


1. Introduction
The Silurian was long considered to have been a period with a relatively stable greenhouse climate, devoid of significant extinction events (see Holland, Reference Holland1991 and Calner, Reference Calner and Ashraf2008 for summary). This view of an ‘idyllic Silurian’ rapidly changed in the 1990s. Prominent extinction events were described among graptolites (Jaeger, Reference Jaeger1991; Koren’, Reference Koren’1993; Urbanek, Reference Urbanek1993; Loydell, Reference Loydell1994; Štorch, Reference Štorch1995a, Reference Štorchb; Melchin, Koren’ & Štorch, Reference Melchin, Koren’ and Štorch1998) and conodonts (Jeppsson, Reference Jeppsson1990, Reference Jeppsson1998; Jeppsson & Aldridge, Reference Jeppsson and Aldridge2000). Later studies on carbon isotopes revealed that some extinctions correlate approximately with positive carbon isotope excursions and may have resulted from extraordinary perturbations in the carbon-ocean cycle (Munnecke, Samtleben & Bickert, Reference Munnecke, Samtleben and Bickert2003; Lehnert et al. Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b; Calner, Reference Calner and Ashraf2008). In addition, oxygen isotopes suggest dramatic fluctuations in water temperature during the Silurian, indicating recurrent periods with an icehouse climate (Lehnert et al. Reference Lehnert, Eriksson, Calner, Joachimski and Buggisch2007a; Calner, Reference Calner and Ashraf2008; Eriksson & Calner, Reference Calner and Ashraf2008). Various climatic and oceanographic models have been proposed to explain extinctions and changes in chemistry in the Silurian oceans (Jeppsson, Reference Jeppsson1990; Bickert et al. Reference Bickert, Pätzold, Samtleben and Munnecke1997; Samtleben, Munnecke & Bickert, Reference Samtleben, Munnecke and Bickert2000). Nevertheless, there is as yet no generally accepted explanation (see discussion in Loydell, Jeppsson & Aldridge, Reference Loydell, Jeppsson and Aldridge2001 and Loydell, Reference Loydell2007). Particular debates have been devoted to high-resolution correlation. The correlation of graptolite extinctions in deeper water shale facies with those of conodonts in shallow water limestone facies, combined with correlation between different palaeogeographical settings, i.e. higher and lower latitudes, is a difficult exercise. The timing, extent and selectivity of extinction in benthic groups are still poorly known (Talent et al. Reference Talent, Mawson, Andrew, Hamilton and Whitford1993; Kříž, Reference Kříž1998, Reference Kříž, Boucot and Lawson1999a, Reference Kříž2010a, Reference Křížb; Manda & Kříž, Reference Manda and Kříž2006; Eriksson, Nilsson & Jeppsson, Reference Eriksson, Nilsson and Jeppsson2009). The question has been posed (Loydell, Jeppsson & Aldridge, Reference Loydell, Jeppsson and Aldridge2001; Manda & Kříž, Reference Manda and Kříž2006; Loydell, Reference Loydell2007; Manda & Frýda, Reference Manda and Frýda2010): are extinctions in graptolites, conodonts and benthos coincident or do they represent separate events? Loydell (Reference Loydell2007) presented evidence to indicate that the Wenlock graptolite and conodont extinction events are not coincident, but for some other events, so far no adequate evidence has been provided.
The middle Ludfordian change in marine faunas is an example of such an extinction event. The Kozlowskii Event is considered to be one of the most significant graptolite extinction events, and has been recorded from Baltica, central-Asian terranes and peri-Gondwanan Bohemia (Koren’, Reference Koren’1993; Urbanek, Reference Urbanek1993; Štorch, Reference Štorch1995a, Reference Štorchb; Manda & Kříž, Reference Manda and Kříž2006). A conodont extinction event, known as the Lau Event in Baltica, Laurentia and East Gondwana (Australia), is roughly coincident with the Kozlowskii Event, but detailed comparison is lacking (Jeppsson, Reference Jeppsson1990; Eriksson & Calner, Reference Eriksson and Calner2008). A positive carbon isotope excursion, the largest in the Phanerozoic, has been identified close to both the Lau conodont extinction Event (Samtleben, Munnecke & Bickert, Reference Samtleben, Munnecke and Bickert2000; Calner & Eriksson, Reference Calner and Eriksson2006; Stricanne, Munnecke & Pross, Reference Stricanne, Munnecke and Pross2006; Kaljo et al. Reference Kaljo, Grytsenko, Martma and Mõtus2007) and the Kozlowskii graptolite extinction (Lehnert et al. Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b). In Bohemia, the Kozlowskii Event is associated with prominent extinctions among benthic faunas (Manda & Kříž, Reference Manda and Kříž2006). Oxygen isotopes suggest progressive cooling accompanying an icehouse period around the time of the extinction (for details see Lehnert et al. Reference Lehnert, Eriksson, Calner, Joachimski and Buggisch2007a).
The Prague Synform (i.e. Barrandian area) of central Bohemia has long been considered a classic area for Silurian graptolite biostratigraphy and a reference area for correlation of the Early Palaeozoic of northwestern peri-Gondwana. Many graptolite biozones and index species were first distinguished and described from this area. The Llandovery, Wenlock and lower Ludlow sedimentary rock sequence, composed largely of graptolite-bearing shales, provides an ideal opportunity for detailed study of the graptolite succession (Bouček, Reference Bouček1953; Přibyl, Reference Přibyl1983; Štorch, Reference Štorch1994, Reference Štorch2006 and further references therein). Similarly, the Pridoli succession, composed of limestones interbedded with shales, has yielded graptolite assemblages, permitting its division based upon graptolites (Přibyl, Reference Přibyl1940; Kříž et al. Reference Kříž, Jaeger, Paris and Schönlaub1986). In the limestone-dominated upper Ludlow strata of the Prague Synform, however, the graptolite biostratigraphy remained of rather poor resolution. The graptolite biozones established by Bouček (Reference Bouček1936) and revised by Přibyl (Reference Přibyl1983) were defined inadequately and, in particular, there were no published descriptions of sections that had been measured and sampled in detail. Comparison between graptolite taxa based upon Bohemian type material preserved as internal moulds in skeletal limestones and specimens flattened in shales was inadequate and resulted in some confusion in stratigraphic correlation. Some of these graptolite taxa, described by Bouček (Reference Bouček1931, Reference Bouček1936) and Přibyl (Reference Přibyl1943, Reference Přibyl1983), are in need of revision.
The most detailed late Ludlow, Ludfordian, graptolite biozonal scheme was elaborated in Poland (Urbanek & Teller, Reference Urbanek and Teller1997 and references therein) and partly applied in other parts of the Eastern European Platform (such as Podolia in Ukraine, Belarus and Latvia). Koren’ & Sujarkova (Reference Koren’ and Sujarkova1997, Reference Koren’ and Sujarkova2004) introduced a different biozonation in Central Asia. The graptolites collected from Kosov Quarry near Beroun (Štorch, Reference Štorch1995a) and from the present Všeradice section enable correlation with both the Eastern European Platform and Central Asia.
In this paper, the shale-dominated succession comprising the middle Ludfordian Kozlowskii Event and post-extinction recovery is described for the first time both from Bohemia and peri-Gondwana. The Všeradice section was exposed in trenches and measured and sampled bed by bed for graptolites. Isolated limestone beds were sampled for conodonts. The sedimentary succession was examined in detail and the carbon isotope record analysed in order to evaluate the depositional environment of the graptolite-bearing rocks and to recognize possible gaps in sedimentation. Scattered shelly faunas recovered from shales and some limestone beds enabled correlation between abundant benthic faunas of the limestone-dominated facies widespread in other Bohemian sections and the widely applicable graptolite biozones recognized herein.
2. Všeradice section
In the Prague Synform, the middle and upper Ludfordian succession usually comprises carbonate rocks with poorly preserved and rare graptolites (Bouček, Reference Bouček1936; Kříž, Reference Kříž1992). The relatively deeper water, oxygen-depleted hemipelagic shale facies, potentially rich in diverse graptolites, is restricted to the southwestern closure and southeastern flank of the Prague Synform. There are only a few poorly exposed sections in the shale-dominated area, usually insufficient for detailed investigations (Horný, Reference Horný1955, Reference Horný1960). The single exception is the section along the field track near Všeradice village. This area is regarded as the deepest preserved part of the basin infill (Horný, Reference Horný1960; Kříž, Reference Kříž1991, Reference Kříž1992).
2.a. Location and previous research
The Všeradice section is located on a gentle, S-facing slope along a field track running NNW from the Bykoš–Všeradice road, NW from Všeradice village (Fig. 1). Rather discontinuous exposures and subcrops may be traced along the field track over a length of about 200 m. Close to the summit, bedrock becomes buried under thick drift deposits. The section represents the shale-dominated, relatively offshore facies of the Silurian succession in the Prague Synform, beginning with the middle Wenlock and terminating with the lower part of the Pridoli Series. Sedimentation here was little influenced by syn-sedimentary basalt volcanic activity, which left a significant record in most other Silurian sections of Bohemia (see e.g. Kříž in Chlupáč et al. Reference Chlupáč, Havlíček, Kříž, Kukal and Štorch1998 for instance).

Figure 1. Position of the area studied within the Czech Republic. (a) Distribution of the Silurian rocks of the Prague Synform; and (b) detailed position of the Všeradice section on a topographic map.
The first note on this important section was published by Horný (Reference Horný1960). Much later, Kříž (Reference Kříž1992) presented a more detailed, but still incomplete log. His section started from marly shales high in the Homerian Cyrtograptus lundgreni Biozone and terminated with the middle Ludfordian intraformational conglomerate (bed no. 13 of this paper). Shale beds with the ostracod Entomis migrans and some bivalves, encountered just below the conglomerate, Kříž (Reference Kříž1992) referred to the ‘B. bohemicus tenuis Interzone’. Kříž et al. (Reference Kříž, Dufka, Jaeger and Schönlaub1993) and later Kozłowska-Dawidziuk, Lenz & Štorch (Reference Kozłowska-Dawidziuk, Lenz and Štorch2001) examined the uppermost Wenlock (upper Homerian) part of the section with particular emphasis on the Lundgreni Event and post-extinction recovery. Piras (Reference Piras2006; unpub. Ph.D. thesis, Univ. degli Studi di Modena e Reggio Emilia, 2006) studied the lower Gorstian beds comprising the Neodiversograptus nilssoni and Lobograptus progenitor graptolite biozones. Budil, Collette & Manda (Reference Budil, Collette and Manda2010) studied the phyllocarid fauna from the lower Ludfordian.
In late 2008 and early 2009, excavations were conducted to expose the supposedly graptolite-rich, shale-dominated section through the upper Ludlow strata. Our data came from two up to 2.5 m deep trenches VA and VB, excavated on both sides of the field track (Fig. 1). Trench VA and the upper part of trench VB exposed a c. 24 m thick middle and upper Ludfordian succession up to the base of the Pridoli Series.
2.b. Material and methods
The shale-dominated Ludfordian was sampled for graptolites in a similar way to the late Homerian interval studied by Kozłowska-Dawidziuk, Lenz & Štorch (Reference Kozłowska-Dawidziuk, Lenz and Štorch2001). Isolated limestone beds and tuffitic intercalations enabled division of the 24 m thick succession into 37 arbitrary units of different stratigraphic thickness (beds 1–37 in ascending order, see Fig. 2). Shaly beds were further subdivided into 10–60 cm thick sub-units. Each sub-unit was sampled bed by bed down the section and all of its fossil samples were placed in the same sample bag(s). The majority of these sampling intervals were about 30 cm thick. Many intervals were rich in graptolites; some others were poor or barren. Every graptolite, at least tentatively determinable, was collected from the rock volume of about 0.03 m3. The lower part of the section exposed by trench VB (see Fig. 2) lacks the arbitrary subdivision, but fossils were collected in the same manner, in intervals (sub-units) of similar thickness. Only a few graptolites were found in the limestone and tuffitic beds.

Figure 2. Chronostratigraphy and lithology of the Všeradice section. Ranges of graptolites, conodonts and selected other fossils are included.
The carbon isotope record of upper Silurian carbonates coming from deeper water environments of the Prague Basin has never been studied, in contrast to the carbonates from shallower environments (Lehnert et al. Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b). The carbonates of the Všeradice section were sampled for determination of δ13Ccarb isotope evolution through the late Ludlow Kozlowskii Event. The isotopic composition of organic carbon from shales occurring between the carbonate beds was not studied owing to the relatively high degree of weathering. The carbon isotope sampling of the Všeradice section (Fig. 2) started about 8.5 m below the late Ludlow Kozlowskii Event. The highest sample was collected from lowermost Pridoli beds (about 2 m above the first appearance (FA) of Neocolonograptus parultimus). Fifteen samples were analysed for δ13C: 12 samples were taken from the Ludlow part of the section and 3 samples from the lowermost Pridoli. A few milligrams of rock powder (preferably micrite) were recovered with a dental drill from rock samples or polished slabs. Carbonate samples were decomposed in a vacuum by 100% phosphoric acid at a temperature of 25°C. The carbon and oxygen isotopic composition of the released CO2 was measured with a Finnigan Mat 251 mass spectrometer. All values are reported in ‰ relative to V-PDB (Vienna Pee Dee Belemnite) by assigning a δ13C value of +1.95 ‰ and a δ18O value of 2.20 ‰ to NSB 19. Accuracy and precision were controlled by replicate measurements of laboratory standards and were better than ± 0.1 ‰ for both carbon and oxygen isotopes.
Conodont samples were collected from the majority of limestone beds and nodules uncovered in both trenches across the interval from Ludfordian to the lowermost Pridoli. A total of 28 samples of average weight between 3 and 5.5 kg was taken mostly from the entire thickness of each limestone bed (bulk samples). The rock was broken into approximately 5 cm sized pieces and processed using standard acid techniques with 10–15% acetic acid. Residues were washed through calibrated double sieves of 2.24 mm (upper) and 90 μm (lower) meshes. Wet residues were washed with acetone and dried. Dried, relatively voluminous residues (of up to 250 ml), with a prevalence of pyrite, goethite and complex silicates (e.g. nagashimalite, macfallite) were concentrated using the heavy liquids diiodomethane (density 3.3 g/cm3) and tribromomethane. Conodonts were obtained from only nine of the limestone beds; no conodonts were extracted from nodules. The conodont material was documented directly in aluminium cells using a Leica D160 digital camera and a Leica MZ7s stereomicroscope, and scanning electron microscope (SEM) images of selected conodont elements (coated with gold) were made by using a Cameca SX100 electron microscope.
Graptolites are housed in the collection of Petr Štorch in the Czech Geological Survey. Non-graptolite macrofauna is deposited in the collection of Štěpán Manda in the Czech Geological Survey (unnumbered specimens in locality collection, prefix SM Všeradice section). The conodont material is temporarily stored in the collection of Ladislav Slavík at the Institute of Geology, Academy of Sciences, Czech Republic and will be later deposited as a part of a larger collection in the National Museum, Prague.
2.c. Sedimentary succession, benthic fauna and depositional environment
The lower part of the section consists of light grey laminated calcareous shale (beds 1–10, Bohemograptus tenuis–Neocucullograptus kozlowskii biozones). Graptolites and pelagic ostracods (Entomis migrans Barrande, Bolbozoe bohemica Barrande) are the most common faunal elements, but their abundance is strongly variable. Pelagic orthocerids (Parakionoceras originale Barrande, Michelinoceras michelini Barrande, Kopaninoceras sp., Arionoceras sp.) and the phyllocarid Ceratiocaris sp. occur only in some beds. Flattened smooth atrypids are present occasionally, but on some bedding planes they occur abundantly. Bivalves are rather rare: Cardiola docens Barrande, C. aff. docens Barrande, C. cf. pectinata Barrande, C. aff. foma Barrande, Butovicela migrans (Barrande) and Tenka aspirans (Barrande). Shale bed no. 3 yielded the trilobite Cromus beaumonti Barrande. Up to 20 cm thick beds of grey argillitic, usually laminated, mudstones occur throughout this part of the section. Limestones contain occasional graptolites and cephalopods; in one case graptolites were found to be oriented by currents and wrapped around a cephalopod shell lying obliquely to the bedding plane – a so-called ‘comet’ (Ferretti & Kříž, Reference Ferretti and Kříž1995). Limestone nodules rich in pyrite, with a maximum diameter of 20 cm occur sporadically. Thin, 1–3 mm thick lenticular laminae of mudstone are common in the shales. Seven beds of yellow-grey, upward fining tuffites, 5 to 25 cm thick, were found in this part of the section. Rare trilobite debris (Otarion sp. and unidentified taxa) was found in some tuffites. In addition, tuffitic bed no. 4 contains close to its base small rounded intraclasts of rusty cephalopod packstone with small cephalopod fragments (Michelinoceras michelini, Pseudocycloceras sp., Kopaninoceras sp., Ophioceras simplex Barrande), common disarticulated bivalves (Cardiola docens) and rare rhynchonellid brachiopods.
The shale-dominated sequence suggests an offshore hemipelagic environment. Some light coloured laminae contain brachiopods and bivalves, which indicates the occasional activity of weak bottom currents ventilating the otherwise anoxic conditions close to the sea bottom. Deposition of shales was interrupted by that of tuffite layers, which also include skeletal fragments derived from shallow water environments (e.g. intraclasts of cephalopod limestone, trilobite fragments).
The shale-dominated sequence ends with a thin bed of fine-grained mud-wackestone overlain by bioturbated tuffitic shale (bed nos 11 and 12), but no body fossils were found.
The section continues with a large lens, up to 160 cm thick, formed of inverse graded intraformational conglomerate (bed no. 13). The matrix consists of coarse, light grey crinoidal grainstone, which passes upward into fine-grained wackestone (Fig. 3). Crinoid ossicles are abraded only in the lower part. Brachiopods are usually disarticulated, but well preserved including fine growth structures in shells. Atrypa evenida Havlíček is the most common: from 126 collected shells, 72 are ventral valves, 53 are dorsal valves, and only one shell is articulated (for a complete list of brachiopods see Kříž, Reference Kříž1992). Fragments of trilobites, Ananaspis fecunda (Barrande), proetids, odontopleurids, corals and gastropods are also present. Pebbles are usually rounded; their abundance and size (maximum diameter is about 25 cm) increase from the base of the bed upward. Pebbles include four lithological types that differ also in their fossil content: (1) small rounded pebbles of dark grey wackestone with cephalopods and the bivalve Cheiopteria glabra Goldfuss (which indicates a latest Nc. kozlowskii Biozone age; see Kříž, Reference Kříž, Boucot and Lawson1999a; Manda & Kříž, Reference Manda and Kříž2006); (2) rounded pebbles of brachiopod-trilobite wackestone with the trilobite Ananaspis fecunda; (3) small angular intraclasts of rusty fine-grained packstone with ostracods, the brachiopod Atrypa sp. and trilobite A. fecunda; (4) intraclasts that represent probably exhumed limestone nodules with Entomis migrans (i.e. Saetograptus linearis–Nc. kozlowskii Zone in age; see Kříž, Reference Kříž, Boucot and Lawson1999a). The conglomerates are overlain by an up to 10 cm thick bed of dark grey mudstone with some cracks on the upper bedding plane.

Figure 3. Microfacies of the carbonate gravity deposits from the Všeradice section. (a) Coarse-grained grainstone with an intraclast of dark grey mudstone with dolomite, bed no. 13. (b) Coarse-grained grainstone with crinoids, trilobites and rugose coral, small rounded intraclasts (wackestone with sponges and ostracod fragments and fine-grained wacke-grainstone), bed no. 13. (c) Partly pyritized rugose coral with partly damaged wall preserved in grainstone, bed no. 13. (d) An intraclast of mudstone with ostracods; smaller ostracod is filled with pyrite, bed no. 13. (e) Intraclastic crinoidal grainstone with cohesive intraclasts of shale and a fragment of tabulate coral, bed no. 22. (f) Detail of contact of matrix with cohesive intraclasts, bed no. 22. Scale bars 2 mm, in (d) 0.5 mm.
The intraformational conglomerate probably represents a canalized debris flow because it forms large lenses in the shale and shows clear inverse grading (e.g. Einsele, Reference Einsele2000, p. 214). The fauna preserved in the matrix of the conglomerate suggests transport from a nearby shallow water locality.
The overlying sedimentary succession comprises dark grey mudstones (particularly nodular) intercalated with dark grey laminated shale (bed nos 14–20, Pseudomonoclimacis latilobus–Slovinograptus balticus Biozone). The shale contains common graptolites and rare cephalopods (e.g. Parakionoceras originale, Michelinoceras michelini, Arionoceras sp.), phyllocarids (Ceratiocaris sp.) and a few benthic remains including scolecodonts, brachiopods (Plectodonta sp.), smooth ostracods and juvenile gastropods. Higher up a distinct tuffitic bed is present. The tuffite is locally eroded and filled with cross-stratified, rusty, upward fining grainstone with cohesive shale intraclasts (bed nos 21–22). The grainstone contains crinoids, trilobites, corals, ostracods, cephalopods and other fossils (see Table 1). This limestone is overlain by a thin bed of tuffite (bed no. 23) and, higher up, shale (bed no. 24). The succeeding strata comprise a mudstone bed overlain by a tuffitic bed (bed nos 25–26).
Table 1. Fauna and selected taphonomic features recorded in limestone bed no. 22 (late Ludfordian, Pseudomonoclimacis latilobus–Slovinograptus balticus Biozone)

Sample weight is c. 15 kg. Abbreviations: S – number of specimens; D and A – number of disarticulated/articulated specimens in brachiopods; C and P – numbers of cephalons/pygidia in trilobite remains. S and D mean deeper/shallow-water faunal elements, respectively. c – common; r – rare. This taphocoenosis is composed of two different faunas mixed together in a gravity flow deposit. The respective faunas originated from relatively shallow and deeper slope settings.
The sequence above the intraformational conglomerates was deposited at the margin of a bathymetric high, as indicated by the shales with benthic fauna and limestone beds. The skeletal limestone filling the depressions in the tuffite probably represents storm event deposits with cross-stratification, normal grading and shale intraclasts (see Fig. 3). Brachiopods are usually disarticulated and large brachiopods and trilobites are broken into small fragments (Table 1). The brachiopod assemblage is dominated by the shallow water brachiopod Kirkidium sp. accompanied by the rather deeper water Jarovathyris canaliculata (see Havlíček & Štorch, Reference Havlíček and Štorch1990); this suggests mixing of two distinct depth-related communities during transport.
The upper part of the studied section (beds 27–43, Pristiograptus fragmentalis and Ne. parultimus–Ne. ultimus biozones) is developed as calcareous shale and platy argillite mudstone, but the shale dominates. One thin bed (no. 36) of grey, upward fining crinoidal wackestone is present. It contains common, minute fragments of brachiopods (atrypids, orthids, leptaenids and strophomenids), trilobites (Otarion sp. and Prionopeltis striata Šnajdr) and disarticulated ostracods. Shale beds yielded graptolites and rare pelagic cephalopods. Bed no. 27 contains also pyritized shells of juvenile cephalopods and gastropods. A thin level with accumulations of the flattened atrypid Lissatrypa sp. was observed in mudstone bed no. 30.
This part of the succession was deposited during deepening, which corresponds with that interpreted in other coeval sections of the Prague Synform (Horný, Reference Horný1955; Kříž et al. Reference Kříž, Jaeger, Paris and Schönlaub1986; Kříž, Reference Kříž1991; Manda & Kříž, Reference Manda and Kříž2006). The sparse or absent benthic fauna suggests usually anoxic conditions at the bottom. Bed no. 36, of fine-grained skeletal limestone, probably reflects a storm event as indicated by its sharp base and upward fining. Rounded crinoid debris and small fragments of disarticulated atrypid brachiopods indicate a significant amount of transport.
2.d. Gaps in sedimentation and source area of conglomerates
The Všeradice section is located in the Western Segment (Kříž, Reference Kříž1992) of the Prague Synform. This is characterized by offshore hemipelagic sedimentation throughout the Silurian as indicated by the shale-dominated sequence ranging from the lowest Llandovery to the Ludlow (Horný, Reference Horný1955, Reference Horný1960). Common tuffite beds throughout the Ludlow succession indicate nearby volcanic activity and deposition of reworked volcaniclastic material. Tuffites in the upper Ps. latilobus–Sl. balticus Biozone are the youngest known manifestation of volcanic activity in the Silurian strata of the Prague Synform, although coeval basalt effusives are unknown. More limestones (wacke-mudstone and gravity deposits) appear in the middle and upper Ludfordian parts of the succession, which is consistent with basinward extension of shallow water sedimentation into the Western Segment of the synform (Horný, Reference Horný1955, Reference Horný1960). This proliferation of carbonates is correlatable with the latest Ludlow lowstand well documented in coeval limestone sequences elsewhere in the Prague Synform (Horný, Reference Horný1955; Kříž, Reference Kříž1991; Kříž in Chlupáč et al. Reference Chlupáč, Havlíček, Kříž, Kukal and Štorch1998; Manda & Kříž, Reference Manda and Kříž2006). Earlier lowstands (those in the late Wenlock and early Ludfordian) are not distinguishable in the sedimentary log in the Všeradice section and elsewhere in the Western Segment (Horný, Reference Horný1955, Reference Horný1960). Kříž (Reference Kříž1991) suggested that intense subsidence occurred in this part of the basin in the Ludlow. However, a relatively thin sedimentary succession, including limestones of the uppermost Ludlow, probably indicates reduced accommodation space. Canalized debris-flow deposits indicate the appearance of nearby shallow water environments (although these are not preserved in the rock record) within the Western Segment, which was isolated from the main shallow platforms located further to the north. Although corresponding with the lowstand recorded in coeval limestone strata, eustatic sea-level oscillation cannot explain fully the appearance of this local shallow water area, i.e. a small-scale carbonate domain surrounded by shale deposition. The submarine high was most probably associated with local, synsedimentary uplift. Sea-level drawdown, associated with the middle Ludfordian Kozlowskii and/or Lau Event initiated only a debris flow from the neighbouring submarine high. Slopes were probably relatively steep, as suggested by the exhumed limestone nodules with ostracods in the debris flow. These nodules (but no surrounding matrix) contain fine framboidal dolomite, which directly documents the early diagenetic origin of the dolomite.
The stratigraphically significant shelly fauna preserved in the limestone debris flow (intraformational conglomerate), the lenticular cross-section of the conglomerate body and the much reduced thickness of the post-Kozlowskii Ludfordian indicate prominent erosion and a gap in sedimentation in the Všeradice section, when compared to coeval strata in the limestone-dominated sections of the Prague Synform.
The temporal and spatial extent of the gap connected with these canalized debris-flow deposits is evident from analysis of faunas in the matrix and pebbles (i.e. it spans the interval between the last appearance (LA) of Nc. kozlowskii until at least the FA of the index trilobite A. fecunda). However, allochthonous fauna from both intraclasts and matrix make correlation with coeval carbonate sequences of the Prague Synform possible (see discussion in Section 3).
2.e. Conodont fauna
The late Silurian conodont biozonation was developed by Walliser (Reference Walliser1964), who included the conodont data from the Prague Synform into his first biozonal framework. Schönlaub (in Chlupáč, Kříž & Schönlaub, Reference Chlupáč, Kříž and Schönlaub1980 and in Kříž et al. Reference Kříž, Jaeger, Paris and Schönlaub1986) furnished crucial conodont data from sections in the upper Ludlow and Pridoli of the Prague Synform and demonstrated taxon ranges. Schönlaub in Kříž et al. (Reference Kříž, Jaeger, Paris and Schönlaub1986) made an extensive summary of conodont faunas obtained from samples from numerous sections and localities that were sampled and studied by him or by previous authors. He recognized a succession of Walliser's biozones for the Ludlow and Pridoli: Ancoradella ploeckensis, Polygnathoides siluricus, Pedavis latialatus, ‘Ozarkodina’ snajdri, ‘Oz.’ crispa and ‘Oz.’ eosteinhornensis. The Ludlow part of this biozonation is in use without large changes; the ‘Oz.’ eosteinhornensis sensu stricto and ‘Oz.’ detorta biozones suggested for the Pridoli by Jeppsson (Reference Jeppsson1988) are problematic or inapplicable (see discussions in Carls, Slavík & Valenzuela-Ríos, Reference Carls, Slavík and Valenzuela-Ríos2007).
The conodont faunas of the Všeradice section (Fig. 4) are studied for the first time in this paper. The conodont sampling was successful only in one third of the samples, which yielded c. 350, mostly fragmented conodont elements. The conodont faunas are rather poor regarding taxonomic diversity and are of low abundance in general, with the exception of the intraformational conglomerate (bed no. 13) and grainstone bed no. 22 where the numbers of conodont elements exceed 80. A relatively rich conodont sample was obtained from bed no. 28 with more than 70 conodont elements. The composition of conodont faunas in all samples is well balanced and variable regarding the size of the elements; also the frequent presence of tiny, simple-cone elements suggests that assemblage composition was not strongly affected by hydrodynamic sorting of particles. The depositional characters of some carbonate layers (i.e. intraformational conglomerate, presence of intraclasts and grainstone fillings) described in the previous Sections suggests the likelihood of stratigraphical mixing of conodont elements, but evidence for conodont redeposition was not found.

Figure 4. Conodonts from the Všeradice section (SEM images). All images are at the same magnification (scale bar represents 500 μm). (a) Kockelella maenniki Serpagli & Corradini, oral view of Pa element, bed no. 2. (b) Kockelella cf. a. absidata, lateral view of Pa element, bed no. 2. (c) Kockelella absidata sardoa Serpagli & Corradini, lateral view of Pa element, bed no. 4. (d) Kockelella absidata ssp., lateral view of Pa element, bed no. 9. (e) Polygnathoides siluricus Branson & Mehl, oral view (ea) and lateral view (eb) of incomplete Pa element from bed no. 9. (f) Wurmiella inclinata (Rhodes), lateral view of Pa element, bed no. 13. (g) W. inclinata, lateral view of Pa element from the base of bed no. 13. (h) W. inclinata, lateral view (ha) and oral view (hb) of Pa element from base of bed no. 13. (i) Wurmiella excavata excavata (Branson & Mehl), lateral view (ia) and oral view (ib) of Pa element from base of bed no. 13. (j) W. cf. excavata, lateral view of Pa element, bed no. 2. (k) Ozarkodina typica, lateral view (ka) and oral view (kb) of Pa element, bed no. 9. (l) Ozarkodina typica Branson & Mehl, lateral view (la) and oral view (lb) of Pa element from the base of bed no. 13. (m) O. typica, lateral view (ma) and oral view (mb) of Pa element from the base of bed no. 13. (n) O. cf. typica, lateral view (na) and oral view (nb) of Pa element, bed no. 22. (o) Oz. typica, lateral view (oa) and oral view (ob) of Pa element from the base of bed no. 13. (p) Oz. typica, lateral view (pa) and oral view (pb) of Pa element from the base of bed no. 13. (q) Delotaxis sp., lateral view of incomplete element from the base of bed no. 13. (r) ‘Ozarkodina’ snajdri–crispa transition, oral view of incomplete Pa element, bed no. 28. (s) Oz. typica, lateral view of Pa element, bed no. 22. (t) ‘Ozarkodina’ snajdri or crispa, lateral view of Pb element, bed no. 22. (u) Oz. typica, lateral view of Pb element from the base of bed no. 13. (v–z) ‘Ozarkodina’ snajdri Walliser, lateral views (va–za) and oral views (vb–zb) of Pa elements from bed no. 22.
The preservation of conodonts in samples is good. Apart from long-ranging simple cones (e.g. belodellids, Panderodus), the conodont material obtained enables recognition of the following conodont genera: Delotaxis, Kockelella, Ozarkodina, Polygnathoides and Wurmiella. Owing to fragmentation, some taxonomic assignments are only to generic level, but several stratigraphically important taxa were recognized and enabled delimitation of the P. siluricus, ‘Oz.’ snajdri and ‘Oz.’ crispa conodont biozones in the middle–upper Ludfordian (see Fig. 4). The stratigraphically lowest evidence for the P. siluricus Biozone is the presence of Kockelella maenniki Serpagli & Corradini in bed no. 2. This species, erected in Sardinia (Serpagli & Corradini, Reference Serpagli and Corradini1998, Reference Serpagli and Corradini1999), is confined to the lower–middle part of the P. siluricus Biozone. Representatives of the genus Kockelella were found also in beds nos 4 and 9. Following the reconstruction of the origin and phylogeny of the genus given by Serpagli & Corradini (Reference Serpagli and Corradini1999), Kockelella absidata sardoa (see Serpagli & Corradini, Reference Serpagli and Corradini1999) found in bed no. 4 ranges into the lower P. siluricus Biozone; Pa elements of the long-ranging Kockelella absidata group were found in beds nos 2 and 9. The record of K. absidata ssp. together with P. siluricus points to the highest stratigraphical occurrence of the K. absidata group (Branson & Mehl, Reference Branson and Mehl1933). The distribution of several taxa of the genus Kockelella and their stratigraphic relations within the P. siluricus Biozone are shown in Slavík, Kříž & Carls (Reference Slavík, Kříž and Carls2010) from the Požáry section.
The interval from bed no. 11 to bed no. 19 yielded mostly representatives of the genus Wurmiella (W. excavata excavata (Branson & Mehl) and Wurmiella inclinata (Rhodes)), and Ozarkodina typica Branson & Mehl (= Hindeodella confluens Jeppsson). Oz. typica particularly shows high intra-specific variability as regards denticulation of the blade, variation in ‘cockscomb’ structure and proportions of the Pa element (length and width of the unit). The stratigraphical evaluation of the changing morphology of the Pa element of Oz. typica, however, requires more material with precise stratigraphical control. The P. latialatus Biozone could have been expected to be found within this interval, but there is no direct evidence for this biozone and there is also no proof that the coniform element of Acodina? sp. may truly belong to the apparatus of Pedavis Klapper & Philip.
The ‘Oz.’ snajdri Biozone is directly documented by several Pa elements of the name-bearer taxon in bed no. 22. Transition Pa elements between ‘Oz.’ snajdri and ‘Oz.’ crispa were obtained from bed no. 28. The element shown in Figure 4r exhibits already a large undulated platform like that seen in typical ‘Oz.’ crispa, but the termination of the blade within the platform is in question. The taxa overlap considerably stratigraphically; however, ‘Oz.’ snajdri appears at a lower stratigraphical level in the Prague Synform (Walliser, Reference Walliser1964; Schönlaub in Chlupáč, Kříž & Schönlaub, Reference Chlupáč, Kříž and Schönlaub1980). Accordingly, the base of the ‘Oz.’ crispa Biozone can be expected to lie close to this stratigraphical level. The generic name of these biozonally diagnostic taxa is placed in quotation marks because of the ambiguity over the generic assignment as explained in Murphy, Valenzuela-Ríos & Carls (Reference Murphy, Valenzuela-Ríos and Carls2004). Bed no. 36 is the highest bed that yielded conodonts, although these are of no biostratigraphical value, the Pridoli age of this level being indicated by graptolites only.
2.f. Graptolite fauna
Graptolite rhabdosomes are uncommon in this hemipelagic, shale-dominated offshore succession, if compared to the rich early Silurian and even early Ludlow taphocenoses. Diversity is moderate to low. Twenty graptolite taxa have been identified from the middle and upper Ludfordian and lowest Pridoli. Despite the relative scarcity, however, graptolites are still playing a dominant role in the preserved pelagic macrofauna, being associated with myodocopid ostracods before the Kozlowskii Event. Shale beds with few graptolite rhabdosomes are rich in ostracods (accompanying cephalopods) and vice versa. The principal graptolite taxa are figured in Figures 5, 6 and 7. All biozones applied in this paper are interval zones defined as an interval between two biohorizons (see International Stratigraphic Guide edited by Salvador, Reference Salvador1994).

Figure 5. Middle and late Ludfordian graptolites from the Všeradice section. (a) Neocucullograptus kozlowskii Urbanek: PŠ 2028, upper part of bed no. 10; (b, f, g, k–n) Slovinograptus balticus (Teller): (b) PŠ 2020, (f) PŠ 2011, (g) PŠ 2018, (k) PŠ 2016, (l) PŠ 2017–1, (m) PŠ 2017–2, (n) PŠ 2015, bed no. 20; (c) Linograptus posthumus R. Richter: PŠ 2029, lowermost part of bed no. 7; (d, e, j, r, s) Pseudomonoclimacis latilobus (Tsegelnjuk): (d) PŠ 2004, (e) PŠ 2023, (j) PŠ 2001, (r) PŠ 2006, (s) PŠ 2003, (d, j, r, s) bed no. 14, (e) bed no. 20; (h) Pristiograptus dubius (Suess): PŠ 2024, bed no. 31; (i) Pseudomonoclimacis kosoviensis (Bouček): PŠ 2009, bed no. 16; (o) Polonograptus chlupaci sp. nov.: PŠ 2030, lowermost part of bed no. 7; (p) Pristiograptus fragmentalis (Bouček): PŠ 2031, bed no. 27; (q) Pseudomonoclimacis dalejensis (Bouček): PŠ 2027b, bed no. 10. Vertical scale bars represent 1 mm.

Figure 6. Middle and late Ludfordian and basal Pridoli graptolites from the Všeradice section. (a, b) Formosograptus formosus (Bouček): (a) PŠ 2025, (b) PŠ 2303, bed no. 37; (c, d) Neocolonograptus parultimus (Jaeger): (c) PŠ 2026, (d) PŠ 2302, bed no. 37; (e) Slovinograptus cf. balticus (Teller): PŠ 2307, bed no. 33; (f) Crinitograptus? sp.: PŠ 2308, bed no. 33; (g–j) Pseudomonoclimacis kosoviensis (Bouček): (g) PŠ 2244; (h) PŠ 2114, (i) PŠ 2101, (j) PŠ 2110a, lowermost part of section VB; (k) Neocucullograptus inexpectatus (Bouček) ssp.: PŠ 2100, lowermost part of the section designated VB; (l) Pseudomonoclimacis aff. kosoviensis (Bouček): PŠ 2304, bed no. 27; (m) Pristiograptus fragmentalis (Bouček): PŠ 2309, bed no. 27; (n) Neocolonograptus ultimus (Perner): PŠ 2310, bed no. 39. Vertical scale bars represent 1 mm.

Figure 7. Middle and late Ludfordian and basal Pridoli graptolites from the Všeradice section. (a) Neocucullograptus inexpectatus (Bouček) ssp.: PŠ 2100, lowermost part of the section designated VB; (b) Linograptus posthumus R. Richter: PŠ 2029, lowermost part of bed no. 7; (c) Formosograptus formosus (Bouček): PŠ 2303, bed no. 37; (d) Neocolonograptus parultimus (Jaeger): PŠ 2302, bed no. 37; (e) Neocolonograptus ultimus (Perner): PŠ 2310, bed no. 39; (f) Slovinograptus cf. balticus (Teller): PŠ 2307, bed no. 33; (g) Bohemograptus tenuis (Bouček): PŠ 2098, lowermost part of section VB; (h) Crinitograptus? sp.: PŠ 2308, bed no. 33; (i) Pseudomonoclimacis dalejensis (Bouček): PŠ 2027b, bed no. 10; (j, k) Pristiograptus fragmentalis (Bouček): (j) PŠ 2309, (k) PŠ 2031, bed no. 27; (l, p) Slovinograptus balticus (Teller): (l) PŠ 2018, (p) PŠ 2016, bed no. 20; (m) Neocucullograptus kozlowskii Urbanek: PŠ 2028, upper part of bed no. 10; (n) Pristiograptus dubius (Suess): PŠ 2024, bed no. 31; (o) Polonograptus chlupaci sp. nov.: PŠ 2030, lowermost part of bed no. 7; (q) Pseudomonoclimacis latilobus (Tsegelnjuk): PŠ 2006, bed no. 14; (r) Pseudomonoclimacis aff. kosoviensis (Bouček): PŠ 2304, bed no. 27; (s) Pseudomonoclimacis kosoviensis (Bouček): PŠ 2110a, lowermost part of section VB; All specimens × 5, vertical scale bar represents 1 mm.
The lower part of the succession described in this paper was exposed in the upper part of the VA trench, on the western side of the field track. It is developed in the form of partly bleached, brownish calcareous shales with Pristiograptus dubius (Suess) sensu lato, Pseudomonoclimacis kosoviensis (Bouček) and Bohemograptus tenuis (Bouček), and referred to the uppermost part of the lower–middle Ludfordian B. tenuis Biozone. The same lithology, but intercalated with several muddy limestone and tuffitic-carbonate beds, continues through the succeeding Neocucullograptus inexpectatus and Neocucullograptus kozlowskii biozones.
The lower part of the 8.7 m thick Nc. inexpectatus Biozone was exposed in the overlapping parts of the neighbouring trenches VB and VA. Small, long-ranging P. dubius, Pseudomonoclimacis dalejensis (Bouček), B. tenuis, large rhabdosomes of Ps. kosoviensis (Bouček) and the biozonal index species Neocucullograptus inexpectatus (Bouček) are common in the shale. Uncommon Linograptus posthumus (R. Richter), Bohemograptus praecornutus Urbanek and Polonograptus chlupaci sp. nov. have been collected from the upper part of the inexpectatus Biozone on the eastern side of the track, in the VA trench.
Higher in the VA trench Nc. inexpectatus is replaced by a more derived form, Neocucullograptus kozlowskii Urbanek, which is the index species of the next biozone. Apart from the different index graptolite, the 3.2 m thick kozlowskii Biozone contains an assemblage similar to that of the inexpectatus Biozone. Polonograptus podoliensis Přibyl, a prominent but uncommon element in the assemblage of the Nc. kozlowskii Biozone in Kosov Quarry near Beroun (Štorch, Reference Štorch1995a) and a few other sections (Přibyl, Reference Přibyl1983), has not been recovered from the Všeradice section.
The continuous graptolite record and sedimentation were interrupted at the base of limestone bed no. 11 and graptolites reappeared only above the intraformational conglomerate (Figs 2, 3, bed no. 13).
The mass extinction of the middle Ludfordian graptolite fauna at the top of the kozlowskii Biozone has been described and discussed by Urbanek (Reference Urbanek1970, Reference Urbanek1993). Although all graptolites with ventrally curved rhabdosomes (Bohemograptus, Neocucullograptus and Polonograptus) vanished, some generalists such as Pristiograptus dubius, Linograptus posthumus and Pseudomonoclimacis kosoviensis reappeared soon after the extinction, together with several newcomers of rather cryptic origin.
Graptolites appearing in the first shale intercalations within the limestone-dominated interval immediately above the intraformational conglomerates belong to a new, post-extinction assemblage of the Ps. latilobus–Sl. balticus Biozone. The assemblage, dominated by Ps. latilobus (Tsegelnjuk), comprises also Sl. balticus (Teller), L. posthumus, Ps. kosoviensis and a slender form of P. dubius.
Higher in the succession Sl. balticus has been replaced by the more slender, weakly S-shaped Sl. cf. balticus (Teller) accompanied by Pseudomonoclimacis aff. kosoviensis, Crinitograptus? sp. and the long-ranging L. posthumus and P. dubius. Rare finds of Pristiograptus fragmentalis (Bouček) in bed no. 27 enable recognition of a specific P. fragmentalis Biozone in the interval between the last joint occurrence of Ps. latilobus and Sl. balticus and the first occurrence of Neocolonograptus parultimus (Jaeger), the latter indicating the base of the Pridoli Series.
In the uppermost shale beds exposed in the trench (beds nos 35–41), Ne. parultimus is accompanied by Neocolonograptus ultimus (Perner), Formosograptus formosus (Bouček), Crinitograptus? sp., L. posthumus, the last P. fragmentalis and a slender form of P. dubius.
2.g. Carbon isotopes
The δ13Ccarb values from the Všeradice section range from –2 to +2 ‰, and in contrast to earlier studied sections of the Prague Basin (Lehnert et al. Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b), no distinct positive δ13C shift was observed above the level of the Kozlowskii Event. The lack of this δ13C positive excursion as well as the relatively low δ13Ccarb values in the Všeradice section (frequently close to or just below zero) may suggest an absence of sedimentary record equivalent to beds with the high positive δ13C values and/or a slight diagenetic overprint. A high content of organic carbon in the shale-dominated succession could cause a lowering of the δ13Ccarb values during diagenetic processes. The decomposition of organic matter yields isotopically light carbon, which can then be precipitated as carbonate, which pushes the δ13Ccarb values down in organic-rich carbonates. Because of that we cannot determine whether the measured low δ13C values represent the primary isotopic composition of the studied micrites or values that have been slightly secondarily lowered by diagenetic overprint, or both. The only positive δ13C values were recorded in an interval just above the late Ludlow Kozlowskii Event roughly to a level about 4.5 m below the FA of ‘Ozarkodina’ snajdri. However, no distinct positive δ13C shift was found in the carbonates from the Všeradice section (Fig. 8).

Figure 8. Correlation of the Všeradice section with a shallow water succession exposed in the New Kosov Quarry section showing lithology, ranges of selected fossils, relative eustatic oscillation and carbon isotope curve. Note that conodonts are not included because conodonts have not been studied in Kosov Quarry. The grey field indicates the interval of a sedimentary break in the Všeradice section. Data from Kosov Quarry are adopted from Kříž (Reference Kříž1992) and Lehnert et al. (Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b).
Previous studies (Lehnert et al. Reference Lehnert, Frýda, Buggisch and Manda2003, Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b) revealed a distinct positive shift of the δ13Ccarb values from the carbonate platform of the Prague Basin. The most complete isotopic peak was recorded from the Kosov Section (Fig. 8) where the positive δ13C shift starts at the level of the late Ludlow Kozlowskii Event (Fig. 8). The values in the subsequent part of the Kosov Section (between 6.7 and 8 ‰) show no indication of a decrease in δ13C. Another, earlier studied section, the Mušlovka Quarry Section, clearly shows that this δ13C positive excursion ends before the FA of ‘Oz.’ snajdri. The level of the late Ludlow Kozlowskii Event was confidently indicated biostratigraphically in the Všeradice section; however, determination of the FAD of ‘Oz.’ snajdri is less certain because only a few conodont elements of the latter conodont species were found. The true FA of ‘Oz.’ snajdri may thus be lower in the section than shown (Fig. 2).
In the Všeradice section, the level of the late Ludlow Kozlowskii Event is just below the intraformational conglomerate (bed no. 13), which probably corresponds to the prominent gap in the sedimentary record (Fig. 2). The absence of a distinct positive shift of the δ13C values in the Všeradice section likely suggests an absence of sedimentary record equivalent to beds with the high positive δ13C values in the Kosov Section (Fig. 8).
3. Graptolite biozones and correlation
Graptolite biozones are used as the primary correlation tool as the conodonts from the section are not sufficient to compose a precise zonal scheme. Whenever possible the shelly fauna and its potential for correlation are commented upon.
3.a. The Bohemograptus tenuis Biozone
Přibyl (Reference Přibyl1983) designated an interval between the demise of the saetograptids and the first occurrence of Neocucullograptus inexpectatus (Bouček) as the ‘Pristiograptus longus’ Biozone with, at its top, the Bohemograptus bohemicus Bed. ‘Pristiograptus longus’ (Bouček) is considered herein to be a junior synonym of Pristiograptus dubius (Suess) sensu lato. Bohemograptus bohemicus (Barrande) is considerably outnumbered by its evolutionary offshoot Bohemograptus tenuis (Bouček) in this level. We prefer to define the entire interval between the last saetograptids and the first Neocucullograptus as the Bohemograptus tenuis Biozone. This interval is probably coeval with the Bohemograptus bohemicus–Neolobograptus auriculatus biozones erected by Teller (Reference Teller1969) and Urbanek (Reference Urbanek1970) in the subsurface Silurian rocks of Poland and further updated by Urbanek & Teller (Reference Urbanek and Teller1997). Holland & Palmer (Reference Holland and Palmer1974) used the B. tenuis Biozone to designate strata with a monospecific bohemograptid fauna overlying the Saetograptus leintwardinensis Biozone in the Welsh Borderland (see Bohemograptus proliferation Zone in Zalasiewicz et al. Reference Zalasiewicz, Taylor, Rushton, Loydell, Rickards and Williams2009). The present B. tenuis Biozone is correlatable also with the B. tenuis Interzone sandwiched between the S. linearis and Polonograptus podoliensis biozones in Tien Shan, Kyrgyzstan (Koren’ & Sujarkova, Reference Koren’ and Sujarkova2004). Saetograptus linearis is a biozonal index graptolite in Bohemia and Polonograptus podoliensis (Přibyl) is an important component in assemblages of the Nc. inexpectatus and Nc. kozlowskii biozones (Přibyl, Reference Přibyl1983; Štorch, Reference Štorch1995a). Finally, in Bohemia the B. tenuis Biozone is used to name the interval between the late S. leintwardinensis Biozone mass extinction and the subsequent development of the new neocucullograptid and polonograptid fauna. The uppermost part of the B. tenuis Biozone, overlain by the FA of Nc. inexpectatus, is shown in the lowermost part of the section illustrated in Figure 2.
3.b. The Neocucullograptus inexpectatus Biozone
The Nc. inexpectatus Biozone of this paper corresponds to the lower part of the Nc. inexpectatus Biozone of Přibyl (Reference Přibyl1983). The stratigraphically lowest neocucullograptid rhabdosomes define the base of the biozone, whereas its top is defined by the FA of Nc. kozlowskii. Apart from Nc. inexpectatus inexpectatus (Bouček, Reference Bouček1931), Urbanek (Reference Urbanek1970) recognized the more advanced subspecies Nc. inexpectatus supernus that certainly occurs in the upper part of the biozone at Kosov Quarry (Štorch, Reference Štorch1995a). Preservation of the specimens found in the Všeradice section, however, is not good enough to enable separation of the two subspecies. Limestone nodules in tuffite bed no. 4 contain the conodont Ozarkodina fundamentata that indicates the P. siluricus Biozone. In addition, a bivalve assemblage with Cardiola docens occurs in the limestone nodules (bed no. 4). This bivalve assemblage is used in correlations of carbonate sequences in peri-Gondwanan Europe; it ranges from the S. linearis Biozone up to the Nc. inexpectatus Biozone (Kříž, Reference Kříž1998, Reference Kříž, Boucot and Lawson1999a, Reference Křížb, Reference Kříž2010a, Reference Křížb).
3.c. The Neocucullograptus kozlowskii Biozone
Přibyl (Reference Přibyl1983) did not recognize the Neocucullograptus kozlowskii Biozone in Bohemia; hence the upper part of his Nc. inexpectatus Biozone correlates with the Nc. kozlowskii Biozone sensu Štorch (Reference Štorch1995a) and the present paper. The Nc. kozlowskii Biozone comprises an interval between the FA and LA of its name-giving species. The present conception of the Nc. kozlowskii Biozone matches well that introduced for the East European Platform by Urbanek (Reference Urbanek1970) and further elaborated by Urbanek & Teller (Reference Urbanek and Teller1997). The same neocucullograptid biozones are probably applicable in Podolia, although Tsegelnjuk (Reference Tsegelnjuk1976, Reference Tsegelnjuk1981) did not use a Nc. inexpectatus Biozone. A limestone bed just above the base of the Nc. kozlowskii Biozone yielded the index conodont Polygnathoides siluricus. The direct correlation between the Nc. kozlowskii Biozone and upper part of the P. siluricus Biozone has been previously documented in Kosov Quarry only (Lehnert et al. Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b). The upper limit of the Nc. kozlowskii Biozone is defined by the sudden extinction of the bohemograptid–polonograptid–neocucullograptid fauna, defined as the Kozlowskii Event by Urbanek (Reference Urbanek1993). Graptolite extinction and facies change indicating shallowing were identified also in Kosov Quarry near Beroun (Štorch, Reference Štorch1995a, Reference Štorchb). There, however, graptolites are missing in the supposed post-extinction recovery interval.
Limestones and intraformational conglomerates that occur just above the graptolite extinction level in the Všeradice section suggest that the extinction coincides with the beginning of shallowing as suggested by Urbanek (Reference Urbanek1993) and Melchin, Koren’ & Štorch (Reference Melchin, Koren’ and Štorch1998). Apart from graptolites, the ostracod Entomis migrans disappeared (similarly as elsewhere in the Prague Synform, see Manda & Kříž, Reference Manda and Kříž2006). This is a striking feature because this ostracod is a common component of pelagic faunas in peri-Gondwanan basins starting from the lower Ludfordian S. linearis Biozone (Siveter, Vannier & Palmer, Reference Siveter, Vannier and Palmer1991; Kříž, Reference Kříž, Boucot and Lawson1999a). Bolbozoe disappeared from shales in the Nc. inexpectatus Biozone. Note that no pelagic ostracods appeared during the post-extinction recovery. Pelagic orthocerids pass through the Kozlowskii extinction Event and Parakionoceras, Kopaninoceras, Arionoceras and Michelinoceras occur throughout the section. Conodonts found in limestone just above the Kozlowskii Event are the same as in the lowest Nc. kozlowski Biozone except for P. siluricus. This suggests that the deeper water conodont assemblage consists mainly of long-ranging taxa and that the extinction thus affected rather shallow water conodont faunas (see also Slavík, Kříž & Carls, Reference Slavík, Kříž and Carls2010). This should be taken into account when correlating deeper water limestones.
Manda & Kříž (Reference Manda and Kříž2006) concluded that the Kozlowskii Event correlates roughly with the conodont extinction known as the Lau Event (Jeppsson, Reference Jeppsson1993). However, no section with adequately combined graptolite and conodont records is known. Consequently, high-resolution correlation of the graptolite and conodont extinctions is doubtful. Moreover the Lau Event is not clearly manifested in the conodont faunas in the Prague Synform and its lower part is entirely missing in the sections (Slavík, Kříž & Carls, Reference Slavík, Kříž and Carls2010).
In the Kosov Section the LA of P. siluricus (defining the Lau Event) is drawn below the LA of Nc. inexpectatus and B. tenuis. Conodonts are, however, rare in this section. The late Ludfordian carbon isotope excursion which is commonly ascribed to the Lau Event (Saltzman, Reference Saltzman2001; Maartma et al. Reference Maartma, Brazauskas, Kaljo, Kaminskas and Musteikis2005; Lehnert et al. Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b) started in the Kosov Section close to the level of the graptolite extinction.
3.d. The Pseudomonoclimacis latilobus–Slovinograptus balticus Biozone
The graptolite assemblage recovered from shaly intercalations within the post-kozlowskii limestone suite is dominated by the two biozonal index species, Pseudomonoclimacis latilobus (Tsegelnjuk, Reference Tsegelnjuk1976) and Slovinograptus balticus (Teller, Reference Teller1966), both widespread in the East European Platform (e.g. Tsegelnjuk, Reference Tsegelnjuk1976; Paškevičius, Reference Paškevičius1979; Urbanek, Reference Urbanek1997). Přibyl (Reference Přibyl1983) described these two species under the junior synonyms ‘Saetograptus insignitus’ and ‘Monograptus abhorrens’, respectively, from a temporary trench in Velký Vrch hill near Koněprusy. He recognized a distinctive ‘S. insignitus’ Biozone also in Kosov Quarry and in a water supply gallery in Praha-Podolí. We are not surprised to find these graptolites in relatively more offshore facies in the Všeradice section. The Ps. latilobus–Sl. balticus Biozone of this paper is an interval between the last occurrence of Nc. kozlowskii at the base and last joint occurrence of Ps. latilobus and Sl. balticus at the top.
The intraformational conglomerate just below the lowest occurrences of Ps. latilobus and Sl. balticus contains trilobites and other shelly fossils of relatively inshore provenance corresponding with the former Ananaspis fecunda Horizon (Horný, Reference Horný1955). Benthic fossils recovered from both matrix and pebbles of the conglomerate belong to the Ananaspis fecunda–Cyrtia postera Assemblage, which therefore pre-dated the Ps. latilobus–Sl. balticus graptolite assemblage. The relict graptolite fauna that survived the maximum lowstand (shallow water limestones with Ananaspis fecunda overlie deeper water shale and mudstone in the Kosov and Mořina areas) was subsequently joined by the new association of Ps. latilobus and Sl. balticus. Although in the Všeradice section the interval between the Kozlowskii extinction and the FA of Ps. latilobus is only about 2.5 m thick, in Kosov Quarry it comprises at least 10 m of strata. This indicates that the post-extinction interval with strongly impoverished or missing graptolites was remarkably long and the recovery of graptolites was delayed after the benthic faunal recovery. Finally, the limestone sequence with the Ananaspis fecunda–Cyrtia postera Assemblage (A. fecunda Horizon of Horný, Reference Horný1955) developed across the basin, correlates with the Ps. latilobus–Sl. balticus Biozone as used here.
Pseudomonoclimacis latilobus and Slovinograptus balticus range together higher up the succession, close to the FA of Neocolonograptus parultimus (Jaeger), the biozonal index of the lowermost Pridoli. The graptolite assemblage of the present Ps. latilobus–Sl. balticus Biozone is quite similar to that of the homonymous zone applied by Urbanek (Reference Urbanek1997) in Poland. It is uncertain, however, how precisely the two latilobus–balticus biozones correlate. The index taxa of the three succeeding biozones, i.e. Uncinatograptus acer Tsegelnjuk, Uncinatograptus protospineus Urbanek and Uncinatograptus spineus Tsegelnjuk, which overlie the Ps. latilobus–Sl. balticus Biozone in Poland (Urbanek, Reference Urbanek1997), are missing in the Všeradice section and elsewhere in peri-Gondwanan Europe. The lack of these biozones may be explained by the palaeogeographical isolation of peri-Gondwanan Europe in the latest Ludlow. In the Všeradice section, however, some gaps in sedimentation should be also considered since the 2.5 m between the LA of Ps. latilobus and the FA of Ne. parultimus corresponds with a c. 120 m thick sequence in Poland (see Urbanek, Reference Urbanek1997, fig. 3). Přibyl (Reference Přibyl1983) recognized a monospecific ‘Pristiograptus’ fecundus Biozone above his ‘insignitus Biozone’ in Kosov Quarry and some other sections of the Prague Synform. ‘Pristiograptus’ fecundus (Přibyl), which is probably a descendant of Ps. latilobus, has not been recovered from the Všeradice section.
Limestone bed no. 22, containing ‘Oz.’ snajdri and other conodont taxa (Fig. 2), immediately overlies the last occurrence of Ps. latilobus. The FA of ‘Oz.’ snajdri is therefore above the FA of Ps. latilobus.
3.e. The Pristiograptus fragmentalis Biozone
Instead of distinctive range biozones, we can apply only another interval biozone, named after the presumably long-ranging Pristiograptus fragmentalis (Bouček) and defined as an interval between the highest joint occurrence of Ps. latilobus and Sl. balticus and the FA of Ne. parultimus. Pristiograptus fragmentalis (Bouček, Reference Bouček1936) is a robust form having a weakly ventrally curved to almost straight proximal-most part of its rhabdosome and considerably overlapping, pristiograptid distal thecae. Our specimens (Figs 5p, 7j, k) are identical with forms figured by Kříž et al. (Reference Kříž, Jaeger, Paris and Schönlaub1986), Urbanek (Reference Urbanek1997, pl. 15, figs 5–6) and, in part, by Koren’ & Sujarkova (Reference Koren’ and Sujarkova1997, pl. 1, figs 1–6; text-fig. 7A–G).
The Pristiograptus fragmentalis Biozone is the uppermost graptolite biozone of the Ludlow in Bohemia (Bouček, Reference Bouček1936). Bouček (Reference Bouček1936, p. 13) noted that P. fragmentalis occurs in cephalopod limestones with the trilobites Cromus intercostatus Barrande and Cerauroides hawlei (Barrande), below the FA of Ne. ultimus at the Lochkov locality. Cr. intercostatus and C. hawlei form part of the distinct Denckmanites–Cromus trilobite Assemblage established by Chlupáč (Reference Chlupáč1987). This assemblage is known only from a few sections near Lochkov village where it is restricted to a single thin bed. Šnajdr (Reference Šnajdr1985) noted that the latter assemblage occurs 11–14 cm below the FA of Ne. parultimus in the Orthoceras Quarry Section. Kříž et al. (Reference Kříž, Jaeger, Paris and Schönlaub1986) reported P. fragmentalis from the uppermost Ludlow just below the FA of Ne. parultimus (in the Koledník and Karlštejn sections). New data from the Všeradice section suggest that the FA of P. fragmentalis in Bohemia falls immediately after the LA of Ps. latilobus.
The Všeradice section seems to be either condensed or interrupted by a discrete unconformity in this interval, since Ne. parultimus, the index of the basal Pridoli, makes its first appearance less than 1 m above the last Sl. balticus. As in other Bohemian sections (Kříž et al. Reference Kříž, Jaeger, Paris and Schönlaub1986), Ne. parultimus is accompanied by Formosograptus formosus in the Všeradice section. In the East European Platform (Paškevičius, Reference Paškevičius1979; Urbanek & Teller, Reference Urbanek and Teller1997), Arctic Canada (Lenz & Kozłowska-Dawidziuk, Reference Lenz and Kozlowska–Dawidziuk2004) and Tien Shan (Koren’ & Sujarkova, Reference Koren’ and Sujarkova1997) Formosograptus formosus appears much lower and even gives its name to a combined biozone in the latest Ludfordian. Similarly, P. fragmentalis appears in Poland earlier; it co-occurs with Ps. latilobus, but disappeared already before the LA of Ps. latilobus.
4. Conclusions
Although late Ludfordian graptolites have rarely left a fossil record sufficient for detailed biostratigraphy and high-resolution correlation, a better understanding is of crucial importance as correlation of the Silurian System relies primarily on graptolite biozones. Benthic faunas and sequence boundaries can be used in correlations within individual basins, especially in stratigraphical intervals where conodont biozones lack the precision of those based on graptolites. Here, four graptolite biozones are distinguished in the latest Ludlow between the B. tenuis Biozone and the base of Pridoli Series. The Všeradice section provides the most complete graptolite record in the peri-Gondwanan realm. The late Ludlow graptolite biozonation previously applied in Bohemia and other parts of peri-Gondwanan Europe resulted rather from uncertain correlations between isolated partial sections.
The Kozlowskii extinction Event and subsequent recovery is recorded for the first time from both Bohemia and peri-Gondwana; this also represents the first record of the event in higher latitudes outside the tropical zone. The Všeradice section is unique also in its sedimentary record. Although situated in an offshore, open sea setting, it enables correlation with benthic and particularly conodont faunas, because it was situated close to a small shallow water area with carbonate sedimentation, which was colonized by benthic assemblages closely related to those inhabiting the carbonate platform situated to the north.
The Kozlowskii Event caused extinction of many graptolite lineages as previously suggested. Bohemograptids, neocucullograptids and Polonograptus all vanished from the fossil record. It is noteworthy that the pelagic orthocerids and nektonic Ceratiocaris passed through the extinction interval (Fig. 9). The pelagic ostracod Entomis, however, became extinct. Although a single taxon, its extinction is of note because it is a widespread, long-ranging generalist. Similarly, the conodont faunas in these deeper water settings were affected only slightly (see also Slavík, Kříž & Carls, Reference Slavík, Kříž and Carls2010). Data from the Všeradice section further support the earlier thoughts of Lehnert et al. (Reference Lehnert, Eriksson, Calner, Joachimski and Buggisch2007a) and Eriksson & Calner (Reference Calner and Ashraf2008) that the Kozlowskii extinction Event coincides with a drop in sea-level and period of icehouse climate.

Figure 9. Kozlowskii Event: graptolite crisis recorded by Bohemian sections. Note that all ventrally curved graptolites disappeared from the fossil record. Most species recorded for the first time from the Pr. fragmentalis Biozone suggest striking evolutionary links between Ludfordian and Pridoli graptolite faunas.
The interval between the graptolite extinction and subsequent recovery and radiation was relatively long (based on sedimentary rock thickness). Eroded shale and the intraformational conglomerate in the Všeradice section indicate a prominent gap in sedimentation during this time. The benthic faunas from the conglomerate (matrix and pebbles of different age), nevertheless, make correlation possible with coeval shallow water carbonate strata (Figs 8–10).

Figure 10. Correlation of late Ludlow graptolite and conodont biozones, bivalve communities and trilobite horizons in Bohemia with conodont biozones and formations in Gotland (including eustatic oscillation and climatic state as suggested by Lehnert et al. Reference Lehnert, Frýda, Buggisch, Munnecke, Nützel, Křiž and Manda2007b). Gaps in discussed Bohemian sections are indicated.
In terms of eustatic sea-level oscillations, the graptolite recovery recorded in the Prague Synform coincides with a period of rising sea-level after major eustatic drawdown. It was delayed far behind the recovery of benthic fauna. The graptolite recovery started after the end of the positive carbon isotope excursion and before the FAD of ‘Oz.’ snajdri. The post-Kozlowskii Event graptolite recovery gave origin to a novel graptolite fauna of Pridoli type. Some graptolite taxa that emerged or reappeared above the Kozlowskii Event crisis are likely ancestors to substantial elements of Pridoli graptolite assemblages (e.g. Sl. balticus gave rise to Sl. beatus (Koren’, Reference Koren’1983; Jaeger in Kříž et al. Reference Kříž, Jaeger, Paris and Schönlaub1986) and Ps. kosoviensis may have evolved into Ps. transgrediens). In turn, Ne. parultimus, although remarkably similar to Ps. latilobus, has been derived from another, less specialized ancestor, after Urbanek (Reference Urbanek1997).
The latest Ludlow F. formosus and/or Uncinatograptus acer, U. protospineus and U. spineus biozones, distinguished in Baltica and elsewhere in lower palaeolatitudes, are missing in Bohemia and indeed in the whole of peri-Gondwana. F. formosus, which makes its lowest occurrence in middle Ludfordian strata in Poland, Lithuania, Kyrgyzstan, Kazakhstan and Arctic Canada, is unknown from below the Ne. ultimus–Ne. parultimus Biozone of the basal Pridoli Series in Bohemia and there are some discrepancies in stratigraphical ranges of some other taxa (e.g. P. fragmentalis). Differences may result from temporary palaeobiogeographical separation rather than an incomplete fossil record in peri-Gondwana owing to a condensed or missing sedimentary rock record.
Precise correlation of late Ludlow conodont and graptolite biozones is not yet possible from published data. Consequently, the correlation of the graptolite Kozlowskii and conodont Lau events is still unknown in detail, although that the two events are approximately coincident is suggested from their position at the beginning of the positive carbon isotope excursion.
5. Systematic palaeontology (by Petr Štorch)
All graptolite taxa previously unknown under the correct name from Bohemia, new species and new combinations are described. Illustrated and described specimens are housed in the official collection of the Czech Geological Survey, Prague (prefix PŠ); type specimens prefixed L are housed in the National Museum, Prague.
Pseudomonoclimacis kosoviensis (Bouček, Reference Bouček1931) Figures 5i, 6g–j, 7s
1931 Monograptus kosoviensis n. sp. Bouček, pp. 2, 15, text-fig. 1c, d.
1936 Monograptus kosoviensis Bouček; Bouček, pp. 10–11, pl. 2, fig. 10; text-fig. 1g–k.
1943 Pristiograptus kosoviensis (Bouček); Přibyl, pp. 24–5, pl. 3, figs 14, 15.
1997 Pristiograptus dubius cf. frequens Jaekel; Rickards & Wright, pp. 219, 221–2, text-fig. 9i, j.
Material. Twenty-six flattened rhabdosomes from bed nos 1a, 1b, 3, 10 and 16 of the Všeradice section (see Fig. 2), several rhabdosomes from the middle part of trench VB below the present log and the type material figured by Bouček (Reference Bouček1931) and Přibyl (Reference Přibyl1943).
Lectotype. Designated by Bouček (Reference Bouček1936): specimen no L 19940 figured by Bouček (Reference Bouček1931, text-fig. 1d); from the level corresponding with the tenuis Biozone of the present biozonal scheme; Koledník Quarry near Beroun, Bohemia.
Description. The sicula is straight or gently ventrally curved, 1.2–1.7 mm long, its apex attaining a level at or below the aperture of th 2. The sicular aperture is concave, 0.22–0.28 mm wide, furnished with a short and slender virgella. The nema is robust, 0.4 mm wide in mature specimens, and extending as much as 30 mm beyond the distalmost thecae. The rhabdosome gradually widens from 0.45–0.6 mm at the apertural margin of th 1, through 0.7–0.8 mm at th 3, 0.85–1.1 mm at th 5, 1.2–1.7 mm at th 10 and 1.55–1.95 mm at th 20 to a maximum of 2.5–2.7 mm attained 40–50 mm from the sicula. Isolated distal fragments are 2.8 mm wide. Proximal thecae are slightly geniculate and overlap for one-third their length. More or less prominent hook-like secondary apertural additions are developed in proximal thecae of mature rhabdosomes with a more than 30–40 mm long thecate part (Fig. 6i). Details are not seen owing to inadequate preservation. The two-thecae repeat distance (2TRD) increases from 1–1.35 mm at th 2 and 1.1–1.45 mm at th 5 to 1.3–1.7 mm at th 10 and then gradually until 2.1–2.3 mm is attained most distally. Proximal thecae number 12–13 in 10 mm of rhabdosome length; long, simply tubular distal thecae number 9.5–8.5 in 10 mm and overlap for almost three-quarters of their length. Distal thecal apertures are perpendicular to the thecal axis or are slightly everted.
Remarks. Our specimens agree in all dimensions with Bouček's (Reference Bouček1931) type material, especially in their distal dorso-ventral width and thecal spacing. The sicular part of the rhabdosome is described for the first time and shows that this species is similar to Pseudomonoclimacis contumax (Bouček, Reference Bouček1936) and ‘Monograptus’ sp. of Rickards & Palmer (Reference Rickards and Palmer1977). The apertural overgrowths observed in our mature rhabdosomes of Ps. kosoviensis (Bouček, Reference Bouček1931) are equally developed in ‘Monograptus’ sp. (of Rickards & Palmer, Reference Rickards and Palmer1977), which may be distinguished solely by its larger, up to 2.5 mm long sicula attaining a level above the aperture of th 2. Pseudomonoclimacis contumax differs from stratigraphically younger Ps. kosoviensis in having a less robust rhabdosome with a straight to slightly dorsally curved proximal part and more closely spaced thecae. Small apertural outgrowths have been found in one mature specimen of Ps. contumax.
Jaeger (Reference Jaeger1978) demonstrated that the hook-like apertural structures in ‘Monograptus’ sp. of Rickards & Palmer (Reference Rickards and Palmer1977) are secondary structures formed during the late astogenetic or gerontic stage of colony development. He also reported similar secondary outgrowths in Colonograptus colonus, Saetograptus chimaera and Monoclimacis haupti. The tendency to thecal isolation and development of lateral apertural lobes, however, may represent, at least in part, natural variability rather than gerontic growth because of its occurrence in immature rhabdosomes.
Although the resemblance of the proximal and mesial thecae of Ps. kosoviensis to Monoclimacis thecae is commonly enhanced by flattening, the distinct thecal geniculation, hook-like apertural growth additions in the proximal thecae of mature rhabdosomes, very rigid nema and proximal part of the rhabdosome lacking ventral curvature clearly differentiate these graptolites from the Pristiograptus dubius group, including Pristiograptus frequens Jaekel figured by Jaeger (Reference Jaeger1991, pl. 25, figs 9, 11, 15). M. kosoviensis Bouček and related taxa are best assignable to the genus Pseudomonoclimacis (Mikhaylova, Reference Mikhaylova, Kaljo and Koren’1976) sensu Koren’ (Reference Koren’, Nikitin and Bandaletov1986), Urbanek (Reference Urbanek1997) and Lenz & Kozłowska-Dawidziuk (Reference Lenz and Kozlowska–Dawidziuk2004).
Pseudomonoclimacis aff. kosoviensis (Bouček, Reference Bouček1931) Figures 6l, 7r
Remarks. Nine rhabdosomes found in the fragmentalis Biozone can be distinguished from typical Ps. kosoviensis by their relatively robust proximal part and lesser distal maximum width (2.1 mm). Thecae are equally overlapping and exhibit the same tendency to develop hook-like apertural outgrowths proximally. Thecae are wider and slightly more widely spaced (2TRD th 10 = 2.1 mm). This stratigraphically distinctive form is left in open nomenclature referring to its close similarity to Ps. kosoviensis until more material is available.
Pseudomonoclimacis latilobus (Tsegelnjuk, Reference Tsegelnjuk1976) Figures 5d, e, j, r, s, 7q
1976 Colonograptus latilobus n. sp. Tsegelnjuk, p. 109, pl. 29, figs 9–11.
1983 Saetograptus (Colonograptus) insignitus n. sp. Přibyl, p. 160, pl. 1, figs 3–8; pl. 4, figs 11–13.
1988 Ludensograptus latilobus (Tsegelnjuk); Tsegelnjuk, p. 82, text-fig. 3.
1988 Monograptus latilobus (Tsegelnjuk); Koren’, Lytochkin & Rinenberg, p. 17, text-figs 9–10.
1997 Pseudomonoclimacis latilobus (Tsegelnjuk); Urbanek, pp. 161–5, pls 17–19; text-figs 45–47.
1997 Monograptus latilobus (Tsegelnjuk); Koren’ & Sujarkova, pp. 75–7, pl. 4, figs 3–4; text-fig. 11g–n.
Material. Twenty-two flattened rhabdosomes from bed nos 14, 16 and 20 of the Všeradice section (see Fig. 2) and several rhabdosomes from Velký Vrch near Koněprusy, including topotypical specimens of ‘S. insignitus’ collected by Alois Přibyl.
Holotype. Specimen no N 1788/11 figured by Tsegelnjuk (Reference Tsegelnjuk1976) on plate 29 as figure 11; from the ‘?leintwardinensis Biozone’ of the Gushcha borehole, Volynia, Ukraine.
Description. The dorsal wall of the medium-sized, 20–30 mm long rhabdosome is straight or slightly ventrally curved in the proximal part. The sicula is also ventrally curved. It is c. 2.0 mm long, including a rarely recognizable slender prosicular part. The readily visible metasicular portion is 1.4–1.5 mm long. Sicular aperture is 0.4 mm wide in profile, deeply concave, furnished with a c. 0.1 mm long dorsal lobe. The first theca initiates 0.2–0.3 mm above the sicular aperture. Th 1 is roughly triangular in outline, with a slightly everted aperture provided with more or less developed, ventrally directed, and paired lateral lobes. Distinct ventro-lateral apertural lobes are present on each theca throughout the rhabdosome. Thecae appear slightly geniculated owing to a gentle excavation on the ventral thecal wall. Flattened thecae exhibit somewhat isolated and introverted apertures with the lateral lobe separated by a notch from the ventral wall of the next theca. The ventral part of the apertural margin is either acuminate, when preserved in profile, or lobate, when obliquely flattened. Distal thecae are 2.1–2.4 mm long, straight and inclined at an angle of 40–50° to the rhabdosome. The rhabdosome is 0.85–0.95 mm wide at the th 1 aperture, 1.15–1.3 mm wide at th 3, 1.3–1.5 mm wide at th 5 and 1.6–1.8 mm wide at th 10. Maximum distal width of the rhabdosome, 2–2.1 mm, is attained by approximately the 14th theca. The 2TRD increases from 1.3–1.45 mm at th 2, to 1.5–1.7 mm at th 10 and then to the distal maximum of 1.75–1.95 mm, i.e. the distal thecae number 10.5–11.5 in 10 mm of rhabdosome length.
Remarks: The present material matches the typical specimens illustrated by Tsegelnjuk (Reference Tsegelnjuk1976, Reference Tsegelnjuk1978) and Urbanek (Reference Urbanek1997). A morphologically less-advanced and stratigraphically younger morphotype, described and illustrated by Urbanek (Reference Urbanek1997, pl. 19) is similar to Pseudomonoclimacis fecundus (Přibyl, Reference Přibyl1943), which can be distinguished by its poorly developed apertural lobes and less robust proximal-most part of the rhabdosome.
Polonograptus chlupaci sp. nov. Figures 5o, 7o, 11a, b
1995a Polonograptus egregius (Urbanek); Štorch, p. 74–5, pl. 4, figs 1–5; text-figs 4d, 6f, i, j, l.
Name. After the late Prof. Ivo Chlupáč, distinguished Czech stratigrapher and palaeontologist.
Material. Eight complete and four incomplete flattened rhabdosomes and several fragments from Kosov Quarry and two incomplete rhabdosomes from the Všeradice section.
Holotype. Specimen no PŠ 631, figured by Štorch (Reference Štorch1995a, pl. 4, fig. 3; text-fig. 6i) and refigured herein in Figure 11a, from the middle Ludfordian Nc. kozlowskii Biozone at Kosov Quarry near Beroun, Bohemia.
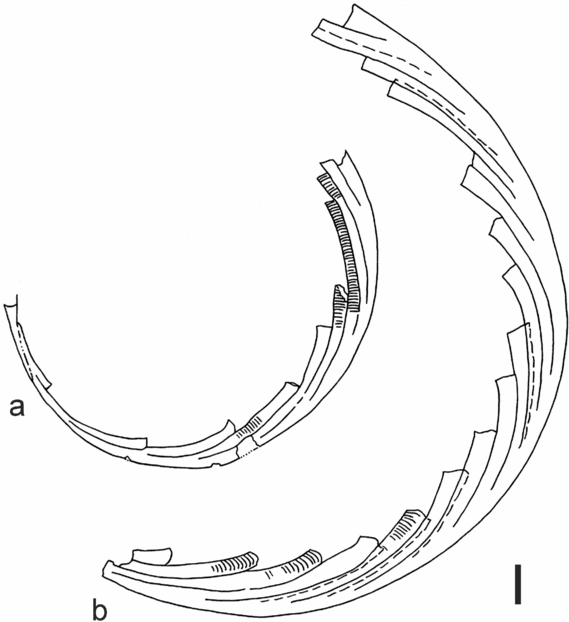
Figure 11. Type material of Polonograptus chlupaci sp. nov. (a) holotype PŠ 631; (b) paratype PŠ 667/1; Nc. kozlowskii Biozone, Kosov Quarry, × 5. Scale bar represents 1 mm.
Diagnosis. Rhabdosome regularly ventrally coiled, almost circular. Sicula 1.3–1.7 mm long; the apex reaches about two-thirds up the 1.2–1.7 mm long th 1. Relatively short th 1 followed by considerably longer (2.4–3.6 mm) th 2. Thecae slender tubes terminated by simple apertures with gentle lateral apertural elevations; 4.4–4.9 mm long distal thecae overlap for about two-thirds their length.
Discussion. This unusual taxon was described by Štorch (Reference Štorch1995a) from the kozlowskii Biozone of the Kosov Quarry as Polonograptus egregius (Urbanek, Reference Urbanek1970). Two almost circular rhabdosomes encountered from the same level in the Všeradice section exhibit the same parameters medially and distally whereas their proximal parts are broken. The specimen shown in Figures 5o and 7o seems to have an incomplete th 1 and the sicula and initial bud are missing.
Urbanek (Reference Urbanek1970) erected ‘Monograptus’ egregius based upon fragmentary material from the kozlowskii Biozone of the Mielnik Borehole in eastern Poland. The holotype is a distal fragment comprising four thecae and the paratype is a sicula with a very incomplete first theca. Urbanek (Reference Urbanek1970, pl. 42, fig. f) published a hypothetical complete development of the first theca based on isolated fragments, but subsequently this ‘reconstruction’ was used as the most essential part in the generic diagnosis of Egregiograptus Rickards & Wright, Reference Rickards and Wright1997. Koren’ & Sujarkova (Reference Koren’ and Sujarkova2004) encountered a rich and moderately well-preserved egregiograptid fauna in Tien Shan (Kyrgyzstan) and considered the extremely long th 1 a principal diagnostic feature of Egregiograptus. Accordingly, Koren’ & Sujarkova referred Bohemian specimens with overall egregiograptid morphology combined with a short th 1 to a new, so far undescribed species of Polonograptus. This form, which is named Polonograptus chlupaci sp. nov. in this paper, can be barely distinguished from egregiograptids when its proximal part with its short th 1 and long th 2 is broken, since the mesial and distal parts of its rhabdosome match Egregiograptus rinellae Koren & Sujarkova, Reference Koren’ and Sujarkova2004, or Egregiograptus egregius Urbanek as illustrated by Tomcyzk, Urbanek & Teller (Reference Tomczyk, Urbanek, Teller and Pachlowa1990). The most similar polonograptid, Polonograptus aloisi Koren’ & Sujarkova, Reference Koren’ and Sujarkova1997, differs from P. chlupaci in having an arcuately curved and more rapidly widening rhabdosome with strongly inclined, shorter and less-overlapping thecae.
Slovinograptus balticus (Teller, Reference Teller1966) Figures 5b, f, g, k–n, 7l, p
1966 Monograptus balticus n. sp. Teller, pp. 556–8, pl. 1, figs 6–11; text-fig. 4a, b.
1974 ‘Monograptus’ balticus Teller; Paškevičius, p. 126, pl. 15, figs 1–5; pl. 19, figs 4–11; pl. 20, figs 1–2.
1976 Wolynograptus balticus (Teller); Tsegelnjuk, pp. 111–12, pl. 33, fig. 5.
1979 Monograptus balticus Teller; Paškevičius, pp. 168–9, pl. 12, figs 4–8; pl. 28, figs 1–12.
1983 ‘Monograptus’ balticus Teller; Tsegelnjuk, pl. 145, fig. 34.
1983 Monograptus (Wolynograptus) abhorrens sp. nov.; Přibyl, pp. 159–60, pl. 2, figs 5–9; pl. 4, figs 5–9.
1988 Monograptus balticus Teller; Koren’, Lytochkin & Rinenberg, p. 17, fig. 1.
1997 Monograptus balticus Teller; Koren’ & Sujarkova, p. 71, pl. 1, figs 9–10; text-fig. 9e–h, (?a).
Material. Twenty rhabdosomes; mostly flattened. Several specimens are pyritized, either with periderm or preserved as internal moulds. An additional four flattened specimens originate from Velký Vrch (type locality of ‘Monograptus (Wolynograptus) abhorrens Přibyl).
Holotype. The specimen figured by Teller (Reference Teller1966, pl. 1, fig. 6) and refigured by Koren’ & Sujarkova (Reference Koren’ and Sujarkova1997, text-fig. 9g) from the upper Ludfordian of the Łeba 3 borehole in northern Poland.
Description. The otherwise straight rhabdosome is slightly dorsally curved in its slender proximal part. The sicula is 1.35–1.45 mm long, and gently ventrally recurved. Its apex reaches approximately two-thirds up th 1. The sicular aperture is concave in outline, 0.18–0.22 mm wide, furnished with a weakly developed dorsal lobe and short virgella. Slender, 1.25–1.45 mm long th 1 diverges at about 5° from the rhabdosome axis with its origin 0.25–0.4 mm above the sicular aperture. Thecal aperture is covered by tightly appressed, transversely extended hood. The succeeding thecae are slender, very gently inclined, straight or gently S-shaped tubes with prominent apertural hoods that may be responsible for the common subscalariform mode of rhabdosome preservation. A thecal length of 2.25 mm was measured in th 10, which overlaps for one-half of the succeeding theca (see also specimen in Fig. 5n). The rhabdosome widens gradually from 0.45 mm at the level of the first thecal aperture, to 0.6–0.7 mm at th 3, and 0.7–0.85 mm at th 5. A width of 0.85–1.2 mm is attained by about th 10 and is maintained in the distalmost parts of the present, still not fully mature rhabdosomes. The 2TRD is 1.7–2.0 mm at th 2, and 1.7–2.4 mm more distally.
Discussion. Tsegelnjuk (Reference Tsegelnjuk1976) assigned Monograptus balticus Teller, Reference Teller1966 and another three species to his new genus Wolynograptus. He chose Wolynograptus valleculosus Tsegelnjuk, Reference Tsegelnjuk1976 as the type species. This choice turned out to be unfortunate since W. valleculosus differs from other species included in this genus in having hooked metathecae without characteristic tightly appressed, globular and transversely extended apertural hoods. Its rhabdosome and described thecal morphology resemble Uncinatograptus acer (Tsegelnjuk, Reference Tsegelnjuk1976) instead. Urbanek (Reference Urbanek1997) included slender wedge-shaped Ludfordian monograptids with thecal apertures covered by tightly appressed hoods in a new subgenus Slovinograptus. Morphological distinctiveness of its sigmoidally curved thecae and close affinities to M. beatus Koren’ and M. microdon R. Richter (see Jaeger in Kříž et al. Reference Kříž, Jaeger, Paris and Schönlaub1986) lead us to promote Slovinograptus to genus level. A successive series leading from Ludfordian Sl. balticus (Teller) through Sl. beatus (Koren’) of Pridoli to Lochkovian Sl. microdon (R. Richter) has been suggested by Koren’ (Reference Koren’1983).
Flattened specimens of Slovinograptus balticus figured by Teller (Reference Teller1966) and Paškevičius (Reference Paškevičius and Obut1974, Reference Paškevičius1979) differ from the present material in their more abrupt widening within the first 5–7 thecae, when measured without the apertural hoods. This apparent difference may be explained by the largely box-like mode of flattening of Polish and Lithuanian specimens, as suggested by pressed-through interthecal septa. The Bohemian specimens, by contrast, have been partly pyritized in calcareous shale and argillitic limestone. Specimen PŠ 2015 (Fig. 5n) is a pyritized internal mould preserved in relief. It is obvious that the dorsal rhabdosome curvature, missing in the specimens of ‘Monograptus abhorrens Přibyl, Reference Přibyl1983’ and specimens of Sl. balticus studied by Urbanek (Reference Urbanek1997, pl. 3, figs 2, 4, 5) is partly due to natural intraspecific variation and, in part, due to the subscalariform orientation of the rhabdosomes. No Bohemian specimen exhibits such pronounced and ventrally extended apertural hoods as does the rhabdosome figured by Koren’ & Sujarkova (Reference Koren’ and Sujarkova1997, text-fig. 9A). Sl. balticus either varies in its rhabdosomal curvature and apertural hood development or still comprises several palaeogeographically isolated populations of so far unrecognized taxonomic status. The type material of ‘Monograptus abhorrens’ has been missing for at least 20 years. Owing to the considerable morphological variability caused by different preservation, observed already within our material from the Všeradice section, we have retained all material in a single species.
Slovinograptus cf. balticus (Teller, Reference Teller1966) Figures 6e, 7f
Remarks. This form is slender, gently S-shaped or almost straight except for a slightly dorsally curved proximal part comprising 2–3 initial thecae. The dorso-ventral width increases from 0.45 mm at th1, through 0.6 mm at th 3 and 0.75 mm at th 5, to 0.85 mm at th 10. The 2TRD of 1.9–2.1 mm is constant throughout the rhabdosome.
Sl. beatus (Koren’, Reference Koren’1983) can be differentiated by its wider, more robust proximal part. Sl. balticus is distinguishable by its ventrally curved to almost straight, more rapidly widening proximal part and by its interthecal septa, which are inclined to the rhabdosome axis. Eleven rhabdosomes of Slovinograptus cf. balticus from the upper fragmentalis and parultimus–ultimus biozones in the Všeradice section help to fill a stratigraphical gap between the late Ludfordian slovinograptids (Sl. hamulosus and Sl. balticus) and forms ranging from the late Pridoli (Sl. beatus (Koren’), Sl. aksajensis (Koren’)) through to the earliest Devonian M. uniformis Biozone (Sl. microdon (R. Richter)).
The narrow wedge-shaped rhabdosomes, ventral thecal walls parallel to the rhabdosome axis or sloping slightly inwards, and dorsal thecal walls terminated by an apertural hood transversely extended into a tabular or semicircular roof make this group of graptolites fairly distinctive.
Crinitograptus? sp. Figures 6f, 7h
Remarks. About 20 dorsally arcuate, slender rhabdosomes of nearly uniform width were collected from a restricted interval of the uppermost fragmentalis and lowermost parultimus–ultimus biozones. Material is poorly preserved but obviously represents a distinctive form previously unknown from Bohemian sections. It is readily recognizable by its prominent apertural hoods combined with long and slender prothecae with ventral wall parallel to the rhabdosome. Short metathecae terminate with ventrally to proximo-ventrally directed apertural hoods which, in some specimens, appear to be transversely extended. Dorso-ventral width is 0.4–0.5 mm across the parallel-sided part of the theca and c. 0.85 mm across the apertural hood. The 2TRD is 2.3–2.55 mm in the majority of our specimens that are, however, either incomplete or just fragments. The proximal end is barely recognizable and the sicula is not preserved.
A similar taxon, designated as Crinitograptus cf. operculatus (Münch), has been encountered by Lenz & Kozłowska-Dawidziuk (Reference Lenz and Kozlowska–Dawidziuk2004) from the lowermost Pridoli of the Arctic Islands of Canada. The Canadian specimens are broader (0.6–0.7 mm without hoods and 1.0–1.2 mm across the hoods) and their thecae are more widely spaced (3 thecae in 5 mm). Until better specimens are available, the Všeradice material has also been tentatively assigned to Crinitograptus owing to the nearly uniform rhabdosome width, slender thecae with ventral walls parallel to the rhabdosome axis and apertural hoods. The hooded apertural structure and prothecae parallel to the rhabdosome, however, suggest that this taxon may have evolved from Slovinograptus instead.
Acknowledgements
The authors greatly appreciate the financial support provided by the Czech Science Foundation through projects 205/09/0703 and 210/10/2351. Many thanks to David Loydell and Michael J. Melchin for valuable comments on the manuscript and improving the English, and Jan Kolda for his field assistance and help with permissions to excavate. Petr Budil kindly identified some trilobites.












