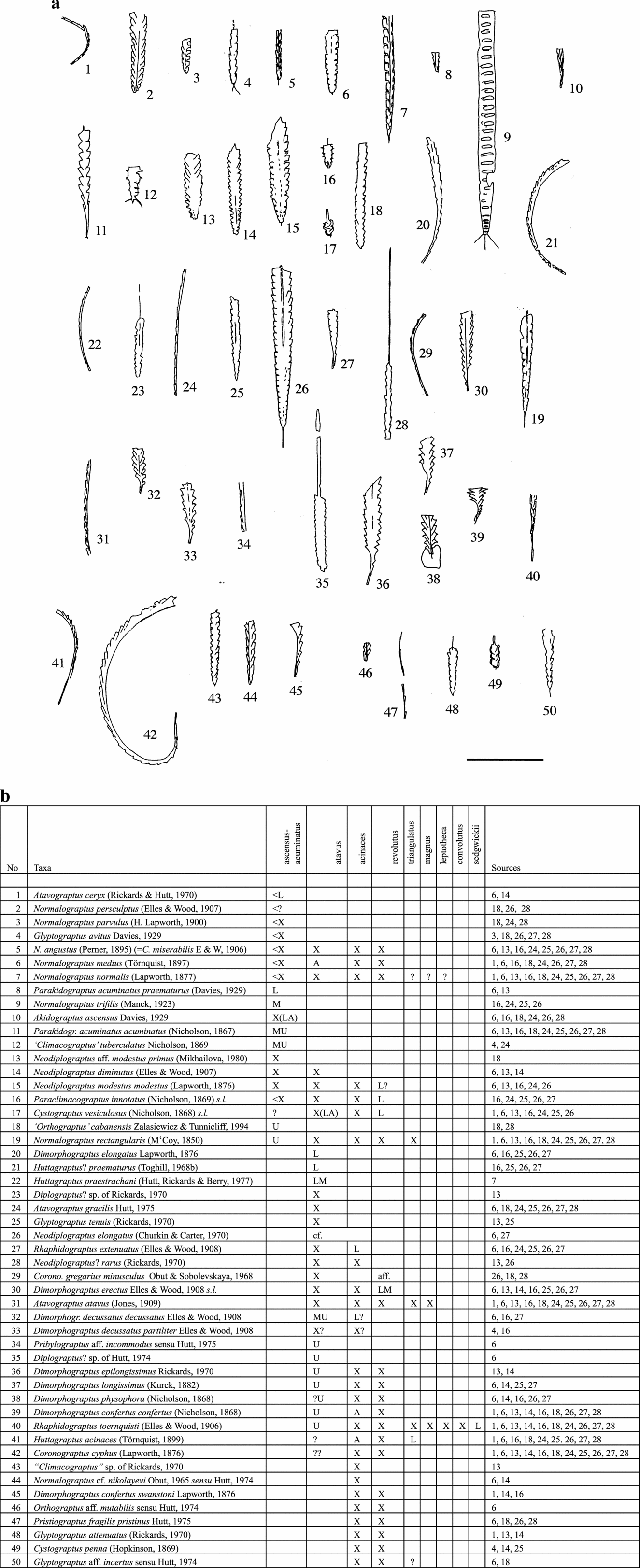
Figure 11. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, ascensus–acuminatus to sedgwickii biozones. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 12. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, atavus Biozone to gemmatus Subzone. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 13. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, triangulatus Biozone to utilis Subzone. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 14. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, convolutus Biozone to galaensis Subzone. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 15. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, runcinatus Subzone to crenulata Biozone. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 16. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, proteus Subzone to riccartonensis Biozone. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 17. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, spiralis to ludensis biozones. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 18. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, dubius to bohemicus biozones. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.

Figure 19. Silurian graptolites (a) and their stratigraphical ranges (b) in Great Britain, scanicus to bohemicus biozones. Scale bar represents 10 mm. For key to symbols in range charts, see Figure 3.
Table 1. Ordovician range chart sources (England & Wales) (for Figs 3–7)

Table 2. Ordovician range chart sources (Scotland) (for Figs 8–10)

Table 3. Silurian range chart sources (for Figs 11–19)

Table 4. Index to taxa; O/EW/no. = Ordovician/England and Wales/taxon number in range chart (Figs 3–7); O/Sc/no. = Ordovician/Scotland/taxon number in range chart (Figs 8–10); S/no. = Silurian/taxon number in range chart (Figs 11–19)

Elles & Wood (Reference Elles and Wood1901–1918) assigned their species to relatively few genera. There has since been an expansion of generic concepts; those used here mostly follow Strachan's (Reference Strachan1996–1997) review of British graptolites. Strachan gave full bibliographic references to the species known to him, and those references are not repeated here, though references are given here for species recognized in Britain subsequent to Strachan's compilation.
These range charts were commenced by JAZ and AWAR in work for the British Geological Survey, substantially developed by LT during Ph.D. studies, and subsequently added to by the other authors. Species recorded in the range charts have been fully documented in papers, monographs or memoirs. Records in conference abstracts are not included.
In the range charts: X – present; A – abundant; L – present in lower part; M – present in middle part; U – present in upper part; cf. – similar to but not certainly identified; aff. – related to but not identical to named species; ? – doubtful record; ?? – very doubtful; < – range extends lower; > – range extends higher.
The thumbnail sketches of the graptolite taxa on the pages facing the charts are intended to remind the general reader of the overall character of the various species. They are not adequate for identification.
8. Ordovician
R. A. Cooper (Reference Cooper1999) discussed the global correlation of Tremadocian graptolites, recognized nine graptolite chronozones, and in his figure 1 showed the ranges of the main types of graptolites in the Tremadocian. In Britain, only a few of those chronozones are recognized. They are distributed between two locally defined stages proposed by Fortey et al. (Reference Fortey, Harper, Ingham, Owen and Rushton1995), the Cressagian below and the Migneintian. In Britain, the upper Tremadocian is almost devoid of graptolites, with two widely recognized trilobite biozones, the Conophrys salopiensis and Angelina sedgwickii zones, occupying most of the Migneintian (Fig. 1, 3b).
8.a. Rhabdinopora flabelliformis Biozone
The appearance of Rhabdinopora flabelliformis (formerly Dictyonema flabelliforme) has for a long time been taken as a marker for the beginning of the Tremadocian. Although the base of the Ordovician, as currently defined, is now taken at the appearance of the conodont Iapetognathus fluctivagus (Cooper, Nowlan & Williams, Reference Cooper, Nowlan and Williams2001), the appearance of the earliest subspecies of Rhabdinopora flabelliformis remains a good approximation to the base of the Ordovician and is applicable in clastic sequences where conodonts are rare (Landing et al. Reference Landing, Bowring, Davidek, Rushton, Fortey and Wimbledon2000).
The lowest of Cooper's biozones, that of Rhabdinopora praeparabola, is not yet known in Britain. The flabelliformis Biozone in Britain appears to include the equivalents of Cooper's parabola and matanensis biozones, and possibly part of the overlying anglica Biozone. At Brin-llin-fawr, North Wales, the occurrence of taxa closely comparable to the early subspecies Rhabdinopora flabelliformis parabola and R. f. desmograptoides at the base of the flabelliformis Biozone (Legrand in Rushton et al. Reference Rushton, Owen, Owens and Prigmore1999, fig. 7.4) suggests that the parabola Biozone may prove recognizable. Higher in the flabelliformis Biozone, R. flabelliformis flabelliformis and R. f. socialis are the most widely recognized forms, and are known from north Wales, Shropshire and numerous boreholes in Warwickshire and Buckinghamshire (Bulman & Rushton, Reference Bulman and Rushton1973; Old, Sumbler & Ambrose, Reference Old, Sumbler and Ambrose1987; Bridge et al. Reference Bridge, Carney, Lawley and Rushton1998). Although Rhabdinopora f. anglica appears to occur high in the flabelliformis Biozone, it is not yet clear whether a separate anglica Biozone can be recognized. The borehole record of Kiaerograptus quasimodo Rushton, Reference Rushton1981 is anomalous because it lies in the midst of borehole records proving the flabelliformis Biozone (Old, Sumbler & Ambrose, Reference Old, Sumbler and Ambrose1987), whereas Kiaerograptus occurs typically in the equivalents of the Migneintian Stage.
8.b. Adelograptus tenellus Biozone
This biozone is recognized by the appearance of Adelograptus (formerly Clonograptus) tenellus and A. hunnebergensis. Cooper (Reference Cooper1999) recognized an Adelograptus Biozone above the Rhabdinopora flabelliformis anglica Biozone, but these are not readily separated in Britain. In the ‘Transition Beds’ of the Shineton Shales of Shropshire, Rhabdinopora flabelliformis subspp., including R. f. anglica, alternate with Adelograptus spp. through a relatively small thickness of strata, above which Rhabdinopora disappears. It appears that an equivalent situation pertains to the English subcrop.
8.c. Higher Tremadocian zones
Above the tenellus Biozone, graptolites are practically absent from strata in Britain until the top of the Tremadocian, and Migneintian biostratigraphy has depended on evidence from trilobites and acritarchs. There are, however, two records of Rhabdinopora flabelliformis subspp. from strata referred to the Conophrys salopiensis trilobite Biozone as recognized on the basis of trilobites and acritarchs: one from the Deanshanger Borehole in Buckinghamshire (Bulman & Rushton, Reference Bulman and Rushton1973), the other from the ‘Upper Dictyonema Band’ in the Gwynant Valley, north of Cadair Idris (Pratt, Woodhall & Howells, Reference Pratt, Woodhall and Howells1995, p. 16). Both records presumably represent a late development of Cooper's ‘Assemblage 3’ (Cooper, Reference Cooper1979) and pre-date his P. antiquus and Kiaerograptus biozones.
8.d. Araneograptus murrayi Biozone
This biozone was first used in Britain by Cooper et al. (Reference Cooper, Rushton, Molyneux, Moore and Webb1995), in reference to the Skiddaw Group from the English Lake District. The A. murrayi Biozone is currently the only recognizable upper Tremadocian graptoloid biozone in the British sequence. The base is taken on the appearance of the biozone fossil accompanied by Acrograptus? cf. sinensis and a declined Didymograptus (s.l.) sp. (Molyneux & Rushton, Reference Molyneux and Rushton1988).
8.e. Tetragraptus phyllograptoides Biozone
The base of this biozone equates to that of the Floian Stage of the Ordovician (Bergström, Löfgren & Maletz, Reference Bergström, Löfgren and Maletz2004). Cooper et al. (Reference Cooper, Rushton, Molyneux, Moore and Webb1995) adopted the T. phyllograptoides Biozone, an established Scandinavian interval (Lindholm, Reference Lindholm1991), based on the similarity between the Skiddaw Group assemblages and those characteristic of the upper part of this biozone in Scandinavia (Maletz, Rushton & Lindholm, Reference Maletz, Rushton and Lindholm1991). The biozone fossil is not known from the Lake District, but this biozone may be recognized by the appearance of Acrograptus? protobalticus, A.? rigoletto and Temnograptus multiplex. Other taxa appearing are Tetragraptus quadribrachiatus and Tetragraptus (Pendeograptus) cf. fruticosus. Species diversity is still relatively low in the phyllograptoides Biozone, although greater than that of the previous biozone, and includes the first tetragraptids and a temnograptid (Fig. 3). This interval has not been recorded elsewhere in Britain (Cooper et al. Reference Cooper, Rushton, Molyneux, Moore and Webb1995, Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004).
In Scotland, graptoloid assemblages from the lower to middle part of the Ordovician sequence resemble those in Australasia more closely than the English and Welsh assemblages. This prompted Stone & Rushton (Reference Stone and Rushton1983) to use the Australasian graptolite biostratigraphical scheme (VandenBerg & Cooper, Reference VandenBerg and Cooper1992) during their work on the faunas of the Ballantrae ophiolite complex. The earliest assemblages from Ballantrae contain Tetragraptus approximatus, either by itself or associated with T. (Pendeograptus) fruticosus. The latter association is characteristic of the lowermost Bendigonian (Be1), but the former could originate from the Lancefieldian (La3). Assemblages from Pinbain that contain T. cf. decipiens, Acrograptus cf. filiformis and Paradelograptus sp. are less definite, but suggest an early Bendigonian (Be1–2) age (Rushton et al. Reference Rushton, Stone, Smellie and Tunnicliff1986).
8.f. Corymbograptus varicosus Biozone
Following recent revision of Skiddaw Group graptoloid biostratigraphy (Cooper et al. Reference Cooper, Rushton, Molyneux, Moore and Webb1995), the C. varicosus Biozone has replaced the previously used Corymbograptus deflexus Biozone (Jackson, Reference Jackson and Moseley1978; Fortey & Owens, Reference Fortey and Owens1990). Cooper et al. (Reference Cooper, Rushton, Molyneux, Moore and Webb1995, Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004) found C. deflexus to be rare in this interval, whereas C. varicosus is abundant. There is no clear definition for the base of the varicosus Biozone, due to the lack of known suitable fossiliferous strata in the Lake District (Rushton et al. Reference Rushton, Owen, Owens and Prigmore1999, p. 266; Cooper et al. Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004, p. 13). This assemblage displays a marked increase in species and generic diversity. New appearances include Baltograptus vacillans attenuatus, Didymograptus aff. balticus, Tetragraptus reclinatus, T. (P.) fruticosus, Expansograptus cf. decens and Acrograptus filiformis. Some species are known only from the upper part of the biozone: C. deflexus, Pseudophyllograptus angustifolius, Dichograptus octobrachiatus sedgwickii, Schizograptus tardifurcatus and Expansograptus cf. similis (Fig. 3).
8.g. Expansograptus simulans Biozone
This biozone straddles the interval between the ‘second’ (Floian) and ‘third’ (Dapingian) stages of the Ordovician (Cooper & Sadler, Reference Cooper, Sadler, Gradstein, Ogg and Smith2004). The former Didymograptus nitidus Biozone was renamed on the basis that the former zone fossil does not occur in this interval in the Skiddaw Group (Cooper et al. Reference Cooper, Rushton, Molyneux, Moore and Webb1995). The base of the biozone is defined on the incoming of Expansograptus simulans and Acrograptus infrequens (Cooper et al. Reference Cooper, Rushton, Molyneux, Moore and Webb1995; Rushton in Cooper et al. Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004). A significant rise in graptolite diversity can be seen at this horizon, with about 33 taxa recorded (Fig. 3). Several new forms are confined to this biozone, including Didymograptus minutus, Tetragraptus postlethwaitei, Adelograptus? divergens and Tetragraptus (Pendeograptus) pendens. Other noteworthy appearances are Azygograptus eivionicus, Azygograptus lapworthi, Pseudobryograptus cumbrensis, Acrograptus? kurcki, Loganograptus logani and Tetragraptus crucifer. Cooper et al. (Reference Cooper, Rushton, Molyneux, Moore and Webb1995) suggested that the presence of Isograptus cf. primulus in this interval indicates a correlation with the Chewtonian Stage (Ch 1–2).
The contemporaneous Scottish taxa differ markedly from those of England and Wales, with the exception of Tetragraptus serra and Tetragraptus reclinatus, which occur in both successions (Fig. 8). Stone & Rushton's (Reference Stone and Rushton1983) work at Ballantrae revealed graptolite assemblages containing Didymograptus cf. protomurchisoni, Tetragraptus bigsbyi cf. askerensis, Expansograptus? aff. geometricus, Didymograptus cf. protoindentus, Sigmagraptus praecursor, Paradelograptus sp., Tetragraptus (P.) fruticosus and Tetragraptus cf. kindlei, which they found consistent with a Chewtonian age.
8.h. Isograptus victoriae victoriae Biozone
The recognition of distinct, biostratigraphically important graptolites within the old Isograptus gibberulus Biozone of the Skiddaw Group led to the proposal of a lower I. victoriae victoriae Biozone and an upper I. gibberulus Biozone (Rushton in Cooper et al. Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004, p. 17). The I. v. victoriae Biozone is characterized by the appearance of the zone fossil, which is confined to this interval. However, the index species is rare in the Lake District, leading to difficulties in identifying the biozonal boundaries. Horizontal didymograptids, such as Expansograptus hirundo, E. cf. nitidus and E. extensus linearis help to characterize the victoriae and gibberulus biozones. Other forms limited to the victoriae Biozone are Phyllograptus densus and Isograptus victoriae cf. maximus, while the appearance of E. hirundo and E. cf. nitidus (Figs 3, 4) is recorded in the Shelve area of Shropshire (Strachan, Reference Strachan1986).
No graptolite assemblage corresponding to this interval has been reported from Scotland (Fig. 8).
8.i. Isograptus gibberulus Biozone
This biozone, as restricted by Rushton (in Cooper et al. Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004, p. 17) in the Skiddaw Group of the English Lake District, is defined by the appearance of I. gibberulus. Cooper et al. (Reference Cooper, Rushton, Molyneux, Moore and Webb1995) suggested that the former I. gibberulus Biozone equated with the Australasian Castlemainian Stage (Ca 1–4), but, as now restricted, it represents the upper part of the Castlemainian (Ca 3–4) (Cooper et al. Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004). The identification of Pseudisograptus dumosus at this horizon from the Aberdaron area of Wales, and Isograptus caduceus cf. imitatus from Grisedale Pike in the Lake District (Jenkins, Reference Jenkins1982) seems to confirm this. Other noteworthy appearances include Pseudisograptus angel, Corymbograptus v-fractus volucer, Tetragraptus bigsbyi bigsbyi, Corymbograptus? uniformis and Xiphograptus svalbardensis (Fig. 4).
In Scotland, an assemblage obtained from the North Ballaird Borehole in the Ballantrae ophiolite complex contained elements comparable to those for this interval in England and Wales (Stone & Strachan, Reference Stone and Strachan1981), but also other taxa (Arienigraptus gracilis, Isograptus caduceus australis, Tylograptus? and Yutagraptus v-deflexus) that indicate a latest Castlemainian or early Yapeenian age (Stone & Rushton, Reference Stone and Rushton2003) (Fig. 8). In a review of this assemblage, Maletz (Reference Maletz2004) referred the supposed A. gracilis to Pseudisograptus initialis Maletz, Reference Maletz2001 and the Tylograptus? to a species of Meandrograptus. However, although Stone & Rushton (Reference Stone and Rushton2004) gave reasons for rejecting the transfer of their specimens of I. caduceus australis to Isograptus caduceus imitatus, they accepted Maletz's suggestion that the North Ballaird assemblage is best restricted to the late Castlemainian (Ca4).
As yet, no graptolite assemblages have been reported from Scotland at a higher stratigraphical level, until the base of the Nemagraptus gracilis Biozone at the base of the Sandbian Stage (base of Upper Ordovician) (Fig. 8).
8.j. Aulograptus cucullus Biozone
The base of the cucullus Biozone equates with that of the Darriwilian Stage (Cooper & Sadler, Reference Cooper, Sadler, Gradstein, Ogg and Smith2004). Rushton (in Cooper et al. Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004, p. 19) renamed the former Didymograptus hirundo Biozone the Aulograptus cucullus Biozone to take into account the fact that Expansograptus hirundo originates much earlier, in the victoriae Biozone. The base of the cucullus Biozone is characterized by the incoming of A. cucullus, and many other taxa originate in the biozone, among them the first cryptograptids, Oelandograptus austrodentatus anglicus, Acrograptus nicholsoni planus, Didymograptus protobifidus, Expansograptus sparsus, Undulograptus cumbrensis, U. sinicus and Eoglyptograptus shelvensis; these include the first biserial graptolites in the British Ordovician sequence (Fig. 4). Assemblages of the A. cucullus Biozone have also been recorded from the Shelve district of Shropshire (Strachan, Reference Strachan1986). The range chart shows a few taxa indicating the upper part of this biozone (Fig. 5); these are Acrograptus acutidens, and possibly Amplexograptus confertus and Cryptograptus tricornis schaeferi.
8.k. Didymograptus artus Biozone
The base of the D. artus Biozone corresponds with that of the locally defined Llanvirn Series. The biozone was established by Fortey & Owens (Reference Fortey and Owens1987) to replace the traditional Didymograptus bifidus Biozone, which was based on incorrect identifications. This interval displays a highly diverse and distinctive graptolite assemblage (Fig. 5), with the majority of taxa appearing at the base of the biozone, which is characterized by the zone fossil and Didymograptus spinulosus (Fortey & Owens, Reference Fortey and Owens1987; Fortey, Beckly & Rushton, Reference Fortey, Beckly and Rushton1990; Rushton in Cooper et al. Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004). Approximately 31 species originate in the biozone, 21 of which have ranges restricted to it. Several genera are recorded for the first time, notably Glossograptus and Climacograptus. The artus Biozone has good potential for further biostratigraphical subdivision. While most elements of the assemblage span the entire biozone, there is a distinguishable middle to upper biozonal assemblage containing Pseudoclimacograptus scharenbergi, P. angulatus, Trichograptus fragilis, Diplograptus hollingworthi and Nicholsonograptus fasciculatus (Fig. 5). A few taxa help differentiate the lower part of the biozone, such as D. protobifidus and Expansograptus sparsus, which originate in the cucullus Biozone but disappear in the lower part of the artus Biozone. The D. artus Biozone has been widely recognized in northern England and Wales (Elles, Reference Elles1940; Strachan, Reference Strachan1986; Fortey & Owens, Reference Fortey and Owens1987; Fortey, Beckly & Rushton, Reference Fortey, Beckly and Rushton1990; Cooper et al. Reference Cooper, Rushton, Molyneux, Moore and Webb1995, Reference Cooper, Fortey, Hughes, Molyneux, Moore, Rushton and Stone2004).
8.l. Didymograptus murchisoni Biozone
Graptolite assemblages characteristic of this biozone have been recorded from the Builth–Llandrindod Wells district of central Wales (Elles, Reference Elles1940; Hughes, Reference Hughes1989; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997), the Shelve area of Shropshire (Strachan, Reference Strachan1986; Hughes, Reference Hughes1989), the Skiddaw Group strata of the Tarn Moor Tunnel in the Lake District (Wadge, Nutt & Skevington, Reference Wadge, Nutt and Skevington1972), the Fishguard area of South Wales (Davies et al. Reference Davies, Waters, Wilby, Williams and Wilson2003) and Abereiddy Bay (Jenkins, Reference Jenkins1987). The base of the murchisoni Biozone sees the incoming of the zone fossil, which is restricted to this biostratigraphical interval (Fig. 5). Other new appearances include Diplograptus foliaceus, Cryptograptus tricornis tricornis, Pseudoclimacograptus angulatus magnus, P. angulatus micidus? and Didymograptus speciosus? (Fig. 5). Pterograptus elegans?, Lasiograptus retusus and Didymograptus nanus are reported only from the lower part of the biozone, whereas Diplograptus? decoratus, Amplexograptus caelatus and Normalograptus brevis appear in its middle to upper part; this suggests that refinement of the biostratigraphy may be possible.
8.m. Hustedograptus teretiusculus Biozone
The base of this biozone is delineated by the disappearance of D. murchisoni (Hughes, Reference Hughes1989), as the zone fossil originates in the middle part of the interval (Fig. 6). The lower part of the biozone therefore is an interregnum. The only taxa to appear at the basal boundary are Dicellograptus divaricatus divaricatus, Climacograptus antiquus and possibly Normalograptus euglyphus. Approximately half of the total species occurring at this biostratigraphical level appear in the middle to upper part of the biozone. These data provide a strong case for a biostratigraphical review of the graptolite faunas found in this interval. However, Hughes (Reference Hughes1989) remarked that Elles's (Reference Elles1940) attempt to subdivide the teretiusculus and gracilis biozones in the Builth area was incorrect, as her refinement was based on taxonomically unsound species and poorly preserved material. The teretiusculus Biozone, as it is presently understood, is recorded from Wales and Shropshire (Elles, Reference Elles1940; Strachan, Reference Strachan1986; Hughes, Reference Hughes1989; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
8.n. Nemagraptus gracilis Biozone
The base of this biozone equates with that of the Sandbian Stage of the Ordovician (Cooper & Sadler, Reference Cooper, Sadler, Gradstein, Ogg and Smith2004), or with that of the Caradoc of British usage. This biostratigraphic interval was first described by Lapworth (Reference Lapworth1879–1880) and defined by the incoming of the dicellograptids and nemagraptids. Subsequent work led to a revised definition of the base of the biozone, which is now taken at the incoming of N. gracilis (Finney & Bergström, Reference Finney, Bergström, Hughes and Rickards1986). Assemblages representing this biozone have been recorded from the Builth region of Wales and the Shelve area of the Welsh Borderland (Elles, Reference Elles1940; Strachan, Reference Strachan1986; Hughes, Reference Hughes1989; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). However, although the position of the base of the gracilis Biozone at Builth Wells is unclear (Hughes, Reference Hughes1989; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997), Bettley, Fortey & Siveter (Reference Bettley, Fortey and Siveter2001) have identified a correlatable base to the biozone west of Shelve and at Meidrim, south Wales. The Anglo-Welsh N. gracilis assemblage is diverse, containing some 26 incoming species, as well as many longer-ranging forms from underlying zones (Fig. 6). Several species appear in the upper part of the biozone, and although only one of these, Lasiograptus pusillus, seems limited to this level, this assemblage may potentially be biostratigraphically useful. Incoming upper zonal taxa include Climacograptus bicornis, Dicranograptus nicholsoni nicholsoni, Amplexograptus perexcavatus and Glossograptus hincksii hincksii.
Graptolites reappear dramatically in the Ordovician sequence of Scotland in the gracilis Biozone. Following the sparse, low-diversity assemblages of the Scottish lower to middle Arenig interval of British usage, and the absence of Llanvirn biozones, a varied and abundant graptolite assemblage of N. gracilis age (about 75 taxa) has been recorded from southern Scotland (Stone, Reference Stone1995; Rushton, Tunnicliff & Tripp, Reference Rushton, Tunnicliff and Tripp1996; Armstrong et al. Reference Armstrong, Rushton, Owen and Floyd1998; Williams et al. Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004), including the Girvan district (Rushton, Reference Rushton2001b). From this biozone upwards, the Scottish graptolite succession is more directly comparable to that of England and Wales than in the Lower Ordovician, although the two biostratigraphical schemes still reflect significant assemblage differences (Fig. 1), making precise correlation difficult (Zalasiewicz, Rushton & Owen, Reference Zalasiewicz, Rushton and Owen1995; Williams et al. Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004). In Scotland, the gracilis Biozone is similarly defined as in England and Wales, although Stone (Reference Stone1995) commented on the importance of Expansograptus? superstes and Dicellograptus intortus to the characteristic gracilis assemblage (Figs 8, 9). D. intortus appears lower in the Anglo-Welsh sequence, however, and the exact position of the base of the gracilis interval cannot be clearly recognized in Scotland (Rushton, Reference Rushton1990; see also Williams et al. Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004). Recent work by Williams et al. (Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004) has restricted the gracilis Biozone as outlined in the next section.
8.o. Diplograptus foliaceus Biozone
The D. foliaceus Biozone is an Anglo-Welsh biostratigraphical unit, traditionally employed for the graptolite assemblages found between those of the underlying gracilis Biozone, and the Dicranograptus clingani Biozone above (Fig. 1). It was formerly known as the Diplograptus multidens Biozone; however, the taxonomic validity of D. multidens has been questioned (Hughes, Reference Hughes1989), and we follow Bettley, Fortey & Siveter (Reference Bettley, Fortey and Siveter2001) in naming the biozone after its suggested senior synonym, D. foliaceus. Much of Elles & Wood's (Reference Elles and Wood1901–1918) and Elles's (Reference Elles1940) earlier work on this biozone from the Shelve inlier and the type sections of the Caradoc area of Wales must be treated with caution, as many of the original taxa have not been found following recent studies (Hughes, Reference Hughes1989). Nonetheless, work carried out by the BGS in central Wales has confirmed the existence of a distinct and stratigraphically useful foliaceus Biozone assemblage (Figs 6, 7) with several newly occurring taxa (Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997; Williams et al. Reference Williams, Davies, Waters, Rushton and Wilby2003a). An upper biozonal assemblage with biostratigraphical potential has been identified (Fig. 7), including Dicranograptus nicholsoni minor and Dicellograptus patulosus.
8.p. Climacograptus bicornis Biozone (in Scotland)
Williams et al. (Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004) recently formalized the use of this zone for Scottish strata, having reviewed all the Scottish early Caradoc graptolite assemblages preserved from the 19th century geological survey of southern Scotland (Peach & Horne, Reference Peach and Horne1899) and from the work of the British Geological Survey during the 1980s and 1990s. The bicornis Biozone corresponds to the range of Climacograptus bicornis, commencing with its appearance in the N. gracilis–C. bicornis faunas from the Glenkiln Shales (in beds that were formerly assigned to the gracilis Biozone) and ranging up almost to the base of the Dicranograptus clingani Biozone in the lower Hartfell Shales (S. H. Williams, Reference Williams1994). M. Williams et al. (Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004) divided the bicornis Biozone into two subzones, the apiculatus–ziczac Subzone and the wilsoni Subzone (Fig. 20), as described below.

Figure 20. Comparison of presently adopted and earlier Scottish zonal schemes for the gracilis to clingani biozonal interval.
Formerly, two zones were recognized in the Scottish strata that follow the last appearance of Nemagraptus gracilis and precede the advent of the Dicranograptus clingani Biozone; these were known as the Climacograptus peltifer and C. wilsoni biozones. There are problems with the recognition of the C. peltifer Biozone, partly because the validity of the index species is doubtful (Strachan, Reference Strachan1971; Riva, Reference Riva and Bassett1976), and also because the original, supposedly diagnostic, peltifer assemblage (Elles, Reference Elles1925) is much less diverse than the underlying N. gracilis-bearing assemblages and contains few elements that do not occur in the gracilis Biozone (Rushton, Reference Rushton1990; Williams et al. Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004). The long-recognized wilsoni Biozone is more distinctive where it is found, but is of relatively local occurrence and is not widely recorded. Williams et al. (Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004) reduced it to a subzone of the bicornis Zone.
The correlation of the zonal units in the gracilis to clingani interval is not a simple one-to-one correlation, because in the new arrangement, the gracilis Biozone has a reduced stratigraphical range, characterized by the range of N. gracilis below the first appearance of C. bicornis. The upper and better-known assemblages (e.g. from the Glenkiln Shale) that were formerly assigned to the gracilis Biozone are now referred to the lower part of the bicornis Subzone, namely the apiculatus–ziczac Subzone (see Figs 8, 9).
8.p.1. Orthograptus apiculatus–Dicranograptus ziczac Subzone (in Scotland)
This subzone is recognized by the first appearances (FADs) of a large number of taxa; Williams et al. (Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004, fig. 2) listed about 40 that first appear at about this level, though because good measured sections are lacking, the FADs are not claimed to be precisely synchronous. The subzone is named after two distinctive species that are fairly widely recorded (Williams et al. Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004, p. 101) and have similar ranges. The apiculatus–ziczac Subzone differs significantly from the peltifer Zone as previously used because N. gracilis is present in its lower part, together with such taxa as Dicranograptus furcatus minimus, Expansograptus? superstes and Hallograptus spp. The assemblages in the lower part of the apiculatus–ziczac Subzone, in which the range of N. gracilis overlaps with those of C. bicornis and the taxa named above, are well known from the Glenkiln Shales of southern Scotland, and were formerly assigned to the extended concept of the N. gracilis Biozone. The upper part of the apiculatus–ziczac Subzone, above the range of N. gracilis, is a weakly characterized interval that may, at least in part, correspond to the peltifer Biozone (Williams et al. Reference Williams, Rushton, Wood, Floyd, Smith and Wheatley2004, p. 105), and such faunas have been recorded in the higher Glenkiln Shales.
8.p.2. Climacograptus wilsoni Subzone (in Scotland)
Lapworth (Reference Lapworth1878, p. 308) established the C. wilsoni Biozone during his work on the Moffat Series of the Southern Uplands, and it remains a recognizable part of the Scottish biostratigraphical scheme (Williams, Reference Williams1994) although it has not been recognized elsewhere in Great Britain (Elles & Wood, Reference Elles and Wood1901–1918; Elles, Reference Elles1925, Reference Elles1937). However, a taxonomic and biostratigraphical review of the wilsoni Biozone by Williams (Reference Williams1994) confirmed the value of this graptolite assemblage in southern Scotland. A type section was designated at Hartfell Spa (Williams, Reference Williams1994) and the biozone has been recognized elsewhere (Stone, Reference Stone1995; Williams, Reference Williams1994). The wilsoni assemblage has fewer taxa than the ‘peltifer assemblage’, and many of them continue from the lower biozone (Fig. 9). The base of the C. wilsoni Biozone is defined by the appearance of the index species; other incoming taxa are Orthograptus calcaratus vulgatus, Corynoides curtus, C. calicularis, Dicellograptus angulatus and Orthograptus of the amplexicaulis group.
8.q. Dicranograptus clingani Biozone
The D. clingani Biozone was established in the Moffat area by Lapworth (Reference Lapworth1878, p. 308) and has been widely recognized in Scotland (Williams, Reference Williams1982a; Stone, Reference Stone1995; Floyd, Reference Floyd1999) and also in Wales (Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997; Howells & Smith, Reference Howells and Smith1997; Young, Gibbons & McCarroll, Reference Young, Gibbons and McCarroll2002; Williams et al. Reference Williams, Davies, Waters, Rushton and Wilby2003a). Work at Hartfell Score in the Southern Uplands of Scotland and Whitland in south Wales led Zalasiewicz, Rushton & Owen (Reference Zalasiewicz, Rushton and Owen1995) to erect two subzones within the clingani interval, clearly defined by distinct graptolite assemblages and recognizable in both regions (Fig. 1), although there is some variation in assemblage composition (Zalasiewicz, Rushton & Owen, Reference Zalasiewicz, Rushton and Owen1995). The potential for subdivision of the biozone in the Southern Uplands had previously been remarked on by Williams (Reference Williams1982a).
8.q.1. Ensigraptus caudatus Subzone
The base of this subzone is defined by the incoming of the index species at Hartfell Score with a large assemblage of other forms (Zalasiewicz, Rushton & Owen, Reference Zalasiewicz, Rushton and Owen1995). The base of this subzone also defines that of the Katian Stage of the Ordovician (Cooper & Sadler, Reference Cooper, Sadler, Gradstein, Ogg and Smith2004). D. clingani is restricted to the caudatus Subzone in both Wales and Scotland (Zalasiewicz, Rushton & Owen, Reference Zalasiewicz, Rushton and Owen1995; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997; Figs 7, 9). In North Wales the caudatus Subzone is present in the Nod Glas Formation (Pratt, Woodhall & Howells, Reference Pratt, Woodhall and Howells1995, p. 49; Young, Gibbons & McCarroll, Reference Young, Gibbons and McCarroll2002, p. 53). In South Wales the assemblage contains similar elements to those found in the Southern Uplands, such as Orthograptus quadrimucronatus and Climacograptus spiniferus. However, E. caudatus itself has been recognized in this region only in the Llanilar–Rhayader area (Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) and doubtfully from Cardigan (Williams et al. Reference Williams, Davies, Waters, Rushton and Wilby2003a). Overall, the caudatus Subzone assemblages of Wales, with about 28 taxa present, are less diverse than those of Scotland, in which about 46 taxa are known.
8.q.2. Dicellograptus morrisi Subzone
In Wales and Scotland, the D. morrisi Subzone has a large and diverse graptolite assemblage, but fewer incoming species than the underlying E. caudatus Subzone (Figs 7, 10). At Hartfell Score, southern Scotland, Zalasiewicz, Rushton & Owen (Reference Zalasiewicz, Rushton and Owen1995) described this interval as a partial-range subzone, the base being defined by the appearance of D. morrisi and the top by the incoming of the linearis Biozone assemblage. At Whitland, South Wales, the base of the subzone was delineated by the incoming of Normalograptus angustus (Perner, 1895) (likely a senior synonym of Climacograptus miserabilis: Loydell, Reference Loydell2007) and possibly D. morrisi, the range of which is limited to this subzone (Zalasiewicz, Rushton & Owen, Reference Zalasiewicz, Rushton and Owen1995). ‘Glyptograptus’ daviesi is the only incoming species definitely reported from the base of the morrisi Subzone in Wales (Fig. 7), but in Scotland several taxa (besides the zone fossil) appear, including Normalograptus mohawkensis, Climacograptus dorotheus, Dicellograptus caduceus and Leptograptus flaccidus spinifer (Fig. 10). Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) did not identify a discrete upper clingani Biozone assemblage in central Wales, but it is present in the Nod Glas of North Wales (Pratt, Woodhall & Howells, Reference Pratt, Woodhall and Howells1995, p. 49) and in coastal sections west of Cardigan, Williams et al. (Reference Williams, Davies, Waters, Rushton and Wilby2003a) found that the morrisi Subzone was better represented than the underlying caudatus Subzone.
8.r. Pleurograptus linearis Biozone
The original work on the graptolite assemblage of this interval was carried out by Lapworth (Reference Lapworth1878) on the upper part of the Lower Hartfell Shales in the Southern Uplands. Williams (Reference Williams1982a) has undertaken the only comprehensive study of the P. linearis Biozone in this area since Elles & Wood's (Reference Elles and Wood1901–1918) review of British graptolites, though Toghill (Reference Toghill1970b) redescribed some of the fauna. Williams (Reference Williams1982a) was able to delineate the base of the biozone for the first time in the North Cliff section of Dob's Linn, and designated it as the type section for the interval base. Taxa diagnostic of the base include the zone fossil, with Climacograptus tubuliferus, possibly Climacograptus styloideus and Leptograptus flaccidus macilentus; however, several forms appear mid-biozone including: Amphigraptus divergens divergens, Leptograptus capillaris and Dicellograptus elegans elegans (Fig. 10). The overall assemblage is quite diverse in Scotland, most species continuing from underlying biozones. Graptolites characteristic of this biostratigraphical interval have been reported in the Rhins of Galloway, southwest Scotland (Stone, Reference Stone1995) and at many sites along-strike to the NE (Floyd, Reference Floyd1999; McMillan, Reference McMillan2002), including Hartfell Score in the Moffat district (Zalasiewicz, Rushton & Owen, Reference Zalasiewicz, Rushton and Owen1995). Floyd (Reference Floyd1999) recorded the linearis Biozone in the Girvan succession.
At Whitland in south Wales, a low-diversity graptolite assemblage, possibly equating (at least in part) with the P. linearis Biozone, was recognized by Zalasiewicz, Rushton & Owen (Reference Zalasiewicz, Rushton and Owen1995), the base of which marks the appearance of large normalograptids. They referred to this biozone as the ‘Normalograptus proliferation interval’. Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) did not record any linearis Biozone assemblages from the Llanilar–Rhayader area, but Williams et al. (Reference Williams, Davies, Waters, Rushton and Wilby2003a) recognized equivalents of the linearis Biozone at Frongoch, southwest of Cardigan, and records of Climacograptus styloideus at Llanystumdwy suggest that the linearis Biozone may be present in North Wales (Harper, Reference Harper1956). Exact correlation between the base of the Ashgill Series as defined in the UK, and the British graptolite biostratigraphical sequence cannot as yet be made, as the Welsh record is poor, and the Lower Ashgill of Scotland is rarely graptolitic (Rushton, Reference Rushton1990). The linearis Biozone currently spans the British Caradoc–Ashgill boundary, as the lower part of the biozone is believed to correlate with the uppermost Caradoc Onnian Substage (see Fig. 1; Ingham, Reference Ingham1966; Ingham & Wright, Reference Ingham and Wright1970). Rickards (Reference Rickards2002) described several species referable to the linearis Biozone from strata as high as the upper Rawtheyan (shelly zone 6) in the typical Ashgill succession of the Cautley district, northern England (see below).
8.s. Dicellograptus complanatus Biozone
The D. complanatus Biozone is another graptolite assemblage biozone initally recognized in the Southern Uplands of Scotland (Dob's Linn) by Lapworth (Reference Lapworth1879–1880a), but rarely recognized since. The work of Williams (Reference Williams1987) on strata representing this interval from the Moffat and Girvan areas has resulted in a greater understanding of the complanatus Biozone assemblage, although the upper and lower limits of the biozone cannot be precisely delineated as the fossiliferous beds lie within strata barren of graptolites. The assemblage is of low diversity and consists mainly of species confined to this level, including the zone fossil and Dicellograptus alector, Orthoretiolites? pulcherrimus, Dicellograptus minor and Glyptograptus occidentalis. Longer-ranging species occur, such as Normalograptus angustus (= Climacograptus miserabilis of older literature) and C. tubuliferus (Fig. 10).
The complanatus Biozone has not been conclusively proven to occur in Wales (Rushton, Reference Rushton1990), and no diagnostic complanatus Biozone species have been recovered. There is no graptolitic evidence for the complanatus Biozone in northern England; indeed there is scarcely space to accommodate it: Rickards (Reference Rickards2002, Reference Rickards2004) placed graptolites from shelly zone 6 (upper Rawtheyan) of the type Ashgill section, Backside Beck, Cautley, northern England (Ingham, Reference Ingham1966; Ingham & Wright, Reference Ingham and Wright1970) in the linearis Biozone and recorded graptolites from shelly zone 7 that could represent the complanatus or the anceps graptolite Biozone. In the Girvan area, however, beds that are correlated with shelly zone 7 have yielded Paraorthograptus pacificus, the index species of the upper subzone of the anceps Biozone (Floyd, Williams & Rushton, Reference Floyd, Williams and Rushton1999). The suggestion by Elles (Reference Elles1925, p. 343) that the complanatus Biozone was ‘a sub-zone of the P. linearis zone. . .’, possibly representing special environmental conditions, might bear renewed examination.
8.t. Dicellograptus anceps Biozone
Lapworth introduced the D. anceps Biozone for the graptolite assemblages of the uppermost Hartfell strata in the Moffat area (Lapworth, Reference Lapworth1878, p. 316). A taxonomic and stratigraphical review of this interval was undertaken by Williams (Reference Williams1982b), who was able to subdivide the biozone into two subzones: a lower Dicellograptus complexus Subzone and an upper Paraorthograptus pacificus Subzone. The base of the D. complexus interval is marked by the incoming of D. complexus and D. anceps. The subzone possesses a relatively small but distinct graptolite assemblage with Dicellograptus minor and Normalograptus angustus (= N. miserabilis) continuing from the previous biozone. New appearances include Anticostia fastigata, Appendispinograptus supernus, Normalograptus normalis and Pleurograptus lui (Fig. 10) while Orthograptus abbreviatus is abundant. The upper part of the complexus Subzone is characterized by the incoming of Amplexograptus latus and Orthoretiograptus denticulatus. The overlying P. pacificus Subzone has a slightly more diverse assemblage, consisting of the same taxa from the underlying complexus Subzone with the addition, at the base, of P. pacificus, Nymphograptus velatus and Plegmatograptus lautus (Fig. 10). Stone (Reference Stone1995) noted anceps Biozone assemblages in southwest Scotland and there are several records along-strike to the NE (McMillan, Reference McMillan2002).
Graptolites of probable D. anceps Biozone age have been recorded in Wales, although no potential for subdivision has been reported (Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). Taxa present include App. supernus, N. normalis and O. abbreviatus (Fig. 7), while D. anceps itself has been recorded at Nant-y-môch (Jones, Reference Jones1909; Cave & Hains, Reference Cave and Hains1986). The anceps Biozone may be present in the north of England, but is not proved.
8.u. Normalograptus extraordinarius Biozone
This biozone, recognized in Scotland but not in England and Wales because of a lack of graptolitic (anoxic) strata, sees the incoming of Normalograptus extraordinarius and Normalograptus? pseudovenustus cf. pseudovenustus. At Dob's Linn this appears in Band E, formerly placed in the anceps Biozone (Williams, Reference Williams, Hughes and Rickards1986), where it overlaps with the latest appearances of taxa such as Dicellograptus anceps, D. ornatus, Appendispinograptus supernus and Paraorthograptus pacificus. Above this, a very thin black bed, between the highest bed of the anceps Biozone and the base of the persculptus Biozone, was termed the Climacograptus? extraordinarius Band (Ingham, Reference Ingham, Bassett, Cocks, Holland, Ingham, Lawson, Rickards and Temple1979; Williams, Reference Williams1983, Reference Williams, Hughes and Rickards1986, Reference Williams1988); the graptolite assemblage in this is small, comprising only the zonal fossil with Climacograptus sp. indet. and Glyptograptus? sp. indet.
8.v. Normalograptus persculptus Biozone
This biozonal assemblage was originally subdivided from the lower part of the acuminatus Biozone by Jones (Reference Jones1909) at Pont Erwyd, central Wales. The interval has since been recognized throughout the UK (Davies, Reference Davies1929; Toghill, Reference Toghill1968a; Rickards, Reference Rickards1970, Reference Rickards1976; Hutt, Reference Hutt1974–1975; Williams, Reference Williams1988; Stone, Reference Stone1995, p. 11; McMillan, Reference McMillan2002, p. 24), where it equates to the local Hirnantian. The persculptus Biozone is now accepted as the highest graptolite assemblage biozone of the Ordovician in the UK biostratigraphical scheme. Indeed, work on the synonomy of N. persculptus and ‘Glyptograptus’ bohemicus led Štorch & Loydell (Reference Štorch and Loydell1996) to recommend that the base of the persculptus Biozone could be further extended to include an extraordinarius Subzone. The base of the interval is taken at the appearance of N. persculptus; other incoming species include Glyptograptus avitus, Atavograptus ceryx, N. parvulus, N. medius and ‘Climacograptus’ tuberculatus (Figs 7, 10).
9. Silurian
9.a. Akidograptus ascensus–Parakidograptus acuminatus Biozone
This is the basal Silurian graptolite biozone of the British succession. Charles Lapworth (Reference Lapworth1878, p. 318) originally recognized it as such, and named it the Diplograptus acuminatus Zone, although Lapworth's (and Elles & Wood's Reference Elles and Wood1901–1918) concept of this biozone embraced also the underlying persculptus Biozone. It is the lowest of six biozones that Lapworth based on the Birkhill Shale Formation succession at Dob's Linn in southern Scotland.
The base of the biozone is marked by the incoming of Akidograptus ascensus and Parakidograptus acuminatus praematurus and, 1.6 m higher, of Parakidograptus acuminatus acuminatus (Melchin, Cooper & Sadler, Reference Melchin, Cooper, Sadler, Gradstein, Ogg and Smith2004). The assemblage as a whole is dominated by biserial graptolites (Fig. 11). Some of the commonest taxa (Normalograptus normalis, N. medius, N. angustus) range up from the persculptus Biozone, and these are joined by newcomers such as Neodiplograptus modestus s.l. and Ne. diminutus. Records of typically Ordovician taxa, most notably those of orthograptids of the truncatus group (Hutt, Reference Hutt1974–1975; Rickards, Reference Rickards1976) have not been substantiated by later work.
The only monograptid known is Atavograptus ceryx, reported as coming up from the persculptus Biozone and rarely recorded (in Scotland) from this interval (Hutt, Reference Hutt1974–1975; Rickards, Reference Rickards1976), though reported by Harper & Williams (Reference Harper and Williams2002) as appearing in the ascensus Biozone. Normalograptus persculptus has been recorded into the upper part of the biozone (Davies, Reference Davies1929; Zalasiewicz & Tunnicliff, Reference Zalasiewicz and Tunnicliff1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997; but see Loydell, Reference Loydell2007).
Informal subdivision of the ascensus–acuminatus Biozone has been made (Rickards, Reference Rickards1976; Zalasiewicz & Tunnicliff, Reference Zalasiewicz and Tunnicliff1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997), and in some studies a formal distinction has been made (e.g. Rong et al. Reference Rong2007). A. ascensus is generally more abundant in, and P. acuminatus praematurus is confined to, the lower part of the biozone, with P. a. acuminatus appearing higher and being more common in the upper part (Toghill, Reference Toghill1968a; Hutt, Reference Hutt1974–1975; see also Loydell, Reference Loydell2007). Atavograptus ceryx has been recorded only in the lower part (though presumably there must be some link between A. ceryx and later atavograptids which appear in the succeeding atavus Biozone). The middle of the biozone has been recognized by the presence of the distinctive short-ranged species Normalograptus trifilis. Although Cystograptus vesiculosus has also been recorded as low as the middle of the biozone (e.g. Rickards, Reference Rickards1976; McMillan, Reference McMillan2002, appendix 1, loc. 48), these records seem to represent C. ancestralis (Štorch, Reference Štorch1996), at least in part (M. Melchin, pers. comm.). The first record of N. rectangularis is in the upper part of the biozone.
9.b. Atavograptus atavus Biozone
Herbert Lapworth (Reference Lapworth1900) originally referred to this unit as the Monograptus tenuis ‘Zone’, from the Rhayader District of Central Wales. He recorded the assemblage of ‘M. tenuis’, Climacograptus scalaris normalis and C. rectangularis. However, Lapworth's zonal species had been misidentified, and Jones (Reference Jones1909) renamed the zone the Monograptus atavus Zone.
From 19 taxa recorded in the ascensus–acuminatus Biozone, the number in the atavus Biozone increases to approximately 30 (Fig. 11). The essential feature of this biozone is the radiation of monograptid and dimorphograptid taxa. The incoming of Atavograptus atavus defines the base of the biozone, and A. gracilis and Huttagraptus? praematurus appear at more or less the same level. Pribylograptus cf. incommodus appears higher in the biozone. The dimorphograptids appear to form a useful temporal succession; thus Rhaphidograptus extenuatus and Dimorphograptus elongatus appear at or near the base of the biozone, while later newcomers include Dimorphograptus confertus confertus, Dimorphograptus decussatus decussatus, Dimorphograptus longissimus and Dimorphograptus epilongissimus (see Hutt, Reference Hutt1974–1975); the common and long-ranging Rhaphidograptus toernquisti originates near the top of the biozone. This suggests the possibility of informal subdivision, as noted by Rickards (Reference Rickards1976), a possibility reinforced by the seeming restriction of Huttagraptus? praematurus to the lower part of the biozone (Zalasiewicz, Williams & Akhurst, Reference Zalasiewicz, Williams and Akhurst2003).
Biserial graptolites continue to be important. Most of the common normalograptids from the ascensus–acuminatus Biozone range up into the atavus Biozone and beyond; these include N. normalis, N. angustus, N. medius and N. rectangularis.
Toghill (Reference Toghill1968a) described a vesiculosus Biozone assemblage from Dob's Linn which equates with that of the atavus Biozone recorded elsewhere (Rickards, Reference Rickards1976). Elles & Wood (Reference Elles and Wood1901–1918) recorded a zone of ‘Mesograptus modestus and Orthograptus vesiculosus’ to account for the assemblage between the acuminatus and cyphus biozones, and many of the taxa that they recorded clearly indicate an Atavograptus atavus Biozone assemblage.
9.c. Huttagraptus acinaces Biozone
Jones's (Reference Jones1909) initial description of the Monograptus rheidolensis Biozone assemblage from the Pont Erwyd district, central Wales, was based on the appearance of M. rheidolensis (= Huttagraptus acinaces), M. (= Pribylograptus) sandersoni, Dimorphograptus confertus swanstoni, ‘Climacograptus hughesi’ (= Metaclimacograptus slalom) and Glyptograptus tamariscus. Jones's (Reference Jones1909) original definition of the biozone adequately describes the core acinaces Biozone assemblage. Many of the earlier Rhuddanian normalograptids and dimorphograptids range up into the biozone (Figs 11, 12).
Although Toghill (Reference Toghill1968a) recorded H. acinaces and G. tamariscus s.l. from his vesiculosus Biozone (= atavus Biozone), both Rickards (Reference Rickards1970) and Hutt (Reference Hutt1974–1975) placed these as key taxa defining the base of the acinaces Biozone, along with ‘M. hughesi’ (= M. slalom). Rickards (Reference Rickards1976) correlated the upper part of Toghill's vesiculosus Biozone with the acinaces Biozone, and suggested an origin for acinaces in the late atavus Biozone, though this has not been substantiated by further work (e.g. Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
The biozone as a whole is less easy to recognize than the atavus Biozone below or the revolutus Biozone above. Subdivision of this interval is not easy, but Pribylograptus incommodus, Huttagraptus strachani, Pristiograptus fragilis pristinus and Pseudorthograptus mutabilis (and questionably Coronograptus gregarius) seem to appear in the upper part of the biozone. In Central Wales (Zalasiewicz & Tunnicliff, Reference Zalasiewicz and Tunnicliff1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997), the zone fossil and associated normalograptids represent the lower part of the biozone, and a more diverse assemblage occurs in the upper part, with the incoming of several species, including Metaclimacograptus slalom and Glyptograptus tamariscus tamariscus, that are commonly recorded at stratigraphically lower levels in other areas.
Toghill (Reference Toghill1968a) did not recognize an acinaces assemblage in the Birkhill Shale at Dob's Linn; the acinaces Biozone appears to be represented by the lower part of his cyphus Biozone (see discussion in Hutt, Reference Hutt1974–1975; Rickards, Reference Rickards1976), as well as part of his underlying vesiculosus Biozone. The acinaces Biozone and the sub- and superjacent biozones are recognized in the Birkhill Shales southwest of Dob's Linn to the Rhins of Galloway (Stone, Reference Stone1995; McMillan, Reference McMillan2002).
9.d. Monograptus revolutus Biozone
The biozone traditionally placed at this level is that of Coronograptus cyphus (e.g. Elles & Wood, Reference Elles and Wood1901–1918; Rickards, Reference Rickards1970; Hutt, Reference Hutt1974–1975; Rickards, Reference Rickards1976). This latter assemblage, originally recognized by H. Lapworth (Reference Lapworth1900) from the Wye Valley sequence of Rhayader, consisted of Coronograptus cyphus cyphus, Monograptus revolutus?, ‘Monograptus’ attenuatus, Pribylograptus sandersoni?, Normalograptus rectangularis and N. normalis.
As C. c. cyphus has been recorded in assemblages broadly referable to the underlying acinaces Biozone (e.g. Zalasiewicz & Tunnicliff, Reference Zalasiewicz and Tunnicliff1994; see also Rickards, Reference Rickards1976), its appearance cannot define the base of the cyphus Biozone without altering the concept of the overall assemblage and also attenuating the underlying acinaces Biozone.
M. revolutus and C. gregarius gregarius appear in the middle of Toghill's (1968a) cyphus Biozone at Dob's Linn. This, combined with an increase in numbers of C. cyphus cyphus at about the same level, suggested to Rickards (Reference Rickards1976) and Hutt (Reference Hutt1974–1975) that only the upper part of Toghill's biozone represents the cyphus Biozone recognized in other parts of Britain. Elles & Wood (Reference Elles and Wood1901–1918) included a cyphus Biozone in their zonal scheme, containing C. c. cyphus and C. g. gregarius. Both these forms have been recorded from the underlying acinaces Biozone (Rickards, Reference Rickards1976; Zalasiewicz & Tunnicliff, Reference Zalasiewicz and Tunnicliff1994), and the appearance of Rhaphidograptus toernquisti, Glyptograptus tamariscus tamariscus, Huttagraptus acinaces and Pribylograptus incommodus in their cyphus Biozone suggests that at least the lower part of it is equivalent to the acinaces Biozone.
Given this rather complicated state of affairs, we consider that the revolutus/austerus group of graptolites provides a more useful delineation, and their appearance at the base of the biozone is noted by most authors (H. Lapworth, Reference Lapworth1900; Jones, Reference Jones1909; Elles & Wood, Reference Elles and Wood1901–1918; Rickards, Reference Rickards1970, Reference Rickards1976; Hutt, Reference Hutt1974–1975; Baker, Reference Baker1981; Zalasiewicz & Tunnicliff, Reference Zalasiewicz and Tunnicliff1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). We therefore propose renaming the zone after M. revolutus, following Bjerreskov (Reference Bjerreskov1975).
Other newly appearing species are Coronograptus gregarius gregarius (although this species possibly originates in the upper acinaces Biozone), Pribylograptus argutus argutus and Metaclimacograptus undulatus (Fig. 12). Some taxa continue from the previous biozone, notably Atavograptus atavus, Huttagraptus acinaces, Normalograptus medius, N. rectangularis, Rhaphidograptus toernquisti, cf. Pseudorthograptus mutabilis, Cystograptus vesiculosus and Metaclimacograptus slalom, but they are mostly in decline and are not recorded from all areas.
A distinct middle to upper part to this biozone can sometimes be recognized, mainly based on the appearance of several glyptograptids, such as G. tamariscus distans, G. t. varians, G. t. linearis and G. t. angulatus. A probable forerunner to the triangulate monograptids, M. difformis, also appears midway through the biozone.
9.e. Monograptus triangulatus Biozone
The Monograptus triangulatus Biozone represents the lowest part of a broad interval formerly known as the Monograptus gregarius Zone (Lapworth, Reference Lapworth1878; Elles & Wood, Reference Elles and Wood1901–1918). The gregarius Biozone was still used in the Britain until the 1960s (Toghill, Reference Toghill1968a). Elles & Wood (Reference Elles and Wood1901–1918) divided the zone into three subzones. The lowest of these, the fimbriatus Subzone, equates with the current triangulatus Biozone. Toghill (Reference Toghill1968a) described the distribution of graptolites in the Birkhill Shales at Dob's Linn, and horizons 33–31 of his gregarius Biozone contain a typical triangulatus zonal assemblage. Sudbury's (Reference Sudbury1958) detailed work on triangulate monograptids from the Rheidol Gorge in central Wales contributed greatly to the concept of the existing triangulatus Biozone. Despite using the gregarius Biozone herself, she also acknowledged a possible three-fold subdivision based on her reassessment of the morphologies and ranges of the triangulate monograptids as a stratigraphically important group. This enabled Rickards (Reference Rickards1976) to outline the distinctive assemblages comprising the triangulatus, magnus and leptotheca biozones.
The base of the triangulatus Biozone is marked by the appearance of several triangulate monograptids including Monograptus triangulatus triangulatus, M. t. separatus and M. t. predecipiens. Other new appearances include Pristiograptus concinnus. Diversity is high at this stratigraphical level, with many taxa continuing from the previous biozone in addition to the new appearances. Some 60 taxa are currently recognized (Figs 12, 13).
Locally, biostratigraphically barren strata are found at the base of the biozone. For example, in the Lake District (Hutt, Reference Hutt1974–1975), central Wales (Zalasiewicz & Tunnicliff, Reference Zalasiewicz and Tunnicliff1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) and at Llanystumdwy, North Wales (Baker, Reference Baker1981), taxa suggesting the middle of the biozone are the first to appear appear above the cyphus Biozone as recognized by those authors (and in this account referred to the revolutus Biozone).
Subdivision of the triangulatus Biozone is possible: the middle to upper parts of the biozone are marked by the incoming of distinctive taxa such as M. triangulatus major, M. t. fimbriatus, Petalolithus minor, Rastrites longispinus, Glyptograptus tamariscus acutus, Campograptus communis communis and Ca. c. rostratus.
9.f. Neodiplograptus magnus Biozone
Jones (Reference Jones1909) originally recognized a ‘magnus band’ from the Rheidol Gorge section. Later studies revealed a magnus Biozone assemblage from the Machynlleth area, Wales (Jones & Pugh, Reference Jones and Pugh1916). The biozone is defined by the appearance of Neodiplograptus magnus. Other new appearances include Pseudoglyptograptus vas (Hutt, Reference Hutt1974–1975), P. barriei, Monograptus chrysalis and Neodiplograptus peggyae. Monograptus triangulatus fimbriatus (that may be a junior synonym of Monograptus pectinatus Richter: Bjerreskov, Reference Bjerreskov1975), M. t. major and M. pseudoplanus are common associates, while M. t. triangulatus may persist into the lower part of the biozone (Baker, Reference Baker1981) (Fig. 13). This biozone has also been recognized in Scotland (Toghill, Reference Toghill1968a).
Many taxa continue from previous biozones. The upper part of the magnus Biozone is poorly represented in Britain, commonly being represented by barren (bioturbated) strata (e.g. Baker, Reference Baker1981; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
9.g. Pribylograptus leptotheca Biozone
Marr & Nicholson (Reference Marr and Nicholson1888) first identified this biozone in the Skelgill Beds of northern England, where it was named the Monograptus argenteus Zone. Elles & Wood (1901–1918) termed it the M. argenteus Subzone of the gregarius Zone, and recorded the incoming of Pribylograptus leptotheca, Monograptus argenteus, Pristiograptus regularis and P. jaculum at this level (although their overall list of species appearances contains a mixture of forms recorded from both higher and lower levels by other authors). Later, Jones & Pugh (Reference Jones and Pugh1916) recorded a similar assemblage from the same stratigraphical interval at Machynlleth in Wales, and they referred to it as the leptotheca Biozone; this has been more widely adopted as the name for the biozone.
Both Monograptus argenteus and Pribylograptus leptotheca have been recorded in Scotland, northern England and Wales (e.g. Hutt, Reference Hutt1974–1975; Baker, Reference Baker1981; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997), and although there are variations in relative abundance, the presence of either may characterize the base of this biozone (Fig. 13). Other newly appearing taxa recorded include Pristiograptus regularis (see Toghill, Reference Toghill1968a), and Pristiograptus jaculum, Glyptograptus serratus and Monograptus cerastus. Rickards (Reference Rickards1976) suggested that Pr. leptotheca originated in the magnus Biozone, although this has not been substantiated by further work (e.g. Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). Hutt (Reference Hutt1974–1975) and Zalasiewicz (Reference Zalasiewicz1992b) identified the incoming of the monoclimacid thecal morphology (e.g. M. imago) at this level in Britain.
A major feature of the leptotheca Biozone is the marked increase in monograptids with hooked thecae, which include Campograptus millepeda and Ca. lobiferus. Several species continue from the previous biozone, and Coronograptus gregarius gregarius was singled out by Hutt (Reference Hutt1974–1975) as being very common.
In many regions, for example, central Wales (Zalasiewicz, Reference Zalasiewicz1990; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997), leptotheca Biozone assemblages occur in thin fossiliferous levels within a generally barren sequence. This makes subdivision of this interval difficult in Britain, although more continuously fossiliferous sequences are found outside Britain (e.g. Štorch, Reference Štorch1998).
9.h. Lituigraptus convolutus Biozone
This biozone was initially described at Skelgill by Marr & Nicholson (Reference Marr and Nicholson1888). At Dob's Linn, this biozone represents Lapworth's (Reference Lapworth1878) original clingani band (lower part of convolutus Biozone) and cometa Zone (upper part of convolutus Biozone).
The broad features of the biozone have been agreed upon by subsequent authors (Toghill, Reference Toghill1968a; Hutt, Reference Hutt1974–1975; Rickards, Reference Rickards1976; Baker, Reference Baker1981). A number of taxa appear at or around the base of the biozone, including L. convolutus itself, Campograptus clingani, Monograptus limatulus, Torquigraptus? decipiens, Cephalograptus cometa cometa, C. tubulariformis, Glyptograptus incertus s.l. and Torquigraptus urceolinus (which Štorch, Reference Štorch1998, suggested might be a junior synonym of M. decipiens) (Figs 13, 14). Other distinctive and useful forms characterizing this biozone include Rastrites spina of Rickards, Reference Rickards1970, Paradiversograptus capillaris, Monoclimacis crenularis and Rastrites peregrinus. Normalograptus scalaris, Metaclimacograptus hughesi, Pristiograptus jaculum and Campograptus lobiferus have been commonly reported, though they extend beyond the biozone. A probable first occurrence of Streptograptus has been noted at this level (Zalasiewicz, Reference Zalasiewicz1996).
Subdivision of this biozone seems achievable, especially given the faunal diversity at this level, but is hampered by the dominance of bioturbated, non-graptolitic strata, particularly at low stratigraphical levels in England and Wales. Several workers (Toghill, Reference Toghill1968a; Hutt, Reference Hutt1974–1975; Rickards, Reference Rickards1976; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) record the incoming of Cephalograptus cometa extrema as a reliable indicator of the upper part of the convolutus Biozone, and the usefulness of the cephalograptid lineage was further emphasized by Štorch (Reference Štorch1998) in his thorough review of the convolutus Biozone in Bohemia. Zalasiewicz (Reference Zalasiewicz1996; see also Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) identified a possible lower subdivision of the convolutus Biozone that contains a narrow form of L. convolutus (= Lituigraptus richteri of Štorch, Reference Štorch1998). At least part of this division might now, however, be considered to represent the upper part of the leptotheca Biozone (cf. Štorch, Reference Štorch1998).
9.i. Stimulograptus sedgwickii Biozone
This biozonal assemblage was originally identified from the upper Birkhill Shale Formation of Dob's Linn by Lapworth (Reference Lapworth1878) and he named it the M. spinigerus Biozone (it is now named after the senior synonym St. sedgwickii). He recorded a diverse fauna, and distinguished an upper and lower subdivision within the biozone.
The base is defined by the appearance of Stimulograptus sedgwickii, which appears together with Neolagarograptus tenuis (Hutt, Reference Hutt1974–1975). Suggestions of a slightly earlier origin for St. sedgwickii (Rickards, Reference Rickards1976) have not been substantiated by further work (Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). Other incoming taxa include Glyptograptus sinuatus crateriformis, Pribylograptus argutus sequens, Glyptograptus packhami and Parapetalolithus kurcki (Rickards, Reference Rickards1970, Reference Rickards1976; Hutt, Reference Hutt1974–1975), together with Torquigraptus linterni and questionably Rastrites gracilis (Williams et al. Reference Williams, Zalasiewicz, Rushton, Loydell and Barnes2003b). Hutt (Reference Hutt1974–1975) also referred to the characteristic abundance of Glyptograptus incertus at the base of the biozone (see Loydell, Reference Loydell1992–1993a for discussion), while Oktavites contortus, Streptograptus ansulosus, Torquigraptus involutus and Metaclimacograptus undulatus are common in, though not confined to, this biozone (Fig. 14).
L. tenuis seems to be restricted to the lower part of the biozone (Rickards, Reference Rickards1976; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) and thus appears to form a useful basis for subdivision (cf. Štorch, Reference Štorch2001; Štorch & Massa, Reference Štorch and Massa2006; Melchin, Reference Melchin2007). At Dob's Linn this species shows marked stratigraphical variations in abundance within its range (Pannell, Clarkson & Zalasiewicz, Reference Pannell, Clarkson and Zalasiewicz2006).
9.j. Stimulograptus halli Biozone
Jones & Pugh (Reference Jones and Pugh1916) first recognized the Stimulograptus halli Biozone in the Machynlleth–Llyfnant area of central Wales, as an upper part of the sedgwickii Biozone. Rickards (Reference Rickards1976) considered that the halli and sedgwickii Biozone assemblages were not sufficiently distinctive to be separately recognizable and, owing to difficulties in distinguishing between Stimulograptus sedgwickii and S. halli, recommended that the halli Biozone be removed from the UK biostratigraphical scheme. Subsequently, however, Loydell (Reference Loydell1991a) was able to differentiate halli Biozone assemblages in the Cwmsymlog Formation of western mid-Wales, and reinstated the biozone. Loydell (Reference Loydell1992–1993a) recorded that species defining the base of the biozone are Stimulograptus halli, Pristiograptus pristinus and Rastrites linnaei (Fig. 14). Other incoming species are Rastrites schaueri, Paradiversograptus capillaris sensu Loydell, ‘Monograptus’ admirabilis and Parapetalolithus praecedens. A number of relatively long-ranging species also occur, including ‘Monograptus’ capis, Torquigraptus involutus, Stimulograptus sedgwickii and Oktavites contortus. Loydell (Reference Loydell1991b) recognized a halli Biozone assemblage similar to that found in Wales in the upper Birkhill Shales in Dob's Linn, southern Scotland.
9.k. Spirograptus guerichi Biozone
This biozone was formerly a part of the turriculatus Biozone, as recorded by Elles & Wood (Reference Elles and Wood1901–1918) and others. Formerly, the lower part of the turriculatus Biozone was separated off as the Rastrites maximus Subzone; this subzone was first postulated at Dob's Linn (Lapworth, Reference Lapworth1878), but Loydell (Reference Loydell1991b) found that R. maximus does not appear to occur there, and the Upper Birkhill Shales at Dob's Linn are now assigned to the halli Biozone (see above). Rickards (Reference Rickards1976) remarked that the maximus Subzone did not occur at the type locality of the turriculatus Biozone at Browgill, in the English Lake District (Marr & Nicholson, Reference Marr and Nicholson1888), but he (Rickards, Reference Rickards1970, Reference Rickards1976) identified the maximus Subzone in the Howgill Fells and considered it to be an important part of the turriculatus Biozone, although the subzone itself was defined only on the presence of R. maximus itself. Toghill (Reference Toghill1968a) described a maximus Subzone assemblage from the upper Birkhill Shales, and Hutt (Reference Hutt1974–1975) recorded one locality containing R. maximus in the Lake District.
Loydell (Reference Loydell1991a,b) did not use the maximus Subzone for the Anglo-Welsh sequence (although he did record the species), and he divided the turriculatus Biozone into six subzones based on his work in western mid-Wales (Fig. 2). This biostratigraphical subdivision was enhanced when the former turriculatus Biozone was divided into two, creating a lower Spirograptus guerichi Biozone, followed by an upper Spirograptus turriculatus Biozone (Loydell, Reference Loydell1992–1993a; Loydell, Štorch & Melchin, Reference Loydell, Štorch and Melchin1993). His six subzones remained in place, with the guerichi–turriculatus biozonal boundary occurring in the middle of the utilis Subzone. Loydell, Štorch & Melchin (Reference Loydell, Štorch and Melchin1993), having observed that Spirograptus turriculatus itself does not appear until the middle of the turriculatus Biozone as formerly understood, defined the guerichi Biozone as ‘the interval from the first appearance of Spirograptus guerichi to that of Spirograptus turriculatus’. Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) employed the new biozones and their subdivisions in the Llanilar–Rhayader district, but noted that not all of Loydell's subzones could be consistently recognized. McMillan (Reference McMillan2002) likewise adopted subdivision into the guerichi and turriculatus biozones. Loydell's (1992–1993a) guerichi and turriculatus biozones and their component subzones are defined by the appearance or relative abundance of their index species.
9.k.1. Paradiversograptus runcinatus Subzone
The base is delineated by the appearance of Paradiversograptus runcinatus and Spirograptus guerichi, with the eponym being abundant (Loydell, Reference Loydell1991a, Reference Loydell1992–1993a). The numbers of P. runcinatus have been enhanced by current sorting, but this does not reduce the usefulness of the subzone, as it can be recognized in Scandinavia (Loydell, Reference Loydell1992–1993a). Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) did not recognize the subzone in the Llanilar–Rhayader district, and assigned some localities to a runcinatus to renaudi interval. ‘Monograptus’ gemmatus, Pristiograptus renaudi, Streptograptus plumosus and Str. pseudoruncinatus appear in this subzone (Fig. 15).
9.k.2. ‘Monograptus’ gemmatus Subzone
Loydell (1991a, Reference Loydell1992–1993a) described a diverse, distinctive assemblage including Streptograptus pseudoruncinatus, Spirograptus guerichi, Rastrites fugax, Glyptograptus fastigatus, G. tamariscus tamariscus and ‘Monograptus’ gemmatus. In addition to these, several species appear at the base of the subzone and are confined to it, for example, the parapetalolithids, Parapetalolithus elongatus, P. globosus and P. regius, with Glyptograptus auritus (Fig. 15). This subzone has been recognized by Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
9.k.3. Pristiograptus renaudi Subzone
This subzone is characterized by abundant Pristiograptus renaudi and Streptograptus strachani (Loydell, Reference Loydell1991a, Reference Loydell1992–1993a), although neither of these taxa originate at the base of this subzone. However, Loydell recorded that Torquigraptus planus and T. cavei first appear here, and they are useful indicators of the renaudi to utilis subzone interval. Other newly appearing species include Monograptus marri and Parapetalolithus hispanicus (Fig. 15). Both Loydell (Reference Loydell1991a) and Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) noted that the transition from the gemmatus Subzone into this subzone has not yet been recognized in a continuous section, and only one faunal assemblage from the Llanilar–Rhayader district has been assigned to this interval (Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
9.l. Spirograptus turriculatus Biozone
In its current, more biostratigraphically restricted usage, the turriculatus Biozone is defined as lying between ‘the first appearance of Spirograptus turriculatus to that of ‘Monograptus’ crispus, or other species indicative of the crispus Biozone’ (Loydell, Reference Loydell1992–1993a). Spirograptus turriculatus is taxonomically distinct from Spirograptus guerichi (Loydell, Štorch & Melchin, Reference Loydell, Štorch and Melchin1993), which characterizes the underlying biozone. As a result, older definitions for the base of the turriculatus biozone (Rickards, Reference Rickards1970, Reference Rickards1976; Hutt, Reference Hutt1974–1975) are no longer applicable, since these now refer to the base of the guerichi Biozone. Loydell's (Reference Loydell1992–1993a) definition of the base of the turriculatus Biozone placed it in the middle of the Stimulograptus utilis Subzone (Fig. 2). One means of avoiding the situation of placing a zonal boundary within a subzone might be to place the base of the turriculatus Biozone at the base of a slightly extended johnsonae Biozone, to be defined by the first appearance, rather than acme, of johnsonae, which coincides with the first appearance of Sp. turriculatus (M. Melchin, pers. comm.); the current definition of the johnsonae Subzone, however, has been found to work well in practice (e.g. Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
9.l.1. Stimulograptus utilis Subzone
Loydell (Reference Loydell1991a) erected this subzone as the fourth subdivision of the old turriculatus Biozone. Subsequently, Loydell (Reference Loydell1992–1993a) divided the utilis Subzone into two parts, marking the boundary between the guerichi and turriculatus biozones. This was based on the incoming of Parapetalolithus tenuis, Spirograptus turriculatus and Streptograptus johnsonae, and an abundance of Monograptus marri and Streptograptus plumosus, in the middle of the subzone (Fig. 15). Several species seem to disappear mid-subzone, including Parapetalolithus kurcki, Rastrites linnaei, Paradiversograptus runcinatus, Parapetalolithus conicus (= Pa. hispanicus) and Pseudostreptograptus williamsi. Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) recorded the utilis Subzone widely in the Llanilar–Rhayader district.
9.l.2. Streptograptus johnsonae Subzone
This subzone is defined on the abundance of Streptograptus johnsonae and common Monograptus bjerreskovae and M. marri (Loydell, Reference Loydell1991a, Reference Loydell1992–1993a), and has been recognized in the Llanilar–Rhayader area (Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). Appearances at the base of the subzone are rare, including only Parapetalolithus schaueri and Pristiograptus schucherti (Loydell, Reference Loydell1992–1993a). As the range charts demonstrate (Fig. 15), a significant proportion of the low-diversity johnsonae Subzone assemblage is composed of relatively long-ranging species.
9.l.3. Torquigraptus proteus Subzone
Torquigraptus proteus appears at the base of this subzone, and its presence and abundance characterizes this interval (Loydell, Reference Loydell1991a, 1992–1993a). As other authors record (Zalasiewicz, Reference Zalasiewicz1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997), appearances also include Monograptus rickardsi (which may be a junior synonym of M. priodon: Loydell, Reference Loydell1992–1993a) and Monoclimacis? galaensis, as well as Pristiograptus huttae (see Loydell, Reference Loydell1992–1993a) and Streptograptus pseudobecki (Zalasiewicz, Reference Zalasiewicz1994; Loydell, Reference Loydell1990). The bulk of this relatively small assemblage (Figs 15, 16) contains taxa from underlying biozones and subzones.
9.l.4. Torquigraptus carnicus Subzone
Zalasiewicz (Reference Zalasiewicz1994) and Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) recognized a further subzone in central Wales, representing the highest part of the turriculatus Biozone. The base of this subzone is characterized by the appearance of T. carnicus, while Streptograptus whitei, S. exiguus and possibly Stimulograptus clintonensis also appear in this interval, as does Monograptus aff. crispus, a narrow precursor to the eponym of the succeeding biozone (Fig. 16). Streptograptus storchi is noted as abundant in the carnicus Subzone (Zalasiewicz, Reference Zalasiewicz1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
9.m. ‘Monograptus’ crispus Biozone
Initially described from the Lake District (Marr & Nicholson, Reference Marr and Nicholson1888), this biozone is defined by the incoming of ‘Monograptus’ crispus (assigned to Streptograptus by Loydell & Maletz, Reference Loydell and Maletz2004), while the distinctive and useful Cochlograptus veles (= M. discus of some authors) appears at about the same level (Fig. 16). In central Wales, the boundary between the crispus Biozone and the underlying turriculatus Biozone was not easily resolved (Zalasiewicz, Reference Zalasiewicz1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997), as both these characteristic species are rare at the base of the biozone. Loydell (Reference Loydell1991a, Reference Loydell1992–1993a) noted also the incoming of Streptograptus? sp. nov. of Howe (M. P. A. Howe, unpub. Ph.D. thesis, Univ. Cambridge, 1982) at or near the base of the biozone. Zalasiewicz (Reference Zalasiewicz1994; see also Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) divided the crispus Biozone into three subzones based on sections in central Wales. As S. crispus does not range into the highest of these either in Britain or elsewhere, the sartorius Subzone is here elevated to biozonal status.
9.m.1. Monoclimacis? galaensis Subzone
Zalasiewicz (Reference Zalasiewicz1994) and Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) recognized this interval as a partial-range subzone, characterized by the overlap of Monoclimacis? galaensis and Torquigraptus carnicus with ‘Monograptus’ crispus and Cochlograptus veles (= Monograptus discus). Stimulograptus clintonensis and Streptograptus exiguus are also common at this level (Fig. 16).
9.m.2. Streptograptus loydelli Subzone
Zalasiewicz (Reference Zalasiewicz1994) described this subzone as a partial-range subzone, defined by the overlap of ‘Monograptus’ crispus and Streptograptus loydelli. He termed it the ‘Monograptus’ crispus Subzone, the name being changed here to reflect the taxon whose incoming defines its base. ‘Monograptus’ crispus is abundant in this subzone, disappearing at its top (Fig. 16). This subzone is well exposed in the Cwm Ystwyth section in central Wales (Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
9.n. Streptograptus sartorius Biozone
The base of this biozone is represented by the incoming of Streptograptus sartorius, together with Torquigraptus pragensis pragensis and Torquigraptus pragensis ruzickai?, and the disappearance of ‘Monograptus’ crispus (Zalasiewicz, Reference Zalasiewicz1994) (Fig. 16). Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) also recorded this biozone at Cwm Ystwyth and described the interval as akin to an interregnum, with low faunal diversity.
9.o. Monoclimacis griestoniensis Biozone
First recognized by Wood (Reference Wood1906) in the Talerddig Grits of Trannon, Wales, this biozone has an assemblage characterized by an incoming of monoclimacids. Its base is defined by the incoming of the zone fossil, Monoclimacis griestoniensis. It has been more usually recognized in central Wales by the incoming of Monoclimacis directa (= Monograptus cf. griestoniensis of Elles & Wood: see Zalasiewicz, Loydell & Štorch, Reference Zalasiewicz, Loydell and Štorch1995), which there seemed to appear at about the same level (Zalasiewicz, Reference Zalasiewicz1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997); Wilson's (D. R. Wilson, unpub. Ph.D. thesis, Univ. Birmingham, 1954) work suggests that it might appear at a slightly lower level in the Howgill Fells.
In central Wales the griestoniensis Biozone can be informally subdivided. A lower interval is characterized by abundant Monoclimacis directa, co-occurring with Monograptus pseudocommunis, Streptograptus aff. loydelli of Zalasiewicz, Reference Zalasiewicz1994 and Torquigraptus pergracilis of Zalasiewicz, Reference Zalasiewicz1994 (Fig. 16). Monoclimacis griestoniensis s.s. dominates the upper part of the biozone (Zalasiewicz, Reference Zalasiewicz1990, Reference Zalasiewicz1994). Monograptus priodon, C. veles (= M. discus) and Pristiograptus nudus range through the biozone.
9.p. Monoclimacis crenulata Biozone
Initially referred to as the Monograptus crenulatus Biozone by Wood (Reference Wood1906) in the Trannon area, the biozone was originally defined on the appearance of the zone fossil (= Monoclimacis crenulata), associated with an undistinctive assemblage, generally of low diversity. More recent work in central Wales has suggested that the incoming of Monoclimacis vomerina may be a more useful indicator of the base of this biozone (Zalasiewicz, Reference Zalasiewicz1994; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). The assemblage includes taxa from underlying biozones including C. veles (= M. discus) and M. priodon (Fig. 16). The Monoclimacis crenulata Biozone formerly included all British strata from the top of the Monoclimacis griestoniensis Biozone to the base of the Wenlock Series. Rickards (Reference Rickards1976) noted that this biozone was rarely found abroad, where a more refined biostratigraphical subdivision could be recognized (Bouček, Reference Bouček1953; Bjerreskov, Reference Bjerreskov1975). Since then, work by Loydell & Cave (Reference Loydell and Cave1993, Reference Loydell and Cave1996) has led to the recognition of more biozones within the upper Telychian of Wales, and the current crenulata Biozone now represents only the lowest part the original biozone.
9.q. Oktavites spiralis Biozone
Graptolite assemblages representing the Oktavites spiralis Biozone were recognized relatively recently in the UK by Loydell & Cave (Reference Loydell and Cave1993, Reference Loydell and Cave1996). Initial biostratigraphical work at Buttington Brick Pit, central Wales (Loydell & Cave, Reference Loydell and Cave1993) and subsequent studies in eastern mid-Wales (Loydell & Cave, Reference Loydell and Cave1996) resulted in division of the long-established crenulata Biozone, and allowed direct comparison with the graptolite biozonation of Bohemia (Bouček, Reference Bouček1953) and Scandinavia (Bjerreskov, Reference Bjerreskov1975). The Oktavites spiralis Biozone has a distinctive faunal assemblage, and Loydell & Cave (Reference Loydell and Cave1993, Reference Loydell and Cave1996) recorded the introduction of several taxa through the biozone. These include the zone fossil (previously commonly given a longer range in the UK (e.g. Rickards, Reference Rickards1976), prior to closer scrutiny of the Telychian spiraliform monograptids, many of which are superficially similar) and Monoclimacis hemmanni, Streptograptus anguinus, Monoclimacis geinitzi, Streptograptus nodifer (referred to Awarograptus by Zalasiewicz & Howe, Reference Zalasiewicz and Howe2003), Barrandeograptus bornholmensis and Stimulograptus vesiculosus (Figs 16, 17).
9.r. Cyrtograptus lapworthi Biozone
The Cyrtograptus lapworthi Biozone was first recorded in Britain by Loydell & Cave (Reference Loydell and Cave1996) from the Banwy River section, central Wales. It equates roughly with the upper part of the spiralis Biozone and the Stomatograptus grandis Biozone of the Czech Republic. Loydell & Cave (Reference Loydell and Cave1996) described a relatively diverse and characteristic graptolite assemblage. Several new species appear at the base of the biozone, including the zone fossil (Fig. 17). Some are confined to its lower and middle parts, suggesting a potential for subdivision; these include Monoclimacis sublinnarssoni, Streptograptus speciosus, Streptograptus wimani and Pristiograptus largus. A number of taxa continue from the previous biozone, and a few appear in the middle of the biozone, notably Monoclimacis basilica and Barrandeograptus pulchellus. C. lapworthi itself marks the first appearance of Cyrtograptus in Britain. In the Llanilar–Rhayader district of central Wales, Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) assigned strata overlying the spiralis Biozone to a ‘spiralis/centrifugus interregnum’, the assemblage being dominated by long-ranging graptolites such as the members of the Monoclimacis vomerina and Monograptus priodon groups and Retiolites geinitzianus.
9.s. Cyrtograptus insectus Biozone
The biostratigraphical succession outlined by Loydell & Cave (Reference Loydell and Cave1996) in the Banwy River section included the first record of the Cyrtograptus insectus Biozone in Britain. The biozone contains a number of taxa whose ranges are limited to the lower–middle or middle–upper parts of this interval, indicating the possibility of further biostratigraphical subdivision (Fig. 17). This biozone is notable for the introduction of Mediograptus: Mediograptus flittoni appears at the base, together with Monoclimacis shottoni, C. insectus and Cyrtograptus sp. of Loydell & Cave; Monoclimacis linnarssoni disappears in the lower part of the biozone. Species defining the middle and upper parts of the biozone in the Banwy section include Monograptus pseudocultellus, Mediograptus morleyae and Mediograptus sp. aff. inconspicuus of Loydell & Cave.
9.t. Cyrtograptus centrifugus Biozone
The Cyrtograptus centrifugus Biozone was first identified in Britain in the Howgill Fells, northern England (Rickards, Reference Rickards1967). Rickards (Reference Rickards1967, Reference Rickards1976) described an assemblage containing C. centrifugus, monoclimacids and monograptids. The base of this biozone has long been equated with the base of the Wenlock Series in the UK, although more recent micropalaeontological evaluation at Hughley Brook suggests that this level actually equates to a horizon within the upper centrifugus or succeeding murchisoni graptolite Biozone (Mullins & Aldridge, Reference Mullins and Aldridge2004). The incoming of the zone fossil defines the biozonal base, although several other species also appear for the first time (Rickards, Reference Rickards1976; Loydell & Cave, Reference Loydell and Cave1996; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997; Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999). These include Mediograptus danbyi, Med. cautleyensis, Monograptus simulatus and Monoclimacis? sp. 1 (sensu Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999) (Fig. 17). Mediograptus inconspicuus and questionably Mediograptus flexuosus appear in the middle to upper part of the biozone. These are accompanied by relatively long-lived taxa ranging from previous zones (Loydell & Cave, Reference Loydell and Cave1996; Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997).
9.u. Cyrtograptus murchisoni Biozone
In many areas of Britain, the murchisoni Biozone assemblage is often indistinguishable from the underlying centrifugus Biozone where the eponym is absent (Rickards, Reference Rickards1967, Reference Rickards1976; Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999; Davies et al. Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997). The biozone was initially recognized by Lapworth (Reference Lapworth1879–1880) from Builth Wells, and contains the index species together with Monograptus priodon and Monoclimacis vomerina. In central Wales, Loydell & Cave (Reference Loydell and Cave1996) and Zalasiewicz & Williams (Reference Zalasiewicz and Williams1999) have recognized taxa such as Monoclimacis vikensis?, Mcl. adunca, Pristiograptus praedubius, Monograptus radotinensis radotinensis, Cyrtograptus bohemicus and Pseudoplegmatograptus? wenlockianus? (Fig. 17). Loydell & Cave (Reference Loydell and Cave1996) noted that species diversity drops markedly in the upper part of the biozone, indicating a potential for future subdivision.
9.v. Monograptus firmus Biozone
The rapid decline in diversity in the upper part of the murchisoni Biozone, coupled with the incoming of M. firmus in the Banwy River section, central Wales, resulted in Loydell & Cave (Reference Loydell and Cave1996) establishing the firmus Biozone in Britain for the first time. Approximately seven species, most of which have long biostratigraphical ranges, are found at this level; these include Monograptus priodon, Monoclimacis vomerina and possibly M. basilica (Fig. 17). Loydell & Cave (Reference Loydell and Cave1996) identified the long-ranging Pristiograptus dubius as appearing in the biozone. Zalasiewicz & Williams (Reference Zalasiewicz and Williams1999) reported the appearance of M. firmus s.l. from strata which they assigned to the upper part of the murchisoni Biozone at Builth Wells in Wales, but they did not recognize a discrete firmus Biozone.
9.w. Monograptus riccartonensis Biozone
The riccartonensis Biozone was first recognized in the UK by Elles (Reference Elles1900) at Builth Wells, south-central Wales. It has since been recognized in many areas of Britain, and notable studies include Rickards's (Reference Rickards1967, 1969) work in northern England, that of Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) in the Llanilar–Rhayader district of central Wales, and Zalasiewicz & Williams's (Reference Zalasiewicz and Williams1999) re-examination of the sequence at Builth Wells. The base of the biozone is generally taken at the incoming of Monograptus riccartonensis, the stratigraphical range of which defines the biozone (Fig. 17); this taxon dominates most assemblages.
Several authors have noted stratigraphical variations in specific composition and diversity in this generally impoverished interval, and this might form the basis of future two- or three-fold subdivision (Rickards, Reference Rickards1967, Reference Rickards1976; White et al. Reference White, Barron, Barnes and Lintern1992; Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999). The middle to upper parts of this biozone are generally more diverse, and include Pristiograptus dubius s.l., Monograptus firmus s.l., M. radotinensis inclinatus and the first appearance of Monograptus antennularius and possibly of Monoclimacis flumendosae.
9.x. Note on middle Wenlock graptolite biostratigraphy
The interval from the upper part of the riccartonensis Biozone through to the lundgreni Biozone is problematical in the British Isles, and a number of biozonal schemes have been proposed. This account (Fig. 2) uses the most recent of these (Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999), which is based upon a re-examination of the historically important (Elles, Reference Elles1900; Jones, Reference Jones1947) Builth district of south-central Wales. The problems, and suggested correlations with earlier schemes and those used elsewhere in the world, are outlined in that paper. The most important difference from widely used earlier biozonations (e.g. Rickards, Reference Rickards1976) is that separate flexilis and ellesae biozones are not recognized, because Monograptus flexilis appears earlier than, and then largely co-exists with, Cyrtograptus rigidus at Builth, while C. ellesae appears later than C. lundgreni (Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999; Williams & Zalasiewicz, Reference Williams and Zalasiewicz2004). The biostratigraphical problems in part reflect imperfect taxonomy and in part originate from, and continue to be exacerbated by, biofacies control. Most UK middle Wenlock sequences (including Builth) are dominated by a few morphologically variable and long-ranging monograptid, monoclimacid and pristiograptid taxa, while the biostratigraphically useful cyrtograptids are rare in comparison to successions such as those of Bohemia (e.g. Štorch, Reference Štorch1994).
9.y. Pristiograptus dubius Biozone
The dubius Biozone was originally defined in the Czech Republic (Bouček, Reference Bouček1960; also Štorch, Reference Štorch1994), and Zalasiewicz & Williams (Reference Zalasiewicz and Williams1999) recognized the biozone in the UK for the first time at Builth Wells. This interval is an interregnum, with recognition largely reliant on identification of the biozones directly above and below. The biozone is typified by variable, long-lived taxa of the Pristiograptus dubius group, as well as by Monograptus priodon/flemingii and Monoclimacis flumendosae (Figs 17, 18). Rare first occurrences recorded at Builth include Mediograptus ex. gr. retroflexus and M. flexilis, the latter first being found midway up the biozone. The dubius Biozone is approximately equivalent to the antennularius Biozone of Rickards (Reference Rickards1965, Reference Rickards1967, Reference Rickards1969) and White et al. (Reference White, Barron, Barnes and Lintern1992).
9.z. Cyrtograptus rigidus Biozone
The rigidus Biozone was originally defined at Builth by Elles (Reference Elles1900) as her symmetricus Zone. Her species C. symmetricus was, however, a misidentification of C. rigidus, and the name of the biozone was subsequently changed. The base of the rigidus Biozone is defined by the appearance of the index fossil, which is limited to the biozone. The rigidus Biozone, as defined herein, extends to the base of the lundgreni Biozone and therefore equates with Elles's (1900) linnarssoni Zone (= flexilis Biozone: Rickards, Reference Rickards1976) as well as her symmetricus (= rigidus) Zone (see Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999).
Other elements of the assemblage include taxa ranging up from the dubius Biozone such as Monograptus flexilis, M. priodon/flemingii, Mediograptus ex. gr. retroflexus, Monoclimacis flumendosae and graptolites of the Pristiograptus dubius group including the distinctively broad P. pseudolatus (Fig. 18).
At Builth (Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999), this unit could not be subdivided, because (1) Monograptus flexilis, one index species of the formerly recognized flexilis/linnarssoni Biozone, in fact appears below C. rigidus, and then has a range largely that overlaps with it; (2) the other index species, Cyrtograptus linnarssoni, seems to be based on an unusually narrow and/or deformed example of C. rigidus s.l.; this taxon has accordingly been widely misidentified, most attributed specimens being referable to C. rigidus; and (3) temporal subspecies of C. rigidus could not be distinguished at Builth. C. rigidus cautleyensis Rickards, Reference Rickards1967 has been interpreted as a late, slender subspecies indicative of the flexilis/linnarssoni Biozone (e.g. Rickards, Reference Rickards1976). The type material of this taxon from the Howgill Fells does seem to be a recognizable slender morphotype (Williams & Zalasiewicz, Reference Williams and Zalasiewicz2004), but it could not be recognized as distinct at Builth; on present evidence it may be a true geographical subspecies restricted to northern England. Finally, the remainder of the assemblage, consisting of abundant pristiograptid, monograptid and monoclimacid graptolites that belong to a few variable, long-ranging taxa, are of limited biostratigraphical use.
9.aa. Cyrtograptus lundgreni Biozone
This biozone was first defined at Builth by Elles (Reference Elles1900). The base of the biozone is recognized by the incoming of the index species. However, re-examination of the Builth succession (Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999) has shown that it overlies the rigidus Biozone directly without the intervening linnarssoni or ellesae biozones that Elles had recognized. Zalasiewicz & Williams (Reference Zalasiewicz and Williams1999; see also Williams & Zalasiewicz, Reference Williams and Zalasiewicz2004) found that C. ellesae, the zonal index species of the ellesae Biozone, in fact appears higher than C. lundgreni, and the ranges of C. ellesae and C. lundgreni overlap (see also Warren, Reference Warren1971; Rickards, Reference Rickards1976); thus we do not recognize the ellesae Biozone in this account. This causes problems regarding the links between biostratigraphy and chronostratigraphy, because the boundary between the Sheinwoodian and Homerian stages was placed at the base of the ellesae Biozone as recognized in the Welsh Borders (Bassett, Rickards & Warren, Reference Bassett, Rickards and Warren1975; see discussion in Zalasiewicz & Williams, Reference Zalasiewicz and Williams1999 and Melchin, Cooper & Sadler, Reference Melchin, Cooper, Sadler, Gradstein, Ogg and Smith2004).
There is some potential for finer subdivision. At the base of the biozone, C. lundgreni was found to overlap with the latest occurrences of Mediograptus ex. gr. retroflexus. There might be also a discrete middle to upper biozonal assemblage including Cyrtograptus ramosus, C. hamatus, C. ellesae and Testograptus testis, although its recognition may be hampered in practice because such taxa are rare in comparison with the more long-ranging pristiograptids, monoclimacids and monograptids (Fig. 18). Davies et al. (Reference Davies, Fletcher, Waters, Wilson, Woodhall and Zalasiewicz1997) and Bassett, Rickards & Warren (Reference Bassett, Rickards and Warren1975) recorded this biozone from central Wales, and Warren et al. (Reference Warren, Price, Nutt and Smith1984) used it in north Wales.
9.bb. Gothograptus nassa Biozone
The nassa Biozone was first adopted for use in Britain by Warren (Reference Warren1971) working in north Wales. It succeeds a major graptolite extinction event, with most of the graptolites characteristic of the preceding lundgreni Biozone not surviving into this interval (Rickards, Reference Rickards1976; Jaeger, Reference Jaeger1991). Indeed, only four taxa have been recorded (Fig. 18), one of which is a first appearance. The assemblage consists of Pristiograptus dubius (and its subspecies ludlowiensis) and Gothograptus nassa, which dominate most assemblages, with P. aff. jaegeri of Holland, Rickards & Warren, Reference Holland, Rickards and Warren1969; reliable records of Pristiograptus jaegeri appear in the upper part of the biozone. G. nassa has been reported beyond its biozone in Britain (Fig. 18), although subsequent detailed work in Arctic Canada, Baltica and the Czech Republic (Porębska, Kozłowska-Dawidziuk & Masiak, Reference Porębska, Kozłowska-Dawidziuk and Masiak2004) suggests that it is confined there to its biozone and the overlying praedeubeli Biozone (not recognized in Britain) of that region; this suggests that careful reassessment of UK material of nassa is needed. Zalasiewicz & Williams (Reference Zalasiewicz and Williams1999) referred only to a nassa–ludensis Biozone representing the uppermost interval of Wenlock strata at Builth Wells; both indicator species are rare in this area and their biostratigraphical ranges cannot be clearly constrained. In general, the low abundance and diversity of graptolites in upper Wenlock strata in the British Isles make it difficult to apply the fine upper Wenlock graptolite biostratigraphy recognized elsewhere in the world (e.g. Lenz et al. Reference Lenz, Noble, Masiak, Poulson and Kozłowska2006).
9.cc. Monograptus ludensis Biozone
Originally recognized as the vulgaris Zone by Wood (Reference Wood1900) at Builth and Long Mountain, this biozone was redefined and renamed by Holland, Rickards & Warren (Reference Holland, Rickards and Warren1969; see also Rickards, Reference Rickards1976).
The base of the ludensis Biozone is taken at the incoming of the biozonal species (Fig. 18) which, in the lower part of the interval, co-occurs with taxa from the preceding G. nassa Biozone. This biozonal assemblage indicates the possibility of a more refined biostratigraphic subdivision (see Rickards, Reference Rickards1976), as the first occurrences of other species range from the middle to upper part of the biozone. These include Holoretiolites (Balticograptus) lawsoni, Pristiograptus auctus, Monograptus deubeli and M. aff. uncinatus orbatus. The loss of P. aff. jaegeri of Holland, Rickards & Warren (Reference Holland, Rickards and Warren1969) midway through the biozone reinforces the distinction between a lower and an upper assemblage within the ludensis Biozone.
9.dd. Neodiversograptus nilssoni Biozone
In recent years, much biostratigraphical work has been undertaken on British strata of Ludlow age by the British Geological Survey on Ludlow successions in central Wales (Schofield et al. Reference Schofield, Davies, Waters, Wilby, Williams and Wilson2004; Barclay et al. Reference Barclay, Davies, Humpage, Waters, Wilby, Williams and Wilson2005). Nevertheless, the biozonal criteria for the series have not changed significantly since Rickards's (Reference Rickards1976) review of Silurian graptolite biostratigraphy. The base of the N. nilssoni Biozone coincides only approximately (Melchin, Cooper & Sadler, Reference Melchin, Cooper, Sadler, Gradstein, Ogg and Smith2004) with the base of the Ludlow Series. Many Lower Ludlow taxa had been identified in Britain, notably from the Lake District and Ludlow itself (Nicholson, Reference Nicholson1868; Hopkinson, Reference Hopkinson1873), before Lapworth (Reference Lapworth1879–1880), working in central Wales and the Welsh Borderlands, designated the ‘zone of Monograptus nilssoni’, which included the whole Lower Ludlow. Further studies, most importantly those of Wood (Reference Wood1900), led to refinement and subdivision of this broad biostratigraphical unit (Rickards, Reference Rickards1967, Reference Rickards1969; Warren, Reference Warren1971).
The nilssoni Biozone shows a much greater specific diversity than the preceding Wenlock biozones, with the appearance and diversification of Saetograptus being especially characteristic. The appearance of the zone fossil defines its base, and taxa such as Monograptus uncinatus orbatus, Plectograptus macilentus, Spinograptus spinosus, Saetograptus colonus colonus, S. varians varians and Bohemograptus bohemicus s.l. also appear at about this level (Fig. 18).
In north Wales, Warren (in Warren et al. Reference Warren, Price, Nutt and Smith1984, p. 54) divided the nilssoni Biozone into lower and upper divisions, and there is potential for further subdivision (see discussion in Rickards, Reference Rickards1976). There is a distinct lower to middle zonal assemblage, with M. ludensis and P. auctus, which disappears in the mid-nilssoni Biozone. A distinct assemblage appears in the middle to upper part of the biozone and contains many new genera and species of Monograptidae, notably Cucullograptus (Lobograptus) progenitor, C. (L.) scanicus, C. (L.) simplex, Monoclimacis micropoma, Mcl. haupti, Pristiograptus vicinus and Crinitograptus crinitus. The saetograptids diversify rapidly in this upper interval with the incoming of Saetograptus roemeri, S. fritschi fritschi, S. leintwardinensis incipiens, S. wandalensis, S. chimaera chimaera, S. chimaera salweyi and S. chimaera semispinosus. This biostratigraphical refinement has been expressed in Poland and the Czech Republic in terms of a lower nilssoni Biozone succeeded by an upper progenitor Biozone (Urbanek, Reference Urbanek1966; Teller, Reference Teller1969; Přibyl, Reference Přibyl1983); Melchin, Cooper & Sadler (Reference Cooper, Sadler, Gradstein, Ogg and Smith2004) and Koren’ et al. (Reference Koren’, Lenz, Loydell, Melchin, Štorch and Teller1996) indeed used this assemblage (and the incoming of C. (L.) scanicus) to define the base of the scanicus Biozone, and this resolution might ultimately prove optimal also for the British succession.
9.ee. Cucullograptus (Lobograptus) scanicus Biozone
Wood (Reference Wood1900) established the scanicus Biozone in the Builth and Ludlow areas, and, while widely employed, this interval remains problematical in the British Isles. Rickards's (Reference Rickards1976) range chart showed no appearances at the base of this interval, and it was characterized by extinctions of many nilssoni Biozone taxa and an increase in the abundance of Cucullograptus (Lobograptus) scanicus and Saetograptus chimaera chimaera (Wood, Reference Wood1900; Rickards, Reference Rickards1976) (Figs 18, 19). Recent work in the Montgomery district of central Wales (White in Cave & Hains, Reference Cave and Hains2001, p. 69) showed a number of appearances there at this level, including most of the subspecies of S. chimaera. The incoming of Pristiograptus tumescens tumescens, Saetograptus clunensis, Pristiograptus welchae and Bohemograptus bohemicus tenuis was shown in the middle part of the scanicus interval by Rickards (Reference Rickards1976).
9.ff. Saetograptus incipiens or Pristiograptus tumescens Biozone
Wood (Reference Wood1900) established this biozone in Wales and the Welsh Borderland and employed both zonal terms. As with the preceding scanicus Biozone, it is somewhat ill-defined and approximates to an interregnum. The biozonal name varies locally according to the relative abundance and/or the presence or absence of Saetograptus incipiens and Pristiograptus tumescens, both of which originate in earlier biozones (see discussion in Rickards, Reference Rickards1976). Graptolite species diversity is reduced in the incipiens/tumescens Biozone compared with previous Ludlow biozones, with approximately ten confirmed taxa (Figs 18, 19), most of them ranging from the preceding biozones. Only one new form appears: Saetograptus? aff. incipiens (Wood, Reference Wood1900). A number of species have been recorded as disappearing in the lower to middle part of the interval (Rickards, Reference Rickards1976), perhaps indicating the possibility of subdivision; these include Monoclimacis haupti, Pristiograptus vicinus, Saetograptus clunensis and S. varians varians. In practice, though, this interval, sharing so many taxa with the scanicus Biozone, is hard to recognize, particularly in spot localities.
9.gg. Saetograptus leintwardinensis leintwardinensis Biozone
This biozone was initially defined by Marr (Reference Marr1892) in the Lake District and has been widely recognized in the UK (Wood, Reference Wood1900; Holland, Lawson & Walmsley, Reference Holland, Lawson and Walmsley1963; Rickards, Reference Rickards1967), largely on the incoming of S. l. leintwardinensis (Fig. 19). Only two other taxa are confirmed in this biozone, namely Saetograptus? aff. incipiens, which continues from the previous biozone, and Saetograptus leintwardinensis primus, which is confined to the lower–middle part of the biozone (Shergold & Shirley, Reference Shergold and Shirley1968). Such a low-diversity assemblage may reflect a deteriorating environment or a deepening evolutionary crisis amongst British graptolites at this time.
9.hh. Bohemograptus proliferation Biozone
This biozone was originally described in the UK by Holland & Palmer (Reference Holland, Palmer, Rickards, Jackson and Hughes1974), when they identified Bohemograptus bohemicus tenuis from strata overlying the leintwardinensis Biozone in the Welsh Borderlands (see also White in Cave & Hains, Reference Cave and Hains2001). This taxon is the youngest graptolite in the UK biostratigraphical sequence, and the sole representative in this biozone (Fig. 19). B. bohemicus tenuis has not been reported from the immediately underlying leintwardinensis Biozone, despite its origination earlier in the Ludlow, in the scanicus Biozone (Rickards, Reference Rickards1976).
Acknowledgements
The authors wish to thank Mike Melchin and Petr Štorch for their meticulous and careful refereeing, that has considerably improved this text, Lisa Barber and Mateusz Zalasiewicz for help with producing the figures, and Jane Holland and Lori Snyder for their patient and painstaking editorial work.
























