


1. Introduction
Since the first discovery of its isolated sclerites (Matthew, Reference Matthew1899), Wiwaxia has established itself among the iconic Burgess Shale problematica. Until recently, its affinity was considered unresolvable (Eibye-Jacobsen, Reference Eibye-Jacobsen2004); its interpretation as a mollusc – whether at a crown-group or stem-group level (Scheltema & Ivanov, Reference Scheltema and Ivanov2002; Scheltema, Kerth & Kuzirian, Reference Scheltema, Kerth and Kuzirian2003; Caron et al. Reference Caron, Scheltema, Schander and Rudkin2006, Reference Caron, Scheltema, Schander and Rudkin2007; Conway Morris & Caron, Reference Conway Morris and Caron2007) – had competed with alternative positions in the annelid crown group or stem group (Walcott, Reference Walcott1911; Butterfield, Reference Butterfield1990, Reference Butterfield2003, Reference Butterfield2006, Reference Butterfield2008; Conway Morris & Peel, Reference Conway Morris and Peel1995), or in the stem group to Annelida + Brachiopoda (Conway Morris & Caron, Reference Conway Morris and Caron2007). Membership of an extinct phylum with remote affiliation to the molluscs had also been proposed (Bengtson & Conway Morris, Reference Bengtson and Conway Morris1984; Conway Morris, Reference Conway Morris1985). A position among Mollusca was finally established through the identification of radula-like mouthparts, a creeping ventral foot and an aculiferan-like scleritome, even if the exact affinity of Wiwaxia within the molluscs remains unresolved (Smith, Reference Smith2012, Reference Smith2014).
Despite the phylogenetic uncertainty, the morphology of Wiwaxia's dorsal imbricating scleritome is well constrained by hundreds of articulated specimens from the Burgess Shale (Walcott, Reference Walcott1911; Conway Morris, Reference Conway Morris1985; Smith, Reference Smith2014), complemented by a handful of further individuals from Utah (Conway Morris et al. in press) and two localities in South China (Sun et al. Reference Sun, Zhao, Peng and Yang2014; Yang et al. Reference Yang, Smith, Lan, Hou and Zhang2014). The Wiwaxia scleritome comprised eight rows of overlapping scale-like sclerites, punctuated in adults by two rows of elongate dorsal spines. Sclerites are arranged in four distinct zones on either side of the body (ventral, lower lateral, upper lateral and dorsal), with a distinct zone of sclerites arming the anterior of the body. Leaf-like, the sclerites comprise a smooth proximal root that opens out into an oval, siculate or spinose blade; this blade has a reinforced margin and edge-parallel ribs, and sometimes bears a pustular ornament (Butterfield & Harvey, Reference Butterfield and Harvey2012; Smith, Reference Smith2014). Elongate internal chambers indicate that they were secreted by microvilli, suggesting a chitinous original composition (Butterfield, Reference Butterfield1990; Smith, Reference Smith2014). This recalcitrant carbonaceous construction gives the sclerites a strong preservation potential in suitable taphonomic regimes, either as Burgess Shale-type macrofossils or as robust acid-extracted microfossils (Small Carbonaceous Fossils, see Butterfield & Harvey, Reference Butterfield and Harvey2012).
The distinctive shape and robust constitution of Wiwaxia sclerites has led to their recognition in a worldwide suite of deposits representing platform to outer shelf environments from Cambrian Series 2 to 3 (Fig. 1; Conway Morris, Reference Conway Morris1985; Butterfield, Reference Butterfield1994; Zhao, Qian & Li, Reference Zhao, Qian and Li1994; Ivantsov, Zhuravlev & Leguta, Reference Ivantsov, Zhuravlev and Leguta2005; Fatka, Kraft & Szabad, Reference Fatka, Kraft and Szabad2011; Sun et al. Reference Sun, Zhao, Peng and Yang2014; Yang et al. Reference Yang, Smith, Lan, Hou and Zhang2014). Thus, although articulated specimens are only known from four sites (Conway Morris Reference Conway Morris1985; Zhao, Qian & Li, Reference Zhao, Qian and Li1994; Sun et al. Reference Sun, Zhao, Peng and Yang2014; Yang et al. Reference Yang, Smith, Lan, Hou and Zhang2014; Conway Morris et al. in press), isolated sclerites extend the range of Wiwaxia to North America (Mount Cap, Stephen, Earlie, Pika and Spence formations; Matthew, Reference Matthew1899; Walcott, Reference Walcott1911; Conway Morris, Reference Conway Morris1985; Conway Morris & Robison, Reference Conway Morris and Robison1988; Butterfield, Reference Butterfield1994; Harvey & Butterfield, Reference Harvey and Butterfield2011; Butterfield & Harvey, Reference Butterfield and Harvey2012), South China (Tsinghsutung and Kaili formations; Zhao, Qian & Li, Reference Zhao, Qian and Li1994, Zhao et al. Reference Zhao, Yuan, Zhu, Guo, Zhou, Yang and Van Iten1999; Harvey et al. Reference Harvey, Ortega-Hernández, Lin, Zhao and Butterfield2012; Sun et al. Reference Sun, Zhao, Peng and Yang2014), Australia (Emu Bay Shale and Monastery Creek Formation; Porter, Reference Porter2004), Siberia (Sinsk Formation; Ivantsov, Zhuravlev & Leguta, Reference Ivantsov, Zhuravlev and Leguta2005) and the Czech Republic (Buchava Formation; Fatka, Kraft & Szabad, Reference Fatka, Kraft and Szabad2011). Nevertheless, Wiwaxia has not been reported from Chengjiang, even though this is among the richest and best sampled Cambrian Lagerstätten (Zhao et al. Reference Zhao, Hu, Caron, Zhu, Yin and Lu2012, Reference Zhao, Caron, Bottjer, Hu, Yin and Zhu2014) and has a broadly equivalent ecological and taxonomic composition to the Burgess Shale biota, where Wiwaxia is known from hundreds of specimens (Conway Morris, Reference Conway Morris1985; Smith, Reference Smith2014). Here, we fill this surprising gap in Wiwaxia's distribution with a report of associated sclerites from Chengjiang.

Figure 1. Distribution of Wiwaxia. (a) Stratigraphic distribution. (b) Palaeogeographic distribution (modified after McKerrow, Scotese & Brasier, Reference McKerrow, Scotese and Brasier1992; Álvaro et al. Reference Álvaro, Elicki, Geyer, Rushton and Shergold2003; Fatka, Kraft & Szabad, Reference Fatka, Kraft and Szabad2011). Numbers in (a) correspond to those plotted in (b).
2. Materials and methods
Our material was collected from the mudstone-dominated Maotianshan Shale Member of the Yu’anshan Formation, Cambrian Series 2, Stage 3, Eoredlichia–Wudingaspis Zone (Chen et al. Reference Chen, Zhou, Zhu and Yeh1996; Zhu et al. Reference Zhu, Li, Zhang, Steiner, Qian and Jiang2001), at the Mafang section, Haikou, Yunnan, SW China (24°46′20″N, 102°35′10″E). Detailed locality information and fossil taphofacies were presented by Zhao et al. (Reference Zhao, Caron, Hu and Zhu2009). The material was prepared using a blade and is deposited in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, China (NIGPAS). Photographs were taken under dark field illumination using a Zeiss Discovery V20 microscope system.
3. Description
The material consists of part and counterpart of a single slab of fine-grained mudstone (NIGPAS160801a/b) preserving compressions of four aligned bundles of sclerites on a single bedding surface (Fig. 2). It exhibits characteristic Chengjiang-type preservation (Zhu, Babcock & Steiner, Reference Zhu, Babcock and Steiner2005); its originally carbonaceous components are reflected by weathered aluminosilicate films associated with superficial iron oxides.
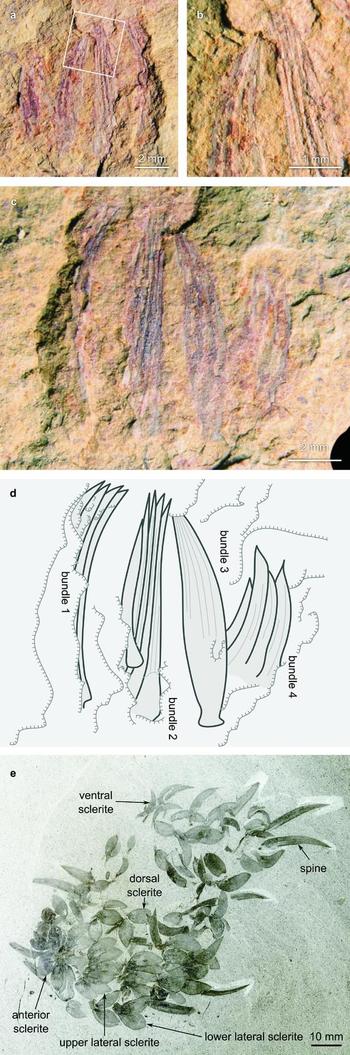
Figure 2. Sclerites of Wiwaxia. (a–d) from the lower Cambrian (Series 2, Stage 3) Maotianshan Shale Member, Yu’anshan Formation (Eoredlichia–Wudingaspis Zone) in the Mafang section, Haikou near Kunming, Yunnan Province; (e) from the middle Cambrian (Series 3, Stage 5) Burgess Shale. (a) Part (NIGPAS160801a). (b) Enlargement of boxed area, showing tips of constituent sclerites in bundle 2. (c) Counterpart (NIGPAS160801b). (d) Interpretative sketch of counterpart. (e) Semi-articulated scleritome from the Burgess Shale (Royal Ontario Museum 56965) illustrating variation in morphology between sclerite zones and splayed habit of upper lateral sclerite zone.
The sclerite bundles contain one to four sclerites of similar size, around 8mm long and 0.8–1.6mm wide; sclerites within a bundle are closely aligned such that their tips and bases are almost adjacent. Sclerites appear to vary in curvature and width between bundles, with the aspect ratio varying from 6:1 to approximately 10:1. This apparent variation may reflect differences in the obliquity of the sclerites’ burial angle, or result from post-mortem deformation. Each sclerite proceeds from a narrow root to a gently curved blade that tapers to a pointed apex with an angle of ~ 20°. Longitudinal ribs adorn the blade, although these are difficult to distinguish from sclerite margins. Individual sclerites can only be discriminated by identifying their tips: four sclerites are evident in the first bundle, five in the second, one in the third and four in the fourth. These counts may represent underestimates; sclerites with occluded tips cannot be identified, and if additional sclerites exist, their margins are difficult to distinguish from ribs.
The identity of this material as Wiwaxia sclerites is indicated by the sclerites’ outlines, longitudinal ribbing, smooth root and limited three-dimensional relief (indicating an originally robust carbonaceous composition). Their close association indicates that they were linked by connective tissue (presumably decayed) when they were buried.
The parallel preservation of multiple bundles recalls a transverse row of sclerites (cf. Smith, Reference Smith2014, fig. 5B). Under this interpretation, the four bundles correspond to the four zones of body sclerites. However, the absence of the fan-like splaying typically associated with non-ventral sclerites (e.g. Fig. 2e) militates against this interpretation, as does the extreme difference between the length:width ratios of the second and third bundles of sclerites (c. 8) and the smaller values in the non-ventral sclerites of W. corrugata (1.8±0.5) and W. foliosa (1.82±0.3) (Fig. 3a). (Detailed measurements from W. herka are not available.)

Figure 3. Comparison of sclerites from Chengjiang with those of other Wiwaxia species. Aspect ratio of sclerites in first (1), second (2), third (3) and fourth (4) sclerite bundles in NIGPAS160801 compared to non-ventral (panel a) and ventral (panel b) sclerites of W. corrugata (c, blue) and W. foliosa (f, orange). Width measurements from NIGPAS160801 may represent underestimates owing to oblique preservation (see text).
More promisingly, the fossil might correspond to iterated bundles of ventral sclerites. This interpretation accounts for the curvature present in all sclerites, the regular spacing of the four bundles and each bundle's limited splaying. The length:width ratio of each bundle fits the profile of the ventral sclerites of W. corrugata and (assuming a similar trend with increasing sclerite size) W. foliosa (Fig. 3b), the latter species being only currently known from diminutive (juvenile?) specimens. (Owing to oblique preservation, particularly in the first two sclerite bundles, measurements of sclerite width are likely underestimates.) We therefore interpret our material as an articulated series of ventral Wiwaxia sclerites of indeterminate species.
4. Discussion
This is the first report of Wiwaxia from the Chengjiang biota, representing the earliest record of the taxon and further expanding its broad geographical range. Its apparent absence from the Chengjiang deposits was a long-standing puzzle, particularly given its prominence in multiple Burgess Shale-type biotas throughout the Cambrian period and the relatively high preservation potential of Wiwaxia sclerites in Burgess Shale-type biotas (Butterfield & Harvey, Reference Butterfield and Harvey2012). Evolution, biostratigraphy and ecology had been cited to account for its absence (Butterfield, Reference Butterfield2003). We can now show that Wiwaxia had evolved and was present in the Chengjiang region by the Cambrian Stage 3, adding it to a growing roster of species that were once considered endemic but are now recognized in both Lagerstätten: taxa such as Primicaris and Misszhouia, previously unique to Chengjiang, have recently been described in the Burgess Shale (Caron et al. Reference Caron, Gaines, Aria, Mángano and Streng2014), whereas taxa such as Nectocaris have been identified in Chengjiang after being considered unique to the Burgess Shale (Smith, Reference Smith2013).
Wiwaxia nevertheless remains a rare component of the Chengjiang fauna – particularly in view of the c. 500 articulated specimens known from the Burgess Shale (Smith, Reference Smith2014). Ecology and taphonomy account for this discrepancy; the Chengjiang deposits represent the relatively shallow-water settings of the continental platform (Zhang, Liu & Zhao, Reference Zhang, Liu and Zhao2008), whereas the majority of Burgess Shale sites sample a basinal setting representing deeper waters off the platform edge (Caron et al. Reference Caron, Gaines, Mángano, Streng and Daley2010, Reference Caron, Gaines, Aria, Mángano and Streng2014). What is more, substantial differences characterize contemporary Burgess Shale communities that are separated by trivial spatial and stratigraphic distances (Caron et al. Reference Caron, Gaines, Mángano, Streng and Daley2010, Reference Caron, Gaines, Aria, Mángano and Streng2014), emphasizing the prominent role of local-scale factors in controlling community composition. As such, Wiwaxia represents a striking example of faunal continuity between these two Lagerstätten, despite distinct environmental settings, a time difference of some 15 million years and substantial geographic separation.
Acknowledgements
Research was supported by the Chinese Academy of Sciences (KZZD-EW-02–2); the National Basic Research Programme of China (2013CB835006); the National Natural Science Foundation of China (41472012, J1210006); the Natural Science Foundation of Jiangsu Province (BK2012893); and Clare College, Cambridge. We thank the anonymous reviewers for their constructive comments and improving the manuscript.



