



1. Introduction
Recent work on early paradoxidids has resulted in the description of several new species from the Anti-Atlas (Morocco), all belonging to the genus Acadoparadoxides (Geyer & Vincent, Reference Geyer and Vincent2015). Those species together with some taxa from other localities (Spain, Turkey, Newfoundland) form a consistent group (sometimes referred to as the Acadoparadoxides mureroensis clade (see e.g. Geyer & Vincent, Reference Geyer and Vincent2015), to be verified in the future with phylogenetics). Until now, this group has never been reported from palaeogeographic areas outside West Gondwana and Avalonia. The Holy Cross Mountains (central Poland), especially their southern region, represent part of the Małopolska Block, a terrane usually reconstructed as being an element of the Baltica palaeocontinent, but with an unclear provenance and a widely disputed age of accretion (e.g. Nawrocki et al. Reference Nawrocki, Dunlap, Pecskay, Krzemiński, Żylińska, Fanning, Kozłowski, Salwa, Szczepanik and Trela2007; Walczak & Belka, Reference Walczak and Belka2017). Previous descriptions of paradoxidids from this area included several species, among which four represented taxa typical of Baltica. This paper provides a revision of this material and shows for the first time the presence of the earliest group of acadoparadoxidines outside West Gondwana coupled with the lack of Scandinavian trilobite taxa. Such revision was possible due to the application of morphometric techniques (geometric morphometrics), a very useful tool becoming increasingly popular in modern palaeontology. This approach included a new application of retrodeformation techniques (since the material from Poland is significantly distorted) that comprises the adaptation of methods used in classical morphometry (e.g. Hughes & Jell, Reference Hughes and Jell1992; Żylińska, Kin & Nowicki, Reference Żylińska, Kin and Nowicki2013) to landmark data.
2. Geological setting and review of the trilobite assemblages
The analysed material was collected in the central and eastern part of the Holy Cross Mountains. This hilly area located within the Trans-European Suture Zone is subdivided into the Kielce and Łysogóry regions, separated by the Holy Cross Fault (Fig. 1). The main outcrops (including Brzechów and Jugoszów–Usarzów) of the traditional ‘Middle’ Cambrian are situated in the southern Kielce Region, which represents an exposed part of the sedimentary cover of the Małopolska Block. During the Cambrian, this crustal element was either a small terrane believed to be the proximal part of the Baltica palaeocontinent (e.g. Nawrocki et al. Reference Nawrocki, Dunlap, Pecskay, Krzemiński, Żylińska, Fanning, Kozłowski, Salwa, Szczepanik and Trela2007) or an exotic terrane rifted from Gondwana and accreted to Baltica during the Cambrian (e.g. Belka et al. Reference Belka, Valverde-Vaquero, Dörr, Ahrendt, Wemmer, Franke and Schäfer2002).

Figure 1. Geological sketch map of the Holy Cross Mountains (HCM) showing the distribution of Cambrian deposits, modified from Orłowski (Reference Orłowski1975, Reference Orłowski1992), with location of the studied outcrops. EEC – East European Craton; TTL – Teisseyre–Tornquist Line.
The Cambrian succession of the Holy Cross Mountains is developed in siliciclastic facies. The analysed material comes from two lithostratigraphic formations: the Słowiec Sandstones and the Usarzów Sandstones (Orłowski, Reference Orłowski1975). Both formations were deposited in a shallow marine environment; for instance, a typical occurrence of the Słowiec Formation bears evidence of a relatively high-energy setting, whereas the lower part of the unit, which occurs in Brzechów, was deposited in calmer conditions (e.g. Kowalczewski, Żylińska & Szczepanik, Reference Kowalczewski, Żylińska and Szczepanik2006). The Usarzów Sandstones developed in a rather low-energy environment (e.g. Kowalczewski, Żylińska & Szczepanik, Reference Kowalczewski, Żylińska and Szczepanik2006).
The first locality with paradoxidid remains, situated near Brzechów village, lies in the western part of the Holy Cross Mountains, c. 10 km to the east of Kielce city (Fig. 2). Cambrian rocks form the core of an inverted anticline and build the bottom of a WNW–ESE-oriented valley mostly covered by fields and grasslands. There is no exposure and the specimens may be found only in loose sandstone blocks on the fields, especially after spring ploughing. The sandstone bedrock was discovered in the mid-1960s by Wiesław Bednarczyk, but presently the location of the trenches made by him cannot be reconstructed (Żylińska & Masiak, Reference Żylińska and Masiak2007). In 2006 another trench was dug close to where the loose blocks with fossils could be found, but it exposed only shales that, based on a low-diversity and low-frequency acritarch assemblage, were assigned to the traditional ‘Middle’ Cambrian; a more precise stratigraphic resolution was not possible and the relation of these rocks to the sandstones yielding macrofossils was not recognized (Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009). The diverse and unique assemblage of fossils found in Brzechów includes trilobites (Czarnocki, Reference Czarnocki1927a, b; Bednarczyk, Reference Bednarczyk1970; Żylińska & Masiak, Reference Żylińska and Masiak2007), brachiopods (Bednarczyk, Reference Bednarczyk1970), imprints of soft-bodied animals (Czarnocki, Reference Czarnocki1927a, b; Stasińska, Reference Stasińska1960; Masiak & Żylińska, Reference Masiak and Żylińska1994) and other remains of organisms (A.Ż., unpublished data). Two species from the Family Paradoxididae Hawle & Corda, Reference Hawle and Corda1847 have been described from this locality: Paradoxides oelandicus Sjögren, Reference Sjögren1872 by Bednarczyk (Reference Bednarczyk1970) and Paradoxides (Acadoparadoxides) cf. mureroensis (Sdzuy, Reference Sdzuy1958) by Żylińska (in Żylińska & Masiak, Reference Żylińska and Masiak2007). The first taxon was the basis for correlating the rocks from this locality with the Scandinavian Acadoparadoxides oelandicus Superzone (Bednarczyk, Reference Bednarczyk1970). Besides paradoxidids, the trilobites from Brzechów include representatives of the Family Ellipsocephalidae Matthew, Reference Matthew1887 (Kingaspidoides sanctacrucensis (Czarnocki, Reference Czarnocki1927a) and Issafeniella orlowinensis (Samsonowicz, 1959)), the Family Palaeolenidae Hupé, Reference Hupé1953 (Palaeolenus medius (Bednarczyk, Reference Bednarczyk1970)) and a member of the Conomicmacca/Myopsolenites clade (Myopsolenites kielcensis (Bednarczyk, Reference Bednarczyk1970)) (see revision in Żylińska & Masiak, Reference Żylińska and Masiak2007 and remarks on Myopsolenites kielcensis in Elicki & Geyer, Reference Elicki and Geyer2013 and Geyer, Reference Geyer2016). The two latter species are important for biostratigraphy, as their presence had led to the correlation of the Brzechów sandstones with the Moroccan Morocconus notabilis Zone (Żylińska & Masiak, Reference Żylińska and Masiak2007). The co-occurrence of Acadoparadoxides oelandicus with Acadoparadoxides cf. mureroensis was additional evidence that the stratigraphic interval represented by the Brzechów sandstones was older than the oldest traditional ‘Middle’ Cambrian strata from Scandinavia, and the base of the Eccaparadoxides insularis Zone in the Holy Cross Mountains was thus placed earlier than in Scandinavia (Żylińska & Masiak, Reference Żylińska and Masiak2007; Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009).

Figure 2. Sketch map of the Brzechów area with location of the Brzechów outcrop.
The second locality, Jugoszów–Usarzów, comprises numerous small outcrops in the eastern part of the Holy Cross Mountains, described in detail by Orłowski (Reference Orłowski1959a, b, Reference Orłowski1964). The Jugoszów–Usarzów section sensu stricto is composed of 25 small outcrops along the Kurówka stream (Fig. 3). Additional outcrops are located in the close vicinity, near Sternalice, Słoptów and Helenów villages (Fig. 4), of which the presently non-existing Sternalice outcrop had yielded abundant specimens of trilobites. Previously described taxa (Orłowski, Reference Orłowski1959a, Reference Orłowski1964; Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009) include eight species of paradoxidids, of which four were known also from Scandinavia, i.e. Acadoparadoxides oelandicus, Acadoparadoxides pinus (Westergård, Reference Westergård1936), Acadoparadoxides torelli (Westergård in: Asklund & Thorslund, Reference Asklund and Thorslund1935) and Eccaparadoxides insularis (Westergård, Reference Westergård1936); one was known from Gondwanan successions, i.e. Acadoparadoxides cf. mureroensis; and three were endemic taxa, i.e. Paradoxides czarnockii Orłowski, 1959, Paradoxides kozlowskii Orłowski, 1959 and Paradoxides samsonowiczi Orłowski, 1959. Besides paradoxidids, the strata exposed in the Jugoszów–Usarzów section and the nearby localities yield numerous trilobites of the Family Ellipsocephalidae (12 species; Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009). Their presence and diversity are similar to those from Gondwanan and Avalonian successions rather than from Baltica (Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009; Żylińska, Reference Żylińska2013a). Other faunal elements known from the succession include brachiopods (Orłowski, Reference Orłowski1964; Jendryka-Fuglewicz, Reference Jendryka-Fuglewicz1992), primitive echinoderms (Orłowski, Reference Orłowski1964; Dzik & Orłowski, Reference Dzik and Orłowski1995), and hyoliths and pelagiellids (Orłowski, Reference Orłowski1964).

Figure 3. Sketch map of the Jugoszów–Usarzów section, modified from Orłowski (Reference Orłowski1964).

Figure 4. Sketch map of the Jugoszów–Usarzów locality with historical outcrops and sections (after Orłowski Reference Orłowski1964).
The hitherto used biostratigraphic scheme for the interval exposed in Brzechów and Jugoszów–Usarzów includes two biozones derived from Scandinavia: the Paradoxides insularis and the Paradoxides pinus zones (e.g. Orłowski, Reference Orłowski1992). The range of the first zone was later extended below to include the part of the Cambrian succession that was correlated with the Morocconus notabilis Zone from Morocco (Żylińska & Masiak, Reference Żylińska and Masiak2007; Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009), but was absent in Scandinavia (e.g. Nielsen & Schovsbo, Reference Nielsen and Schovsbo2015). The lowermost part of this zone was to be represented in Brzechów, whereas the remaining part was supposed to occur in Jugoszów–Usarzów.
3. Material and methodology
3.a. Material
Most of the analysed material was collected by Stanisław Orłowski and is now reposited in the Stanisław Józef Thugutt Geological Museum, Faculty of Geology, University of Warsaw. Additional material from Brzechów was collected by Wojciech Kozłowski and is reposited in the collection of the Department of Historical and Regional Geology, Institute of Basic Geology at the Faculty of Geology, University of Warsaw. Orłowski's collection includes 280 specimens of paradoxidids from Brzechów and Jugoszów–Usarzów. Except for two partial carapaces, all specimens are isolated sclerites: cranidia, pygidia, hypostomata and parts of thoraces. The material is preserved as both internal and external moulds and has experienced significant tectonic deformation, which, however, is simple; the axes of the specimens have not been bent and remain straight (Fig. 5a). Moreover, the specimens are only marginally affected by compaction (Fig. 5b). The former determination of the material includes the following species (names as on labels): Paradoxides (Acadoparadoxides) cf. mureroensis, Paradoxides (Acadoparadoxides) oelandicus, Paradoxides (Acadoparadoxides) pinus, Paradoxides (Eccaparadoxides) insularis, Paradoxides (Eccaparadoxides) torelli, Paradoxides czarnockii, Paradoxides kozlowskii and Paradoxides samsonowiczi. Kozłowski's collection contains nine specimens from Brzechów (isolated cranidia, pleurae, and one incomplete pygidium). Likewise, the material is preserved as internal and external moulds, and is distorted. The specimens were labelled as Paradoxides (Acadoparadoxides) cf. mureroensis and Paradoxides (Acadoparadoxides) oelandicus.

Figure 5. Examples of the style of deformation expressed by trilobite cranidia from the study area. (a) Strongly skewed specimen of Acadoparadoxides sp., MWGUW ZI/29/3252; (b) Side view of cranidium of Acadoparadoxides kozlowskii (Orłowski, 1959) with very low degree of compaction, original of Orłowski (Reference Orłowski1964), pl. 2, fig. 2, and Żylińska & Szczepanik (Reference Żylińska and Szczepanik2009), pl. 4, fig. 9; MWGUW ZI/29/3210. Scale bars equal 5 mm.
3.b. Methodology
Revision of distorted material requires special tools to remove the deformation and restore the original relations between the specimens. There are numerous cases in which retrodeformation techniques have been successfully applied to tectonically deformed trilobites (e.g. Hughes & Jell, Reference Hughes and Jell1992; Żylińska, Kin & Nowicki, Reference Żylińska, Kin and Nowicki2013). Similar tools have already been used to evaluate some features in the Paradoxididae (Esteve, Reference Esteve2014; Rushton, Weidner & Ebbestad, Reference Rushton, Weidner and Ebbestad2016); however, the outcome of these analyses, which used linear measurements, had rather minor influence on the research results, especially in the latter cited paper. A principal component analysis (PCA) was not able to show differences between trilobite species distinguished by qualitative features (including the well-known and well-defined taxa in Rushton, Weidner & Ebbestad, Reference Rushton, Weidner and Ebbestad2016). Although the analyses were not free from technical mistakes (e.g. Rushton, Weidner & Ebbestad, Reference Rushton, Weidner and Ebbestad2016 included overlapping measurements in the PCA) which could have influenced the obtained results, above all, traditional morphometric analyses are obviously insufficient for this group. One reason may be the subtlety of the differences between particular paradoxidid species (e.g. Geyer & Vincent, Reference Geyer and Vincent2015). A possible solution is the application of a more sensitive method, i.e. geometric morphometrics (e.g. Zelditch, Swiderski & Sheets, Reference Zelditch, Swiderski and Sheets2012). The risk of potential inconsistencies in this case is that the ‘noise’ coming from preservation may be overrepresented and may obscure the true morphological variation in the sample.
In geometric morphometrics, PCA may be used in a similar way to its use in classical morphometrics (e.g. Hughes & Jell, Reference Hughes and Jell1992; Żylińska, Kin & Nowicki, Reference Żylińska, Kin and Nowicki2013). One of the components may be interpreted as resulting from deformation caused by tectonics and be removed from the analysed sample to bring back the original variance. The component can be detected manually, by visualizing shapes that correspond to the maximal/minimal value of the component, or by examining the correlation between the components and the length:width proportions. In the latter case, high correlation may suggest that this component represents tectonic deformation. After removing the ‘tectonic’ component, the remaining components can be analysed with various tools, including cluster analysis and distance visualizations, such as Nonmetric Multidimensional Scaling (NMDS).
Angielczyk & Sheets (Reference Angielczyk and Sheets2007) presented a comprehensive validation of numerical retrodeformation techniques in geometric morphometrics. They recognized PCA as a method that does not successfully remove tectonic variance from the dataset. The major problem with PCA application was the distribution of tectonic variance between several components. Removal of only one component did not significantly improve the outcome, whereas removal of more than one component resulted in also throwing out part of the variance that was not connected with deformation. It should be noted, however, that the cited study was based on a single dataset and did not consider a combination of retrodeformation techniques. The latter approach may be a possible solution for the problem of distribution of tectonic variance by joining PCA with another technique: reflection and averaging of bilaterally symmetric landmarks, i.e. symmetrization. To test this approach, we performed a simulation using the R programming language (R Core Team, 2016) – the geomorphotest package (Nowicki, Reference Nowicki2016). As a measure of efficiency of retrodeformation we used a correlation (Pearson's r coefficient) of the Euclidean distances between retrodeformed specimens in a principal component (PC) space and the distances between non-deformed specimens. This statistic was selected because the clustering methods that were to be used to determine the classification of the specimens were based on the distances between the specimens. The test sample contained 1000 datasets, each with 30 specimens. Each specimen had 20 landmarks and was characterized by axial symmetry. The test was repeated nine times, with different deformation ranges. We followed the deforming procedure used by Angielczyk & Sheets (Reference Angielczyk and Sheets2007) – application of a deformation matrix in the form:
 $$\begin{equation*} \left| {\begin{array}{@{}*{2}{c}@{}} {a{\rm{cos\Theta }}}&{{\rm{sin\Theta }}}\\ {{\rm{sin\Theta }}}&{\left( {1/a} \right){\rm{cos\Theta }}} \end{array}} \right| \end{equation*}$$
$$\begin{equation*} \left| {\begin{array}{@{}*{2}{c}@{}} {a{\rm{cos\Theta }}}&{{\rm{sin\Theta }}}\\ {{\rm{sin\Theta }}}&{\left( {1/a} \right){\rm{cos\Theta }}} \end{array}} \right| \end{equation*}$$
where coefficient a is a normal strain and Θ is a shear strain.
In the first three approaches, deformation was restricted to the applied matrices; in the next six approaches some amount of random noise was added to the deformation. Noise was added to the deformation coefficients as a random value, different for each landmark and each coefficient, and derived from normal distribution with a mean 0 and a standard deviation of 0.01 for groups 4–6 and 0.03 for groups 7–9 (Θ was converted to radians). The angle of the deformation was applied randomly to each specimen within each dataset. Coefficient a was derived from the interval 0.95–1. For groups 1, 4 and 7, Θ was derived from the interval 1–10°. For groups 2, 5 and 8, Θ was derived from the interval 10–20°. For groups 3, 6 and 9, Θ was derived from the interval 1–20°. The deformation parameters are summarized in Table 1. In the simulations we intentionally ignored variation related to ontogenetic development. In the Paradoxididae, the shape of the glabella (an element considered in Section 5 of this paper) is quite stable during late ontogeny (more mature meraspid and holaspid stages; see illustrations in Westergård, Reference Westergård1936; Šnajdr, Reference Šnajdr1958). In our analysis, only PC3 (5.1% of variance) and PC8 (0.9% of variance) may be correlated with the size of the cranidia, although this correlation is weak (for PC3: r=−0.379, p-value = 0.0354; for PC8: r=−0.384, p-value = 0.0327). Also, in general, the paradoxidids have a similar ontogenetic development in mature stages (reduction of the relative sizes of the pre-glabellar field and the palpebral lobes) and the species from Poland follow this trend. Ontogenetic allometry is therefore unlikely to account for the interspecific differences documented herein.
Table 1. Parameters of deformation in the simulated landmark sets

Removing the PC with the highest correlation with the length:width proportions from the sets without symmetrization does not improve the correlation of the distances between the specimens in the non-deformed sets and the retrodeformed sets (Fig. 6). In most cases, both the correlation of the deformed set and the correlation of the retrodeformed set are close to 0; therefore, this technique does not improve the outcome in comparison to the non-retrodeformed samples. A meaningful improvement may be observed in the datasets with symmetrization (Fig. 7). In the three groups with no deformation noise (groups 1–3), this improvement may be observed in almost all cases, and the best outcome appears with higher Θ values. The same pattern may be observed in groups with higher values of the deformation noise (groups 4–6). In the groups with intermediate levels of deformation noise (0.01), the results remain promising: most cases show quite high improvement of correlation in comparison to the sets without retrodeformation. Unfortunately, in the third group (7–9), with the highest amount of noise added to the deformation, only part of the sets was successfully retrodeformed, but even in this case, almost all sets displayed better performance than in the non-retrodeformed sets.

Figure 6. Comparison of the correlation with distances between the specimens in non-deformed sets. x-axis: correlation with deformed sets; y-axis: correlation with sets retrodeformed without symmetrization. Ellipses represent the 95% confidence interval.

Figure 7. Comparison of the correlation with distances between the specimens in non-deformed sets. x-axis – correlation with deformed sets; y-axis: correlation with sets retrodeformed with symmetrization. Ellipses represent the 95% confidence interval.
The results clearly show that removal of a PC that is best correlated with the length:width proportions does not improve the outcome when the axial symmetry is not restored in the specimens before the procedure. Symmetrization improves the results, but can be safely applied only to data in which deformation is relatively simple, without a high amount of noise added to the deformation. Such simple deformation may be found in sandstones. On the other hand, it seems that in fine-grained rocks (shales), the complexity of deformation, which often includes bending and flattening, precludes usage of the described technique. The problem of combined retrodeformation in datasets with ontogenetic variance, and also the validation of more complex outputs of retrodeformation is beyond the scope of this paper, but successful application in this case may be taken as encouragement for solving these issues.
Due to the poor preservation of the studied specimens, only three cranidia were sufficiently complete to provide data for geometric morphometrics analysis. To overcome this problem, the analysis focused exclusively on the glabellae and aimed at detecting differences among them. Twenty-seven specimens from various outcrops in the Jugoszów–Usarzów locality and four specimens from Brzechów comprised a dataset sufficient to acquire landmarks from the glabellae. Data for the analyses were collected from digital photographs of the specimens (coated with ammonium chloride before photographing) using TPS software (Rohlf, Reference Rohlf2016). Each photograph was preprocessed before landmark acquisition by restoring the symmetry of the cranidia (using a technique similar to that presented by Srivastava & Shah, Reference Srivastava and Shah2006) in order to allow for estimation of the missing landmarks. The specimens were not fully retrodeformed; instead, only the axis of symmetry was restored by shearing the photographs. A dataset of 11 landmarks and 26 semi-landmarks was collected from each glabella (Table 2; Fig. 8). In the analysis, we used landmarks situated on both sides of the glabellae to avoid the transfer of variance from the glabellar sides to the axial landmarks. This was crucial, since the applied retrodeformation techniques are based on the assumption that the axis of symmetry is preserved as a straight line. All further actions were performed using the R programming language (R Core Team, 2016). Pairs of landmarks/semi-landmarks on the opposite sides of the glabella were averaged, or, if only one landmark/semi-landmark of the pair was preserved, the missing landmark/semi-landmark was estimated (according to the positions of all existing landmarks) using the algorithm proposed by Klingenberg, Barluenga & Meyer (Reference Klingenberg, Barluenga and Meyer2002). Both actions were performed using the OSymm function (Haber, Reference Haber2011). Averaging of the semi-landmarks was possible due to embedding of fans (radii of circles) on the photographs. Since axial symmetry was restored in the specimens, the semi-landmarks were located in homologous positions on both sides of the glabellae (in a single specimen; the homology between specimens was achieved with sliding semi-landmarks), defined by the intersection of the described curve and the symmetric radii. Landmarks and semi-landmarks were superimposed (Generalized Procrustes Analysis). Semi-landmarks were slided with the Procrustes distance criterion applied. Both actions were performed using the geomorph package (Adams & Otárola-Castillo, Reference Adams and Otárola-Castillo2013; Adams et al. Reference Adams, Collyer, Kaliontzopoulou and Sherratt2016). The same package was used for PCA application. Major data manipulation, including symmetrization, was performed using the morphoutils package (Nowicki, Reference Nowicki2015).
Table 2. Location of landmarks and semi-landmarks on the glabella of the studied paradoxidids


Figure 8. Location of collected landmarks and semi-landmarks on a paradoxidid glabella.
4. Revision of former taxonomic assignments
Among the taxa described in the previous papers (Orłowski, Reference Orłowski1959a, Reference Orłowski1964; Bednarczyk, Reference Bednarczyk1970; Żylińska & Masiak, Reference Żylińska and Masiak2007; Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009) occurring in Jugoszów–Usarzów (eight species) and Brzechów (two species), most taxa were diagnosed based only on cranidia, with the exception of two endemic species represented only by pygidia (Paradoxides kozlowskii and Paradoxides samsonowiczi). Except for a relatively detailed description of the differences between specimens assigned previously to Paradoxides (Acadoparadoxides) oelandicus and Paradoxides (Acadoparadoxides) cf. mureroensis given by Żylińska & Masiak (Reference Żylińska and Masiak2007), none of the cited papers presents a comparison between the paradoxidid species, therefore the diagnostic features extracted from Orłowski Reference Orłowski(1959a) are compared in Table 3. Specimens assigned by Orłowski (Reference Orłowski1964) to Paradoxides torelli are the only ones that differ significantly from the remaining paradoxidids; however, they are very poorly preserved and were not included in the comparison. Their species assignment is not possible; additionally, Weidner, Rushton & Ebbestad (Reference Weidner, Rushton and Ebbestad2014) have shown that they definitely cannot be attributed to Acadoparadoxides torelli as currently known from Scandinavia. Differences between the types of cranidia (excluding the specimens assigned to P. torelli) described by Orłowski (Reference Orłowski1959a) are doubtful. The major character emphasized by Orłowski (Reference Orłowski1959a) for cranidia assigned to Paradoxides insularis and Paradoxides czarnockii was the presence of the S3 and S4 glabellar furrows. In reality, these furrows are present also on the glabellae of specimens assigned by Orłowski to Paradoxides oelandicus and Paradoxides pinus (Reference OrłowskiOrłowski 1959a, pl. 1, figs 2, 4, 6, 7). In all cases, the furrows are not well marked, unlike in the type specimens of Eccaparadoxides insularis from Sweden, in which they are clearly developed (e.g. Westergård Reference Westergård1936, pl. 7 figs 1, 2a, 5, 8, 9). This difference is evidently not caused by variable preservation of the material from the Holy Cross Mountains and the material from Scandinavia. Another difference between Eccaparadoxides insularis from Scandinavia and the paradoxidid species in question is the disparity in the length:width proportions of the cranidia. All cranidia assigned by Orłowski (Reference Orłowski1959a) to Paradoxides insularis show traces of tectonic shortening along the axis, therefore this character is inappropriate for distinguishing between the taxa.
Table 3. Diagnostic differences between the specimens of Paradoxides oelandicus, Paradoxides pinus, Paradoxides insularis and Paradoxides czarnockii from the Holy Cross Mountains after Orłowski (Reference Orłowski1959a)
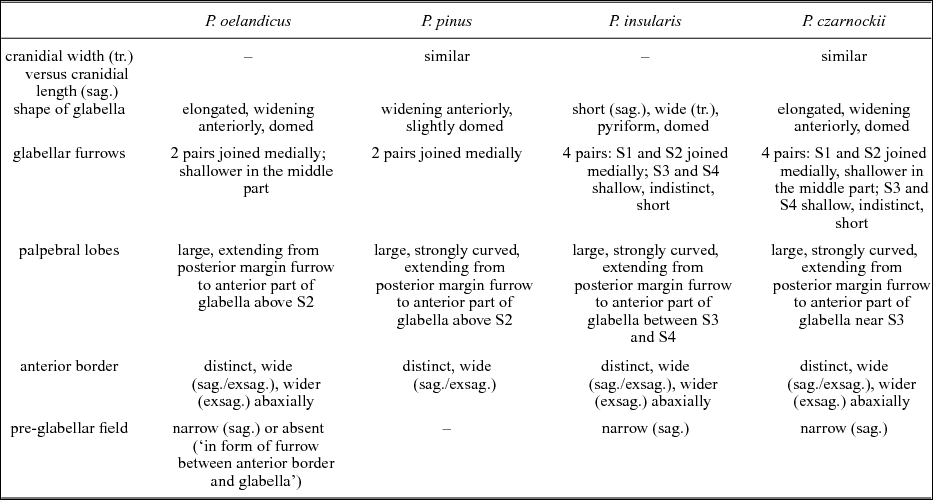
All the characters listed for specimens from Brzechów assigned to Paradoxides (Acadoparadoxides) oelandicus and Paradoxides (Acadoparadoxides) cf. mureroensis by Żylińska & Masiak (Reference Żylińska and Masiak2007) follow the same pattern: the specimens assigned to Paradoxides (Acadoparadoxides) cf. mureroensis are always narrower (Table 4). Owing to the presence of tectonic deformation, these differences are insufficient to distinguish between the taxa. The difference in the shape of glabellar furrows can be noted on only one specimen and seems to be rather an artefact than a true character.
Table 4. Diagnostic differences between the specimens of Paradoxides (Acadoparadoxides) cf. mureroensis and Paradoxides (Acadoparadoxides) oelandicus from the Holy Cross Mountains after Żylińska & Masiak (Reference Żylińska and Masiak2007)

Pygidia from Jugoszów–Usarzów were assigned to two distinct morphotypes. Paradoxides kozlowskii differs from Paradoxides samsonowiczi in the expression of several characters so that in this case the differentiation described by Orłowski (Reference Orłowski1959a, Reference Orłowski1964) appears to be correct. A single pygidium from Brzechów (Żylińska & Masiak, Reference Żylińska and Masiak2007, fig. 11j) is unfortunately incomplete; the posterior part is missing, making a precise taxonomic assignment impossible.
None of the pygidia from Jugoszów–Usarzów can be assigned to Scandinavian species that were described on the basis of cranidia. Moreover, the previously suggested diversities of cranidia and of pygidia are not compatible with one another. The diversity of the pygidia suggests that there should be two morphotypes of cranidia, whereas the number of distinguished morphotypes of cranidia is much larger. Cranidia assigned by Orłowski (Reference Orłowski1959a, Reference Orłowski1964) to Paradoxides torelli may be excluded from this analysis. Their abundance is very low (unlike the abundance of pygidia and other morphotypes of cranidia). Additionally, they bear features that appear to suggest their assignment to different genera than the pygidia (see Section 7 below).
5. Morphometric analysis
In order to determine morphotypes in tectonically deformed specimens, a PCA was performed using landmarks and semi-landmarks on the glabellae. In the first analysis, 27 specimens from Jugoszów–Usarzów were processed. Previous assignments of the analysed dataset include (names according to labels): 13 specimens of Paradoxides (Acadoparadoxides) oelandicus, six specimens of Paradoxides (Acadoparadoxides) pinus, four specimens of Paradoxides (Acadoparadoxides) cf. mureroensis, three specimens of Paradoxides (Eccaparadoxides) insularis and one specimen of Paradoxides czarnockii. The first component is responsible for 74% of variance (Table 5) in the sample and is associated with the length:width proportions of the glabella (correlation: r=−0.96; Fig. 9a); thus it may be treated as variance caused primarily by tectonic deformation. After removing PC1, the rest of the components represent a sufficient approximation of the diversity of the glabellae (as proven in the simulations). The diversity of the pygidia suggested that at least two morphotypes of cranidia should be detectable in the sample. Since PC2 was not connected with deformation (correlation with the length:width proportions of the glabella: r=0.23, p-value = 0.24) nor with ontogeny (correlation with centroid sizes: r=0.03, p-value = 0.87) and represented 10.81% of the variation, it should contain the major morphological diversity. To discover the morphotypes that would potentially be present in the sample, we applied a fuzzy C-Means Clustering algorithm (for details see description of the cmeans function in Meyer et al. Reference Meyer, Dimitriadou, Hornik, Weingessel and Leisch2017). To check the best number of clusters we performed 1000 repetitions of the procedure, each time calculating the partition coefficient, the partition entropy and the proportion exponent (for details see description of the fclustIndex function in Meyer et al. Reference Meyer, Dimitriadou, Hornik, Weingessel and Leisch2017). All indexes confirmed that the best outcome is obtained for two clusters (Fig. 10a). Fuzzy clustering allowed including all PC (except PC1) in the analysis and emphasized the presence of uncertainty in assigning the cranidia. As expected, PC2 was the component that subdivided the sample into two clusters. In the case of several specimens with middle values of PC2 it was not possible to validate their membership (the probabilities of belonging to both clusters were around 0.5), but in most cases the characters located on the other parts of the glabellae allowed such assignment. In two cases (specimens MWGUW ZI/29/3289 and MWGUW ZI/29/3323), characters other than the glabella suggested the assignment to morphotype B, despite the slightly higher probability of being the member of the morphotype A cluster; in the systematic part this is expressed with a question mark. Assignment of one specimen (MWGUW ZI/29/3378) remains unclear. As shown on the PC2 and PC3 scatterplot, the first group (morphotype A) contains widening-forward glabellae, with rather straight lateral margins, and the second group (morphotype B) contains glabellae with (sub)parallel lateral margins in the posterior portion and a bulb-shaped frontal portion (Fig. 11a). It should be emphasized that in this case the number of morphotypes of cranidia in the analysed succession corresponds with the number of morphotypes of pygidia. Both morphotypes of pygidia and both morphotypes of cranidia occur in the lower part of the Jugoszów–Usarzów succession, whereas pygidia described as Paradoxides kozlowskii and cranidia with bulb-shaped glabellae occur in its upper part.
Table 5. Eigenvalues and proportion of variance for the first ten PCs obtained for specimens from Jugoszów–Usarzów
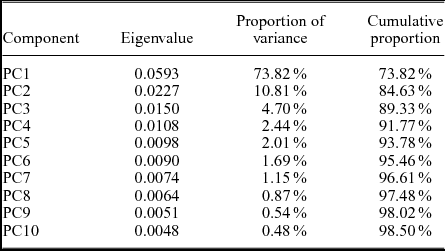

Figure 9. Correlation of PC1 with glabellar proportions with thin plate splines showing glabellar shapes corresponding to the minimum and maximum value of PC1, for (a) specimens from Jugoszów–Usarzów, and (b) specimens from Jugoszów–Usarzów and Brzechów combined. Inner circle represents the probability of being a member of morphotype A (Acadoparadoxides samsonowiczi); outer circle represents the probability of being a member of morphotype B (Acadoparadoxides kozlowskii).

Figure 10. Results of 1000 tests of fuzzy c-means clustering validate indexes (partition coefficient: highest best; partition entropy: lowest best; and proportion exponent: highest best) for (a) specimens from Jugoszów–Usarzów, and (b) specimens from Jugoszów–Usarzów and Brzechów combined. Points indicate the mean values, error bars indicate the range of ±3 standard deviations.

Figure 11. Scatterplots showing the relationship of PC2 and PC3 with thin plate splines showing glabellar shapes corresponding to the minimum and maximum value of PC2. (a) Specimens from Jugoszów–Usarzów; (b) specimens from Jugoszów–Usarzów and Brzechów combined. Inner circle represents the probability of being a member of morphotype A (Acadoparadoxides samsonowiczi); outer circle represents the probability of being a member of morphotype B (Acadoparadoxides kozlowskii).
Four specimens from Brzechów (two previously assigned to Paradoxides (Acadoparadoxides) oelandicus and two previously assigned to Paradoxides (Acadoparadoxides) cf. mureroensis)) were added to the analysed sample, and the PCA was repeated (Table 6). The interpretations are robust: the first component still has the same features that allow it to be interpreted (with the same reservations as in the previous analysis) as resulting from tectonic deformation (correlation: r=0.94; Fig. 9b). Moreover, the subdivision between the two morphotypes of glabellae is also present. All specimens from Brzechów can be assigned to morphotype A, with specimens having a glabella that widens forwards and has straight lateral margins. This assignment can be seen on the scatterplot of PC2 and PC3 (Fig. 11b). In this case, PC2 is still uncorrelated with deformation (correlation with the length:width proportions of the glabella: r=0.27, p-value = 0.14) or ontogeny (correlation with centroid sizes: r=0.16, p-value = 0.38). Again, fuzzy clustering provided the best results for two clusters (Fig. 10b). The PCA scores from the full set of specimens with PC1 excluded have also been visualized using NMDS (Fig. 12). The stress for the scaling was 0.13, which is an acceptable value (Clarke, Reference Clarke1993). The absence of the ‘bulb-shaped’ glabellae in Brzechów may suggest that the strata containing only one morphotype (glabellae with parallel sides) are older than the strata from the Jugoszów–Usarzów succession, which conforms with the conclusions in Żylińska & Masiak (Reference Żylińska and Masiak2007) and Żylińska & Szczepanik (Reference Żylińska and Szczepanik2009).
Table 6. Eigenvalues and proportion of variance for the first ten PCs obtained for specimens from Jugoszów–Usarzów and Brzechów combined
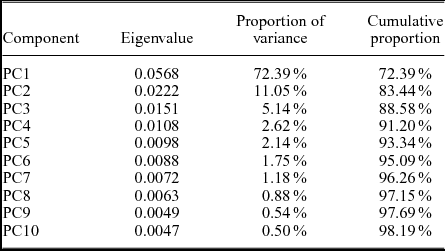

Figure 12. Visualization of PCA (with PC1 excluded) using NMDS for the studied specimens. Inner circle represents the probability of being a member of morphotype A (Acadoparadoxides samsonowiczi); outer circle represents the probability of being a member of morphotype B (Acadoparadoxides kozlowskii).
As mentioned earlier, ontogenetic development is unlikely to have influenced the outcome of the morphometric analysis. Sizes of the glabellae (sagittal length of the glabella with occipital ring) included in the analysis varied from 7 to 30 mm for morphotype A and from 6 to 34 mm for morphotype B.
The presence of two species (and one additional taxon, previously described as Paradoxides torelli in the studied samples) is supported by the diversity of both cranidia and pygidia. The differences in the cranidia are very subtle, so assignment is often difficult. The differences in the pygidia are much more obvious. It is important to emphasize that the preservation of the material does not allow for a more detailed description, but it cannot be excluded that the true diversity is higher. Recognition of cryptic species may be possible only with careful and precise collection of samples from the continuous section, such as the one described from Morocco by Geyer & Vincent (Reference Geyer and Vincent2015); thus the diversity presented herein seems to be the best hypothesis based on presently available data.
6. Institutional abbreviations and terminology
MWGUW: Stanisław Józef Thugutt Geological Museum (Muzeum Geologiczne im. Stanisława Józefa Thugutta), Faculty of Geology, University of Warsaw, Warsaw, Poland; WK: collection of Wojciech Kozłowski in the archives of the Department of Historical and Regional Geology, Institute of Basic Geology, Faculty of Geology, University of Warsaw, Warsaw, Poland. The terminology used herein follows that used in Whittington (Reference Whittington, Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Geyer, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot and Kelly1997).
7. Systematic descriptions
Class TRILOBITA Walch, Reference Walch1771
Order REDLICHIIDA Richter, Reference Richter, Dittler, Joos, Korschelt, Linck, Oltmanns and Schaum1932
Family PARADOXIDIDAE Hawle & Corda, Reference Hawle and Corda1847
Genus Acadoparadoxides Šnajdr, Reference Šnajdr1957
Type species. Paradoxides sacheri (Barrande, Reference Barrande1852) from the Cambrian of Bohemia; SD by Šnajdr (Reference Šnajdr1957, p. 146).
Diagnosis. For characters defining Acadoparadoxides, see Geyer & Vincent (Reference Geyer and Vincent2015).
Remarks. Modern ideas about Acadoparadoxides in general follow the original concept of Šnajdr (Reference Šnajdr1957, Reference Šnajdr1958), modified by Sdzuy (Reference Sdzuy1967) and Šnajdr (Reference Šnajdr1986, Reference Šnajdr1987), although the subdivision between Acadoparadoxides (Acadoparadoxides) and Acadoparadoxides (Baltoparadoxides) as indicated in the treatise (Dean & Rushton, Reference Dean, Rushton, Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Geyer, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot and Kelly1997, pp. O474–5) has recently been criticized by Geyer & Vincent (Reference Geyer and Vincent2015). This paper generally follows the summarized concept provided by the latter authors, although another subdivision within this genus that may have strong impact on biostratigraphic interpretations is demonstrated below.
Species assigned to this genus from Morocco (Geyer, Reference Geyer1993, Reference Geyer1998; Geyer & Vincent, Reference Geyer and Vincent2015), Spain (Sdzuy, Reference Sdzuy1958), Turkey (Dean, Reference Dean2005), Newfoundland (Geyer & Landing, Reference Geyer and Landing2001) and possibly Siberia (specimens described as other genera in Solov'ev, Reference Solov'ev1969; see discussion in Geyer & Vincent, Reference Geyer and Vincent2015) differ significantly from the taxa described from the Czech Republic (Šnajdr, Reference Šnajdr1957, Reference Šnajdr1958, Reference Šnajdr1986, Reference Šnajdr1987) and from Scandinavia (e.g. Weidner, Rushton & Ebbestad, Reference Weidner, Rushton and Ebbestad2014). The main differences are as follows: the species from Bohemia and especially the species from Scandinavia have the anterior part of the cranidial border narrower (tr.) in comparison to the total width (tr.) of the cranidium in the middle point of the palpebral lobes; and the palpebral lobes of the Czech and Scandinavian taxa are wider (tr.), longer (exsag.) and more strongly curved. Important differences are displayed between the pygidia of Scandinavian species and other acadoparadoxidines. The variety of Scandinavian pygidia includes rounded or slightly elongated forms with spines (Acadoparadoxides oelandicus and Acadoparadoxides bidentatus (Westergård, Reference Westergård1936)); hexagonal forms with an optional indentation in the posterior margin (Acadoparadoxides torelli (Westergård in: Asklund & Thorslund, Reference Asklund and Thorslund1935)); and elongated forms with an indentation in the posterior margin (Acadoparadoxides pinus (Westergård, Reference Westergård1936)). In comparison, acadoparadoxidines from other regions have rounded, suboval or pyriform pygidia without spines. Geyer & Vincent (Reference Geyer and Vincent2015) proposed a cladogram for acadoparadoxidine pygidia that includes only non-Scandinavian species. This grouping seems to be quite well defined, although it has not been tested with phylogenetic analysis. Phylogenetic relations between this group and the Scandinavian acadoparadoxidines remain unclear and in fact the latter group has many characters in common with the genus Eccaparadoxides Šnajdr, Reference Šnajdr1957, especially with the Scandinavian species Eccaparadoxides insularis and Eccaparadoxides? thorslundi Rushton, Weidner & Ebbestad, Reference Rushton, Weidner and Ebbestad2016, and with the Scandinavian taxa assigned to Hydrocephalus (i.e. H. vikensis Rushton & Weidner, Reference Rushton and Weidner2007 and H. spinulosus Rushton, Weidner & Ebbestad, Reference Rushton, Weidner and Ebbestad2016). True relations between Scandinavian taxa need a phylogenetic revision, but it is probable that A. torelli, E?. thorslundi, H. vikensis and H. spinulosus may form one clade.
A possible revised internal subdivision of Acadoparadoxides should be developed with phylogenetic data. Until then, informal grouping may be applied, with the major groups described as follows:
1. The earliest acadoparadoxidines comprising species with a wider (tr.) anterior part of the cranidium, smaller and thinner (tr.) palpebral lobes and pygidia that are rounded, suboval or pyriform. This group includes: Acadoparadoxides briareus Geyer, Reference Geyer1993, Acadoparadoxides levisettii Geyer & Vincent, Reference Geyer and Vincent2015, Acadoparadoxides cf. mureroensis, Acadoparadoxides nobilis Geyer, Reference Geyer1998, Acadoparadoxides ovatopyge Geyer & Vincent, Reference Geyer and Vincent2015 and Acadoparadoxides pampalius Geyer & Vincent, Reference Geyer and Vincent2015, from Morocco and partially from SpainFootnote *; Acadoparadoxides deani Geyer & Vincent, Reference Geyer and Vincent2015 from Turkey; and Acadoparadoxides harlani (Green, Reference Green1834) from Newfoundland. Some Siberian species may be included in this group, but a major revision of this material is necessary. The species from the Holy Cross Mountains presented herein also fit well in this group and are the first taxa assigned to this group that come from outside Gondwana.
2. The later group of acadoparadoxidines is much more diverse. It includes species with a narrower (tr.) anterior part of the cranidium and wider (tr.), longer (exsag.) and slightly more curved palpebral lobes. The pygidia in the Bohemian taxa (Acadoparadoxides sacheri and Acadoparadoxides sirokyi (Šnajdr, Reference Šnajdr1986)) are rounded and quite similar to the later species from the first group (e.g. Acadoparadoxides harlani). The pygidia of the Scandinavian forms include various shapes: rounded or slightly elongated forms with spines, hexagonal forms with a variably present indentation in the posterior margin, and elongated forms with an indentation in the posterior margin. It is probable that this group contains several clades, and a phylogenetic analysis should be performed to resolve this problem. Nevertheless, these species differ significantly from representatives of the first group.
The relations, both phylogenetic and palaeobiogeographic, between these two groups remain unclear. Only their stratigraphic relations can be described, as reflected in the names of the groups (earlier versus later acadoparadoxidines), but the nature of how the older group was replaced by the younger group remains unknown. In Scandinavia, the lack of the earliest acadoparadoxidines could be caused by palaeobiogeographic factors (true absence of this group in Baltica), or by the lack of strata of similar age to the interval with the earliest acadoparadoxidines in Morocco, Spain and the Holy Cross Mountains (Hawke Bay unconformity; e.g. Nielsen & Schovsbo, Reference Nielsen and Schovsbo2015). Moreover, data about the later acadoparadoxidines from Morocco have never been published. In a broader view, the middle Cambrian fauna of Baltica (Scandinavia) and West Gondwana (Morocco) clearly seems to reflect different palaeobiogeographic regions, and the fossils from the Holy Cross Mountains are more closely related to the fauna from Morocco (see palaeobiogeographic part of the discussion).
Acadoparadoxides kozlowskii (Orłowski, Reference Orłowski1959)
Figures 5b, 13
pars 1957 Paradoxides forms of the ölandicus group; Orłowski, figs 4, 6 (non figs 1–3 = Acadoparadoxides slowiecensis; non fig. 5 = Acadoparadoxides samsonowiczi).
pars 1959a Paradoxides oelandicus Sjögren, Reference Sjögren1872; Orłowski, pp. 441–2, pl. 1, figs 1–3 (non fig. 4 = Acadoparadoxides samsonowiczi; non fig. 5 = Paradoxididae gen. et sp. indet.).
pars 1959a Paradoxides pinus Holm; Orłowski, p. 442, pl. 1, fig. 6 (non fig. 7 = Acadoparadoxides samsonowiczi; non fig. 8 = Paradoxididae gen. et sp. indet.).
1959a Paradoxides insularis Wgård [sic]; Orłowski, p. 443, pl. 2, figs 2, 3.
1959a Paradoxides kozlowskii n. sp.; Orłowski, p. 444, pl. 2, figs 6a, b, 7.
pars 1964 Paradoxides oelandicus Sjögren; Orłowski, pl. I, figs 3, 4, 6–8 [non fig. 5 = Acadoparadoxides samsonowiczi; non figs 1, 9–11 = Paradoxididae gen. et sp. indet.).
1964 Paradoxides pinus Holm; Orłowski, pl. II, figs 1–3.
1990 Paradoxides kozlowskii Orłowski, 1959; Bednarczyk, Lendzion & Orłowski, p. 58, pl. XVI, fig. 9.
2009 Paradoxides (Acadoparadoxides) oelandicus (Sjögren, Reference Sjögren1872) [sic]; Żylińska & Szczepanik, pl. 4, fig. 7.
2009 Paradoxides (Eccaparadoxides) insularis Westergård, Reference Westergård1936; Żylińska & Szczepanik, pl. 4, fig. 8.
2009 Paradoxides (Eccaparadoxides) pinus Westergård, Reference Westergård1936; Żylińska & Szczepanik, pl. 4, fig. 9.
pars 2013a Paradoxides (Acadoparadoxides) oelandicus Sjögren, Reference Sjögren1872; Żylińska, fig. 7A (non fig. 6F = Acadoparadoxides samsonowiczi).
2013a Paradoxides (Eccaparadoxides) insularis Westergård, Reference Westergård1936; Żylińska, fig. 7D.
2013a Paradoxides (Eccaparadoxides) pinus Westergård, Reference Westergård1936; Żylińska, fig. 7G.
Holotype. Pygidium MWGUW ZI/29/3217, illustrated as specimen no. 465 in Orłowski (Reference Orłowski1959a, pl. II, fig. 7) and Bednarczyk, Lendzion & Orłowski (Reference Bednarczyk, Lendzion and Orłowski1990, pl. XVI, fig. 9).
Other material. Thirty-seven cranidia: MWGUW ZI/29/3210, ZI/29/3211, ZI/29/3220, ZI/29/3222, ZI/29/3244, ZI/29/3250, ZI/29/3254, ZI/29/3255, ZI/29/3260, ZI/29/3280, (?) ZI/29/3289, ZI/29/3292, ZI/29/3298, ZI/29/3301, ZI/29/3305, ZI/29/3311, ZI/29/3316, ZI/29/3320, (?) ZI/29/3323, ZI/29/3324, ZI/29/3325, ZI/29/3328, ZI/29/3334, ZI/29/3338, ZI/29/3340, ZI/29/3343, ZI/29/3348-ZI/29/3350, ZI/29/3355, ZI/29/3357, ZI/29/3370, ZI/29/3392, ZI/29/3397, ZI/29/3406, ZI/29/3416, ZI/29/3428; eleven pygidia: MWGUW ZI/29/3266, ZI/29/3272, ZI/29/3286, ZI/29/3304, ZI/29/3319, ZI/29/3322, ZI/29/3335, ZI/29/3379, ZI/29/3383, ZI/29/3395, ZI/29/3413.
Occurrence. Cambrian Series 2 and 3 boundary interval (Acadoparadoxides samsonowiczi – Acadoparadoxides kozlowskii Assemblage Zone) in the Usarzów Sandstone Formation from the Jugoszów–Usarzów locality, eastern part of the Holy Cross Mountains, Poland.
Description. Cranidium wider (tr., measured across the widest point of the palpebral lobes) than long (sag.), c. 140% of the cranidial length (sag.). Presence of tectonic deformation precludes providing exact range of proportions. Glabella pyriform, widening in the anterior part, anteriorly to S2. Occipital ring with small node at two-thirds of its length (sag.) from SO. Width (tr.) of L1 similar to or slightly larger than width (tr.) of occipital ring. Width (tr.) of L2 similar to or slightly larger than L1. Anterior part of glabella widens (tr.) forwards, widest (tr.) slightly anterior to S4; shape suboval, domed. In the widest (tr.) part reaching c. 132–150% of the width (tr.) of the occipital ring. SO with lateral sections straight; central portion curving anteriorly. S1 bent adaxially, joined in the middle of glabella. S2 arched towards the anterior, also connected across the glabella. SO, S1 and S2 distinct, deep; S1 and S2 shallower and wider (sag.) in the middle part. Anterior margin of S1 in the middle part of glabella occasionally bent forward. S3 and S4 furrows indistinct, shallow and short. Posterior border furrow broad (exsag.). Posterior branch of facial suture shorter than anterior branch, rarely preserved, ε located very close to the posterior border furrow. Palpebral lobes well developed (32–46% of the cranidial length (sag.)), arched, extending from posterior border furrow to S4. Palpebral lobes widest (tr.) in posterior part, narrower anteriorly, in anterior part forming a small arc towards the glabella. Palpebral furrows indistinct, slightly deeper in the posterior and anterior parts. Larger specimens with relatively smaller and less arched palpebral lobes (Fig. 13c). Fixigenae relatively wide (tr.), of more than half glabellar width (measured across the middle of the palpebral lobes). Anterior branches of facial sutures long, directed outwards and slightly forwards from anterior tips of ocular sutures, turning inwards to the front slightly posterior to anterior border furrow. Width (tr.) of anterior part of cranidium similar to width (tr.) of cranidium across centre of palpebral lobes, c. 129–156% of the cranidial width (tr.) between the most abaxial positions of the ocular sutures. Anterior border wider (exsag.) than pre-glabellar field, slightly narrower (sag.) in front of glabella. Pre-glabellar field narrow (sag.), narrower (sag.) than the anterior border. Anterior border furrow distinct, slightly bent in the abaxial parts.

Figure 13. Acadoparadoxides kozlowskii (Orłowski, 1959). (a) MWGUW ZI/29/3210, original of Orłowski (Reference Orłowski1964), pl. 2, fig. 2 and Żylińska & Szczepanik (Reference Żylińska and Szczepanik2009), pl. 4, fig. 9; (b) MWGUW ZI/29/3348, original of Orłowski (Reference Orłowski1964), pl. 2, fig. 3; (c) MWGUW ZI/29/3280; (d) MWGUW ZI/29/3311, original of Orłowski (Reference Orłowski1959a), pl. 1, fig. 1; (e) MWGUW ZI/29/3222, original of Orłowski (Reference Orłowski1964), pl. 2, fig. 1a–b; (f) MWGUW ZI/29/3266; (g) MWGUW ZI/29/3272, original of Orłowski (Reference Orłowski1959a), pl. 2, fig. 6a–b. Scale bars equal 5 mm.
Pygidium pyriform, widening (tr.) posteriorly. Axial part wide (tr.) (wider than the pleural part), with single axial ring defined by furrows. Platform large, extending almost to posterior margin, with a constant width (tr.). Rhachis poorly marked on the available specimens, reaching three-quarters of pygidial length (sag.). Single pair of pleural furrows on pleural field. Lateral pygidial margins directed strongly backwards, curving strongly adaxially at the end of the platforms creating lobes on both sides. Posterior margin wide (tr.), straight or with small median indentation.
Remarks. Due to the mode of preservation (isolated elements), a reconstruction of the entire exoskeleton was not possible. However, the overlapping stratigraphic ranges of one cranidium morphotype and one pygidium morphotype (the pygidia were described by Reference OrłowskiOrłowski 1959a as Paradoxides kozlowskii) in the upper part of the Jugoszów–Usarzów section allow a connection of these elements. As a result, they were assigned to Acadoparadoxides kozlowskii. The other cranidial and pygidial morphotypes are classified together as Acadoparadoxides samsonowiczi (see below for detailed description and discussion). Other parts of the exoskeleton were not diverse enough to discriminate them and to assign them to the described species.
Both Acadoparadoxides kozlowskii and A. samsonowiczi can be included in the earlier group of acadoparadoxidines as described above. Species from the Holy Cross Mountains are very similar to the taxa described from the Anti-Atlas of Morocco (Geyer, Reference Geyer1998; Geyer & Vincent, Reference Geyer and Vincent2015) and from Spain (Sdzuy, Reference Sdzuy1958). From the Scandinavian (A. oelandicus, A. pinus, A. torelli) and Bohemian groups (A. sacheri, A. sirokyi), the cranidia of the species from the Holy Cross Mountains differ in possessing a wider (tr.) anterior part of the cranidium and narrower (tr.) palpebral lobes. The pygidia of the Polish species differ from the variety of Scandinavian morphotypes, whereas the pygidia of the Czech taxa have a more rounded shape compared to the Polish species.
Some specimens described herein as Acadoparadoxides kozlowskii were assigned by Orłowski (Reference Orłowski1959a) to Paradoxides insularis. This assignment is considered incorrect, because they differ from Eccaparadoxides insularis from Scandinavia (Westergård, Reference Westergård1936) in possessing a much wider (tr.) anterior border and much less distinct S3 and S4 furrows. In addition, the pygidium of the Polish species differs in shape from the elongated pygidium of E. insularis with a deep indentation on the posterior margin.
Acadoparadoxides kozlowskii is closely related to a number of Moroccan species (Geyer, Reference Geyer1998; Geyer & Vincent, Reference Geyer and Vincent2015), although in some aspects it differs significantly enough to be distinguished from them. The material from Morocco is preserved in muddy fine-grained sandstones, thus the compaction of this material is much more extreme (the specimens are almost flat), likewise the preservation results in a different mode of deformation that includes cracking and bending of the test. These factors hamper comparisons and preclude the usage of advanced morphometric techniques (e.g. geometric morphometrics) in such comparisons. Acadoparadoxides kozlowskii differs from Acadoparadoxides pampalius in possessing slightly longer palpebral lobes that reach a bit further anteriorly. Palpebral lobes of the Polish species are also narrower (tr.) in the anterior part and slightly more bent. The anterior part of the glabella of A. kozlowskii is wider (tr.) in comparison to the rest of the glabella, but because this element is usually strongly deformed by compaction in the Moroccan specimens, this character should be treated with caution in the comparisons. The widest (tr.) part of the pygidium in A. kozlowskii is situated more posteriorly; the platform of A. pampalius is much shorter (sag.). Acadoparadoxides kozlowskii differs from Acadoparadoxides levisettii in the proportions of the anterior border width (tr.) versus the cranidial width (tr.) in the middle point of the palpebral lobes – in the Polish species these parameters are almost equal, whereas in the Moroccan species the anterior border width (tr.) is distinctly larger. The palpebral lobes are slightly shorter (exsag.) in A. levisettii. The pygidia are quite similar, but A. kozlowskii has a much longer (sag.) platform. The Polish species differs from Acadoparadoxides cf. mureroensis from Morocco in having slightly longer (exsag.) palpebral lobes. The pygidia of the two taxa differ distinctly in their outline, which is pyriform in A. kozlowskii and suboval in A. cf. mureroensis. From Acadoparadoxides nobilis, the species differs in a wider (tr.) posterior part of the glabella and less distinct curvatures on the outer parts of the anterior border. The pygidium of A. nobilis has a much larger posterior part of the pleural field, a very distinct indentation in the posterior margin and a shorter (sag.) platform. Acadoparadoxides kozlowskii has more arched palpebral lobes than Acadoparadoxides ovatopyge. In addition, the Polish species has more subparallel lateral margins of the glabella along L1. Pygidia of the two species differ in outline (pyriform in A. kozlowskii; suboval in A. ovatopyge), and the Moroccan species has a shorter (sag.) platform.
Differences between the cranidia of A. kozlowskii and A. samsonowiczi are not very distinct. The anterior part of glabella of A. kozlowskii expands laterally stronger than in A. samsonowiczi. The widest (tr.) part of the glabella in A. kozlowskii is located slightly anterior to S4, whereas in A. samsonowiczi it may sometimes lie slightly more posteriorly, between S3 and S4. The palpebral lobes of A. kozlowskii are slightly longer (tr.) and their anterior end lies closer to S4. The anterior margin of S1 is more strongly curved anteriorly in the median part of the glabella of A. kozlowskii. The adaxial curvature of the anterior branch of the facial suture lies closer to the anterior border furrow in A. kozlowskii (compare Fig. 13a, b, e with Fig. 14e). Pygidia of the two species can be easily distinguished by comparing their outlines: the pygidium of A. kozlowskii is pyriform, whereas that of A. samsonowiczi is suboval. In the latter species the widest (tr.) part of the pygidium lies in its middle, whereas in A. kozlowskii it is situated near the posterior end of the pygidium. The posterior margin of the pygidium of A. samsonowiczi is gently curved in comparison to the straight margin in A. kozlowskii. The differences between A. kozlowskii and Acadoparadoxides slowiecensis (Czarnocki, Reference Czarnocki1927a, b) (another acadoparadoxidine from the Holy Cross Mountains, previously known as Paradoxides slowiecensis; a revision will be provided in a subsequent paper – J. Nowicki & A. Żylińska in prep.) include: a wider (sag., exsag.) anterior border in A. kozlowskii; a less curved and wider (tr.) anterior part of the palpebral lobes in A. slowiecensis; a pre-glabellar field in adult specimens that is less reduced in A. kozlowskii; the pygidium with a larger, more distinct platform and a different, pyriform outline in A. kozlowskii compared to the suboval outline in the pygidial platform in A. slowiecensis.

Figure 14. Acadoparadoxides samsonowiczi (Orłowski, 1959). (a) WK40, original of Żylińska & Masiak (Reference Żylińska and Masiak2007), fig. 11c; (b) MWGUW ZI/29/0972, original of Żylińska & Masiak (Reference Żylińska and Masiak2007), fig. 11a–b; (c) MWGUW ZI/29/0959, original of Żylińska & Masiak (Reference Żylińska and Masiak2007), fig. 11g and Nawrocki et al. (Reference Nawrocki, Dunlap, Pecskay, Krzemiński, Żylińska, Fanning, Kozłowski, Salwa, Szczepanik and Trela2007), fig. 8.15; (d) MWGUW ZI/29/3288, original of Orłowski (Reference Orłowski1964), pl. 1, fig. 5; (e) MWGUW ZI/29/3212, original of Orłowski (Reference Orłowski1959a), pl. 2, fig. 4a–b and Żylińska & Szczepanik (Reference Żylińska and Szczepanik2009), pl. 4, fig. 10; (f) MWGUW ZI/29/3246, original of Orłowski (Reference Orłowski1964), pl. 2, fig. 9; (g) MWGUW ZI/29/3218, original of Orłowski (Reference Orłowski1959a), pl. 2, fig. 8a–b (holotype). Scale bars equal 5 mm.

Figure 15. Hydrocephalus? sp. MWGUW ZI/29/3341, original of Orłowski (Reference Orłowski1964), pl. 2, fig. 4. Scale bar equals 5 mm.
Acadoparadoxides samsonowiczi (Orłowski, 1959)
Figure 14
pars 1957 Paradoxides forms of the ölandicus group; Orłowski, fig. 5 (non figs 1–3 = Acadoparadoxides slowiecensis; non figs 4, 6 = Acadoparadoxides kozlowskii).
pars 1959a Paradoxides oelandicus Sjögren, Reference Sjögren1872; Orłowski, pp. 441–2, pl. I, fig. 4 (non figs 1–3 = Acadoparadoxides kozlowskii; non fig. 5 = Paradoxididae gen. et sp. indet.).
pars 1959a Paradoxides pinus Holm; Orłowski, p. 442, pl. I, fig. 7 (non fig. 6 = Acadoparadoxides kozlowskii; non fig. 8 = Paradoxididae gen. et sp. indet.).
1959a Paradoxides czarnockii n. sp.; Orłowski, pp. 443–4, pl. II, figs 4, 5.
1959a Paradoxides samsonowiczi n. sp.; Orłowski, p. 445, pl. II, fig. 8a, b.
pars 1964 Paradoxides oelandicus Sjögren; Orłowski, pl. I, fig. 5 (non figs 3, 4, 6–8 = Acadoparadoxides kozlowskii; non figs 1, 9–11 = Paradoxididae gen. et sp. indet.).
1964 Paradoxides czarnockii Orłowski, 1959; Orłowski, pl. II, figs 6, 7.
1964 Paradoxides samsonowiczi Orłowski, 1959; Orłowski, pl. II, figs 8, 9.
1970 Paradoxides oelandicus Sjögren; Bednarczyk, p. 34, pl. 1, fig. 1.
1990 Paradoxides czarnockii Orłowski, 1959; Bednarczyk, Lendzion & Orłowski, p. 58, pl. XVI, fig. 6.
1990 Paradoxides samsonowiczi Orłowski, 1959; Bednarczyk, Lendzion & Orłowski, p. 58, pl. XVII, fig. 11.
2007 Acadoparadoxides oelandicus (Sjögren); Nawrocki et al., fig. 8b.14.
2007 Acadoparadoxides cf. mureroensis (Sdzuy); Nawrocki et al., fig. 8b.15.
2007 Paradoxides (Acadoparadoxides) cf. mureroensis (Sdzuy, Reference Sdzuy1958); Żylińska & Masiak, pp. 679, 681, fig. 11d, g.
2007 Paradoxides (Acadoparadoxides) oelandicus Sjögren, Reference Sjögren1872; Żylińska & Masiak, pp. 681, 682, fig. 11a–c, e, i (non fig. 11f, h = Paradoxididae gen. et sp. indet.).
?2007 Paradoxides (Acadoparadoxides) sp.; Żylińska & Masiak, p. 682, fig. 11j.
2009 Paradoxides (Acadoparadoxides) czarnockii Orłowski, Reference Orłowski1959a; Żylińska & Szczepanik, pl. 4, fig. 10.
pars 2013a Paradoxides (Acadoparadoxides) oelandicus Sjögren, Reference Sjögren1872; Żylińska, fig. 6F (non fig. 7A = Acadoparadoxides kozlowskii).
2013a Paradoxides (Acadoparadoxides) czarnockii Orłowski, 1959; Żylińska, fig. 7J.
Holotype. Pygidium MWGUW ZI/29/3218, illustrated as specimen no. 3 in: Orłowski (Reference Orłowski1959a, pl. II, fig. 8a, b), and Bednarczyk, Lendzion & Orłowski (Reference Bednarczyk, Lendzion and Orłowski1990, pl. XVII, fig. 11).
Other material. Thirty-seven cranidia: WK36, WK40, WK59, MWGUW ZI/29/0959, ZI/29/0961, ZI/29/0962, ZI/29/0972, ZI/29/3212, ZI/29/3248, ZI/29/3249, ZI/29/3257, ZI/29/3259, ZI/29/3263, ZI/29/3288, ZI/29/3293, ZI/29/3294, ZI/29/3297, ZI/29/3299, ZI/29/3303, ZI/29/3307-ZI/29/3310, ZI/29/3312, ZI/29/3317, ZI/29/3318, ZI/29/3331, (?) ZI/29/3333, ZI/29/3346, ZI/29/3359, ZI/29/3369, ZI/29/3375, ZI/29/3387, ZI/29/3415, ZI/29/3419, ZI/29/3425, ZI/29/3443; five pygidia: MWGUW ZI/29/3245, ZI/29/3246, ZI/29/3247, ZI/29/3281, ZI/29/3313.
Occurrence. Cambrian Series 2 and 3 boundary interval (Acadoparadoxides samsonowiczi – Acadoparadoxides kozlowskii Assemblage Zone) in the Słowiec Sandstone Formation from Brzechów, central part of the Holy Cross Mountains, Poland; and Usarzów Sandstone Formation from the Jugoszów–Usarzów locality, eastern part of the Holy Cross Mountains, Poland.
Description. Cranidium wider (tr., measured across the widest point of the palpebral lobes) than long (sag.), around 140% of the cranidial length (sag.). The presence of tectonic deformation precludes providing the exact range of proportions. Glabella with straight borders, widening (tr.) anteriorly quite evenly. Occipital ring with small node in the centre (tr.), in two-thirds of its length (sag.) from SO. LO is the narrowest (tr.) of the glabellar lobes, L1 slightly wider (tr.), L2 also wider (tr.), anterior part of glabella widest (tr.) anteriorly to S3, often (but not always) posteriorly to S4. Anterior part of the glabella suboval in outline, domed, widest (tr.) part reaching c. 130–151% of the width (tr.) of the occipital ring. SO straight laterally, in the middle part arched anteriorly. SO, S1 and S2 distinct, deep. S1 transglabellar, shallowing medially and broadening (exsag., sag.). S2 also transglabellar, shallowing and broadening in the middle of the glabella. S3 and S4 shallow, indistinct and short. Posterior branches of facial sutures short, ε located very close to posterior border furrow. Palpebral lobes well developed (36–46% of the cranidial length (sag.)), arched, extending from near posterior border furrow to frontal lobe of glabella, adjacent to S3 and S4, widest in posterior part and narrowing anteriorly. Palpebral lobes between S3 and S4 slightly curved forward to form arches directed to glabella. Palpebral furrows indistinct, slightly deeper in the posterior and anterior parts. Fixigenae quite wide (tr.), more than half glabellar width (measured across the middle of the palpebral lobes). Anterior branches of facial suture longer than posterior parts, directed abaxially and slightly anteriorly from anterior ends of ocular suture, then curving adaxially transversely opposite S4. Anterior width of the cranidium around 129–156% of the cranidial width (tr.) between the most abaxial positions of the ocular sutures. Anterior border wider (exsag.) than pre-glabellar field, slightly narrower (sag.) in front of glabella. Pre-glabellar field narrower (sag.) than anterior border. Anterior border furrow distinct, with small bendings in the abaxial parts.
Pygidium suboval, widest (tr.) in the middle part. Axial part wide (tr.) (wider than the pleural part) with one axial ring moderately well defined by with furrows. Platform distinct, large (reaching almost to posterior margin of pygidium), with constant width (tr.). Rhachis indistinct, four-fifths of pygidial length (sag.). Pleural field with single pair of pleural furrows. Pygidial margin curved subevenly for the entire length.
Remarks. The correlation between morphotypes of pygidia and cranidia from the Jugoszów–Usarzów locality has been described in the remarks under Acadoparadoxides kozlowskii. Specimens from Brzechów have been assigned to A. samsonowiczi, but this assignment is uncertain because of the lack of complete pygidia. The only known pygidium from this locality lacks the posterior part, which is crucial for the determination. The cranidia from Brzechów fit well into the morphotype described as A. samsonowiczi, although the differences in the acadoparadoxidine cranidia are less distinct. Thus a diagnosis based solely on the cranidia is less reliable. This is also the reason for choosing the name for the species after the name given to the pygidia (Paradoxides samsonowiczi) and not for cranidia that were described in the same paper (Orłowski, Reference Orłowski1959a) as Paradoxides czarnockii.
Acadoparadoxides samsonowiczi is closely related to A. kozlowskii and can also be assigned to the earliest group of acadoparadoxidines. The reasons for this assignment and the differences between this group and the species of Acadoparadoxides from Scandinavia and Bohemia are given in the remarks for A. kozlowskii. The differences between A. samsonowiczi and Eccaparadoxides insularis are also presented therein.
Like Acadoparadoxides kozlowskii, A. samsonowiczi resembles the species from Morocco (Geyer, Reference Geyer1998; Geyer & Vincent, Reference Geyer and Vincent2015). The differences between the cranidia are subtle and difficult to recognize due to the different mode of preservation. Cranidia of A. samsonowiczi differ from A. pampalius in a less widening (tr.) anterior part of glabella. Also the anterior parts of the facial sutures have a different shape, bending towards the anterior much more anteriorly in A. pampalius. Palpebral lobes of A. samsonowiczi are slightly narrower (tr.) in the anterior part. Pygidia are similar in outline, but in A. samsonowiczi the platform is distinctly longer (sag.), almost reaching to the posterior margin of the pygidium. The cranidia of A. samsonowiczi differ from those of A. levisettii in proportions of the width (tr.) of the anterior margin of cranidium versus cranidial width (tr.) in the middle of the palpebral lobes. In the Polish species these dimensions are similar, whereas in the Moroccan species the anterior width (tr.) is larger. The curve in the anterior part of the facial suture is situated more anteriorly in A. levisettii (according to the position of the anterior border furrow). Pygidia differ in shape (suboval in A. samsonowiczi versus pyriform in A. levisettii). Moreover, the platform in the pygidium of the Moroccan species is shorter. The pygidial posterior margin in A. samsonowiczi lacks an indentation, which is sometimes developed in A. levisettii. The Polish species differs from A. cf. mureroensis from Morocco in possessing a less wide (tr.) anterior part of glabella. The pygidium of A. samsonowiczi has a longer (sag.) platform. The widest part of the pygidium is situated more posteriorly in A. cf. mureroensis. From A. nobilis the species from Poland differs in the shape of the anterior branch of the facial suture (in a similar way to A. pampalius and A. levisettii). The curvature in the outer part of the anterior margin furrow is slightly less distinct in A. samsonowiczi. The anterior part of the glabella is wider (tr.) in A. nobilis. The pygidia differ distinctly in their outline (pyriform with an indentation on the posterior margin in the Moroccan species versus suboval in the Polish species). The platform is longer (sag.) in A. samsonowiczi. From A. ovatopyge, the cranidia of A. samsonowiczi differ in possessing a smaller width (tr.) of the anterior part of the glabella and in the shape of the anterior branches of the facial sutures (similarly to the other Moroccan species). The pygidia are very similar, but those of the Polish species have a longer (tr.) platform.
The differences between A. samsonowiczi and A. kozlowskii have been pointed out in the description of the latter species. The differences between cranidia of A. samsonowiczi and A. slowiecensis are very similar to those between A. kozlowskii and A. slowiecensis. The pygidium of A. samsonowiczi is more similar to the pygidium of A. slowiecensis, but has a more circular outline in the latter species and the platform is distinctly larger in A. samsonowiczi.
Genus Hydrocephalus Barrande, Reference Barrande1846
Type species. Hydrocephalus carens Barrande, Reference Barrande1846 from the Cambrian of Bohemia, OD.
Hydrocephalus? sp.
Figure 15
1959a Paradoxides torelli Holm; Orłowski, p. 443, pl. II, fig. 1.
pars 1964 Paradoxides torelli Holm; Orłowski, pl. II, fig. 4 (non fig. 5 = Paradoxididae gen. et sp. indet.).
Material. Two fragmentary cranidia; MWGUW ZI/29/3341 and ZI/29/3414.
Occurrence. Cambrian Series 2 and 3 boundary interval (Acadoparadoxides samsonowiczi – Acadoparadoxides kozlowskii Assemblage Zone) in the Usarzów Sandstone Formation from the Jugoszów–Usarzów locality, eastern part of the Holy Cross Mountains, Poland.
Description. Specimens known only from fragments of cranidia, which show glabellae, fixigenae, palpebral lobes and part of the anterior margin. Glabella pyriform, strongly widening (tr.) anteriorly from S2. Small node situated on occipital ring, near its posterior margin. Widths (tr.) of LO, L1 and L2 similar. Anterior part large, suboval, domed, widest anterior to S4. SO distinct, deep, straight laterally, curving anteriorly in the median part. S1 and S2 distinct, deep, transglabellar, shallow in the middle of the glabella. S1 straight, directed posteriorly from near axial furrows, in the middle part almost orthogonal to axis. S2 arched anteriorly. S3 and S4 short and indistinct. Posterior border furrow wide (exsag.). Length of posterior branches of facial sutures unknown. Fixigenae wide (tr.), up to 60% of glabellar width (tr.) (measured across the middle of the palpebral lobes). Palpebral lobes of smaller specimen long, with nearly equal curvature of abaxial and adaxial margins, but with small additional flexure in the anterior parts, widest in the central section, tapering towards both ends. Palpebral furrow quite shallow, but well defined. Palpebral lobes extend from posterior margin furrows to glabella at L4. Palpebral lobes of larger specimen shorter and narrower in relative size, but similar in shape. Anterior margin unknown except for the part in front of the glabella, which tapers distinctively towards the sagittal line. Pre-glabellar field absent.
Remarks. Two specimens from the Jugoszów–Usarzów locality do not fit with the characters seen in Acadoparadoxides samsonowiczi and A. kozlowskii. The differences include the shape of the palpebral lobes (wider in the smaller specimen and more reduced in the larger specimen compared to the two acadoparadoxidine species) and the anterior margin (which is distinctly wider in the two species). In addition, the glabellae of these two specimens are narrower (tr.) in the posterior parts.
Weidner, Rushton & Ebbestad (Reference Weidner, Rushton and Ebbestad2014) pointed out that specimens described previously by Orłowski (Reference Orłowski1959a, Reference Orłowski1964) as Paradoxides torelli do not match the characters of this species. The shape of the anterior border, the palpebral lobes of the larger specimen and the shape of the glabella suggest that these specimens may be assigned to Hydrocephalus, although poor preservation precludes a confident assignment. Moreover, the genus Hydrocephalus is in need of a revision (J. Nowicki, unpub. Ph.D. thesis, Univ. Warsaw, 2016; J. Nowicki & A. Żylińska in prep.), and this assignment should be regarded as an interim solution.
8. Implications for biostratigraphy and palaeobiogeography
8.a. Biostratigraphy
Revision of the paradoxidids from Brzechów and from the Jugoszów–Usarzów locality has major impact on the biostratigraphic scheme for the Holy Cross Mountains in the analysed stratigraphic interval. The former biozones, Eccaparadoxides insularis and Ptychagnostus praeccurens – Acadoparadoxides pinus (see e.g. Orłowski, Reference Orłowski1992; Żylińska, Reference Żylińska2013a), cannot be used further, since the species that were used to define them are not present in the analysed succession. The traditional ‘Middle’ Cambrian interval of the Holy Cross Mountains is replete with endemic trilobite species, and the paradoxidids also follow this trend. Correlation on the species level is impossible, so a new local biozonation has to be developed. The new scheme is also based on paradoxidids, the remaining trilobites being of much lower stratigraphic resolution (Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009). Although the previous scheme included two zones, the boundary between them cannot be maintained; according to the revised systematics, it would be determined by the last appearance datum (LAD) of A. samsonowiczi so that the stability of this boundary is questionable. Placing the zonal boundary on the first appearance datum (FAD) of A. kozlowskii is also inappropriate. This bioevent probably lies between the interval represented in Brzechów and the interval represented in the Jugoszów–Usarzów locality, and its true position remains unrecognized. Moreover, the material from Brzechów, assigned to A. samsonowiczi, consists only of cranidia so that the assignment of these specimens is less confident. The present state of knowledge allows determining the Acadoparadoxides samsonowiczi – Acadoparadoxides kozlowskii Assemblage Zone encompassing both ranges of these species. The paradoxidids are accompanied by several other trilobite species. Conomicmacca kielcensis and ‘Palaeolenus’ medius occur only in Brzechów and may suggest the lower part of the Acadoparadoxides samsonowiczi – Acadoparadoxides kozlowskii Assemblage Zone.
Correlation of the Acadoparadoxides samsonowiczi – Acadoparadoxides kozlowskii Zone is difficult, but not impossible. It has been done partly by Żylińska & Masiak (Reference Żylińska and Masiak2007), Żylińska & Szczepanik (Reference Żylińska and Szczepanik2009) and summarized in Żylińska Reference Żylińska(2013a) (see also Sundberg et al. Reference Sundberg, Geyer, Kruse, McCollum, Pegel’, Żylińska and Zhuravlev2016). Correlation of the Brzechów trilobites with the Morocconus notabilis Zone of Morocco is also supported by the revised paradoxidids. Both A. samsonowiczi and A. kozlowskii fit very well in the earliest group of acadoparadoxidines described above. These species are endemic; however, they are closely related to the Moroccan species described by Geyer & Vincent (Reference Geyer and Vincent2015). This relationship and the differences between the forms from the Holy Cross Mountains and Scandinavia also permit correlation of the Jugoszów–Usarzów succession with the Morocconus notabilis Zone (Fig. 16). The occurrence of acadoparadoxidines in the upper part of the Morocconus notabilis Zone in the Anti-Atlas successions (Geyer & Vincent, Reference Geyer and Vincent2015) suggests that the analysed sequence from the Holy Cross Mountains may be related to this part. Exact correlation is impossible, but the presented resolution is sufficient to show that the analysed interval is older than considered previously (e.g. Orłowski, Reference Orłowski1992) and, according to the recent report on the Cambrian Series 2 and 3 boundary (Sundberg et al. Reference Sundberg, Geyer, Kruse, McCollum, Pegel’, Żylińska and Zhuravlev2016), in fact at least partially represents Cambrian Series 2 if the FAD of Oryctocephalus indicus is chosen to define the base of Series 3. Therefore, the boundary between Cambrian Series 2 and 3 in the Holy Cross Mountains would fall within the newly proposed Acadoparadoxides samsonowiczi – Acadoparadoxides kozlowskii Zone, but cannot be precisely indicated. The absence of Acadoparadoxides cf. mureroensis in the Holy Cross Mountains confirms that this species is not as widely dispersed as assumed previously by some authors (Gozalo et al. Reference Gozalo, Dies Alvarez, Gamez Vintaned, Zhuravlev, Bauluz, Subías, Chirivella Martorell, Mayoral, Gursky, Andrés and Liñán2013). New data from the Holy Cross Mountains support rather the view presented by Geyer & Vincent (Reference Geyer and Vincent2015).

Figure 16. Revised biostratigraphic scheme for the Cambrian Series 2 and 3 boundary interval in the Holy Cross Mountains (Poland) and its correlation with the biostratigraphic scheme for the same interval in Morocco (right column; Geyer & Landing, Reference Geyer, Landing, Geyer and Landing2006). The name of the uppermost zone in the Holy Cross Mountains is after J. Nowicki (unpub. Ph.D. thesis, Univ. Warsaw, 2016) and J. Nowicki & A. Żylińska (in preparation).
8.b. Palaeobiogeography
The faunistic connection of the paradoxidids from the Holy Cross Mountains with taxa from West Gondwana fits well in the general trend of biogeographic affinities of the trilobites from the studied interval. The diversity of the ellipsocephalids is very similar to that known from West Gondwanan and Avalonian successions (Żylińska & Szczepanik, Reference Żylińska and Szczepanik2009; Żylińska, Reference Żylińska2013a); of similar affinities is also Conomicmacca kielcensis from Brzechów (Geyer & Landing, Reference Geyer and Landing2004; Żylińska & Masiak, Reference Żylińska and Masiak2007; Elicki & Geyer, Reference Elicki and Geyer2013). Since there are no direct faunal links with Baltica, the occurrence of such exotic fauna has to be resolved. Fairly recent palaeogeographic reconstructions by Torsvik & Cocks (Reference Torsvik, Cocks, Harper and Servais2013) indicate that the distance between the present Anti-Atlas region and the putative location of the Małopolska Block near the western margin of Baltica does not support the possibility of dispersion that would lead to the presence of Gondwana-derived fauna in the Holy Cross Mountains succession. The position of the Małopolska Block in the neighbourhood of Baltica, proven in various ways (Nawrocki et al. Reference Nawrocki, Dunlap, Pecskay, Krzemiński, Żylińska, Fanning, Kozłowski, Salwa, Szczepanik and Trela2007), is still a matter of discussion (e.g. Walczak & Belka, Reference Walczak and Belka2017). The position of Gondwana and Baltica presented by Torsvik & Cocks (Reference Torsvik, Cocks, Harper and Servais2013) is also questionable, due to the extreme movement of West Gondwana from low latitudes that favoured the existence of a carbonate platform (e.g. Landing et al. Reference Landing, Geyer, Brasier and Bowring2013) with archaeocyathan reefs (Kerner & Debrenne, Reference Kerner, Debrenne, Harper and Servais2013) in Cambrian Epoch 2 to the South Pole in Cambrian Epoch 3 (Torsvik & Cocks, Reference Torsvik, Cocks, Harper and Servais2013). It is possible that the spatial relations between Baltica, the Małopolska Block and West Gondwana were different than previously believed. The dispersal of Gondwana/Avalonia-related trilobite taxa into the Małopolska Block could have been maintained by oceanic currents (Żylińska, Reference Żylińska2013a, b), and if so, then the Małopolska Block should have faced West Gondwana. The occurrence of paradoxidids similar to those present in the Anti-Atlas successions in the Cambrian of the Holy Cross Mountains provides additional evidence for this interpretation. The possible configuration of Baltica with the adjacent Małopolska Block and with West Gondwana would include a different position of the Małopolska Block near Baltica, or Baltica facing Gondwana with its western margin; and a position of Baltica closer to the Atlas sector of West Gondwana in the Cambrian. The latter configuration implies the location of an oceanic current similar to that presented by Servais et al. (Reference Servais, Danelian, Alexander, Harper and Munnecke2014), but allowing for the transportation of trilobite larvae from the Gondwanan margin into the Baltic realm. The existence of a current transporting larvae is supported by the presence of the Gondwana-related species such as Rectifrontinella olhae Konstantinenko, 2001 (a member of the Conomicmacca/Myopsolenites clade: see Żylińska & Masiak, Reference Żylińska and Masiak2007), in the Ukraine (Carpathian Foredeep) (Drygant & Konstantinenko, Reference Drygant and Konstantinenko2001). The hypothesis of long transportation of paradoxidid larvae is also supported by a recent study by Laibl, Esteve & Fatka (Reference Laibl, Esteve and Fatka2017), in which protaspids of Bohemian paradoxidids are interpreted as lecitotrophic larvae. This mode of life would make the larvae more able to survive transport and allow them to reach distant continental shelves like those of Baltica.
An alternative concept of the palaeogeographic position of the Małopolska Block during the Cambrian also does not resolve the apparent disparate distribution of the fauna. In this view, the Małopolska Block was a part of the Cadomian orogen bordering Amazonia that rifted from Gondwana before the early Cambrian and accreted to Baltica during Furongian–Tremadocian times (Sandomierz Phase) (Belka et al. Reference Belka, Valverde-Vaquero, Dörr, Ahrendt, Wemmer, Franke and Schäfer2002). This concept is supported by geochemical data (Walczak & Belka, Reference Walczak and Belka2017), suggesting a Gondwanan affinity of the detritic material with cooling ages similar to those of the Amazonian terrane basement, and by geophysical data that allow for a Cadomian interpretation of the Małopolska Block basement (Narkiewicz & Petecki, Reference Narkiewicz and Petecki2017). Nawrocki et al. (Reference Nawrocki, Dunlap, Pecskay, Krzemiński, Żylińska, Fanning, Kozłowski, Salwa, Szczepanik and Trela2007) also indicated a non-Baltic (Cadomian) age of the material from the middle Cambrian and younger strata, but their interpretation assumed supply of this type of material from the Uralid margin of Baltica and not from Gondwana. The data from Walczak & Belka (Reference Walczak and Belka2017) are incomplete, as not all Cambrian localities have been analysed, for instance the Słowiec Hill. Belka et al. (Reference Belka, Valverde-Vaquero, Dörr, Ahrendt, Wemmer, Franke and Schäfer2002) and Walczak & Belka (Reference Walczak and Belka2017) also use the biogeographic interpretation of Cambrian brachiopods as evidence for early Cambrian connections with Gondwana and Avalonia, but the present knowledge about this group definitely does not allow such interpretation (Bassett, Popov & Holmer, Reference Bassett, Popov, Holmer, Crame and Owen2002; Winrow & Sutton, Reference Winrow and Sutton2014). Nevertheless, the possibility of West Gondwanan affinities of the Małopolska Block should be reconsidered, since the trilobite fauna has strong connections with taxa from this realm. Palaeomagnetic data support the position of the Małopolska Block close to Baltica, but the accuracy of this method for the Cambrian was questioned (Landing et al. Reference Landing, Geyer, Brasier and Bowring2013) and the confidence interval of the latitude was estimated to be c. 20° and (in some cases) even 40° (Smith, Reference Smith2001). Because Walczak & Belka (Reference Walczak and Belka2017) suggest that the Małopolska Block was not in the vicinity of the West African part of Gondwana during the Cambrian, the scenario with the position of the Małopolska Block as an independent terrane situated in Cambrian Epoch 2 in closer proximity to Gondwana still requires a mechanism for the dispersal of fauna, of which oceanic currents seem the most probable factors. The conclusions drawn by Walczak & Belka (Reference Walczak and Belka2017) and herein are based on completely different data. The problem of the configuration of palaeogeographic elements in the Cambrian seems to be still an open case, and one of the further research topics should be the revision of brachiopods and further work on the revision of the trilobites.
9. Conclusions
Revised paradoxidids from the Holy Cross Mountains imply several important changes in the interpretation of the succession previously described as the traditional ‘Middle’ Cambrian:
(1) The previous descriptions of eight species have been corrected; they have been replaced by two species (Acadoparadoxides kozlowskii and Acadoparadoxides samsonowiczi). Moreover, one taxon is retained in open nomenclature (Hydrocephalus? sp.). Both acadoparadoxidines represent an earliest group of Acadoparadoxides and are closely related to the taxa from the Gondwanan realm rather than from Baltica (Scandinavia). Members of this clade have been described outside Gondwana for the first time.
(2) Due to the absence of diagnostic taxa, the previous Eccaparadoxides insularis and Acadoparadoxides pinus – Pentagnostus praeccurrens zones have been replaced by the Acadoparadoxides samsonowiczi – Acadoparadoxides kozlowskii Assemblage Zone.
(3) The A. samsonowiczi – A. kozlowskii Assemblage Zone can be correlated with the Moroccan Morocconus notabilis Zone, and thus points to an interval located near the Cambrian Series 2 – 3 boundary defined by the FAD of Oryctocephalus indicus Reed, Reference Reed1910. Defining the precise position of this boundary within the A. samsonowiczi – A. kozlowskii Zone is impossible. Former correlations with the Scandinavian Acadoparadoxides oelandicus Superzone have to be disclaimed.
(4) The presence of Gondwana-related paradoxidids is consistent with the present knowledge about other trilobites from this interval in the Holy Cross Mountains. Thus, the palaeogeographical position of the Małopolska Block has to be revised. The hypothesis presented herein assumes a position closer to the Gondwana margin (but does not conclude on the distance from Baltica) and the influence of oceanic currents in the distribution of trilobite larvae from Gondwana to the Małopolska Block.
Acknowledgements
This study was prepared as a result of project 2014/13/N/ST10/02693 funded by the National Science Centre (NCN), Poland, and is part of a Ph.D. thesis by J.N. prepared at the Faculty of Geology, University of Warsaw. Fieldwork was additionally supported by the Institute of Basic Geology, Faculty of Geology, University of Warsaw. The manuscript of this paper gained significantly from insightful and constructive reviews by Gerd Geyer and an anonymous reviewer, to whom we offer our gratitude. We appreciate the help of Małgorzata Bieńkowska-Wasiluk with the photographs, and Ireneusz Walaszczyk for his constructive remarks and discussions on an earlier draft of the manuscript.






















